- Protein splicing

Содержание
- 2. Observation: Nuclear RNA pool consists of very high molecular weight species as well as lower molecular
- 3. Experiment: Treat cells with UV for varying periods of time. Thymidine dimers will form, blocking transcription.
- 4. Experiment: Treat cells with UV for varying periods of time. Thymidine dimers will form, blocking transcription.
- 5. RNA is unstable – it can cleave itself. RECAP (2) Self-splicing introns utilize this suicidal tendency
- 6. Splicing in eukaryotes probably relies on the same chemistry as self-splicing group II introns. RECAP (3)
- 7. The spliceosome is made up of 5 small nuclear ribonucleoprotein subunits + > 100 proteins. These
- 8. Structures of the Spliceosomal snRNAs U1, U2, U4, U5 RNA Pol II transcripts TriMethyl G Cap
- 9. The earliest snRNP to bind to the pre-mRNA is U1, which uses its snRNA to base-pair
- 10. The U2 snRNP binds to the branchpoint via RNA/RNA base-pairs to create a bulged A residue.
- 11. The protein U2AF (U2 Auxiliary Factor) binds to the Polypyrimidine tract and the AG of the
- 12. Splice sites do not always perfectly match the consensus sequences. Thus, the base-pairing interactions between the
- 13. The interactions of U1 with the 5’ splice site and U2 with the branchpoint were proven
- 14. The full spliceosome is formed from the pre-spliceosome by the addition of the U4/U5/U6 Tri-snRNP.
- 15. In the U4/U6 Di-snRNP and the U4/U5/U6 Tri-snRNP, the U4 and U6 snRNAs are base-paired to
- 16. After the formation of the full spliceosome, the U1 and the U4 snRNPs are detached and
- 17. In the catalytically active spliceosome, the U2, U5 and U6 snRNAs make very specific contacts with
- 18. The two transesterification reactions of splicing take place in the mature spliceosome.
- 19. After the second transesterification reaction, the spliceosome comes apart. The snRNPs are recycled, and the spliced
- 20. The lariat intron is debranched by Debranching Enzyme returning it to a typical linear state. This
- 21. Mobile genetic elements provide an example of RNP complexes in which proteins and RNAs cooperate for
- 22. In the catalytically active spliceosome, the U2, U5 and U6 snRNAs make very specific contacts with
- 23. A tale of the U5 protein, Prp8. Prp8 mutants are splicing defective. Many Prp8 mutations suppress
- 24. Crystal structure of Prp8 reveals a cavity of appropriate dimensions to position spliceosomal RNAs for catalysis.
- 25. Splicing is dynamic, with sequential regulated alterations in RNA:RNA and RNA:protein interactions
- 26. DEAD-box helicases found at every step
- 27. Splicing error rates range from 1 in 1000 to 1 in 100,000 DEAD-box RNA helicases implicated
- 28. Monomeric (vs. “AAA” ATPases) RNA-dependent ATPases ~300 aa domain with 7 signature motifs (e.g. eponymous tetrapeptide)
- 29. The story of one helicase: PRP16 Prp16 is required for the second chemical step: - Immunodeplete
- 30. The story of one helicase: PRP16 Prp16-1 mutant was identified in a screen for reduced-fidelity mutants:
- 31. Hypothesis: Prp16 promotes fidelity 1) branchpoint mutations -> slow conformational rearrangement -> rejection 2) suppressor mutations
- 32. How to discriminate between “correct” vs. “incorrect”? A “slow” spliceosome -> ATP-dependent rejection of WT substrate.
- 33. PRP16: functions at 2 steps PRP16 binds before 5’ss cleavage and acts as a sensor to
- 34. Questions How are the splice sites identified? How are the intervening sequences removed?
- 35. How are the splice sites identified? In higher eukaryotes, there isn’t much sequence information encoded in
- 36. How are the splice sites identified? Minor spliceosome, consists of U11, U12, U4atac, U6atac, and U5
- 37. 2.4 Mb 260 kb intron Human Dystrophin gene Genes in higher eukaryotes have many exons and
- 38. The same primary transcript can be spliced many different ways (estimated 90% of genes experience alternative
- 39. Because of the intron length and lack of specificity of splice sites, most introns contain numerous
- 40. How are the splice sites identified? x outcomes of 5’ ss mutants 1. activates cryptic 5’
- 41. How are the splice sites identified? beta-globin mutants that create a new 3’ ss within an
- 42. In multicellular organisms, exons are recognized as units prior to assembly of the spliceosome across the
- 43. How are the splice sites identified? A U2AF Exon 1 U1 snRNP RS 70K RS SF2
- 44. Differential size distributions of exons (~50 to 300 nt) vs. introns ( SR protein - preferentially
- 45. Cross-exon bridging interactions involve SR domains of U2AF, U170K And 1 or more SR-family proteins ~12
- 46. Vertebrate external exons
- 47. Splicing is co-transcriptional and all introns assayed are spliced within 5-10 minutes of transcription of the
- 48. Defining an exon involves the specific stabilization or destabilization of splice site recognition Stabilization: exon inclusion
- 49. Regulation of alternative splicing involves the specific stabilization or destabilization of splice site recognition Stabilization: exon
- 50. How would you identify cis-regulatory sequences responsible for alternative splicing ? Examine RNA Splicing
- 51. Four classes of splicing regulatory elements: Exonic Splicing Enhancers, Exonic Splicing Silencers (ESS), Intronic Splicing Enhancers
- 52. How would an Intronic Splicing Silencer work
- 53. SR proteins generally bind ESE, ESS, ISE, and ISSs
- 54. The SR Proteins are a family of proteins with a common domain structure of 1 or
- 55. SR Proteins bind to specific RNA elements using their RNA binding domains similar to those in
- 56. Characterization of an ESE and SR protein in flies Sex differentiation in flies controlled by AS
- 57. hnRNP contain RRMs but not SR domain Can block sterically, tighter binding affinity than U2AF hnRNP
- 58. SR Proteins bind to CTD of polII: promote co-transcriptional splicing?
- 59. CTD of RNA pol II plays important role in pre-mRNA splicing (Kornblihtt et al, 2004)
- 60. Does splice site strength affect alternative splicing?
- 61. A connection between chromatin and splicing include exonIIIc by repress exonIIIb include exonIIIb, repress exon IIIc,
- 62. mRNA export - formation of an export competent mRNP Sees formation of mRNP as transcription commences
- 63. (Stutz & Izaurralde,2003) Factors involved in mRNA export are co-transcriptionally recruited THO complex: major role in
- 64. (Cullen, 2003) (Sub2p) (Yra1p) (Mtr2p) (Mex67p) (yeast homolog is indicated in parentheses) Proteins involved in the
- 65. (Linder & Stutz, 2001) Sub2, Yra1p and hnRNP proteins such as Npl3p associate co-transcriptionally with the
- 66. Genetic approach to identify genes involved in mRNA export process (Lei et al, 2003) Mutagenized cells
- 67. (Stutz & Izaurralde, 2003) Nuclear mRNA accumulation is observed after shifting mex67 TS mutant to the
- 68. Yra1p and Nab2p are essential for mRNP docking to the Mlp export gate at the nuclear
- 69. (Vinciguerra & Stutz, 2004) The perinuclear Mlp1p protein contributes to mRNP surveillance by retaining unspliced transcripts
- 71. Скачать презентацию
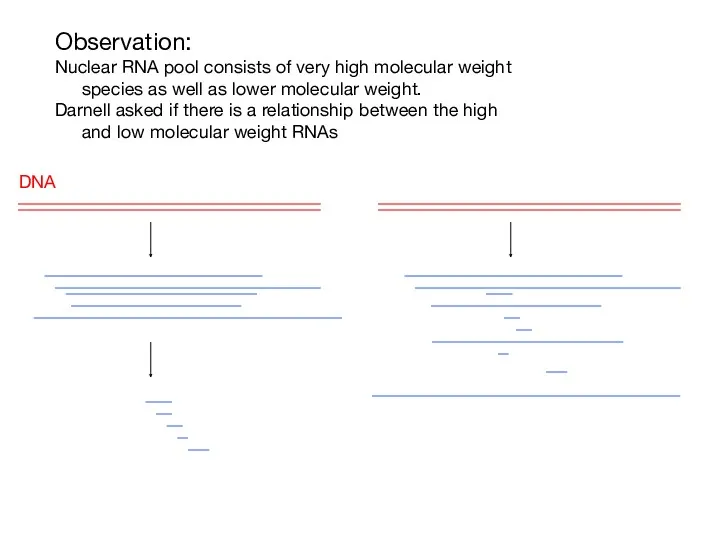
Observation:
Nuclear RNA pool consists of very high molecular weight species as
Observation:
Nuclear RNA pool consists of very high molecular weight species as
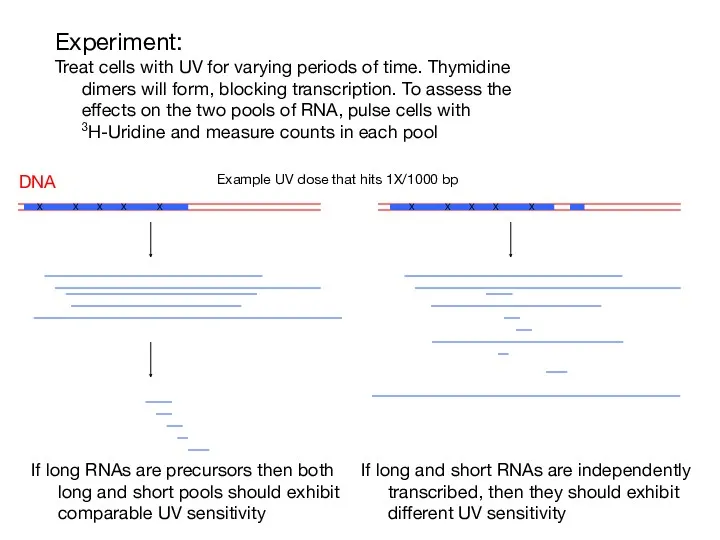
Experiment:
Treat cells with UV for varying periods of time. Thymidine dimers
Experiment:
Treat cells with UV for varying periods of time. Thymidine dimers
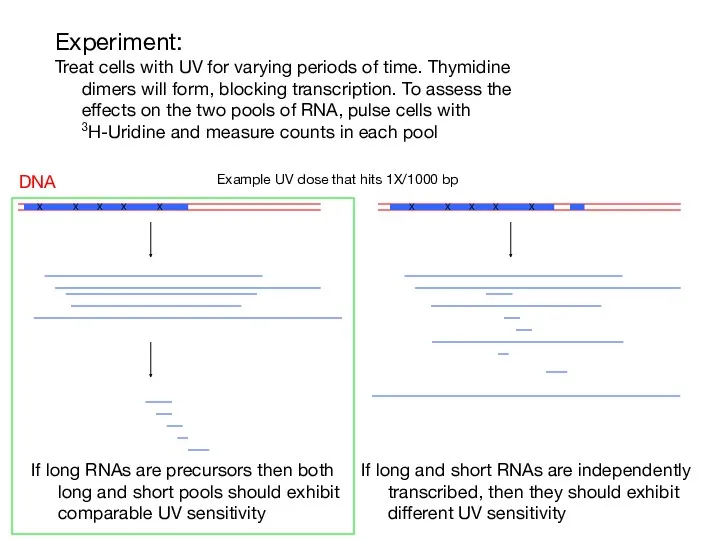
Experiment:
Treat cells with UV for varying periods of time. Thymidine dimers
Experiment:
Treat cells with UV for varying periods of time. Thymidine dimers
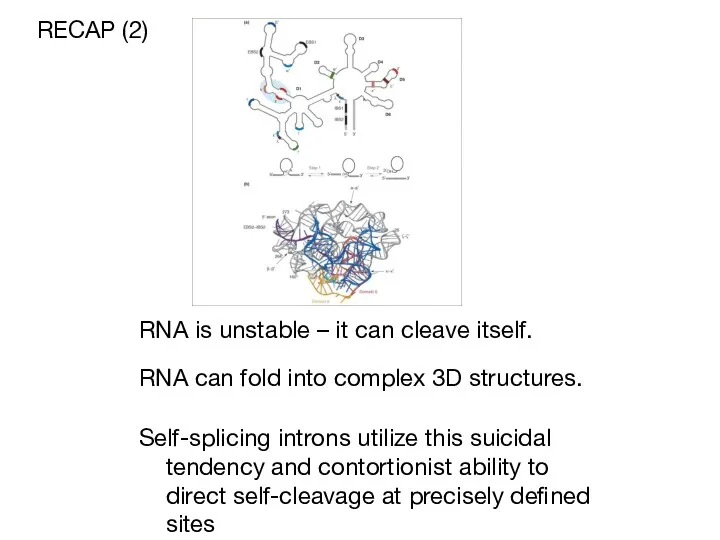
RNA is unstable – it can cleave itself.
RECAP (2)
Self-splicing introns
RNA is unstable – it can cleave itself.
RECAP (2)
Self-splicing introns

Splicing in eukaryotes probably relies on the same chemistry as self-splicing
Splicing in eukaryotes probably relies on the same chemistry as self-splicing
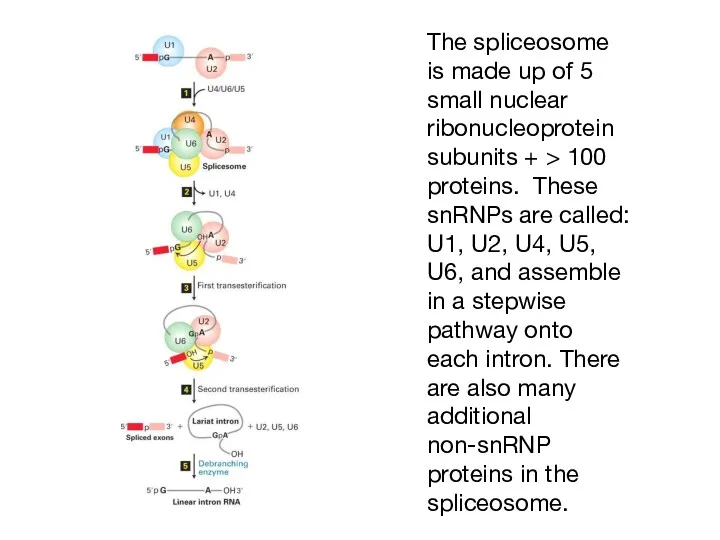
The spliceosome is made up of 5 small nuclear ribonucleoprotein subunits
The spliceosome is made up of 5 small nuclear ribonucleoprotein subunits
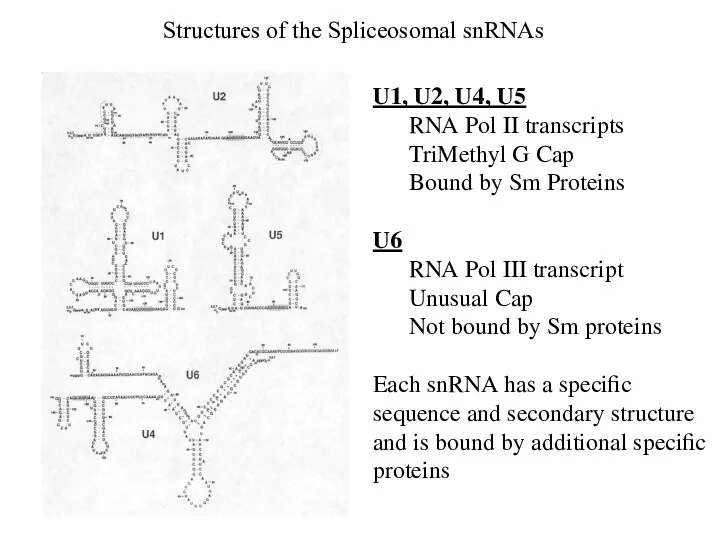
Structures of the Spliceosomal snRNAs
U1, U2, U4, U5
RNA Pol II transcripts
TriMethyl
Structures of the Spliceosomal snRNAs
U1, U2, U4, U5
RNA Pol II transcripts
TriMethyl
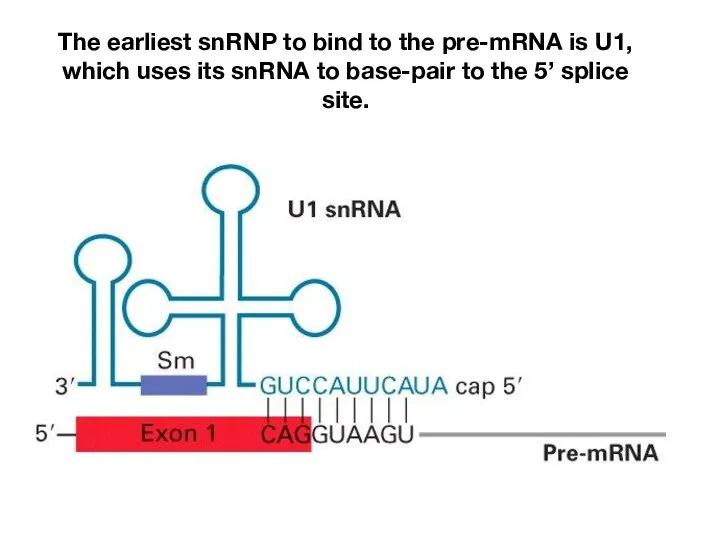
The earliest snRNP to bind to the pre-mRNA is U1, which
The earliest snRNP to bind to the pre-mRNA is U1, which
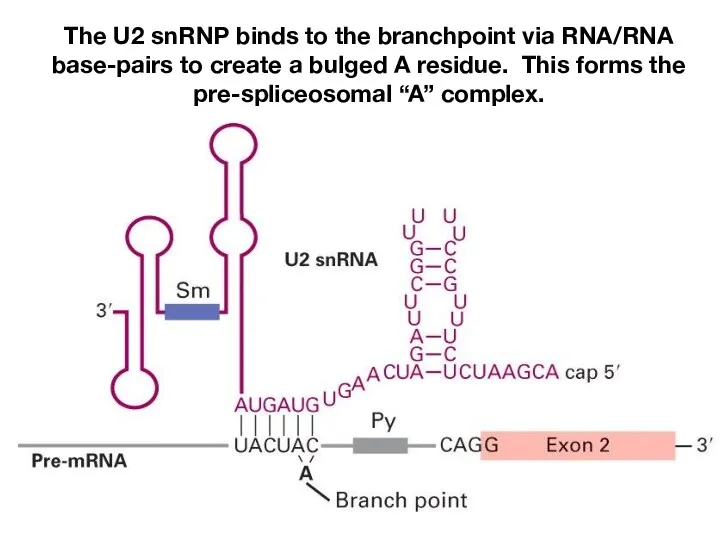
The U2 snRNP binds to the branchpoint via RNA/RNA base-pairs to
The U2 snRNP binds to the branchpoint via RNA/RNA base-pairs to
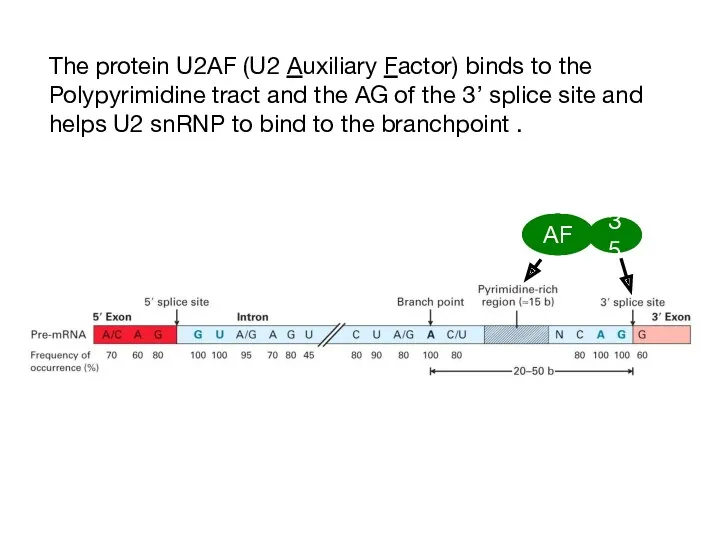
The protein U2AF (U2 Auxiliary Factor) binds to the Polypyrimidine tract
The protein U2AF (U2 Auxiliary Factor) binds to the Polypyrimidine tract
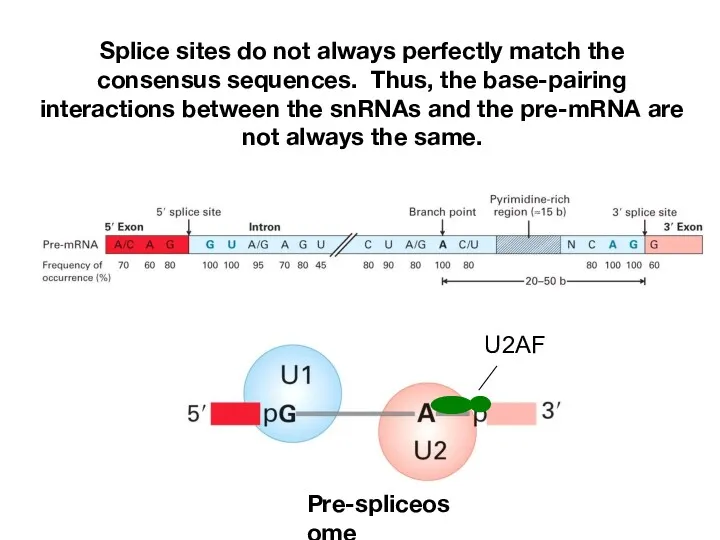
Splice sites do not always perfectly match the consensus sequences. Thus,
Splice sites do not always perfectly match the consensus sequences. Thus,
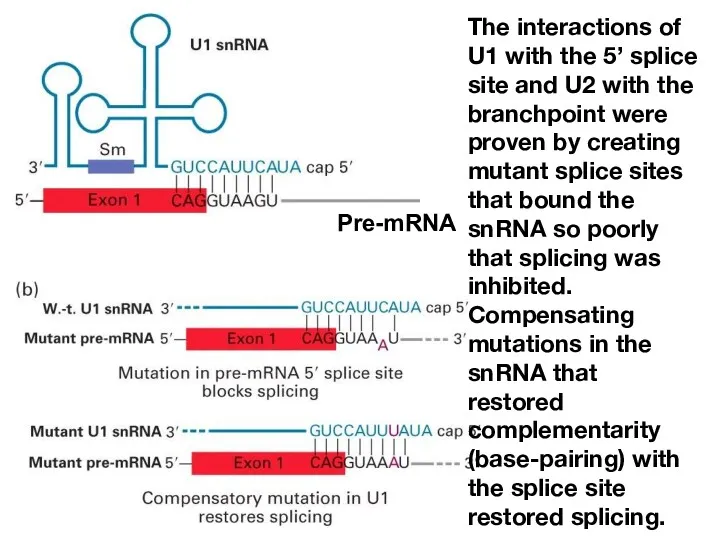
The interactions of U1 with the 5’ splice site and U2
The interactions of U1 with the 5’ splice site and U2
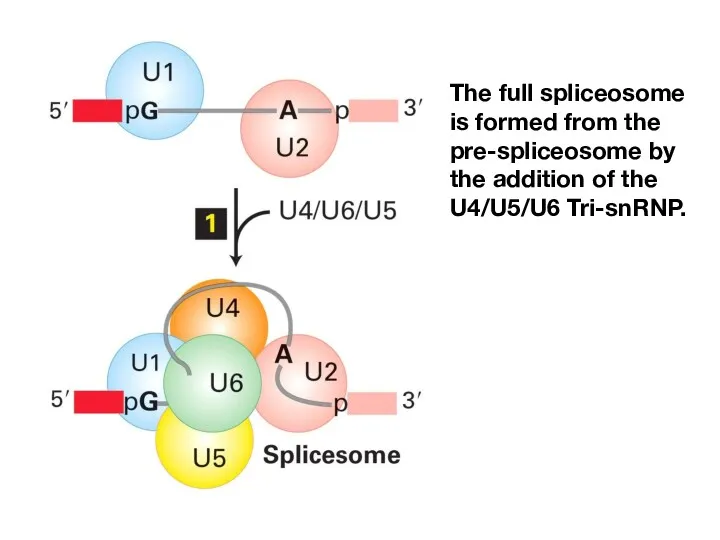
The full spliceosome is formed from the pre-spliceosome by the addition
The full spliceosome is formed from the pre-spliceosome by the addition
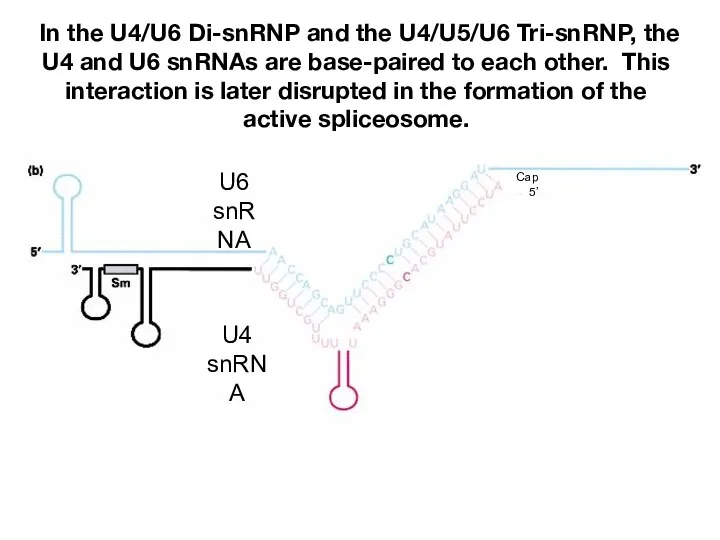
In the U4/U6 Di-snRNP and the U4/U5/U6 Tri-snRNP, the U4 and
In the U4/U6 Di-snRNP and the U4/U5/U6 Tri-snRNP, the U4 and
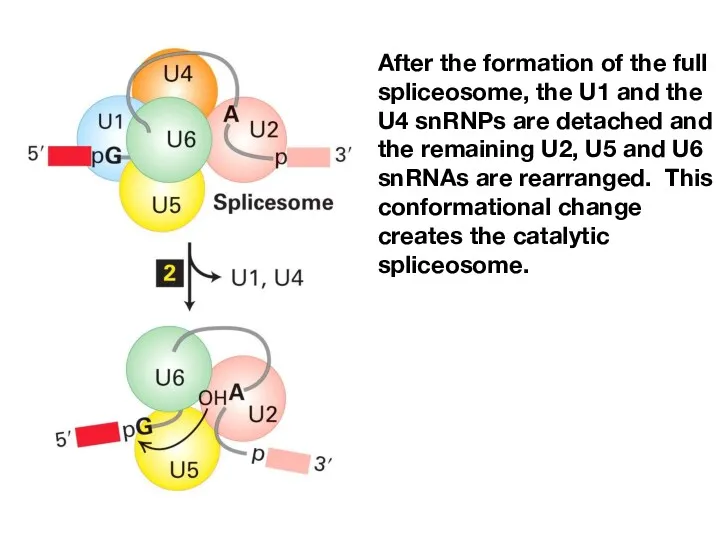
After the formation of the full spliceosome, the U1 and the
After the formation of the full spliceosome, the U1 and the
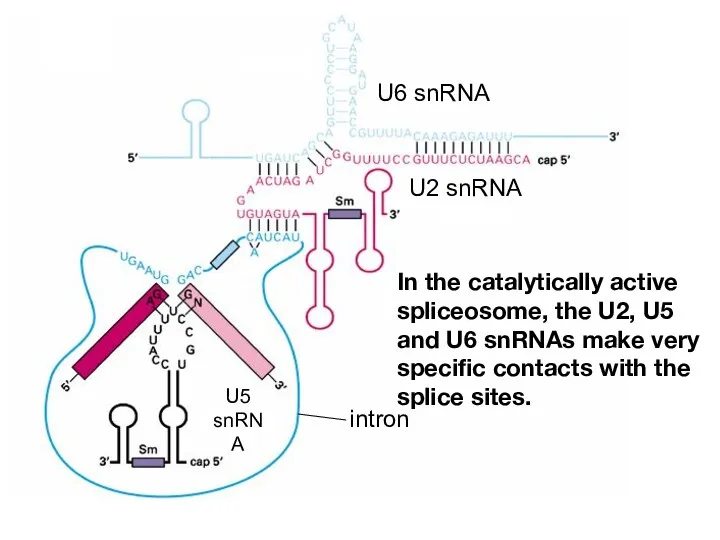
In the catalytically active spliceosome, the U2, U5 and U6 snRNAs
In the catalytically active spliceosome, the U2, U5 and U6 snRNAs
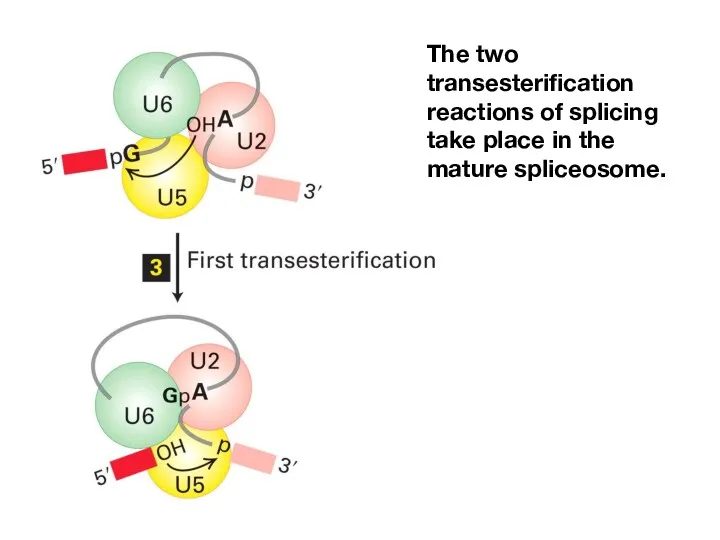
The two transesterification reactions of splicing take place in the mature
The two transesterification reactions of splicing take place in the mature
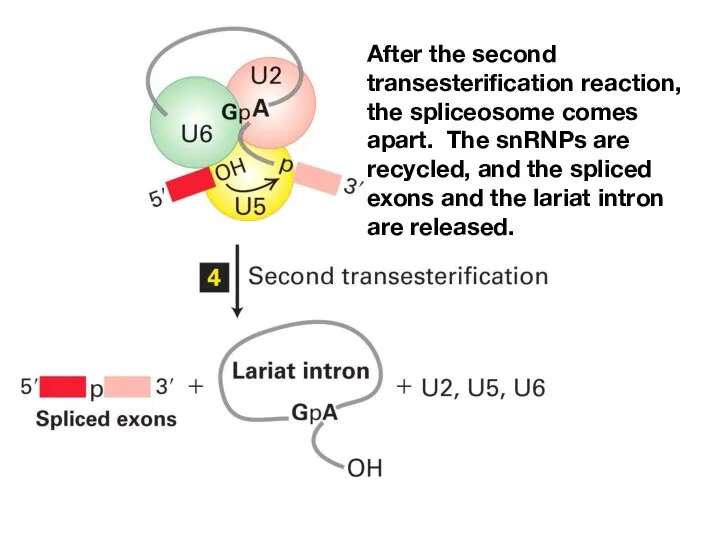
After the second transesterification reaction, the spliceosome comes apart. The snRNPs
After the second transesterification reaction, the spliceosome comes apart. The snRNPs
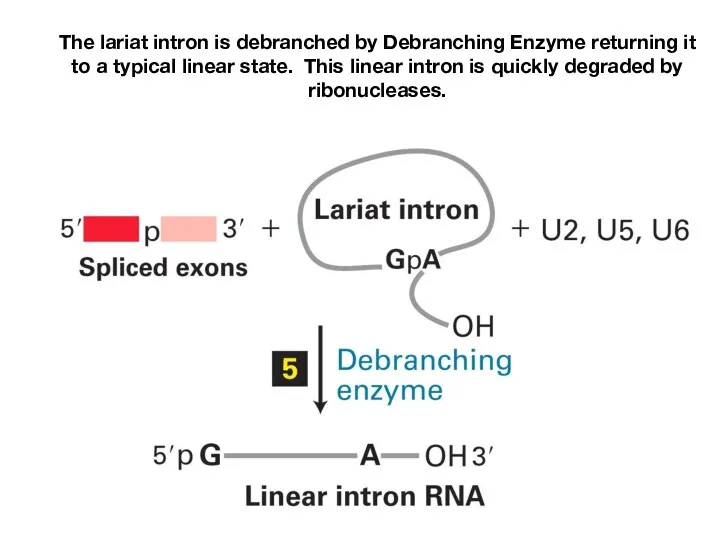
The lariat intron is debranched by Debranching Enzyme returning it to
The lariat intron is debranched by Debranching Enzyme returning it to
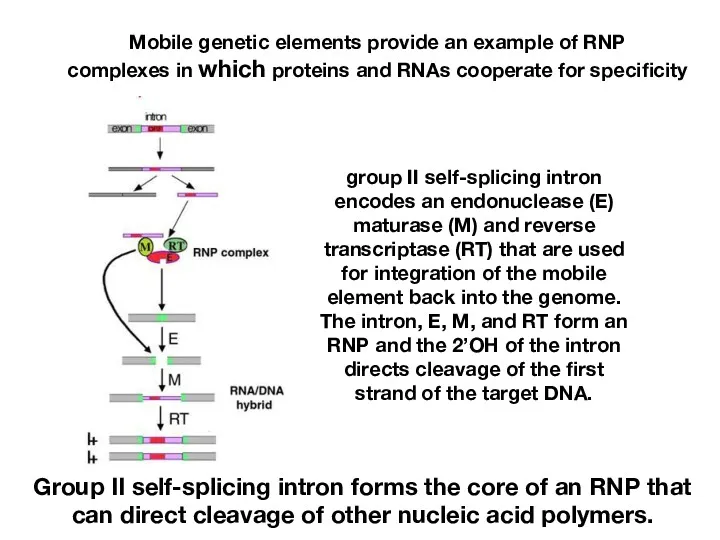
Mobile genetic elements provide an example of RNP
complexes in which proteins
Mobile genetic elements provide an example of RNP
complexes in which proteins
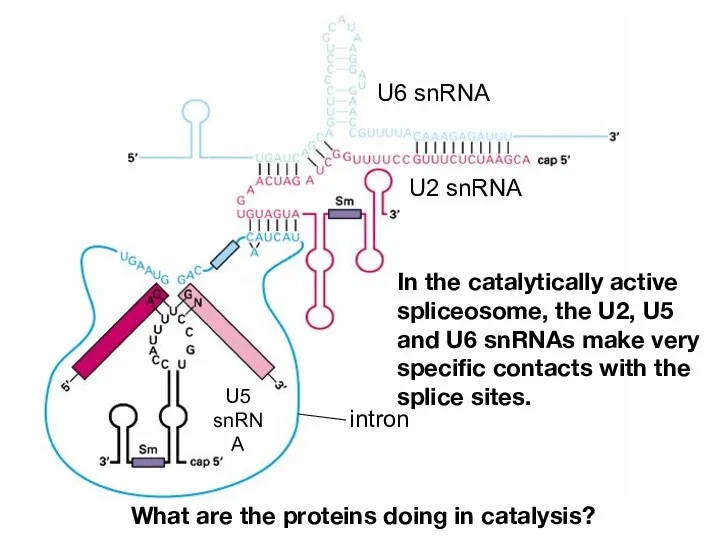
In the catalytically active spliceosome, the U2, U5 and U6 snRNAs
In the catalytically active spliceosome, the U2, U5 and U6 snRNAs
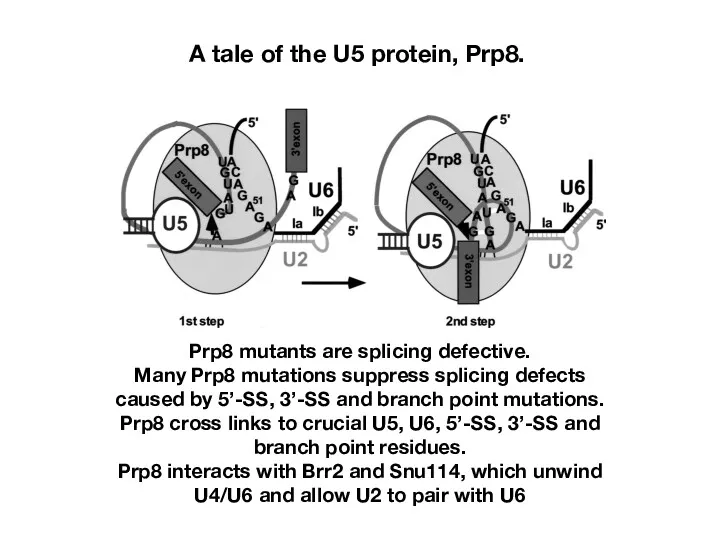
A tale of the U5 protein, Prp8.
Prp8 mutants are splicing defective.
Many
A tale of the U5 protein, Prp8.
Prp8 mutants are splicing defective.
Many
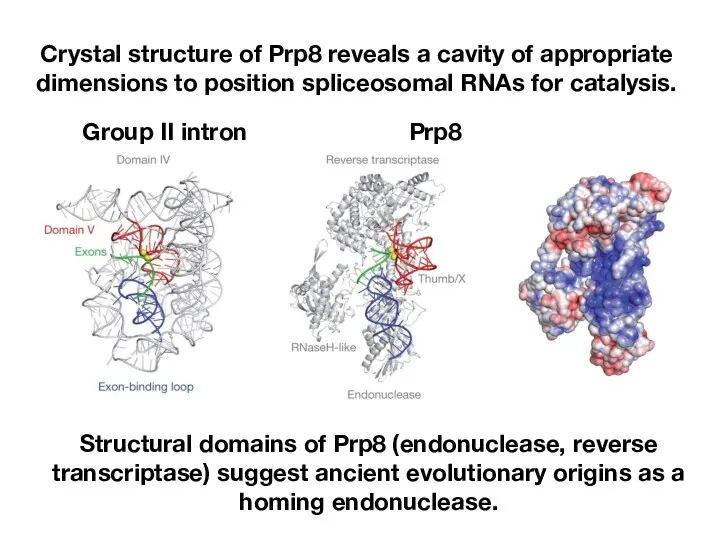
Crystal structure of Prp8 reveals a cavity of appropriate
dimensions to position
Crystal structure of Prp8 reveals a cavity of appropriate
dimensions to position
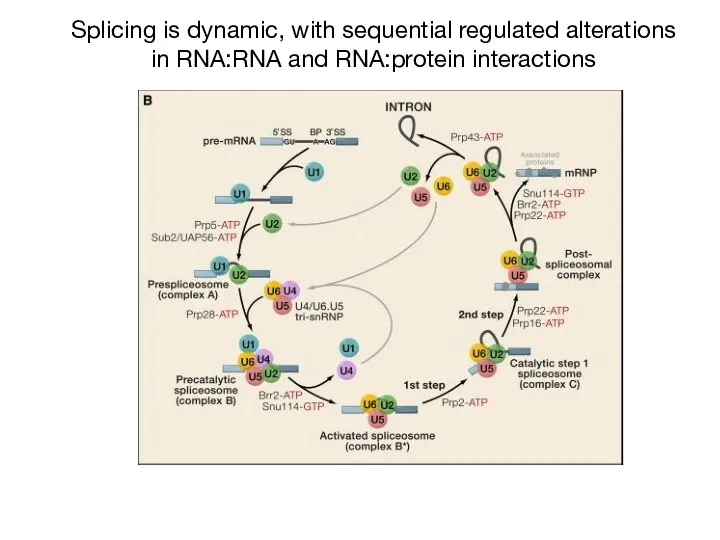
Splicing is dynamic, with sequential regulated alterations
in RNA:RNA and RNA:protein interactions
Splicing is dynamic, with sequential regulated alterations
in RNA:RNA and RNA:protein interactions
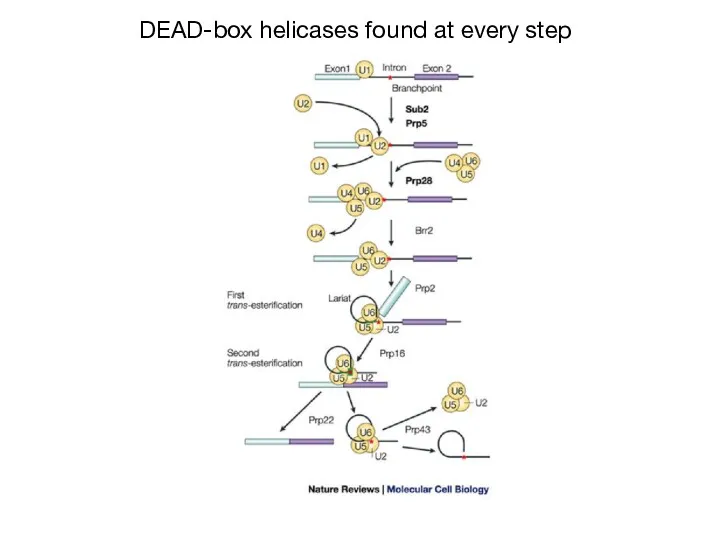
DEAD-box helicases found at every step
DEAD-box helicases found at every step
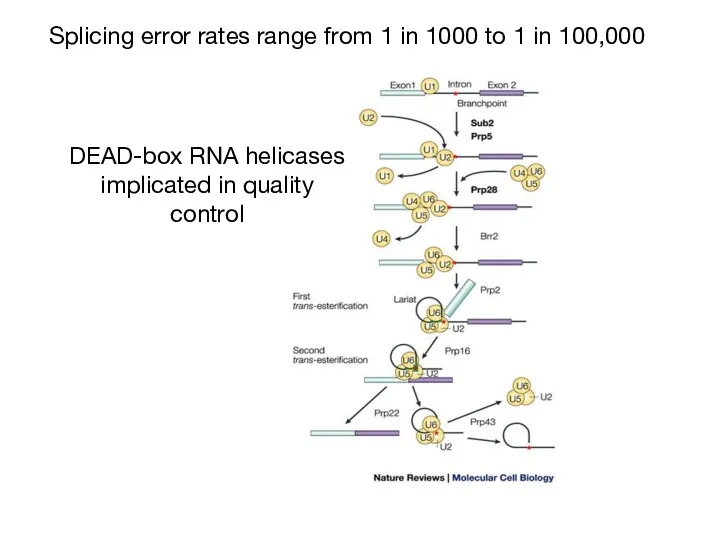
Splicing error rates range from 1 in 1000 to 1 in
Splicing error rates range from 1 in 1000 to 1 in

Monomeric (vs. “AAA” ATPases)
RNA-dependent ATPases
~300 aa domain with 7 signature motifs
Monomeric (vs. “AAA” ATPases)
RNA-dependent ATPases
~300 aa domain with 7 signature motifs

The story of one helicase: PRP16
Prp16 is required for the second
The story of one helicase: PRP16
Prp16 is required for the second
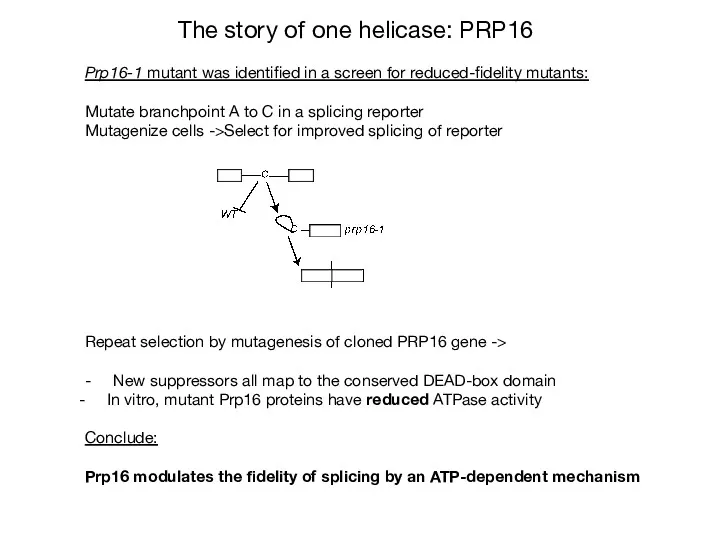
The story of one helicase: PRP16
Prp16-1 mutant was identified in a
The story of one helicase: PRP16
Prp16-1 mutant was identified in a
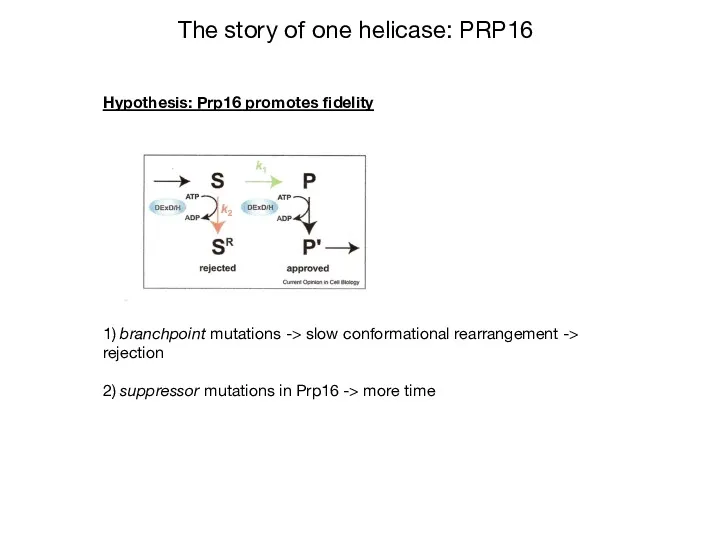
Hypothesis: Prp16 promotes fidelity
1) branchpoint mutations -> slow conformational rearrangement ->
Hypothesis: Prp16 promotes fidelity
1) branchpoint mutations -> slow conformational rearrangement ->
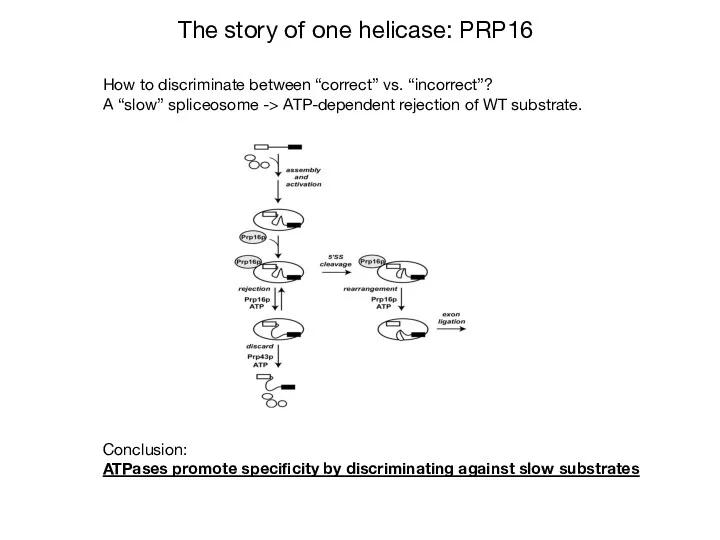
How to discriminate between “correct” vs. “incorrect”?
A “slow” spliceosome -> ATP-dependent
How to discriminate between “correct” vs. “incorrect”?
A “slow” spliceosome -> ATP-dependent
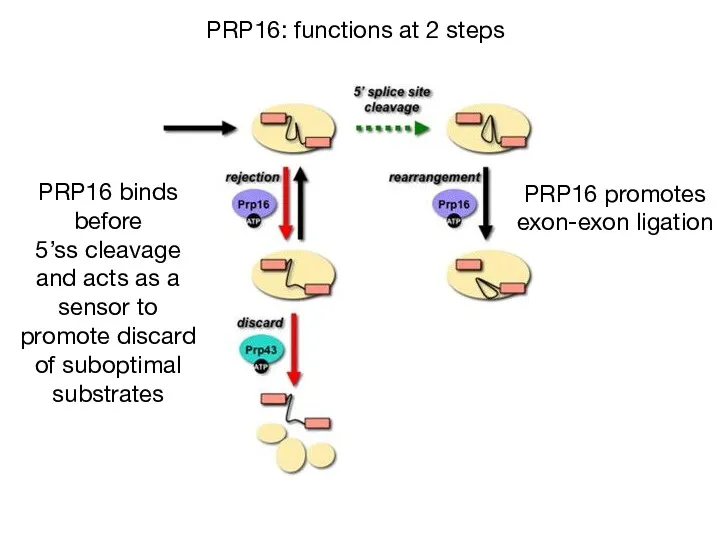
PRP16: functions at 2 steps
PRP16 binds before
5’ss cleavage and acts as
PRP16: functions at 2 steps
PRP16 binds before
5’ss cleavage and acts as

Questions
How are the splice sites identified?
How are the intervening
Questions
How are the splice sites identified?
How are the intervening

How are the splice sites identified?
In higher eukaryotes, there isn’t
How are the splice sites identified?
In higher eukaryotes, there isn’t

How are the splice sites identified?
Minor spliceosome, consists of U11,
How are the splice sites identified?
Minor spliceosome, consists of U11,
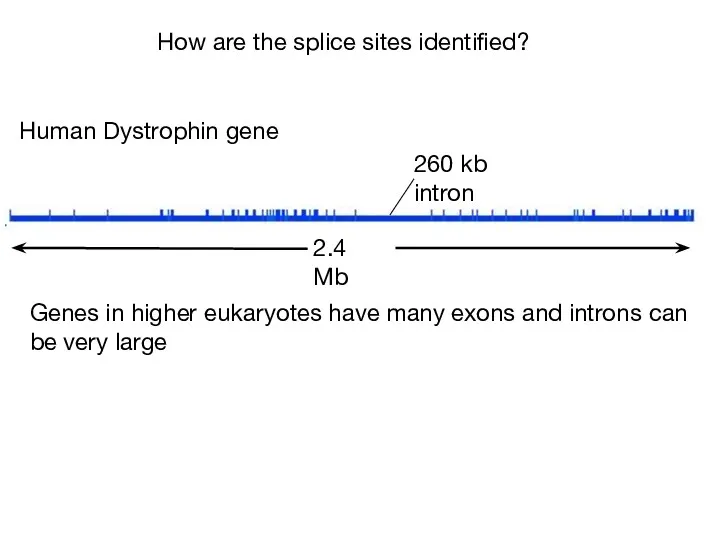
2.4 Mb
260 kb intron
Human Dystrophin gene
Genes in higher eukaryotes have many
2.4 Mb
260 kb intron
Human Dystrophin gene
Genes in higher eukaryotes have many
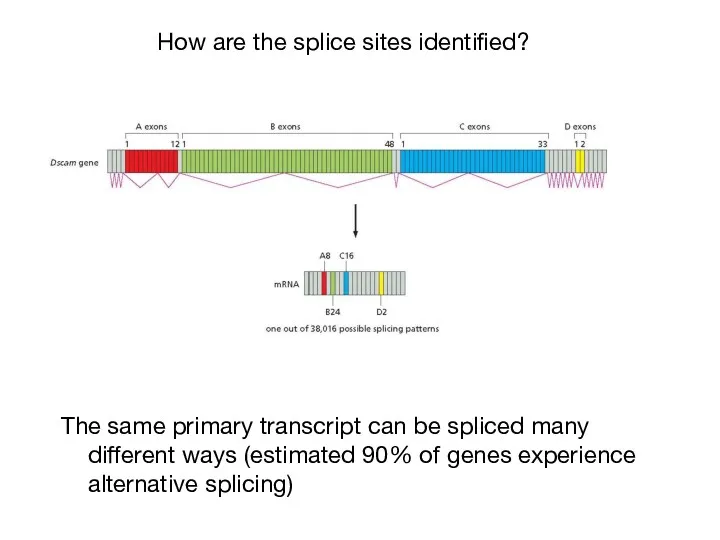
The same primary transcript can be spliced many different ways (estimated
The same primary transcript can be spliced many different ways (estimated
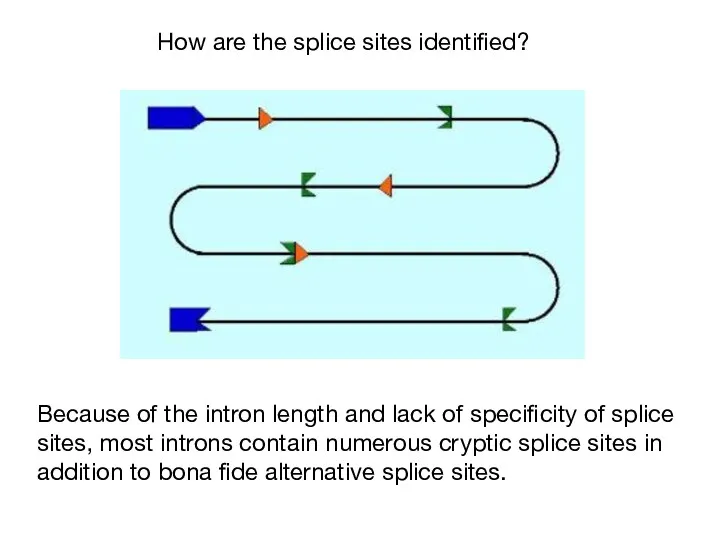
Because of the intron length and lack of specificity of splice
Because of the intron length and lack of specificity of splice
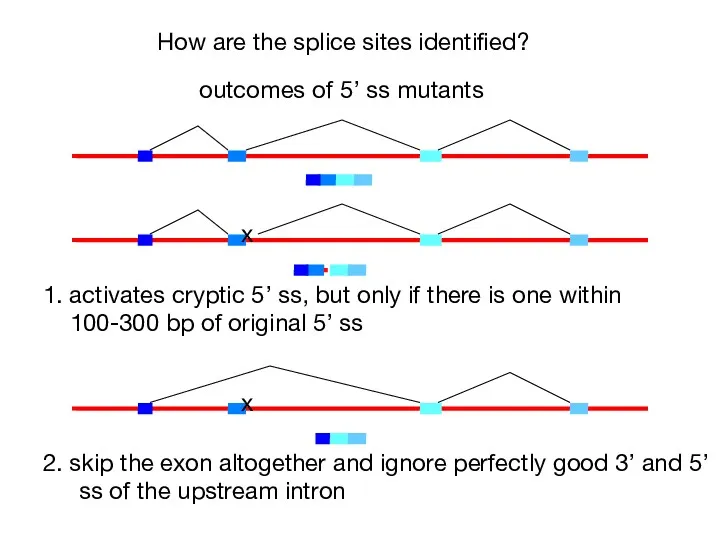
How are the splice sites identified?
x
outcomes of 5’ ss mutants
1.
How are the splice sites identified?
x
outcomes of 5’ ss mutants
1.
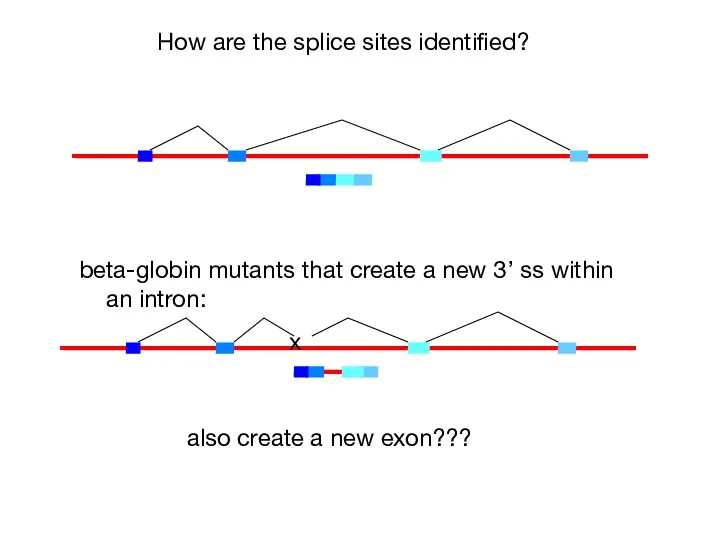
How are the splice sites identified?
beta-globin mutants that create a
How are the splice sites identified?
beta-globin mutants that create a
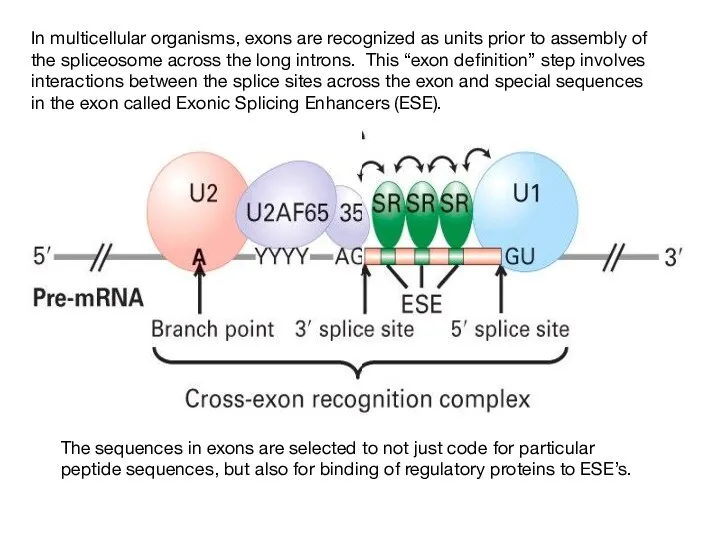
In multicellular organisms, exons are recognized as units prior to assembly
In multicellular organisms, exons are recognized as units prior to assembly
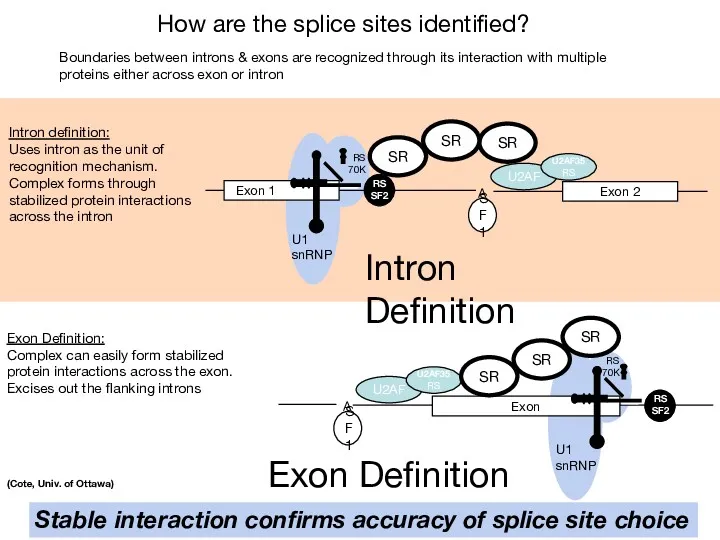
How are the splice sites identified?
A
U2AF
Exon 1
U1
snRNP
RS
70K
RS
SF2
U2AF35
RS
SF1
Exon 2
SR
Intron definition:
Uses
How are the splice sites identified?
A
U2AF
Exon 1
U1
snRNP
RS
70K
RS
SF2
U2AF35
RS
SF1
Exon 2
SR
Intron definition:
Uses
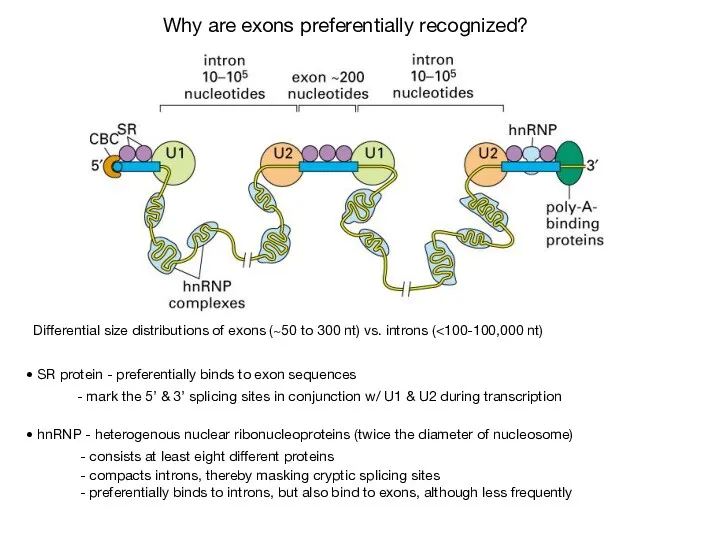
Differential size distributions of exons (~50 to 300 nt) vs. introns
Differential size distributions of exons (~50 to 300 nt) vs. introns
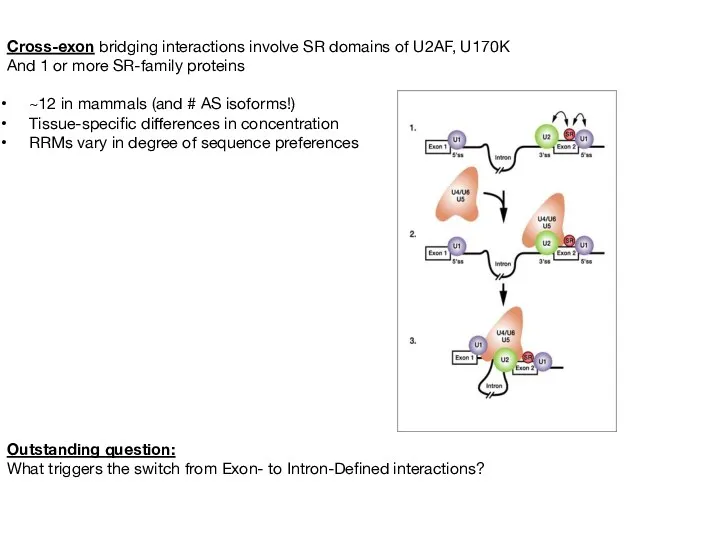
Cross-exon bridging interactions involve SR domains of U2AF, U170K
And 1 or
Cross-exon bridging interactions involve SR domains of U2AF, U170K
And 1 or
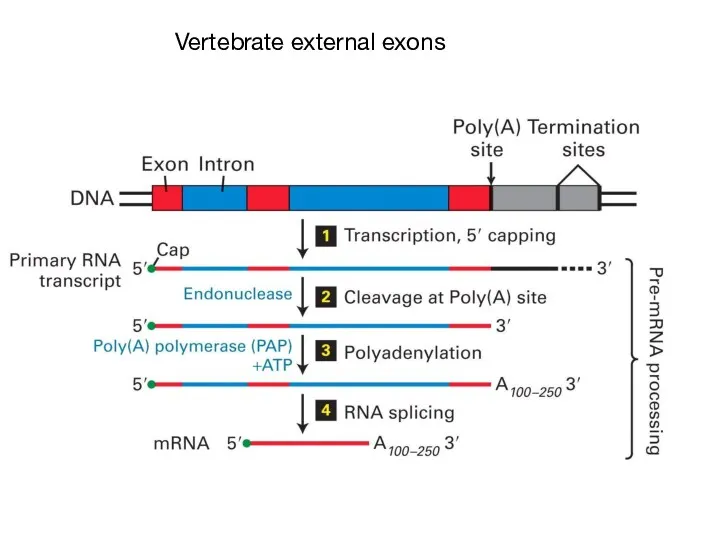
Vertebrate external exons
Vertebrate external exons
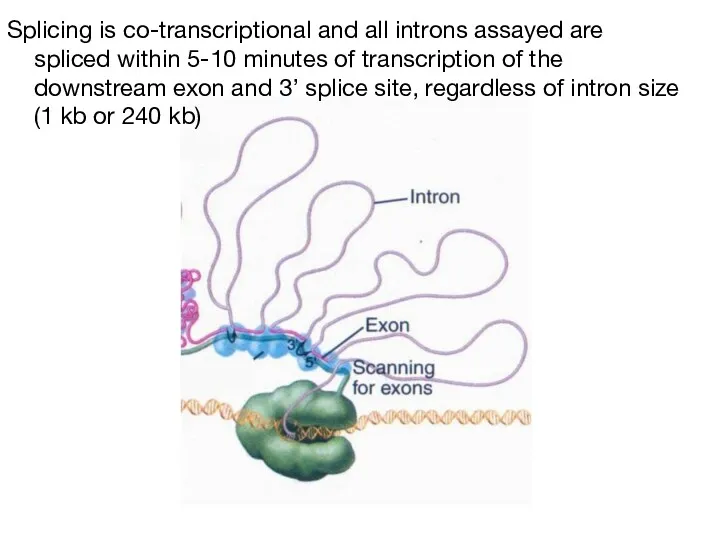
Splicing is co-transcriptional and all introns assayed are spliced within 5-10
Splicing is co-transcriptional and all introns assayed are spliced within 5-10

Defining an exon involves the specific stabilization or destabilization of splice
Defining an exon involves the specific stabilization or destabilization of splice

Regulation of alternative splicing involves the specific stabilization or destabilization
Regulation of alternative splicing involves the specific stabilization or destabilization
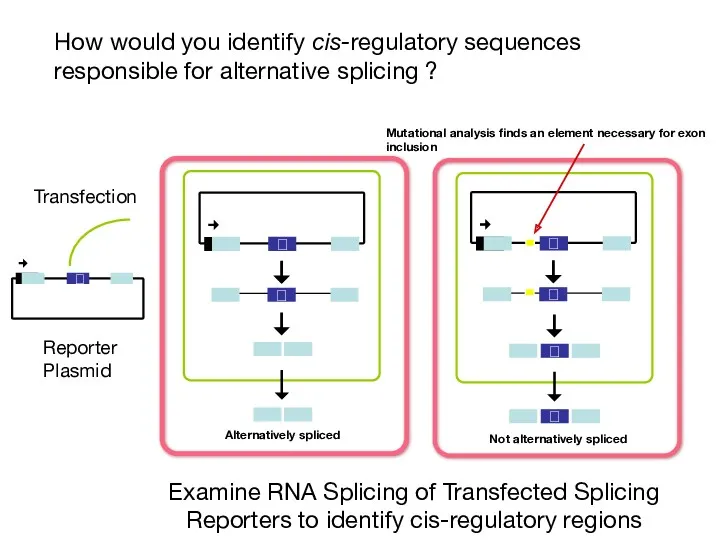
How would you identify cis-regulatory sequences responsible for alternative splicing ?
Examine
How would you identify cis-regulatory sequences responsible for alternative splicing ?
Examine

Four classes of splicing regulatory elements: Exonic Splicing Enhancers, Exonic Splicing
Four classes of splicing regulatory elements: Exonic Splicing Enhancers, Exonic Splicing
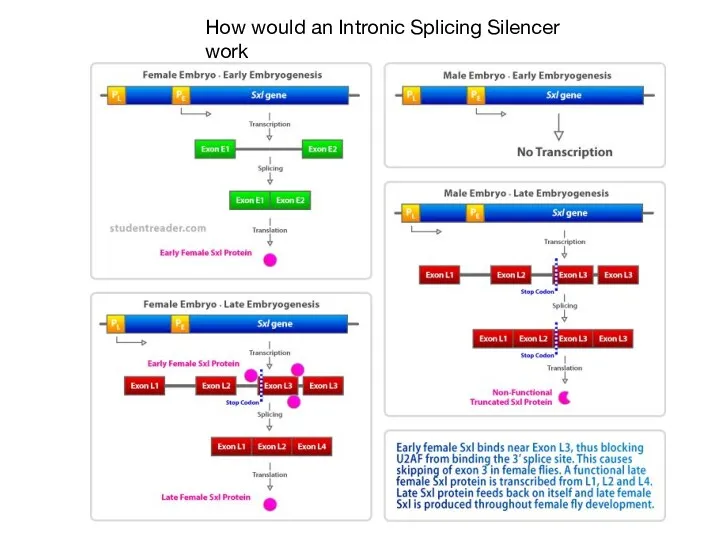
How would an Intronic Splicing Silencer work
How would an Intronic Splicing Silencer work
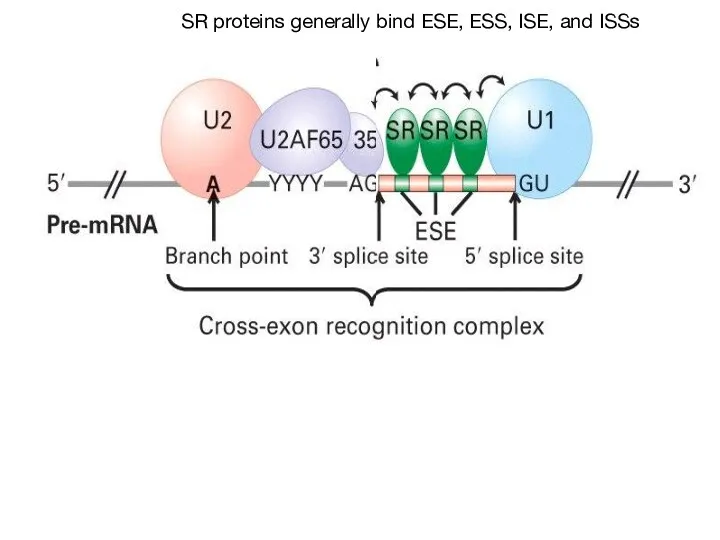
SR proteins generally bind ESE, ESS, ISE, and ISSs
SR proteins generally bind ESE, ESS, ISE, and ISSs
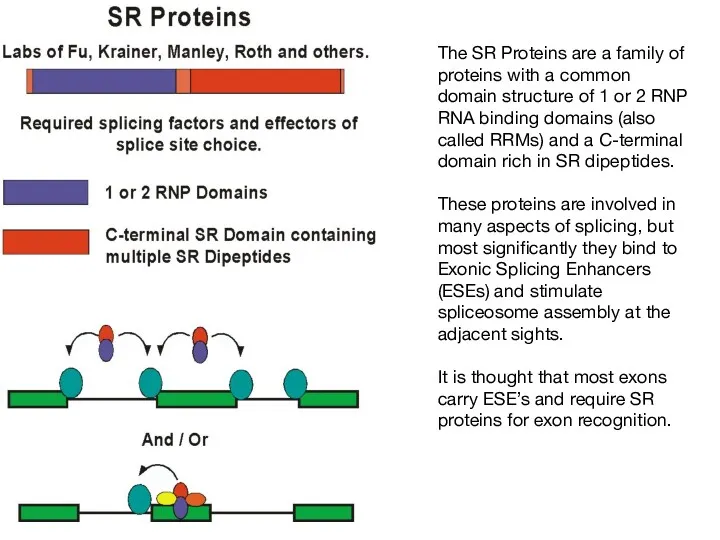
The SR Proteins are a family of proteins with a common
The SR Proteins are a family of proteins with a common
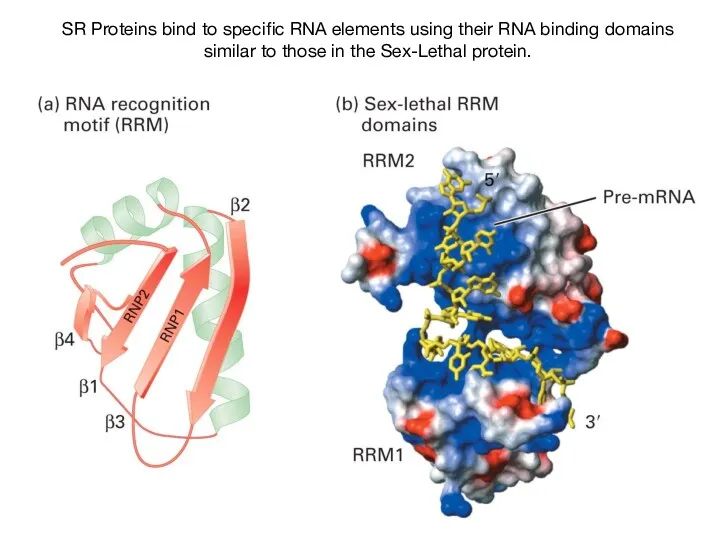
SR Proteins bind to specific RNA elements using their RNA binding
SR Proteins bind to specific RNA elements using their RNA binding

Characterization of an ESE and SR protein in flies
Sex differentiation in
Sex differentiation in
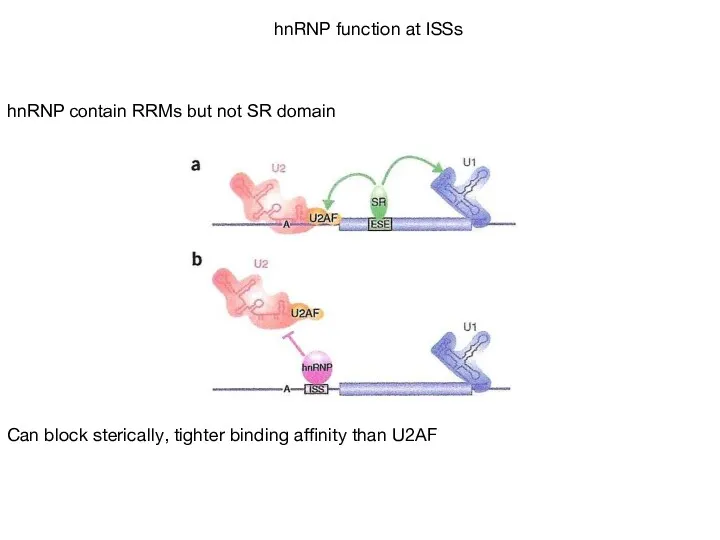
hnRNP contain RRMs but not SR domain
Can block sterically, tighter binding
hnRNP contain RRMs but not SR domain
Can block sterically, tighter binding
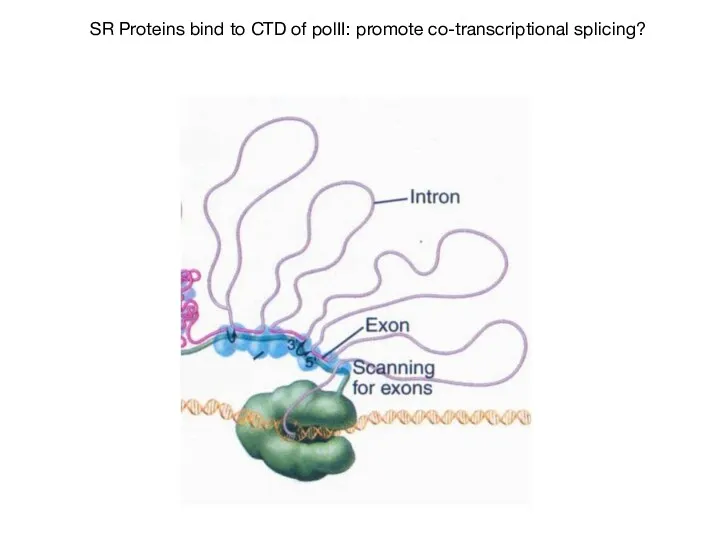
SR Proteins bind to CTD of polII: promote co-transcriptional splicing?
SR Proteins bind to CTD of polII: promote co-transcriptional splicing?
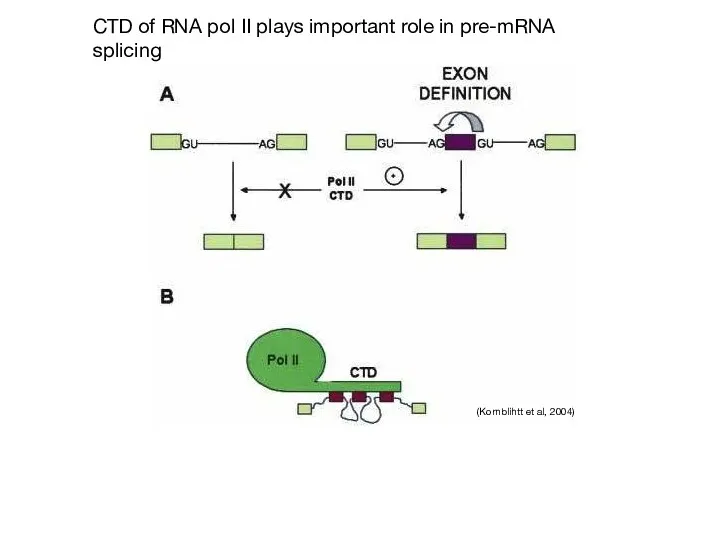
CTD of RNA pol II plays important role in pre-mRNA splicing
(Kornblihtt
CTD of RNA pol II plays important role in pre-mRNA splicing
(Kornblihtt
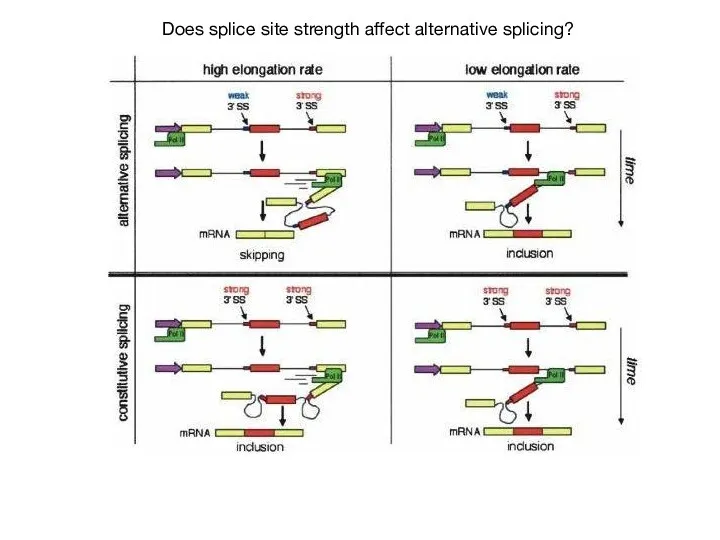
Does splice site strength affect alternative splicing?
Does splice site strength affect alternative splicing?
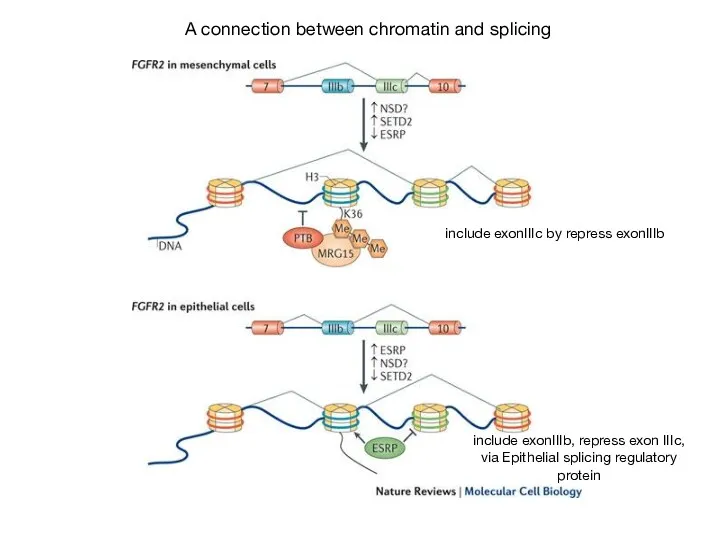
A connection between chromatin and splicing
include exonIIIc by repress exonIIIb
include exonIIIb,
A connection between chromatin and splicing
include exonIIIc by repress exonIIIb
include exonIIIb,

mRNA export - formation of an export competent mRNP
Sees formation of
mRNA export - formation of an export competent mRNP
Sees formation of
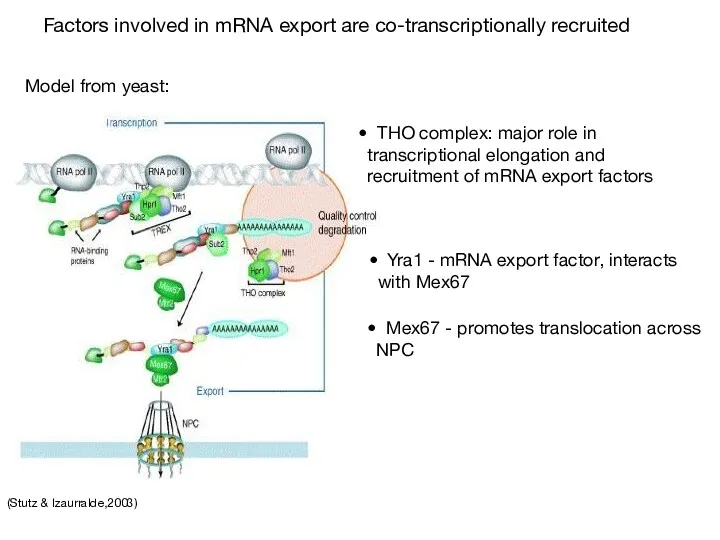
(Stutz & Izaurralde,2003)
Factors involved in mRNA export are co-transcriptionally recruited
THO
(Stutz & Izaurralde,2003)
Factors involved in mRNA export are co-transcriptionally recruited
THO
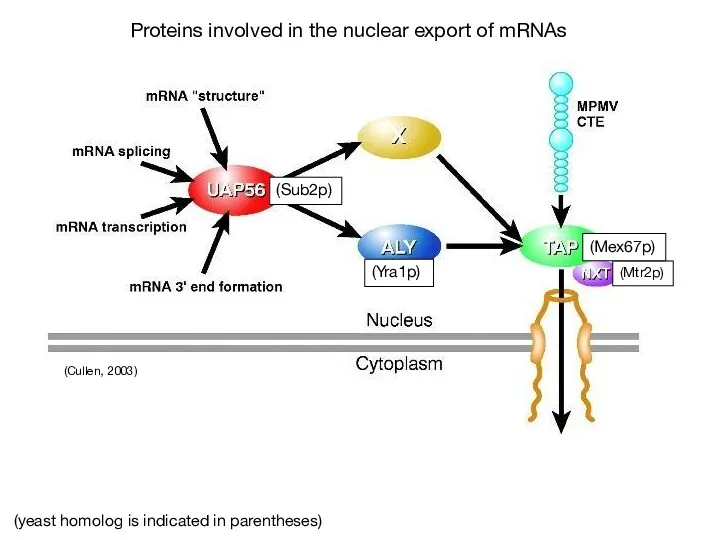
(Cullen, 2003)
(Sub2p)
(Yra1p)
(Mtr2p)
(Mex67p)
(yeast homolog is indicated in parentheses)
Proteins involved in the nuclear
(Cullen, 2003)
(Sub2p)
(Yra1p)
(Mtr2p)
(Mex67p)
(yeast homolog is indicated in parentheses)
Proteins involved in the nuclear
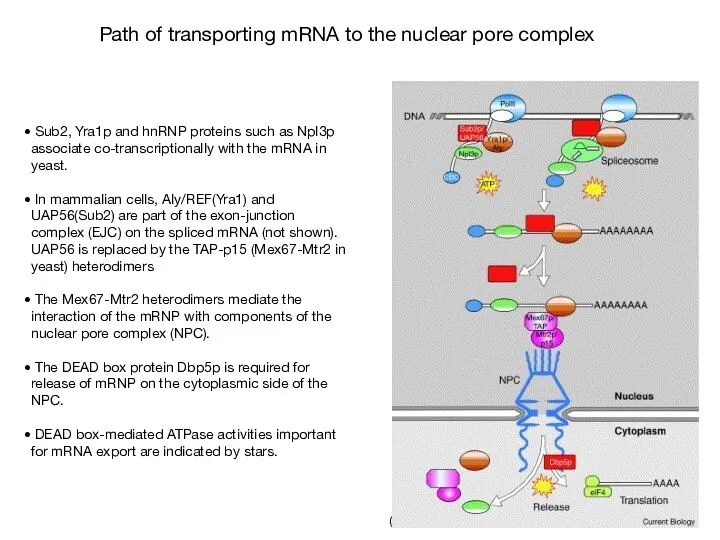
(Linder & Stutz, 2001)
Sub2, Yra1p and hnRNP proteins such as
(Linder & Stutz, 2001)
Sub2, Yra1p and hnRNP proteins such as
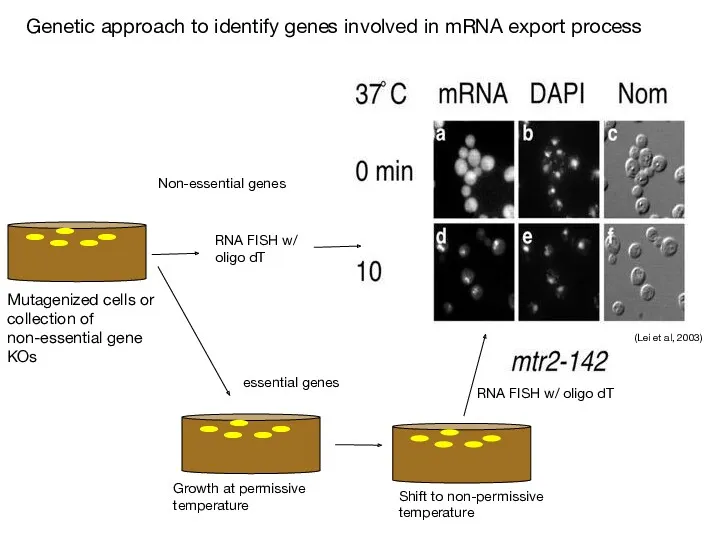
Genetic approach to identify genes involved in mRNA export process
(Lei et
Genetic approach to identify genes involved in mRNA export process
(Lei et
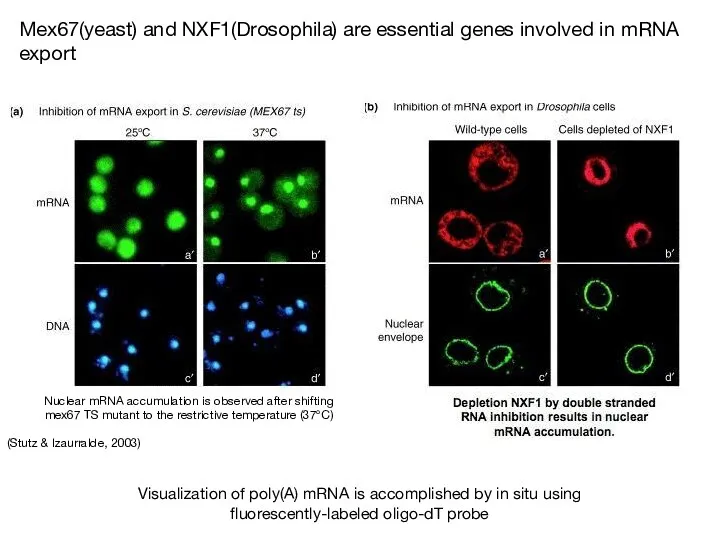
(Stutz & Izaurralde, 2003)
Nuclear mRNA accumulation is observed after shifting mex67
(Stutz & Izaurralde, 2003)
Nuclear mRNA accumulation is observed after shifting mex67
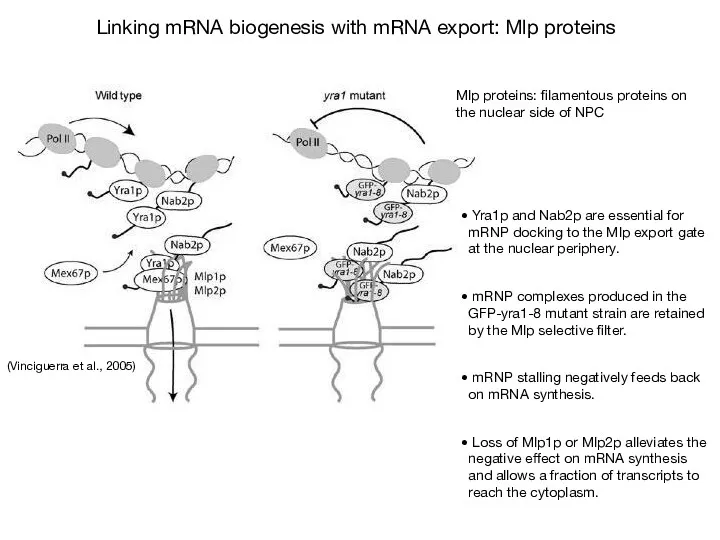
Yra1p and Nab2p are essential for mRNP docking to the
Yra1p and Nab2p are essential for mRNP docking to the
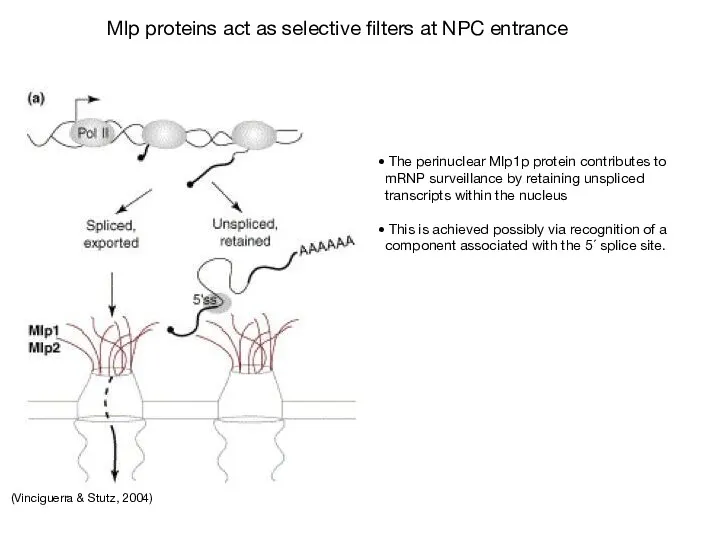
(Vinciguerra & Stutz, 2004)
The perinuclear Mlp1p protein contributes to mRNP
(Vinciguerra & Stutz, 2004)
The perinuclear Mlp1p protein contributes to mRNP
 Выделительная система организма. Гигиена почек
Выделительная система организма. Гигиена почек Перелётные птицы
Перелётные птицы Принципы применения БАД
Принципы применения БАД Манул. Кот, победивший время
Манул. Кот, победивший время Сорные растения и методы борьбы с ними
Сорные растения и методы борьбы с ними Строение цветка
Строение цветка Строение растительной клетки
Строение растительной клетки Участие прокариот в превращениях соединений биогенных элементов
Участие прокариот в превращениях соединений биогенных элементов Индивидуальное развитие животных
Индивидуальное развитие животных Органы кровообращения. Строение сердца
Органы кровообращения. Строение сердца Деление клетки. Митоз
Деление клетки. Митоз Как паук костюмчик менял
Как паук костюмчик менял Вегетативное размножение растений
Вегетативное размножение растений Микробиология. Болезнетворные (патогенные) микроорганизмы
Микробиология. Болезнетворные (патогенные) микроорганизмы Эволюция пищеварительной системы животных
Эволюция пищеварительной системы животных Человек как результат биологической и социокультурной эволюции
Человек как результат биологической и социокультурной эволюции Вегетативное размножение.
Вегетативное размножение. Химическая организация клетки
Химическая организация клетки Ткани растений
Ткани растений Умови та види зберігання картоплі
Умови та види зберігання картоплі Біорізноманіття нашої планети як наслідок еволюції
Біорізноманіття нашої планети як наслідок еволюції Свет как экологический фактор
Свет как экологический фактор Wonderful wild animals
Wonderful wild animals Цианобактерии. Отличия от бактерий
Цианобактерии. Отличия от бактерий Водорастворимые витамины
Водорастворимые витамины Углеводы
Углеводы Пчелы
Пчелы Способы размножения растений. Размножение споровых растений
Способы размножения растений. Размножение споровых растений