- Развитие до имплантации

Содержание
- 2. Мышь от оплодотворения до имплантации: 1 – 5 сутки развития
- 3. Эмбрионы мыши и человека
- 4. Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского генома (генома ооцита) к геному
- 5. У большинства видов животных процесс MZT медленный: в течение многих клеточных циклов (до 2-3 десятков циклов)
- 6. Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC 2015 August 11. Возрастание уровня
- 7. Изменение экспрессии в раннем развитии эмбрионов мыши
- 8. Активация генома эмбриона - участие материнских генов Филогенетическая дистанция между белками ZAR1 у разных видов позвоночных:
- 9. Профиль экспрессии Zscan4 во время доимплантационного развития: А: whole mount in situ hybridization; В: qRT-PCR analysis
- 10. Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and ES cells by the whole
- 11. Определение 1: Тотипотентность – способность клетки развиваться в целый организм Определение 2: Тотипотентность - способность клетки
- 12. После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза), (в том числе DNMTs, Ago2)
- 13. Выбор судьбы клетками раннего эмбриона: решение в 2 этапа 1 этап: внутри или снаружи (вопрос: за
- 15. Дифференцировка бластоцисты: ОСТ-4
- 16. 32 бластомера и более Компактизация и формирование морулы
- 17. Период компактизации Переход от 8 к 16 Переход от 16 к 32 Клеток мало для формирования
- 18. Компактизация и поляризация клеток эмбриона: как круглые клетки становятся плоскими? Fierro-González et al., Cadherin-dependent filopodia control
- 19. Протеин-киназа Hippo (у млекопитающих называется Mst1/2); Этот каскад контролирует пролиферацию и апоптоз; Мутации в этом гене
- 20. Hippo сигналинг: преобразование позиционной информации в молекулярный каскад Gata3 – ранний маркер клеток ТЭ Oct4, Sox2
- 21. Становление полярности эмбриона на стадиях дробления: апикально-базальные различия в строении кортикального слоя. Ajduk and Zernicka-Goetz Molecular
- 22. Hippo сигналинг недостаточен, необходима «внутренняя» среда Щелевые контакты Адгезионные контакты → сигналиг через цитоплазматические тирозин-киназы Фокальные
- 23. Дробление клеток раннего эмбриона: насколько клетки идентичны? Эксперименты по исследованию асимметрии ооцита и раннего эмбриона овцы
- 24. Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell stage mouse blastomeres divide either
- 25. Асимметричная локализация в клетках: aPKC, Par3 и ezrin ezrin aPKC aPKC Par3 Louvet et al., 1996
- 26. Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте Gardner, 2002
- 27. Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути REGULATORY NETWORKS IN EMBRYO-DERIVED PLURIPOTENT STEM CELLS
- 28. Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль в кавитации и формировании бластоцисты:
- 29. Классические представления: В ранней бластоцисте все клетки ВКМ с самого начала различаются по положению, в результате
- 30. FGFR1 Gata6, FGFR1-4 Преимплантационный эпибласт Примитивная эндодерма 2008: PrE - предшественники первичной энтодермы (гипобласта) мигрируют к
- 31. Bischoff et al., 2008 События в жизни преимплантационного эмбриона: что влияет на время их наступления? Эмбрион
- 32. Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых структур: клетки эмбриона до компактизации
- 34. Скачать презентацию
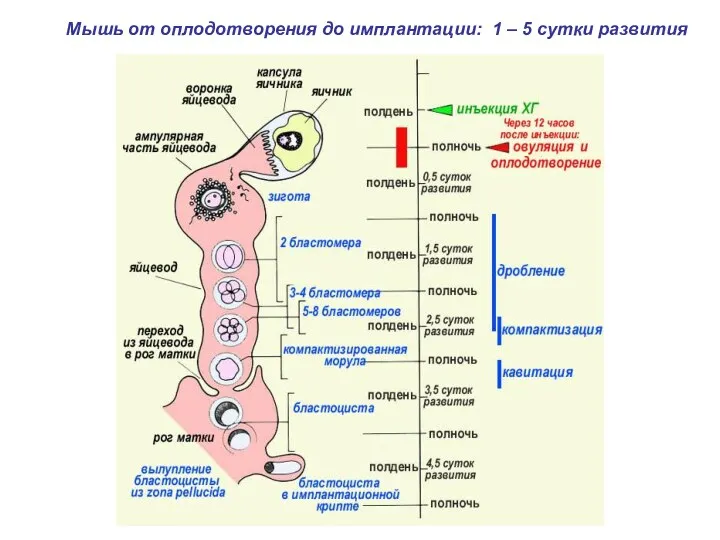
Мышь от оплодотворения до имплантации: 1 – 5 сутки развития
Мышь от оплодотворения до имплантации: 1 – 5 сутки развития

Эмбрионы мыши и человека
Эмбрионы мыши и человека
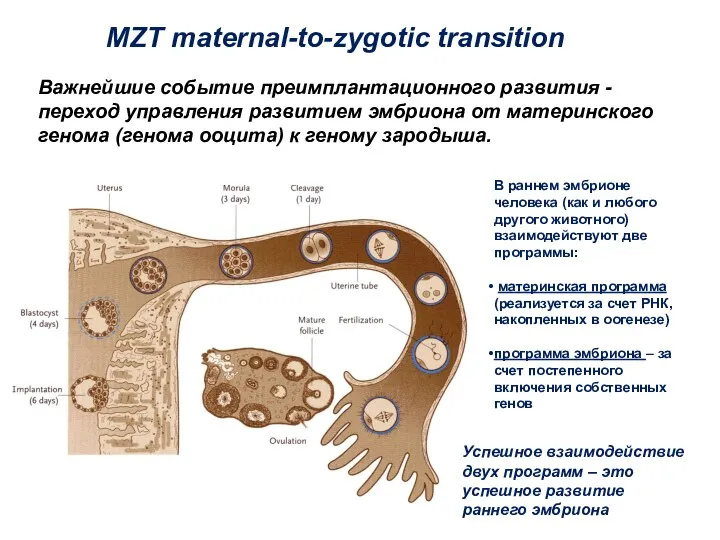
Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского
Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского
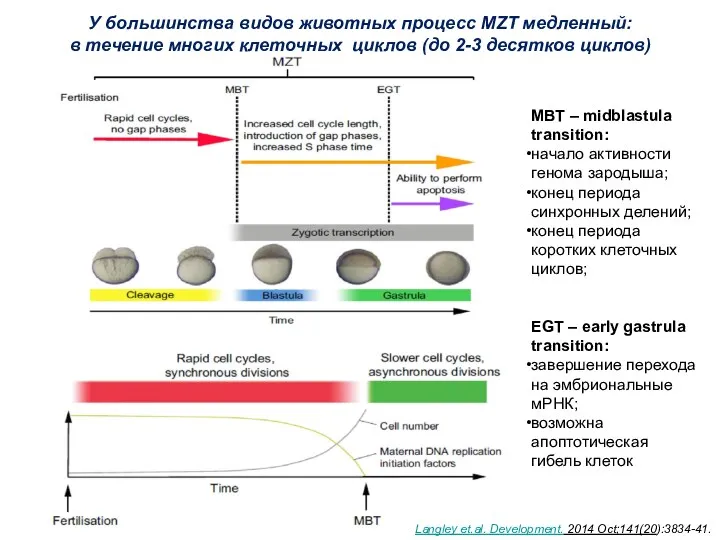
У большинства видов животных процесс MZT медленный:
в течение многих клеточных
У большинства видов животных процесс MZT медленный:
в течение многих клеточных
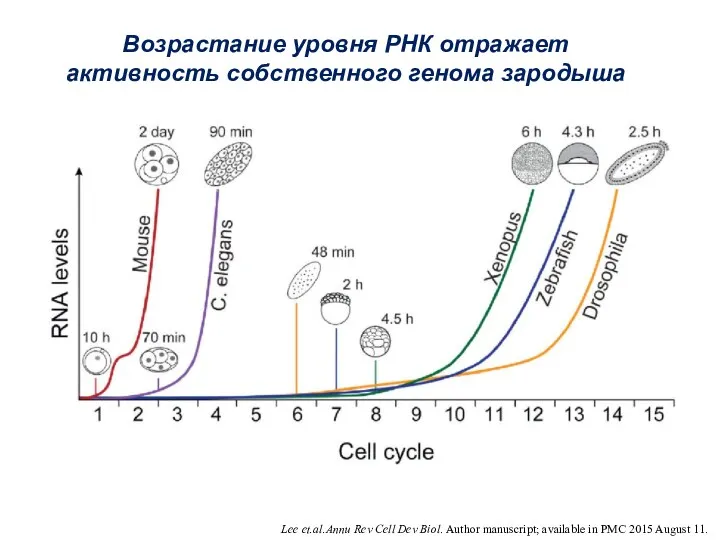
Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC
Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC
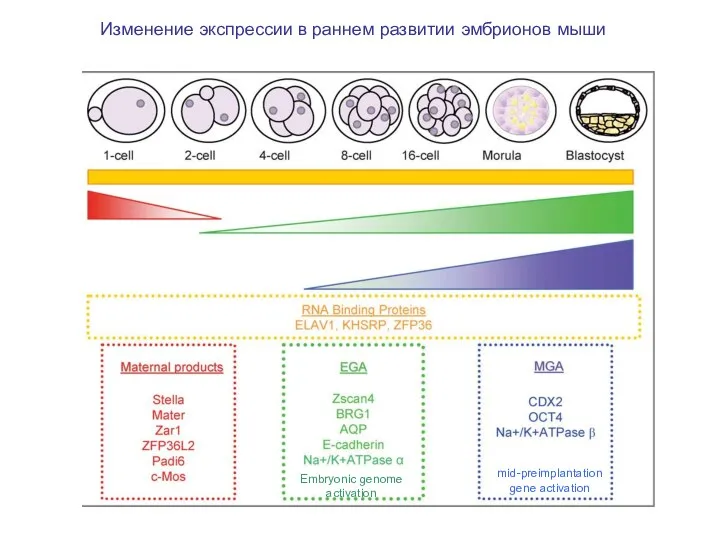
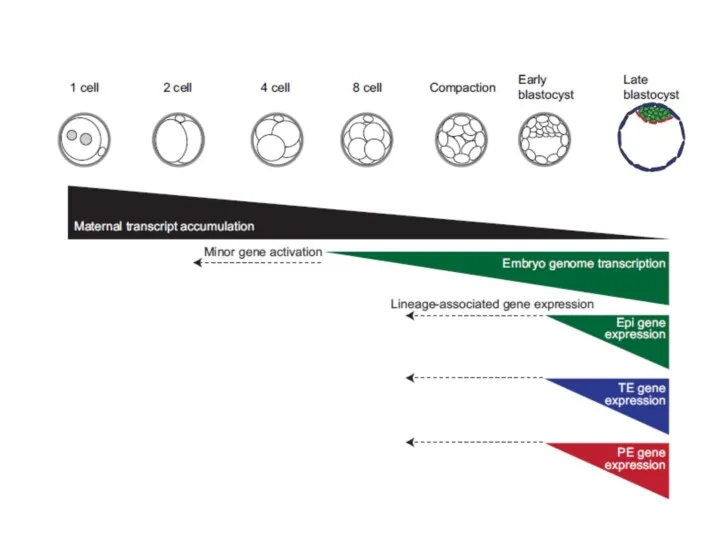
Изменение экспрессии в раннем развитии эмбрионов мыши
Изменение экспрессии в раннем развитии эмбрионов мыши

Активация генома эмбриона -
участие материнских генов
Филогенетическая дистанция между белками ZAR1 у
Активация генома эмбриона -
участие материнских генов
Филогенетическая дистанция между белками ZAR1 у
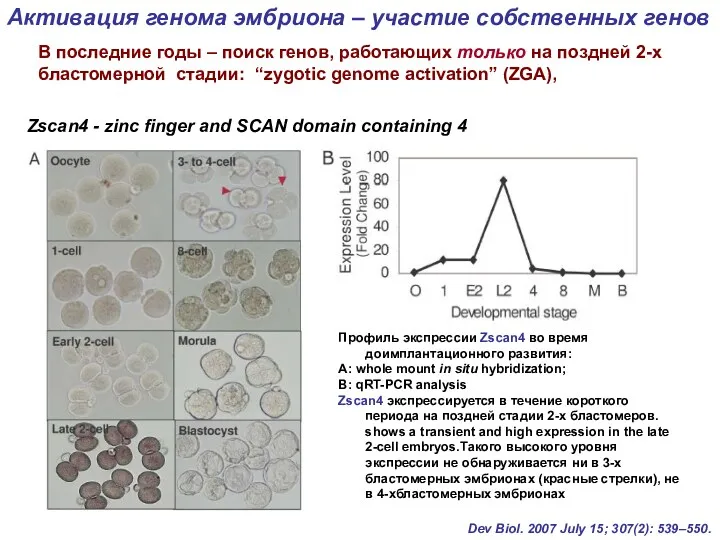
Профиль экспрессии Zscan4 во время доимплантационного развития:
А: whole mount in situ
Профиль экспрессии Zscan4 во время доимплантационного развития:
А: whole mount in situ
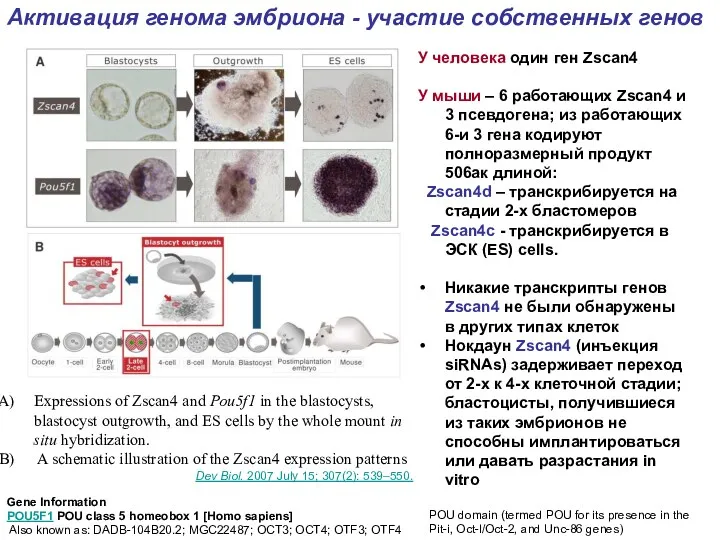
Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and
Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and

Определение 1: Тотипотентность – способность клетки развиваться в целый организм
Определение 2:
Определение 1: Тотипотентность – способность клетки развиваться в целый организм
Определение 2:
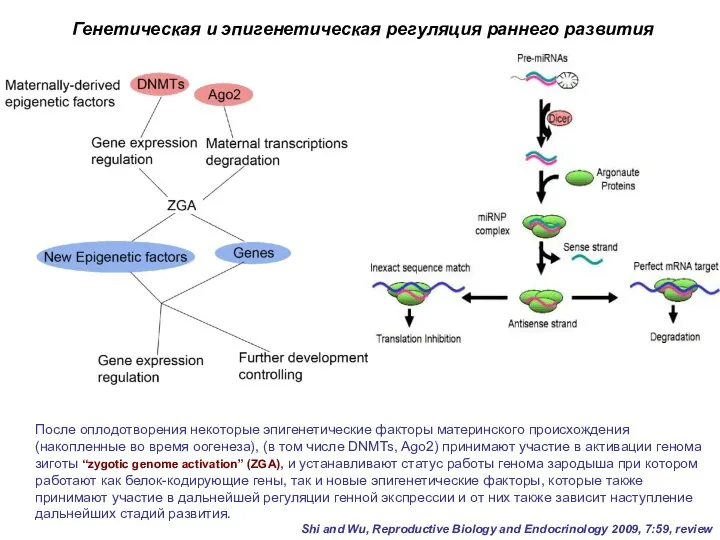
После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза),
После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза),
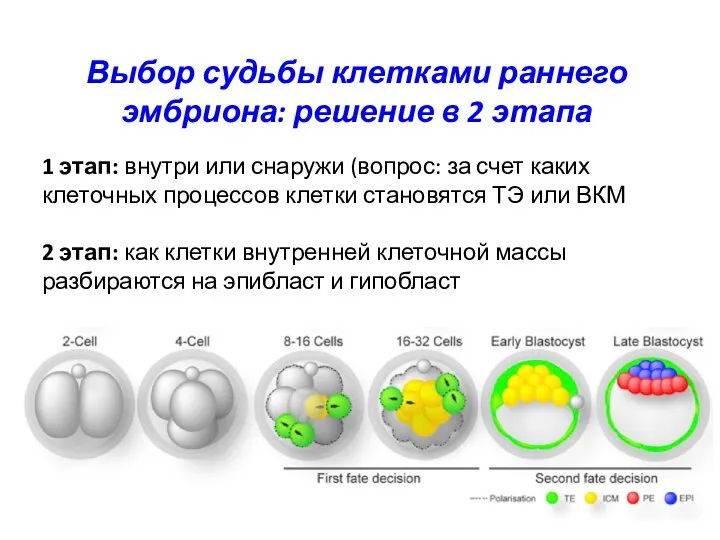
Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри
Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри
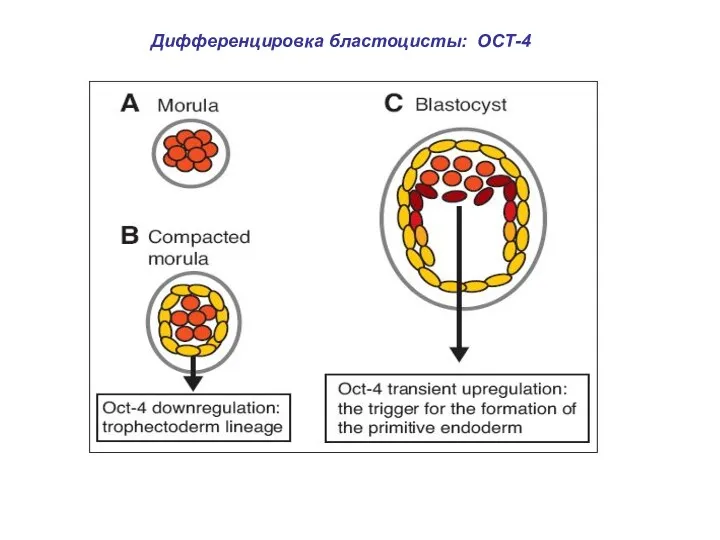
Дифференцировка бластоцисты: ОСТ-4
Дифференцировка бластоцисты: ОСТ-4
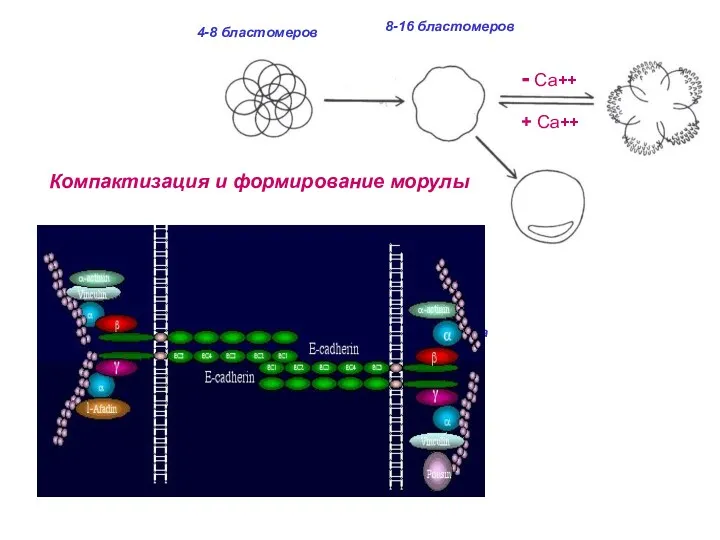
32 бластомера
и более
Компактизация и формирование морулы
32 бластомера
и более
Компактизация и формирование морулы
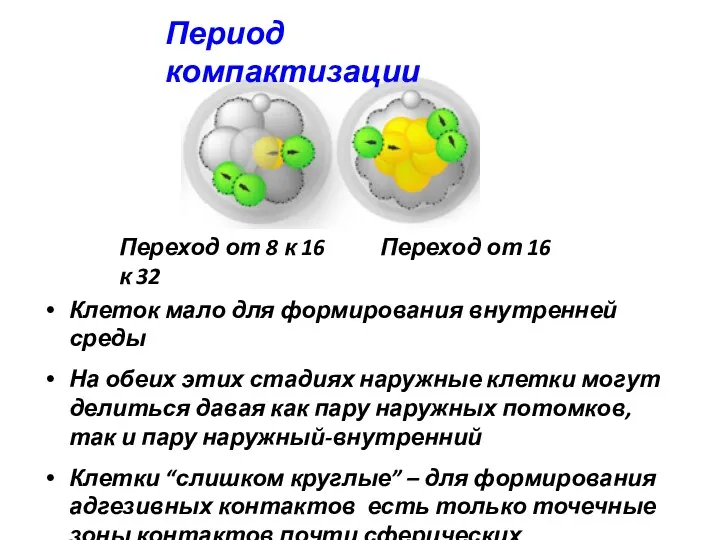
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
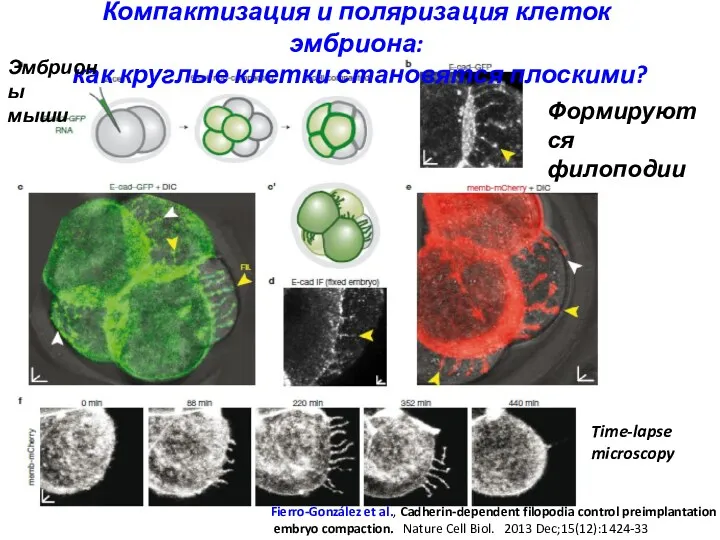
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
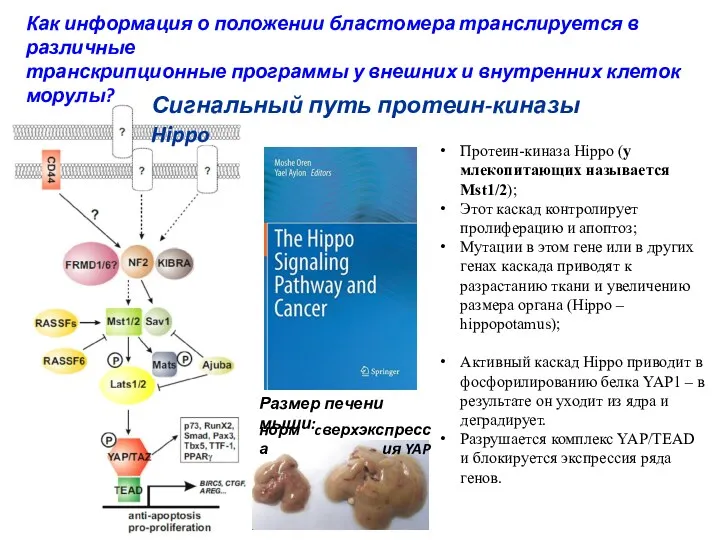
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
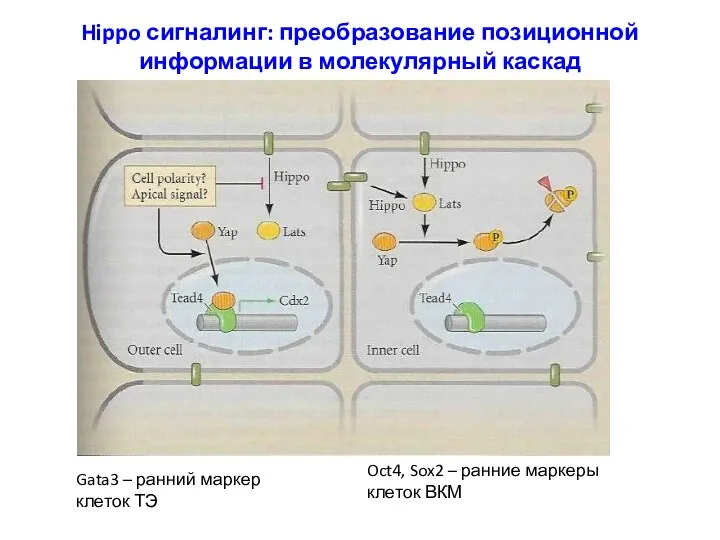
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер

Становление полярности эмбриона на стадиях дробления:
апикально-базальные различия в строении кортикального
Становление полярности эмбриона на стадиях дробления:
апикально-базальные различия в строении кортикального
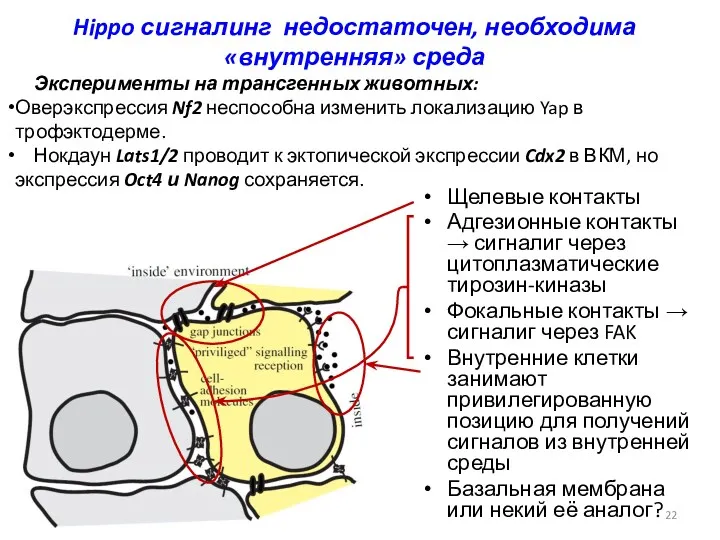
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через
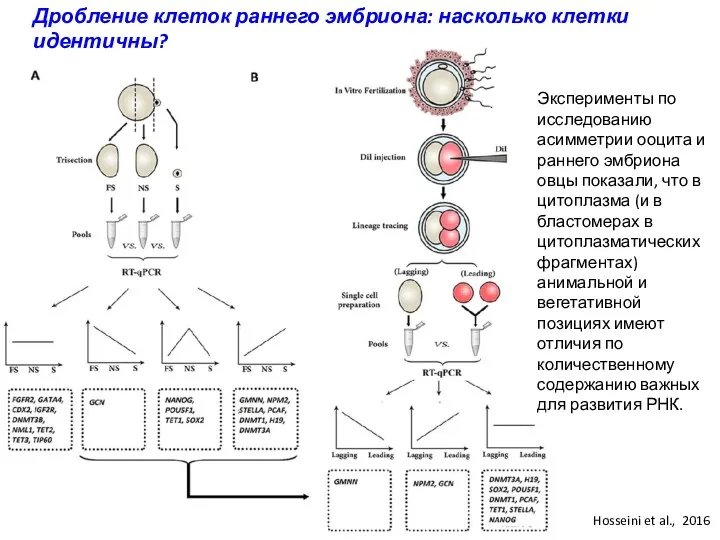
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
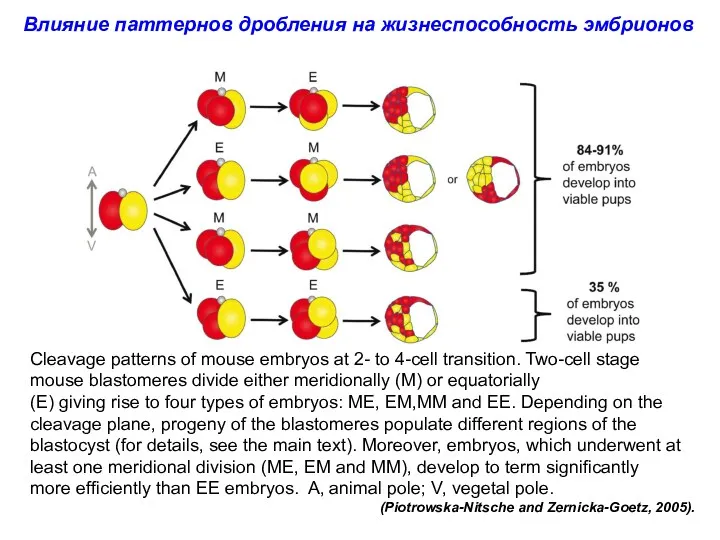
Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell
Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell
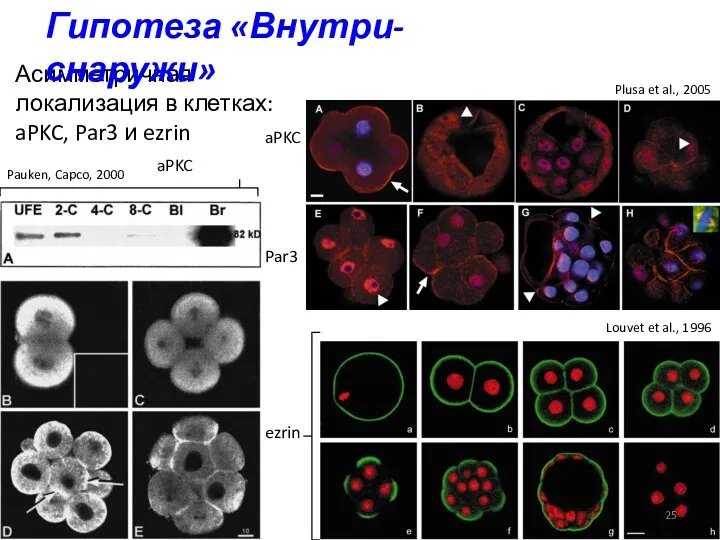
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
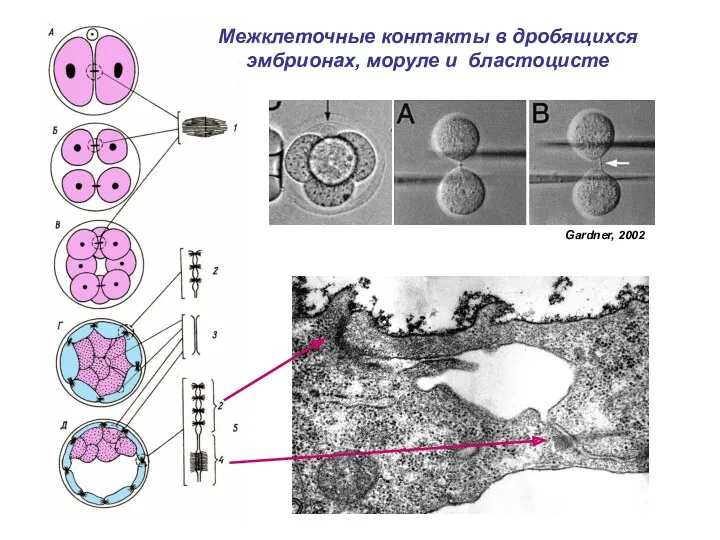
Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте
Gardner, 2002
Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте
Gardner, 2002
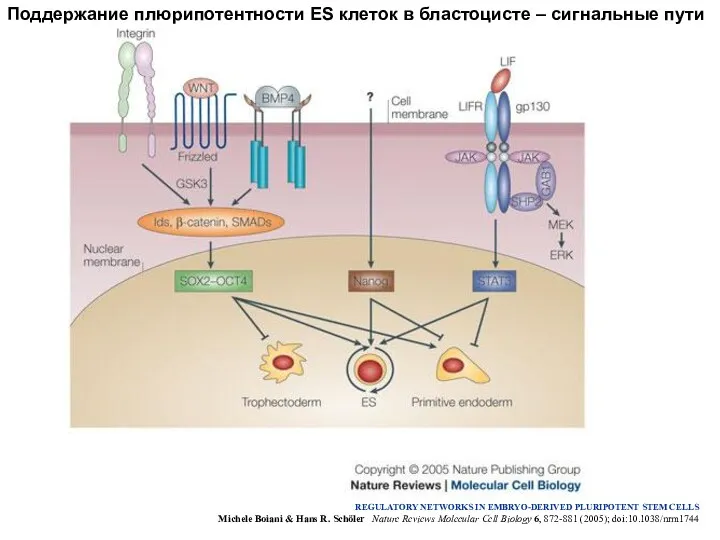
Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути
REGULATORY NETWORKS IN
Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути
REGULATORY NETWORKS IN
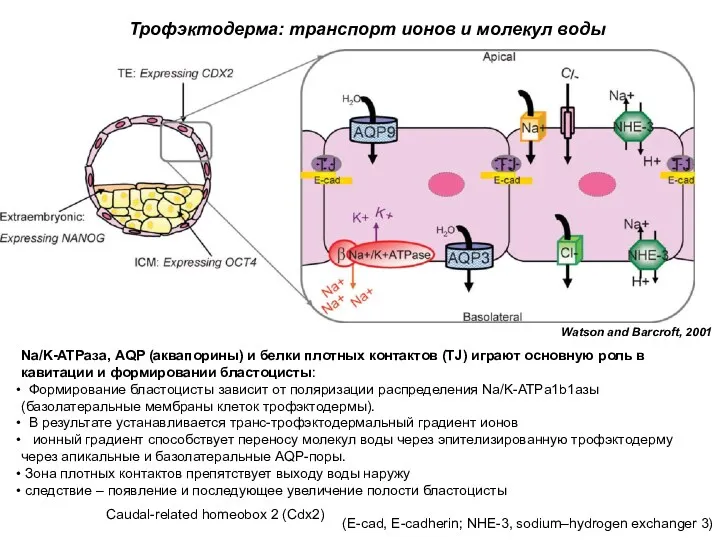
Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль
Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль

Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
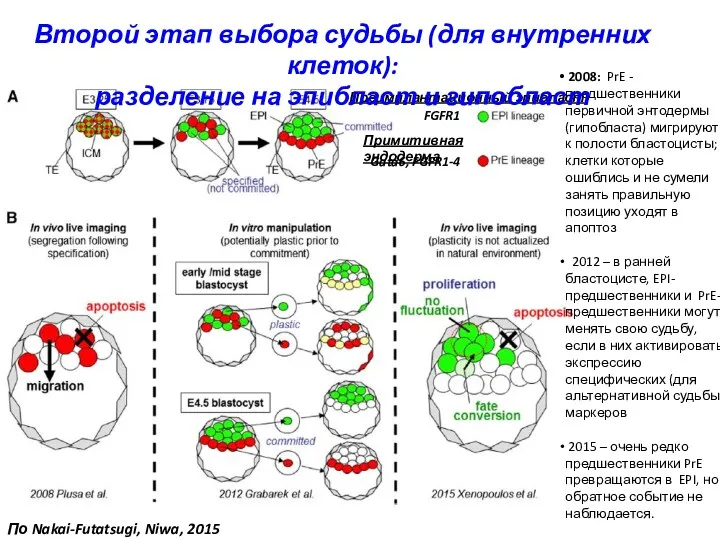
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Примитивная эндодерма
2008: PrE - предшественники первичной энтодермы
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Примитивная эндодерма
2008: PrE - предшественники первичной энтодермы
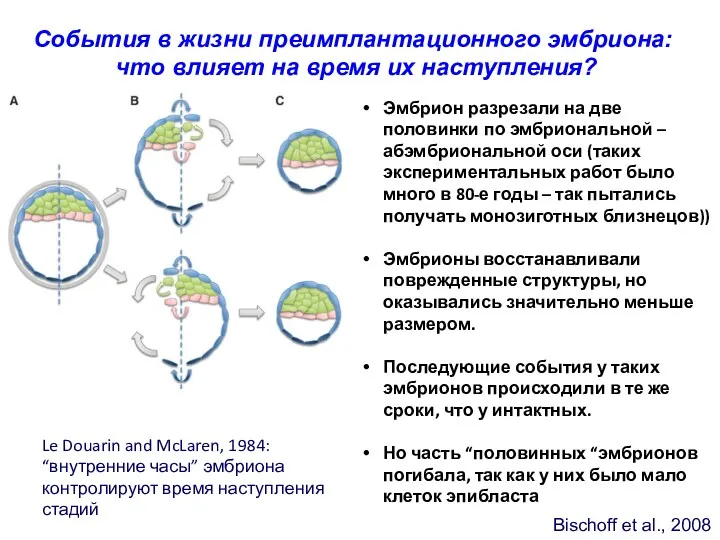
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
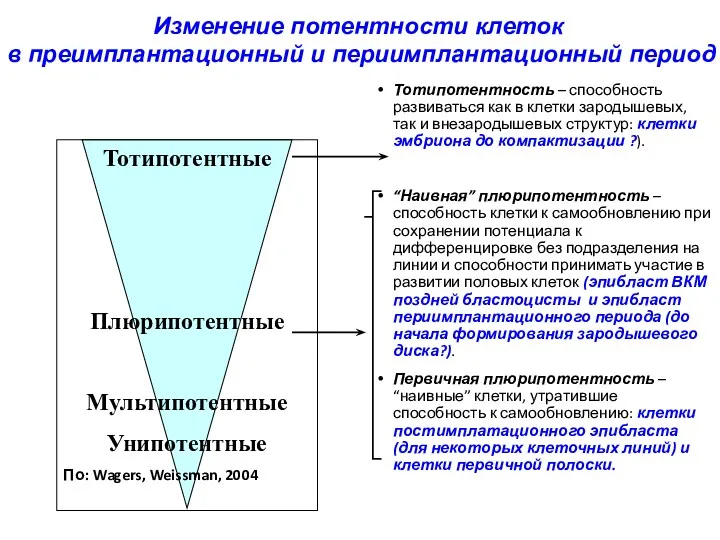
Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых
Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых
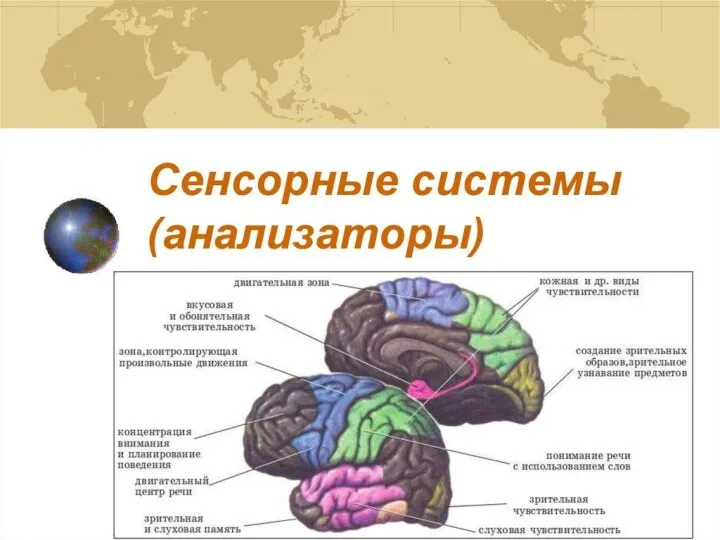 Сенсорные системы (анализаторы)
Сенсорные системы (анализаторы) Поживна цінність, якість та особливості використання в годівлі тварин зернових кормів. Лекція 13
Поживна цінність, якість та особливості використання в годівлі тварин зернових кормів. Лекція 13 Популяції живих організмів та їх основні характеристики
Популяції живих організмів та їх основні характеристики Уход и содержание хомячков
Уход и содержание хомячков Побег и почки
Побег и почки Информация и ее роль в обществе
Информация и ее роль в обществе Признаки наступления на реке
Признаки наступления на реке Природа богата и разнообразна
Природа богата и разнообразна Жасуша теориясы
Жасуша теориясы КВН. Прокариоты
КВН. Прокариоты Червона книга України
Червона книга України Биохимическая трансформация веществ
Биохимическая трансформация веществ Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли
Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли Биология развития
Биология развития Общая характеристика сенсорных систем
Общая характеристика сенсорных систем Внеклассное мероприятие: Птицы нашей Родины в форме КВН
Внеклассное мероприятие: Птицы нашей Родины в форме КВН среда обитания 9 класс
среда обитания 9 класс Эндокринные железы. Общая характеристика
Эндокринные железы. Общая характеристика Генетика пола. Наследование признаков, сцепленных с полом
Генетика пола. Наследование признаков, сцепленных с полом Эволюционное учение. Микро- и макроэволюция. ССТЭ. Генетические процессы в популяциях человека
Эволюционное учение. Микро- и макроэволюция. ССТЭ. Генетические процессы в популяциях человека Омыртқалы жануарлардың қозғалуы
Омыртқалы жануарлардың қозғалуы Полушария мозга
Полушария мозга Физиология движений. Рефлекторный уровень организации движений
Физиология движений. Рефлекторный уровень организации движений Структура ОГЭ по биологии
Структура ОГЭ по биологии Кровеносная система. Кровь
Кровеносная система. Кровь Наследственная изменчивость. 9-10 класс
Наследственная изменчивость. 9-10 класс Влияние стимуляторов роста на проращивание семян томатов
Влияние стимуляторов роста на проращивание семян томатов Теория систем. Функциональная система
Теория систем. Функциональная система