- Регуляция дыхания

Содержание
- 3. РЕГУЛЯЦИЯ ДЫХАНИЯ Лекция № 16 Тема: Лечебный факультет 2008 / 2009 учебный год 13 января 2009
- 4. Литература основная Физиология человека Под редакцией В.М.Покровского, Г.Ф.Коротько Медицина, 2003 (2007) г. С. 366–378.
- 5. Дополнительно Физиология человека Под редакцией В.М.Покровского, Г.Ф.Коротько Медицина, 2003 (2007) г. С. 21–25.
- 6. «Ежик шел-шел, забыл как дышать, Сел под дерево и умер» Из английского юмора
- 7. Вопрос 1
- 8. Под регуляцией внешнего дыхания понимают: Под регуляцией внешнего дыхания понимают: генерацию базового дыхательного паттерна внешнего дыхания.
- 9. Говоря о регуляции в рамках физиологической системы, следует выяснить: что регулируется (какие параметры) как регулируется
- 10. Основные регулируемые параметры в системе дыхания Парциальное давление кислорода (pO2) в артериальной крови. Парциальное давление углекислого
- 11. Как регулируется ? Регуляция осуществляется через изменение паттерна дыхания (частоты и глубины), ведущее к оптимизации вентиляции
- 12. Основные виды (кибернетические) регуляции дыхания по отклонению (отрицательная обратная связь с хеморецепторов при отклонении pO2, pCO2,
- 13. Вопрос 2
- 14. Общий план строения (структура) системы управления Управляющая часть Управляемая часть Каналы связи
- 15. Или по Дж.Уэсту (J.West, 1988; с изменениями): Центральный регулятор Эффекторы Рецепторы
- 16. Центральный регулятор дыхания (управляющую часть системы) Включает: дыхательные центры ствола головного мозга, гипоталамус, лимбическую систему, кору
- 17. Эффекторы (управляемую часть системы) Составляют: дыхательные мышцы
- 18. Рецепторы системы дыхания Хеморецепторы (центральные и периферические) Механорецепторы (легких, дыхательных мышц)
- 19. Общую схему регуляции дыхания можно разбить на детали, например,
- 20. Или так : Схема регуляции дыхания
- 21. Вопрос 3
- 22. ствол головного мозга К стволу мозга (truncus cerebri) относят продолговатый мозг, мост средний мозг. На рисунке
- 23. Дыхательные центры ствола головного мозга Варолиева моста Пневмотаксический (1) Апнейстический (2) Продолговатого мозга вентролатеральный (3) Дорсомедиальный
- 24. Медуллярный дыхательный центр Синонимы: бульбарный дыхательный центр. Расположен в ретикулярной формации в области дна IV желудочка
- 25. Большой вклад в изучение его работы внесли М.Флуранс (1822 г.), Н.А. Миславский (1885 г.), Р.Баумгартен (1956
- 26. Медуллярный дыхательный центр состоит из групп нейронов локализованных в двух основных зонах продолговатого мозга: в дорсомедиальных
- 27. Дыхательные центры Варолиева моста
- 28. Пневмотаксический центр моста (Т.Лумсден, 1960). (греч.: πυέω - дышать, τάξις - расположение в порядок, устроение). Расположен
- 29. Апнейстический центр (греч.: α- - в сложных словах - не- + πυέω - дышать). расположен в
- 30. Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием методом перерезки 1 – пневмотаксический
- 31. Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием методом перерезки
- 32. Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием методом перерезки
- 33. Перерезка на уровне I. Удаление коры головного мозга и мозжечка не оказывает заметного влияния на глубину
- 34. Перерезка на уровне II. Полная поперечная перерезка по нижней границе среднего мозга и по верхней границе
- 35. Результаты описанных перерезок позволяют предполагать, что информация, поступающая по афферентным волокнам блуждающих нервов, может использоваться структурами
- 36. Перерезка на уровне III. Полная поперечная перерезка приблизительно на уровне между верхней третью и нижними двумя
- 37. Апнейстическим дыханием называют редкие затрудненные судорожные дыхательные движения с длительными паузами на высоте вдоха.
- 38. Таким образом, без афферентной информации, поступающей по блуждающим нервам, и без влияния вышележащих структур, группа нейронов,
- 39. Пневмотаксический и апнейстический центры моста, вероятно, являются тоническими пейсмекерами, координирующими функцию ритмических пейсмекеров продолговатого мозга.
- 40. Перерезка на уровне IV. При перерезке ствола мозга между нижним краем моста и продолговатым мозгом можно
- 41. Последующая перерезка блуждающего нерва не оказывает заметного влияния на проявления предшествующей перерезки. Это приводит к предположению,
- 42. Перерезка на уровне V. Перерезка мозга по нижней границе продолговатого мозга приводит к полной остановке дыхания
- 43. Вопрос 4
- 44. Автоматия дыхательного центра ствола головного мозга спобность обеспечить смену вдоха и выдоха за счет своих внутренних
- 45. Генерация дыхательного ритма В течение дыхательного цикла выделяют три фазы активности дыхательных нейронов: инспираторную постинспираторную экспираторную
- 46. Генерация дыхательного ритма
- 47. Дыхательные нейроны
- 48. Дыхательные нейроны ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха (расположены в вентролатеральнои группе);
- 49. Дыхательные нейроны ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха (расположены в вентролатеральнои группе);
- 50. Дыхательные нейроны постинспираторные нейроны — интернейроны, активные в первой половине выдохи, тормозящие как инспираторные, так и
- 51. Инспираторная фаза Соответствует вдоху. Обусловлена последовательной активацией ранних, полных и поздних инспираторных нейронов, что сопровождается линейным
- 52. Постинспираторная фаза Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными нейронами, которые тормозят как
- 53. Экспираторная фаза Соответствует второй половине выдоха (активная экспирация) и обусловлена активацией экспираторных нейронов, иннервирующих мотонейроны мышц
- 54. Вопрос 5
- 55. Гуморальные факторы, участвующие в регуляции дыхания pO2, pCO2 , pH стимулируют легочную вентиляцию гиперкапния (МВЛ увеличивается
- 56. Периферические хеморецепторы: Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их концентрация максимальна в синокаротидной
- 57. КАРОТИДНЫЙ СИНУС [carotid sinus] (Греч.: καρόω - погружать в глубокий сон, лишать чувств, оглушать; лат.: sinus
- 58. Периферические хеморецепторы: хеморецепторные клетки (вторичные рецепторы) контактируют с капиллярами клубочкови друг с другом посредством щелевых контактов
- 59. Афферентная импульсация от них проводится по нерву Циона—Людвига (ветвь X нерва) от аортального тельца и нерву
- 60. реагируют на все три стимула; имеют высокую чувствительность к изменению pO2 артериальной крови (особенно к его
- 61. Периферические хеморецепторы: Хеморецепторы находятся под эфферентным контролем ЦНС; Имеют короткое латентное время действия на дыхательный центр
- 62. Механизм возбуждения
- 63. Механизм возбуждения снижение pO2 и повышение pCO2 и pH приводит к уменьшению внутриклеточного рН, что уменьшает
- 64. Центральные хеморецепторы (хеморецепторная зона дыхательного центра): Локализуются на переднебоковой поверхности продолговатого мозга и моста в виде
- 65. Центральные хеморецепторы (хеморецепторная зона дыхательного центра): Отличаются высокой чувствительностью к изменению рН (пороговые колебания рН примерно
- 66. Сравнительная характеристика раздражителей дыхательной системы
- 68. Вопрос 6
- 69. Гипоталамус Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме. Регулирует дыхание для обеспечения поведенческих
- 70. Лимбическая система Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.
- 71. Кора больших полушарий По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно на спинальные моторные центры
- 72. Синдром "Проклятие Ундины" ("La Malediction d'Ondine") встречающийся при нарушении работы дыхательного центра. При нем человек вообще
- 73. Вопрос 7
- 74. С механорецепторов легких регулируется частота и глубина дыхания: Рецепторы растяжения легких Медленно адаптируются. Расположены в гладких
- 75. Ирритантные рецепторы Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов растяжения легких; Расположены в эпителии
- 76. Юкстаальвеолярные рецепторы (J-рецепторы) Локализуются в интерстиции альвеол у капилляров, реагируют на давление жидкости в межклеточном пространстве
- 77. Проприорецепторы дыхательных мышц Участвуют в компенсации дыхательных нагрузок: Локализуются в дыхательной мускулатуре, преимущественно в межреберных мышцах;
- 78. Вопрос 8 С. 373 – 375.
- 79. Вопрос 9 С. 375 – 376.
- 80. Вопрос 10 С. 376 – 377.
- 81. Вопрос 11 С. 377.
- 83. Скачать презентацию


РЕГУЛЯЦИЯ ДЫХАНИЯ
Лекция № 16
Тема:
Лечебный факультет
2008 / 2009 учебный год
13 января 2009
РЕГУЛЯЦИЯ ДЫХАНИЯ
Лекция № 16
Тема:
Лечебный факультет
2008 / 2009 учебный год
13 января 2009

Литература основная
Физиология человека
Под редакцией
В.М.Покровского,
Г.Ф.Коротько
Медицина, 2003 (2007) г.
С. 366–378.
Литература основная
Физиология человека
Под редакцией
В.М.Покровского,
Г.Ф.Коротько
Медицина, 2003 (2007) г.
С. 366–378.

Дополнительно
Физиология человека
Под редакцией
В.М.Покровского,
Г.Ф.Коротько
Медицина, 2003 (2007) г.
С. 21–25.
Дополнительно
Физиология человека
Под редакцией
В.М.Покровского,
Г.Ф.Коротько
Медицина, 2003 (2007) г.
С. 21–25.

«Ежик шел-шел, забыл как дышать,
Сел под дерево и умер»
Из английского юмора
«Ежик шел-шел, забыл как дышать,
Сел под дерево и умер»
Из английского юмора

Вопрос 1
Вопрос 1

Под регуляцией внешнего дыхания понимают:
Под регуляцией внешнего дыхания понимают:
генерацию базового дыхательного
Под регуляцией внешнего дыхания понимают:
Под регуляцией внешнего дыхания понимают:
генерацию базового дыхательного

Говоря о регуляции в рамках физиологической системы, следует выяснить:
что регулируется
Говоря о регуляции в рамках физиологической системы, следует выяснить:
что регулируется

Основные регулируемые параметры
в системе дыхания
Парциальное давление кислорода (pO2) в
Основные регулируемые параметры
в системе дыхания
Парциальное давление кислорода (pO2) в

Как регулируется ?
Регуляция осуществляется через изменение паттерна дыхания (частоты и
Как регулируется ?
Регуляция осуществляется через изменение паттерна дыхания (частоты и

Основные виды (кибернетические) регуляции дыхания
по отклонению (отрицательная обратная связь с хеморецепторов
Основные виды (кибернетические) регуляции дыхания
по отклонению (отрицательная обратная связь с хеморецепторов

Вопрос 2
Вопрос 2
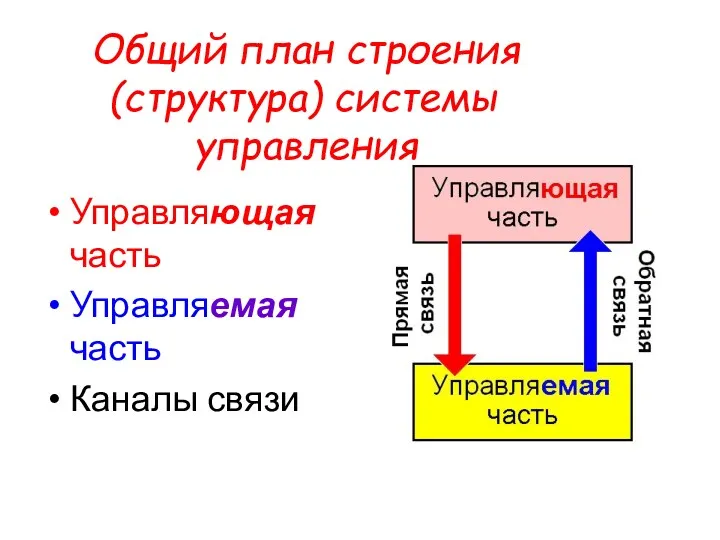
Общий план строения (структура) системы управления
Управляющая часть
Управляемая часть
Каналы связи
Общий план строения (структура) системы управления
Управляющая часть
Управляемая часть
Каналы связи
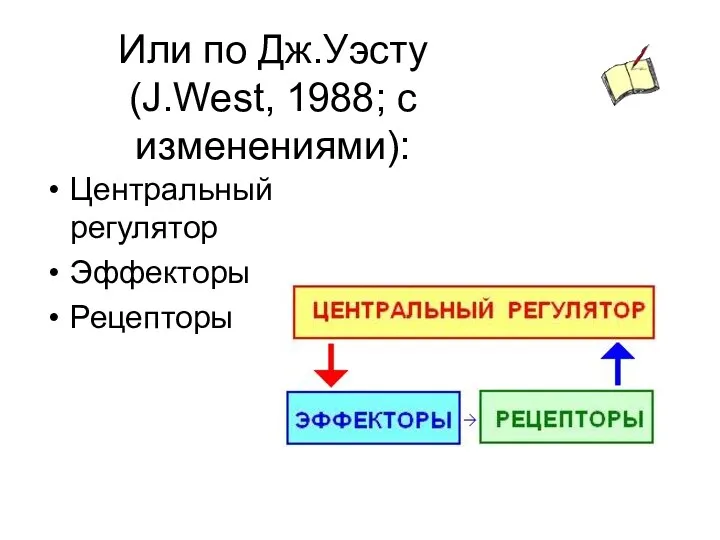
Или по Дж.Уэсту (J.West, 1988; с изменениями):
Центральный регулятор
Эффекторы
Рецепторы
Или по Дж.Уэсту (J.West, 1988; с изменениями):
Центральный регулятор
Эффекторы
Рецепторы

Центральный регулятор дыхания (управляющую часть системы)
Включает:
дыхательные центры ствола головного мозга,
гипоталамус,
лимбическую систему,
кору
Центральный регулятор дыхания (управляющую часть системы)
Включает:
дыхательные центры ствола головного мозга,
гипоталамус,
лимбическую систему,
кору

Эффекторы (управляемую часть системы)
Составляют:
дыхательные мышцы
Эффекторы (управляемую часть системы)
Составляют:
дыхательные мышцы

Рецепторы системы дыхания
Хеморецепторы (центральные и периферические)
Механорецепторы (легких, дыхательных мышц)
Рецепторы системы дыхания
Хеморецепторы (центральные и периферические)
Механорецепторы (легких, дыхательных мышц)
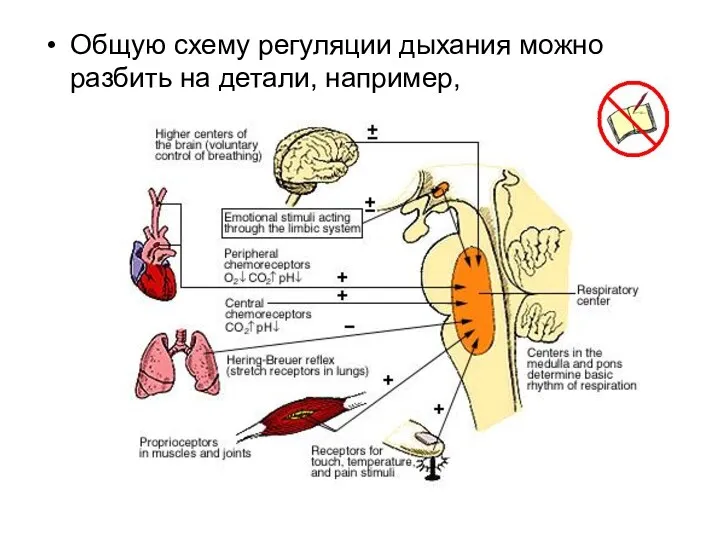
Общую схему регуляции дыхания можно разбить на детали, например,
Общую схему регуляции дыхания можно разбить на детали, например,
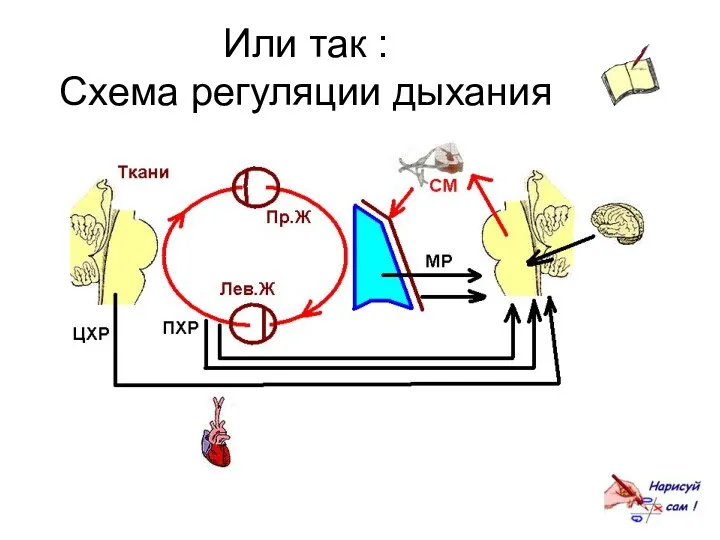
Или так :
Схема регуляции дыхания
Или так :
Схема регуляции дыхания

Вопрос 3
Вопрос 3
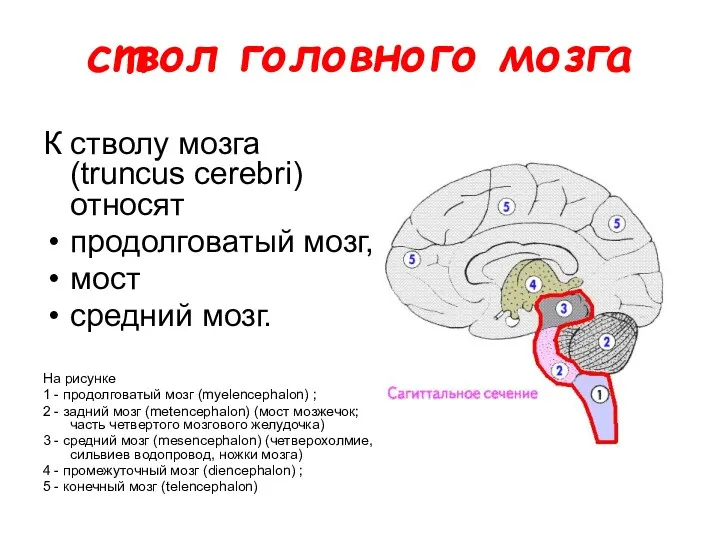
ствол головного мозга
К стволу мозга (truncus cerebri) относят
продолговатый мозг,
мост
ствол головного мозга
К стволу мозга (truncus cerebri) относят
продолговатый мозг,
мост
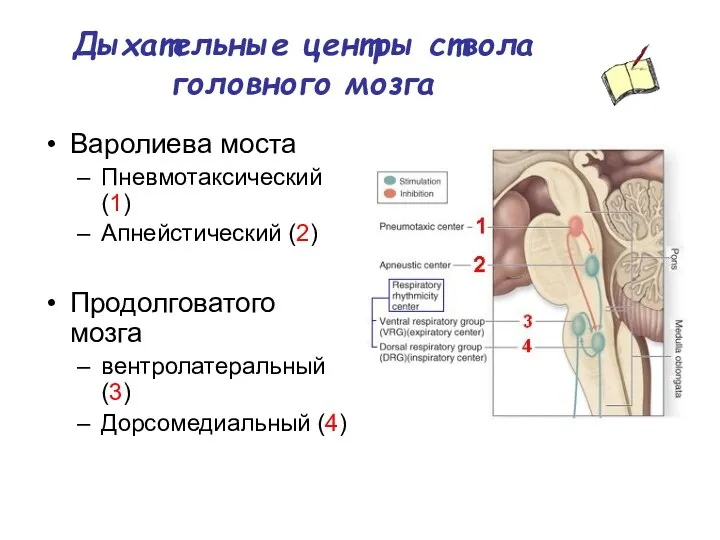
Дыхательные центры ствола головного мозга
Варолиева моста
Пневмотаксический (1)
Апнейстический (2)
Продолговатого мозга
вентролатеральный
Дыхательные центры ствола головного мозга
Варолиева моста
Пневмотаксический (1)
Апнейстический (2)
Продолговатого мозга
вентролатеральный

Медуллярный дыхательный центр
Синонимы: бульбарный дыхательный центр.
Расположен в ретикулярной формации в области
Медуллярный дыхательный центр
Синонимы: бульбарный дыхательный центр.
Расположен в ретикулярной формации в области

Большой вклад в изучение его работы внесли
М.Флуранс (1822 г.),
Н.А.
Большой вклад в изучение его работы внесли
М.Флуранс (1822 г.),
Н.А.

Медуллярный дыхательный центр состоит из групп нейронов локализованных в двух основных
Медуллярный дыхательный центр состоит из групп нейронов локализованных в двух основных
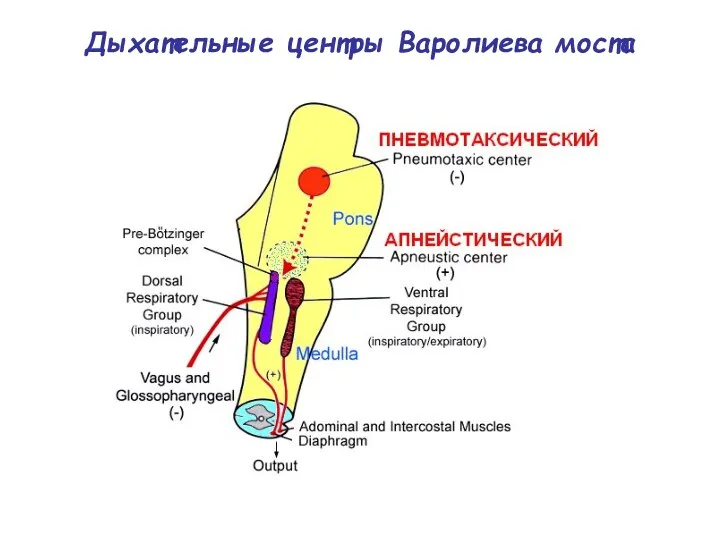
Дыхательные центры Варолиева моста
Дыхательные центры Варолиева моста

Пневмотаксический центр моста (Т.Лумсден, 1960).
(греч.: πυέω - дышать, τάξις - расположение
Пневмотаксический центр моста (Т.Лумсден, 1960).
(греч.: πυέω - дышать, τάξις - расположение

Апнейстический центр
(греч.: α- - в сложных словах - не- + πυέω - дышать).
расположен в
Апнейстический центр
(греч.: α- - в сложных словах - не- + πυέω - дышать).
расположен в
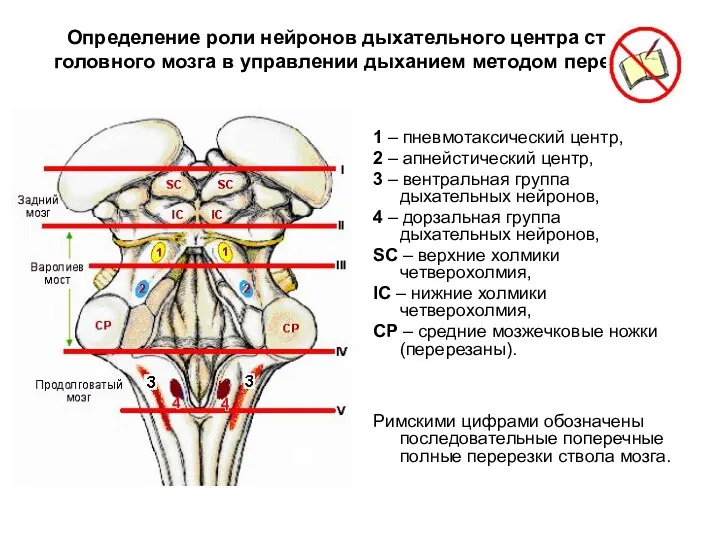
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием
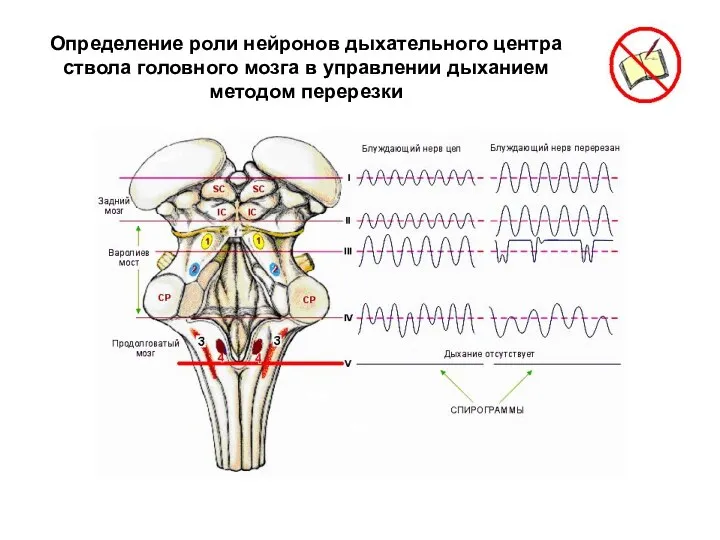
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием
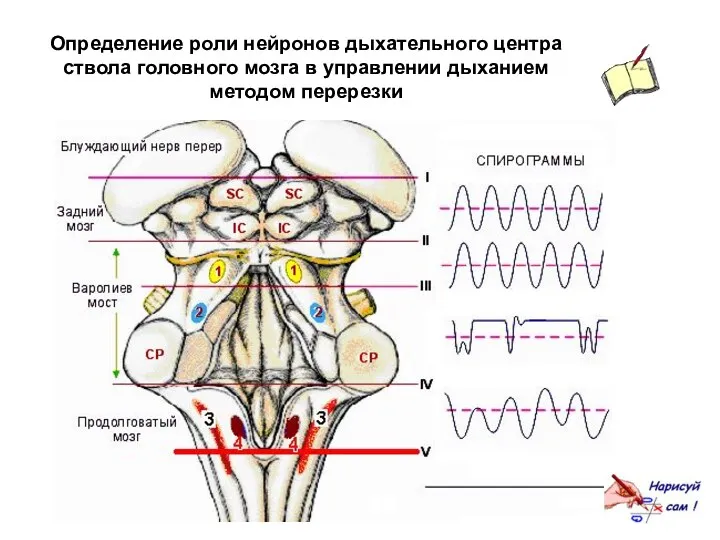
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием
Определение роли нейронов дыхательного центра ствола головного мозга в управлении дыханием

Перерезка на уровне I. Удаление коры головного мозга и мозжечка не
Перерезка на уровне I. Удаление коры головного мозга и мозжечка не

Перерезка на уровне II. Полная поперечная перерезка по нижней границе среднего
Перерезка на уровне II. Полная поперечная перерезка по нижней границе среднего

Результаты описанных перерезок позволяют предполагать, что информация, поступающая по афферентным волокнам
Результаты описанных перерезок позволяют предполагать, что информация, поступающая по афферентным волокнам

Перерезка на уровне III. Полная поперечная перерезка приблизительно на уровне между
Перерезка на уровне III. Полная поперечная перерезка приблизительно на уровне между

Апнейстическим дыханием называют редкие затрудненные судорожные дыхательные движения с длительными паузами
Апнейстическим дыханием называют редкие затрудненные судорожные дыхательные движения с длительными паузами

Таким образом, без афферентной информации, поступающей по блуждающим нервам, и без
Таким образом, без афферентной информации, поступающей по блуждающим нервам, и без

Пневмотаксический и апнейстический центры моста, вероятно, являются тоническими пейсмекерами, координирующими функцию
Пневмотаксический и апнейстический центры моста, вероятно, являются тоническими пейсмекерами, координирующими функцию

Перерезка на уровне IV. При перерезке ствола мозга между нижним краем
Перерезка на уровне IV. При перерезке ствола мозга между нижним краем

Последующая перерезка блуждающего нерва не оказывает заметного влияния на проявления предшествующей
Последующая перерезка блуждающего нерва не оказывает заметного влияния на проявления предшествующей

Перерезка на уровне V. Перерезка мозга по нижней границе продолговатого мозга
Перерезка на уровне V. Перерезка мозга по нижней границе продолговатого мозга

Вопрос 4
Вопрос 4

Автоматия дыхательного центра ствола головного мозга
спобность обеспечить смену вдоха и выдоха
Автоматия дыхательного центра ствола головного мозга
спобность обеспечить смену вдоха и выдоха

Генерация дыхательного ритма
В течение дыхательного цикла выделяют три фазы активности дыхательных
Генерация дыхательного ритма
В течение дыхательного цикла выделяют три фазы активности дыхательных
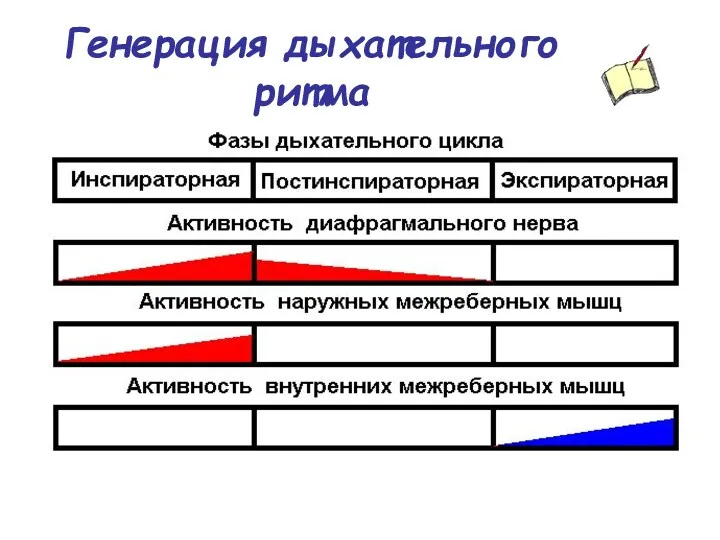
Генерация дыхательного ритма
Генерация дыхательного ритма
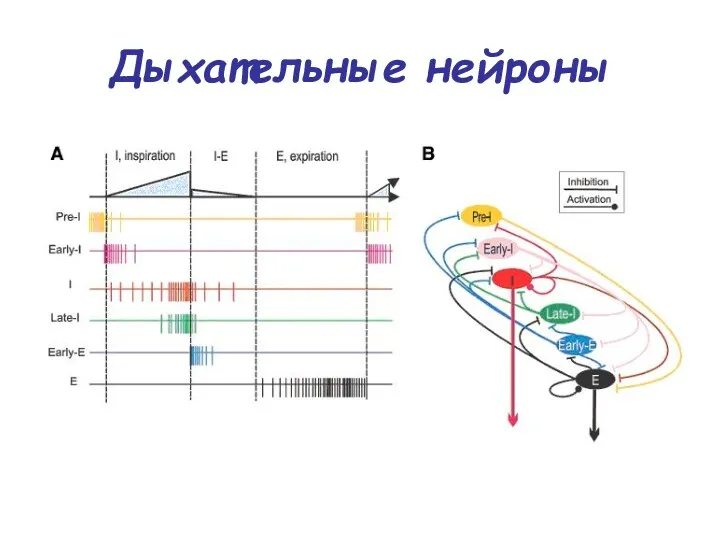
Дыхательные нейроны
Дыхательные нейроны

Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха
Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха

Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха
Дыхательные нейроны
ранние инспираторные нейроны — интернейроны, которые активны в начале вдоха

Дыхательные нейроны
постинспираторные нейроны — интернейроны, активные в первой половине выдохи, тормозящие
Дыхательные нейроны
постинспираторные нейроны — интернейроны, активные в первой половине выдохи, тормозящие

Инспираторная фаза
Соответствует вдоху.
Обусловлена последовательной активацией ранних, полных и поздних инспираторных нейронов,
Инспираторная фаза
Соответствует вдоху.
Обусловлена последовательной активацией ранних, полных и поздних инспираторных нейронов,

Постинспираторная фаза
Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными
Постинспираторная фаза
Соответствует первой половине выдоха (пассивная экспирация) и обусловлена особыми постинспираторными

Экспираторная фаза
Соответствует второй половине
выдоха (активная экспирация) и обусловлена активацией экспираторных нейронов,
Экспираторная фаза
Соответствует второй половине выдоха (активная экспирация) и обусловлена активацией экспираторных нейронов,

Вопрос 5
Вопрос 5

Гуморальные факторы, участвующие в регуляции дыхания
pO2, pCO2 , pH
стимулируют легочную
Гуморальные факторы, участвующие в регуляции дыхания
pO2, pCO2 , pH
стимулируют легочную
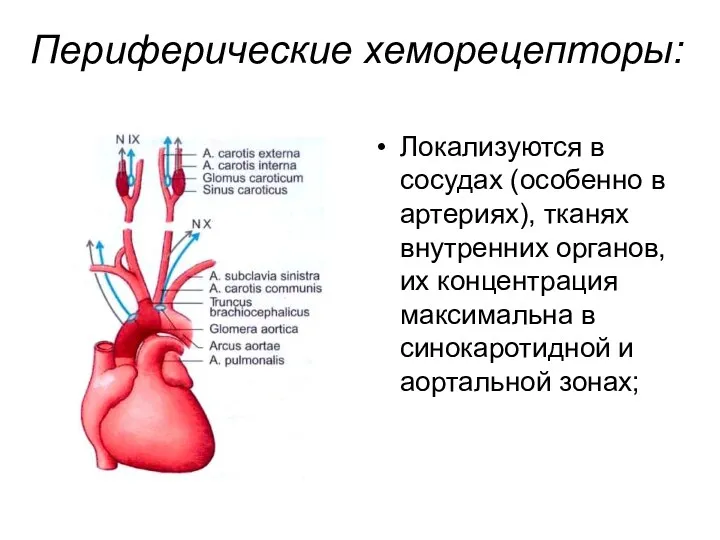
Периферические хеморецепторы:
Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их
Периферические хеморецепторы:
Локализуются в сосудах (особенно в артериях), тканях внутренних органов, их
![КАРОТИДНЫЙ СИНУС [carotid sinus] (Греч.: καρόω - погружать в глубокий](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/373347/slide-56.jpg)
КАРОТИДНЫЙ СИНУС
[carotid sinus]
(Греч.: καρόω - погружать в глубокий сон, лишать
КАРОТИДНЫЙ СИНУС
[carotid sinus]
(Греч.: καρόω - погружать в глубокий сон, лишать
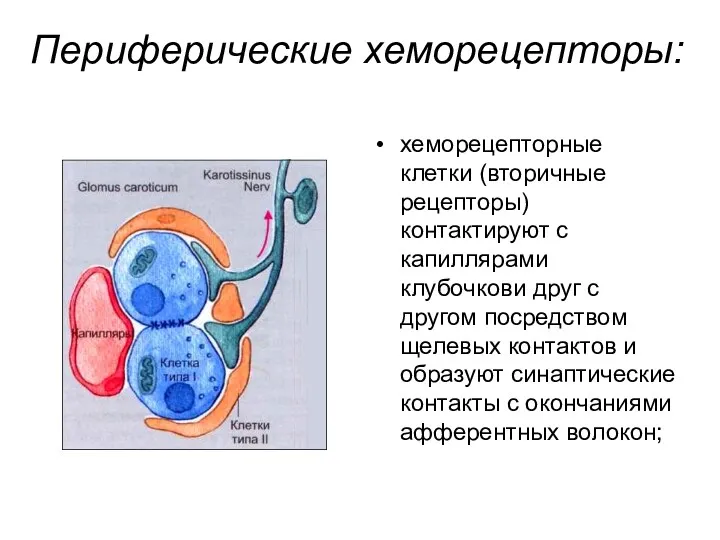
Периферические хеморецепторы:
хеморецепторные клетки (вторичные рецепторы) контактируют с капиллярами клубочкови друг с
Периферические хеморецепторы:
хеморецепторные клетки (вторичные рецепторы) контактируют с капиллярами клубочкови друг с
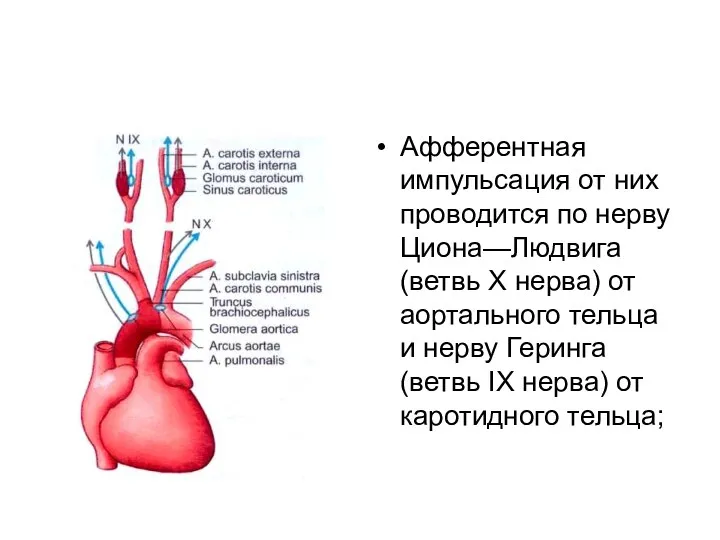
Афферентная импульсация от них проводится по нерву Циона—Людвига (ветвь X нерва)
Афферентная импульсация от них проводится по нерву Циона—Людвига (ветвь X нерва)
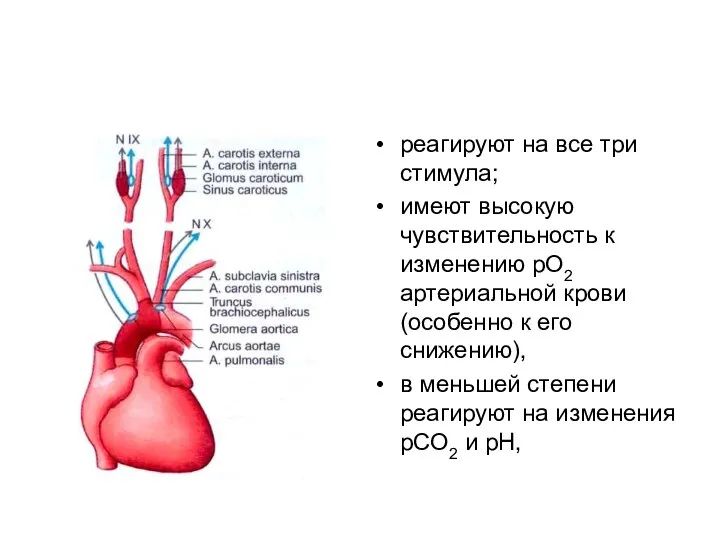
реагируют на все три стимула;
имеют высокую чувствительность к изменению pO2 артериальной
реагируют на все три стимула;
имеют высокую чувствительность к изменению pO2 артериальной

Периферические хеморецепторы:
Хеморецепторы находятся под эфферентным контролем ЦНС;
Имеют короткое латентное время действия
Периферические хеморецепторы:
Хеморецепторы находятся под эфферентным контролем ЦНС;
Имеют короткое латентное время действия
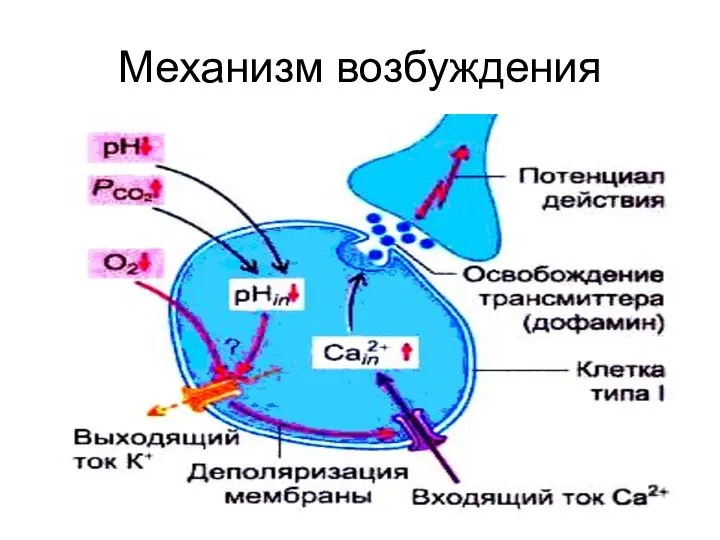
Механизм возбуждения
Механизм возбуждения

Механизм возбуждения
снижение pO2 и повышение pCO2 и pH приводит к уменьшению
Механизм возбуждения
снижение pO2 и повышение pCO2 и pH приводит к уменьшению
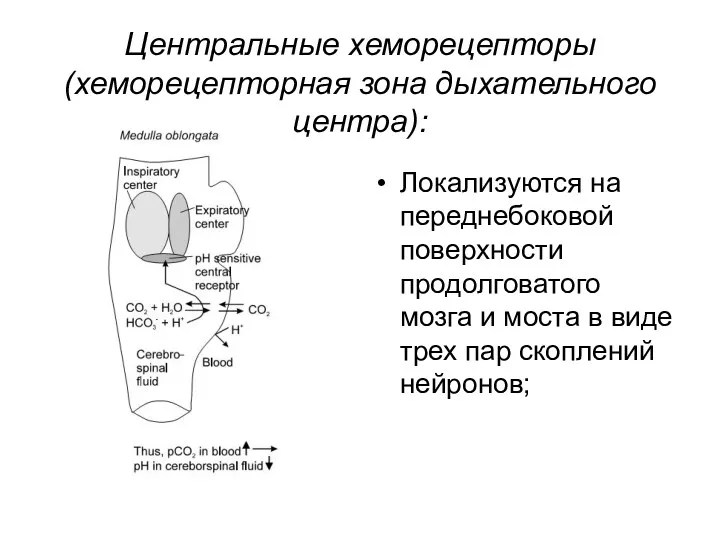
Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Локализуются на переднебоковой поверхности продолговатого мозга
Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Локализуются на переднебоковой поверхности продолговатого мозга

Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Отличаются высокой чувствительностью к изменению рН
Центральные хеморецепторы (хеморецепторная зона дыхательного центра):
Отличаются высокой чувствительностью к изменению рН
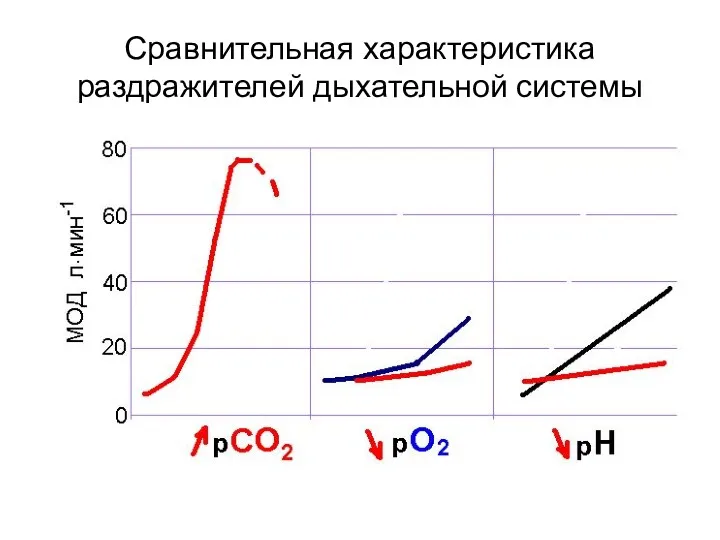
Сравнительная характеристика раздражителей дыхательной системы
Сравнительная характеристика раздражителей дыхательной системы


Вопрос 6
Вопрос 6

Гипоталамус
Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме.
Регулирует дыхание
Гипоталамус
Осуществляет связь дыхания с обменом веществ и терморегуляцией в организме.
Регулирует дыхание

Лимбическая система
Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.
Лимбическая система
Осуществляет связь дыхания с вегетативной регуляцией внутренних органов и эмоциями.

Кора больших полушарий
По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно
Кора больших полушарий
По пирамидным путям, минуя дыхательный центр, оказывает влияние непосредственно

Синдром "Проклятие Ундины" ("La Malediction d'Ondine")
встречающийся при нарушении работы дыхательного центра.
Синдром "Проклятие Ундины" ("La Malediction d'Ondine")
встречающийся при нарушении работы дыхательного центра.

Вопрос 7
Вопрос 7

С механорецепторов легких регулируется частота и глубина дыхания:
Рецепторы растяжения легких
Медленно
С механорецепторов легких регулируется частота и глубина дыхания:
Рецепторы растяжения легких
Медленно

Ирритантные рецепторы
Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов
Ирритантные рецепторы
Быстро адаптирующиеся рецепторы, порог раздражения которых выше, чем рецепторов

Юкстаальвеолярные рецепторы (J-рецепторы)
Локализуются в интерстиции альвеол у капилляров, реагируют на
Юкстаальвеолярные рецепторы (J-рецепторы)
Локализуются в интерстиции альвеол у капилляров, реагируют на

Проприорецепторы дыхательных мышц
Участвуют в компенсации дыхательных нагрузок:
Локализуются в дыхательной мускулатуре, преимущественно
Проприорецепторы дыхательных мышц
Участвуют в компенсации дыхательных нагрузок:
Локализуются в дыхательной мускулатуре, преимущественно

Вопрос 8
С. 373 – 375.
Вопрос 8
С. 373 – 375.

Вопрос 9
С. 375 – 376.
Вопрос 9
С. 375 – 376.

Вопрос 10
С. 376 – 377.
Вопрос 10
С. 376 – 377.

Вопрос 11
С. 377.
Вопрос 11
С. 377.
 Подкласс гамамелидиды
Подкласс гамамелидиды Ріст рослин в умовах невагомості
Ріст рослин в умовах невагомості VITAMINS. History of the discovery of vitamins
VITAMINS. History of the discovery of vitamins Семейство розоцветные
Семейство розоцветные Обмен веществ - метаболизм, общая характеристика
Обмен веществ - метаболизм, общая характеристика Красная книга Саратовской области
Красная книга Саратовской области Состав и строение клеток
Состав и строение клеток Моделирование. Биоритмы
Моделирование. Биоритмы Птичьи посиделки
Птичьи посиделки Проводящие пути ЦНС
Проводящие пути ЦНС Семейства растений
Семейства растений Презентация Вид. Критерии вида
Презентация Вид. Критерии вида Игра Угадай, кто спрятался. Насекомые
Игра Угадай, кто спрятался. Насекомые Условия роста и развития растений
Условия роста и развития растений Растения из Красной книги Челябинской области
Растения из Красной книги Челябинской области Залози змішаної секреції, порушення діяльності та їх профілактика
Залози змішаної секреції, порушення діяльності та їх профілактика Перелетные птицы
Перелетные птицы Факторы, влияющие на митоз
Факторы, влияющие на митоз Метаморфозы корней
Метаморфозы корней Отдел покрытосеменные, или цветковые
Отдел покрытосеменные, или цветковые Пагін. Його будова
Пагін. Його будова Отряд млекопитающие
Отряд млекопитающие Назвать изображенных животных и определить к какому отряду млекопитающих они принадлежат
Назвать изображенных животных и определить к какому отряду млекопитающих они принадлежат План-конспект урока по биологии
План-конспект урока по биологии Метаболизм глицерофосфолипидов. Метаболизм холестерола. (Лекция 10)
Метаболизм глицерофосфолипидов. Метаболизм холестерола. (Лекция 10) Строение и функции клеточной мембраны
Строение и функции клеточной мембраны Биология как наука
Биология как наука Постэмбриональный период развития организмов
Постэмбриональный период развития организмов