- Регуляция обмена веществ

Содержание
- 2. А бывает и еще хуже: только что человек соберется съездить в Кисловодск, – тут иностранец прищурился
- 3. Основные задачи регуляции метаболизма и клеточных функций: внутриклеточное согласование метаболических процессов; межклеточное согласование обмена веществ в
- 4. Внутренняя управляющие сигналы образуются и действуют внутри одной и той же клетки Внешняя управляющие сигналы поступают
- 5. Пример внутренней регуляции метаболизма АМФ фосфорилаза АТФ (аллостерический аллостерический активатор) ингибитор гликогенолиз
- 6. Сигнальные молекулы - эндогенные химические соединения, которые, в результате взаимодействия с рецепторами, обеспечивают внешнее управление метаболическими
- 7. Характерные особенности сигнальных молекул: Малый период жизни Высокая биологическая активность Уникальность действия Наличие эффекта усиления Один
- 8. Клетку, имеющую специализированный воспринимающий рецептор для данной сигнальной молекулы называют клеткой-мишенью. Сигнальная молекула, способная взаимодействовать с
- 9. Способы управления биохимическими процессами управление экспрессией генов (управление количеством определенных белков, ферментов); управление активностью ранее синтезированных
- 10. Основные виды регуляторных эффектов сигнальных молекул (СМ): Эндокринный – СМ поступает с током крови из желез
- 11. Рабочая классификация сигнальных молекул По химической природе Органические (производные аминокислот, белково-пептидной природы, стероиды) Неорганические (оксид азота
- 12. Общие этапы действия сигнальных молекул: I. Распознавание сигнала рецептором клетки-мишени II. Передача сигнала (трансдукция) и его
- 13. Особенности механизма действия липофильных сигнальных молекул: взаимодействие с внутриклеточным рецептором; регуляторный эффект вызван изменением экспрессии генов
- 14. Факторы, необходимые для действия липофильных СМ: Сами СМ Воспринимающий внутриклеточный рецептор, связанный с шапероном. Энхансер или
- 15. Этапы действия липофильных СМ: Проникновение СМ внутрь клетки. Связывание СМ с внутриклеточным рецептором. Освобождение шаперона (запуск
- 16. шаперон протеолиз изменения конформации белка прекращение эффекта Механизм действия липофильных органических сигнальных молекул
- 17. Особенности механизма действия липофобных сигнальных молекул взаимодействие с поверхностным рецептором клетки; сигнал передается от рецептора внутрь
- 18. цАМФ-зависимый механизм действия. Необходимые факторы: Водорастворимая СМ Поверхностный рецептор клетки-мишени Внутриклеточный трансдуктор – G-белок, имеет α-,
- 19. Этапы стимулирующего цАМФ-зависимого механизма действия Взаимодействие СМ с мембранным рецептором Изменение конформации Gs -белка Замена ГДФ
- 21. Ингибирующий цАМФ-зависимый механизм действия СМ Взаимодействие СМ с мембранным рецептором Изменение конформации Gi-белка Замена ГДФ на
- 22. цАМФ-зависимые механизмы действия СМ: стимулирующий ингибирующий
- 23. цГМФ-зависимый механизм действия Рецептор Гуанилатциклаза (образует цГМФ) Протеинкиназа G (активируется цГМФ) Фосфодиэстераза (разрушает цГМФ) Фосфатаза (дефосфорилирует
- 25. Тирозинкиназный механизм действия Димеризация рецепторов после взаимодействия с сигнальной молекулой; Аутофосфорилирование остатков тирозина внутриклеточного домена рецептора;
- 26. Активация внутриклеточных сигнальных путей EGF EGF Фосфорилирование остатков тирозина Рецептор с тирозинкиназной активностью P P P
- 27. Ras-путь трансдукции сигнала
- 28. цГМФ ГТФ -NO Активация протеинкиназы, клеточный ответ ГЦ цит NO цГМФ-зависимый механизм NO
- 29. ГОРМОНЫ - это сигнальные молекулы, которые синтезируются в клетках эндокринных желез, выделяются во внутреннюю среду организма
- 30. Классификация гормонов По химическому строению пептидные (белковые), производные аминокислот, стероидные По локализации синтеза Гормоны центральных эндокринных
- 31. ГОРМОНЫ ГИПОТАЛАМУСА ГИПОТАЛАМУС является компонентом и своеобразным «выходным каналом» лимбической системы. Это отдел промежуточного мозга, контролирующий
- 33. ГОРМОНЫ ГИПОФИЗА В гипофизе выделяют переднюю (аденогипофиз) и заднюю доли (нейрогипофиз). Гормоны аденогипофиза можно разделить на
- 34. Простые белки Гормон роста (соматотропный гормон, ГР, СТГ) является полипептидом, состоящим из 191 аминокислоты. Секреция гормона
- 35. Метаболические эффекты гормона роста: Влияние на белковый обмен: увеличение транспорта аминокислот в клетки-мишени; стимуляция синтеза белка;
- 36. Влияние гормона роста на рост скелета и мягких тканей осуществляется через инсулиноподобные факторы роста (ИФР), которые
- 37. Гипосекреция гормона роста в детском возрасте проявляется гипофизарным нанизмом (карликовость) Гиперсекреция у детей проявляется гигантизмом, у
- 38. Пролактин Синтезируется лактотрофными клетками аденогипофиза, состоит из 199 аминокислотных остатков. Основная физиологическая функция пролактина - стимуляция
- 39. ГЛИКОПРОТЕИНЫ тиреотропный гормон (ТТГ), фолликулостимулирующий гормон (ФСГ), лютеинизирующий гормон (ЛГ)
- 40. Тиреотропный гормон (ТТГ) Клетки-мишени - тиреоциты. Функции: - повышает синтез трийодтиронина и тироксина; - повышает гидролиз
- 41. Фолликулостимулирующий гормон (ФСГ): В женском организме клетки-мишени - фолликулярные клетки, В мужском организме клетки-мишени - клетки
- 42. Лютеинизирующий гормон (ЛГ) В женском организме клетки-мишени - клетки желтых тел, В мужском организме клетки-мишени -
- 43. ПЕПТИДЫ СЕМЕЙСТВА ПРООПИОМЕЛАНОКОРТИНА Проопиомеланокортин – это белок, состоящий из 265 аминокислотных остатков. После отщепления сигнального пептида
- 44. Проопиомеланокортин
- 45. Адренокортикотропный гормон – полипептид, состоящий из 39 аминокислотных остатков. Мишенью для него является кора надпочечниковю Он
- 46. β- эндорфины находятся в гипофизе в ацетилированном состоянии и неактивны, но активны в нервной ткани, где
- 47. β-липотропин стимулирует липолиз. Меланоцитстимулирующие гормоны стимулируют меланогенез (образование пигмента - меланина).
- 48. ГОРМОНЫ ЗАДНЕЙ ДОЛИ ГИПОФИЗА Вазопрессин (антидиуретический гормон, АДГ) и окситоцин. Они образуются в гипоталамусе, транспотируются в
- 49. Эти гормоны – нонапептиды со сходной первичной структурой.
- 50. Антидиуретический гормон Клетками-мишенями для АДГ являются клетки гладкой мускулатуры сосудов и клетки почечных канальцев. В результате
- 51. Окситоцин стимулирует сокращение гладкой мускулатуры матки (стимуляция родовой деятельности) стимулирует сокращение миоэпителиальных клеток молочных желез (вызывает
- 52. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ В щитовидной железе синтезируются гормоны, которые являются йодированными производными тирозина (йодтиронины). К ним
- 54. Биосинтез йодтиронинов Синтез йодтиронинов происходит в составе белка – тиреоглобулина, который находится в фолликулах щитовидной железы.
- 55. Йод поступает в организм с пищей и водой. Суточная потребность в йоде составляет 150- 200 мкг.
- 56. Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина также под влиянием тиреопероксидазы. Происходит образование монойодтирозинов
- 57. Эти гормоны поступают в кровь и связываются с тироксинсвязывающим глобулином, который является основным транспортным белком для
- 58. Влияние йодтиронинов на метаболические процессы Обмен белков В физиологической концентрации йодтиронины усиливают биосинтез белков, но в
- 59. Нарушения секреции тиреоидных гормонов Гипотиреоз развивается вследствие дефицита йодтиронинов при недостаточности функции щитовидной железы (хронический аутоиммунный
- 60. Эндемический зоб (нетоксический зоб) часто встречается у людей, живущих в районах, где содержание йода в воде
- 61. Гипертиреоз возникает при повышенной продукции йодтиронинов вследствие развития опухоли, тиреоидите, избыточном поступлении йода и йодсодержащих препаратов,
- 62. ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ Паратгормон синтезируется в паращитовидных железах и состоит из 84 аминокислотных остатков. Гормон хранится
- 63. При связывании ПТГ с рецепторами клеток костной ткани их активность увеличивается: ускоряется образование щелочной фосфатазы, коллагеназы,
- 64. Гиперсекреция ПТГ Избыточная секреция ПТГ приводит к повышенному выходу кальция и фосфатов в кровь, усилению реабсорбции
- 65. Кальцитонин – полипептид, состоящий из 32 аминокислот. Образуется в С-клетках паращитовидных желез и К-клетках щитовидной железы.
- 66. ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ смешанная железа Эндокринная функция - островки ЛАНГЕРГАНСА 4 гормона - 4 вида клеток:
- 67. Инсулин - это пептид, состоящий из 51 аминокислотного остатка в двух цепях ПРОИНСУЛИН --- 84 АК
- 68. МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА Клетки-мишени: клетки печени, миоциты, жировой ткани Рецепторы находятся на поверхности мембраны клеток
- 69. ВЛИЯНИЕ НА МЕТАБОЛИЗМ: Углеводный обмен: А) снижает уровень ГЛЮКОЗЫ В КРОВИ Б)повышение транспорта глюкозы через мембрану
- 70. 2. ЛИПИДНЫЙ ОБМЕН: ингибирует липолиз - стимулирует липогенез из углеводов
- 71. 3. Обмен белков: - Стимулирует синтез белков -Стимулирует транспорт аминокислот в клетку -Активизирует процессы транскрипции и
- 72. Стимулирует пролиферацию клеток за счёт повышения секреции фактора роста фибробластов (ФРФ), тромбоцитарного фактора роста (ФРТ), фактора
- 73. Дефицит эффектов инсулина проявляется в виде сахарного диабета. Сахарный диабет 1 типа - связан с нарушением
- 74. Признаки дефицита эффектов инсулина : 1. ГИПЕРГЛИКЕМИЯ 2. ПОЛИФАГИЯ 3. ПОЛИДИПСИЯ 4. ГЛЮКОЗУРИЯ ПОЛИУРИЯ ГИПЕРАЗОТЕМИЯ 7.
- 75. ГЛЮКАГОН -полипептид, состоящий из 29 аминокислот. МЕХАНИЗМ ДЕЙСТВИЯ. Клетки-мишени: гепатоциты. Рецепторы-?на поверхности мембраны клеток. Действует через
- 76. ВЛИЯНИЕ НА МЕТАБОЛИЗМ: Повышает концентрацию глюкозы в крови Усиливает процессы глюконеогенеза 3. Повышает интенсивность липолиза
- 77. СОМАТОСТАТИН - пептид, состоящий из 14 аминокислот. Подавляет образование других гормонов ПЖ Функция ПАНКРЕАТИЧЕСКОГО ПОЛИПЕПТИДА Регуляция
- 78. ГОРМОНЫ НАДПОЧЕЧНИКОВ. Мозговой слой А) Адреналин В) Норадреналин образуются из аминокислоты тирозина
- 79. Схема синтеза катехоламинов
- 80. МЕХАНИЗМ ДЕЙСТВИЯ Клетки мишени: клетки печени, скелетных мышц, сердца, слюнных желез, матки. Рецепторы находятся на поверхности
- 81. ВЛИЯНИЕ НА МЕТАБОЛИЗМ 1. Повышает уровень глюкозы в крови. 2. Адреналин действует не только в печени,
- 82. Корковый слой надпочечников более 30 стероидов--- 3 группы: ГЛЮКОКОРТИКОИДЫ МИНЕРАЛОКОРТИКОИДЫ ПОЛОВЫЕ ГОРМОНЫ Все производные - ЦИКЛОПЕНТАНПЕРГИДРОФЕНАНТРЕНА
- 83. МИНЕРАЛОКОРТИКОИДЫ-- альдостерон 1. ГЛЮКОКОРТИКОИДЫ - кортикостерон, кортизол, кортизон В организме человека наибольшую роль играет кортизол
- 85. МЕХАНИЗМ ДЕЙСТВИЯ Рецепторы находятся внутри клетки Механизм действия липофильный Гормон-R?ген---?белок ГЛЮКОКОРТИКОИДЫ: В печени, почках усиливают синтез
- 86. ВЛИЯНИЕ НА МЕТАБОЛИЗМ - повышают глюконеогенез, - усиливают липолиз, - усиливают образование кетоновых тел, - понижают
- 87. 2. Минералокортикоиды: повышают реабсорбцию натрия из первичной мочи - Усиливают секрецию калия в первичную мочу -
- 88. Изменения метаболизма при гипер- и гипофункции коры надпочечников Гиперпродукция глюкокортикоидов повышение уровня АКТГ (болезнь Иценко-Кушинга); избыточный
- 89. Признаки гиперкортицизма : Гипергликемия, снижение толерантности к глюкозе (стимуляция глюконеогенеза) «стероидный диабет»; Усиление катаболизма белков (уменьшение
- 90. Гипофункция коры надпочечников Острая надпочечниковая недостаточность: Нарушения обмена электролитов (потеря ионов натрия, хлора с мочой, потеря
- 91. Первичная надпочечниковая недостаточность (болезнь Аддисона) Туберкулезное или аутоиммунное поражение коры надпочечников; Снижение массы тела; Снижение АД;
- 92. Половые гормоны — гормоны стероидной природы, определяющие у человека и животных половую дифференцировку в эмбриональном периоде,
- 93. Мужские половые гормоны вырабатываются в основном в мужских половых железах — интерстициальных клетках Лейдига семенников (95%),
- 94. Превращение прегненолона в тестостерон катализируется пятью микросомальными ферментами и может протекать двумя путями: через образование дегидроэпиандростерона
- 95. Секреция андрогенов Суточная секреция тестостерона у мужчин составляет в норме примерно 5 мг и сохраняется на
- 96. Транспорт Тестостерон транспортируется в крови в основном в комплексе с альбумином (40%) и специфически связывающим половые
- 97. ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ Синтез. 1. Превращение ХС в прегненолон катализирует в митохондриях клеток теки фолликулов холестеролдесмолаза
- 98. Прогестерон выделяется главным образом жёлтым телом в лютеиновую фазу и фетоплацентарным комплексом во время беременности. В
- 99. Секреция Регуляция секреции 1. Из гипоталамуса импульсно секретируется гонадотропин-рилизинг-гормон (декапептид). 2. Гонадолиберин стимулирует в гипофизе синтез
- 100. В начале менструального цикла ФСГ и ЛГ вызывают развитие первичных фолликулов. Созревающий фолликул секретирует эстрогены, которые
- 101. При наступлении беременности, жёлтое тело продолжает функционировать и секретировать прогестерон, однако на более поздних этапах беременности
- 102. Транспорт. Примерно 95% циркулирующих в крови эстрогенов связано с транспортными белками — СГСБ (секс-гормонсвязывающий белок) и
- 103. Эстрогены через ядерные рецепторы регулируют транскрипцию свыше 50 структурных генов. Эстрогены: стимулируют развитие тканей, участвующих в
- 104. Прогестерон: влияет в основном на репродуктивную функцию организма; увеличивает базальную температуру тела на 0,2-0,5 С, которое
- 106. Скачать презентацию

А бывает и еще хуже: только что человек соберется съездить в
А бывает и еще хуже: только что человек соберется съездить в

Основные задачи регуляции метаболизма и клеточных функций:
внутриклеточное согласование метаболических процессов;
Основные задачи регуляции метаболизма и клеточных функций:
внутриклеточное согласование метаболических процессов;

Внутренняя
управляющие сигналы образуются и действуют внутри одной и той же
Внутренняя
управляющие сигналы образуются и действуют внутри одной и той же
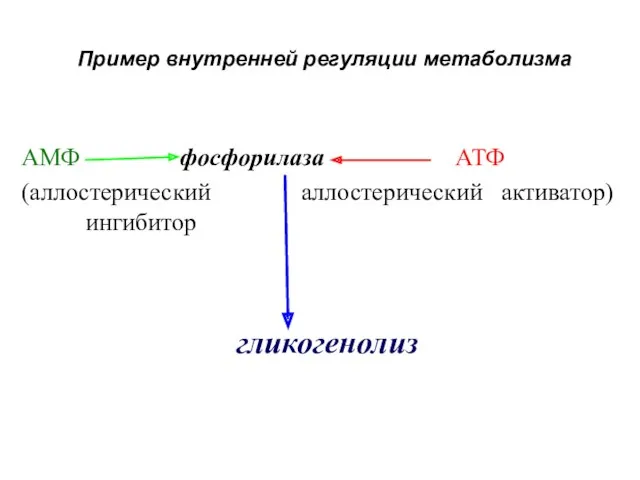
Пример внутренней регуляции метаболизма
АМФ фосфорилаза АТФ
(аллостерический аллостерический активатор) ингибитор
гликогенолиз
Пример внутренней регуляции метаболизма
АМФ фосфорилаза АТФ
(аллостерический аллостерический активатор) ингибитор
гликогенолиз

Сигнальные молекулы - эндогенные химические соединения, которые, в результате взаимодействия с
Сигнальные молекулы - эндогенные химические соединения, которые, в результате взаимодействия с

Характерные особенности сигнальных молекул:
Малый период жизни
Высокая биологическая активность
Уникальность действия
Характерные особенности сигнальных молекул:
Малый период жизни
Высокая биологическая активность
Уникальность действия

Клетку, имеющую специализированный воспринимающий рецептор для данной сигнальной молекулы называют клеткой-мишенью.
Сигнальная
Клетку, имеющую специализированный воспринимающий рецептор для данной сигнальной молекулы называют клеткой-мишенью. Сигнальная

Способы управления биохимическими процессами
управление экспрессией генов (управление количеством определенных белков, ферментов);
управление
Способы управления биохимическими процессами
управление экспрессией генов (управление количеством определенных белков, ферментов);
управление

Основные виды регуляторных эффектов сигнальных молекул (СМ):
Эндокринный – СМ поступает с
Основные виды регуляторных эффектов сигнальных молекул (СМ):
Эндокринный – СМ поступает с

Рабочая классификация сигнальных молекул
По химической природе
Органические (производные аминокислот,
белково-пептидной природы,
Рабочая классификация сигнальных молекул
По химической природе
Органические (производные аминокислот,
белково-пептидной природы,

Общие этапы действия сигнальных молекул:
I. Распознавание сигнала рецептором клетки-мишени
II. Передача
Общие этапы действия сигнальных молекул: I. Распознавание сигнала рецептором клетки-мишени II. Передача

Особенности механизма действия липофильных сигнальных молекул:
взаимодействие с внутриклеточным рецептором;
регуляторный эффект вызван
Особенности механизма действия липофильных сигнальных молекул:
взаимодействие с внутриклеточным рецептором;
регуляторный эффект вызван

Факторы, необходимые для действия липофильных СМ:
Сами СМ
Воспринимающий внутриклеточный рецептор, связанный с
Факторы, необходимые для действия липофильных СМ:
Сами СМ
Воспринимающий внутриклеточный рецептор, связанный с

Этапы действия липофильных СМ:
Проникновение СМ внутрь клетки.
Связывание СМ с внутриклеточным рецептором.
Освобождение
Этапы действия липофильных СМ:
Проникновение СМ внутрь клетки.
Связывание СМ с внутриклеточным рецептором.
Освобождение
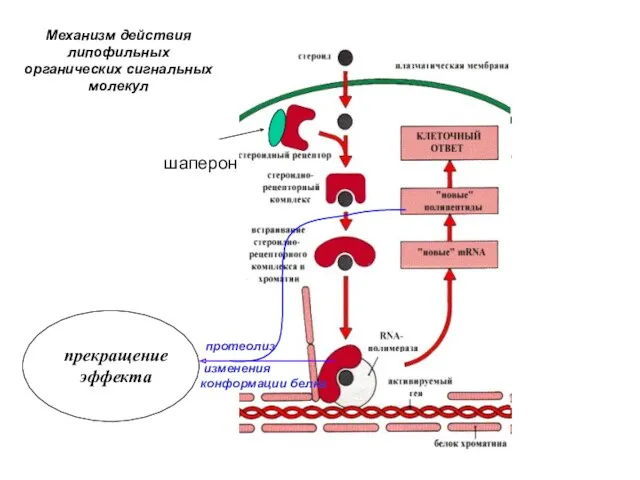
шаперон
протеолиз
изменения конформации белка
прекращение эффекта
Механизм действия липофильных органических сигнальных
шаперон
протеолиз
изменения конформации белка
прекращение эффекта
Механизм действия липофильных органических сигнальных

Особенности механизма действия
липофобных сигнальных молекул
взаимодействие с поверхностным рецептором клетки;
липофобных сигнальных молекул
взаимодействие с поверхностным рецептором клетки;

цАМФ-зависимый механизм действия. Необходимые факторы:
Водорастворимая СМ
Поверхностный рецептор клетки-мишени
Внутриклеточный трансдуктор – G-белок,
цАМФ-зависимый механизм действия. Необходимые факторы:
Водорастворимая СМ
Поверхностный рецептор клетки-мишени
Внутриклеточный трансдуктор – G-белок,

Этапы стимулирующего цАМФ-зависимого механизма действия
Взаимодействие СМ с мембранным рецептором
Изменение конформации Gs
Этапы стимулирующего цАМФ-зависимого механизма действия
Взаимодействие СМ с мембранным рецептором
Изменение конформации Gs
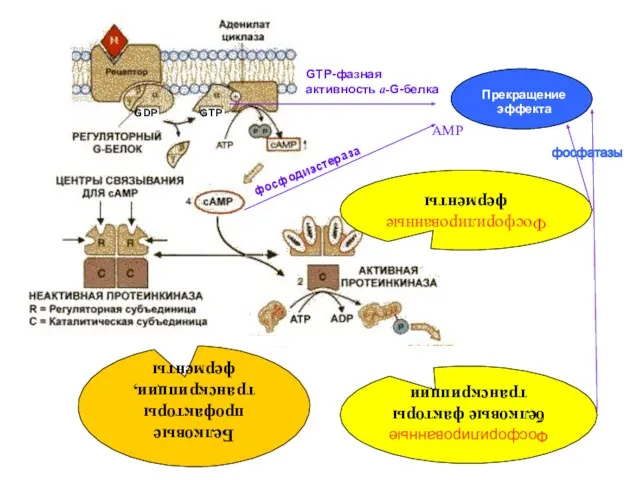

Ингибирующий цАМФ-зависимый механизм действия СМ
Взаимодействие СМ с мембранным рецептором
Изменение конформации Gi-белка
Замена
Ингибирующий цАМФ-зависимый механизм действия СМ
Взаимодействие СМ с мембранным рецептором
Изменение конформации Gi-белка
Замена
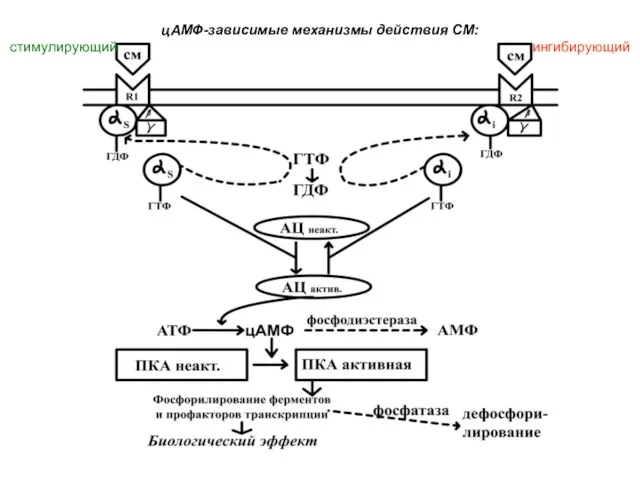
цАМФ-зависимые механизмы действия СМ:
стимулирующий ингибирующий
цАМФ-зависимые механизмы действия СМ:
стимулирующий ингибирующий

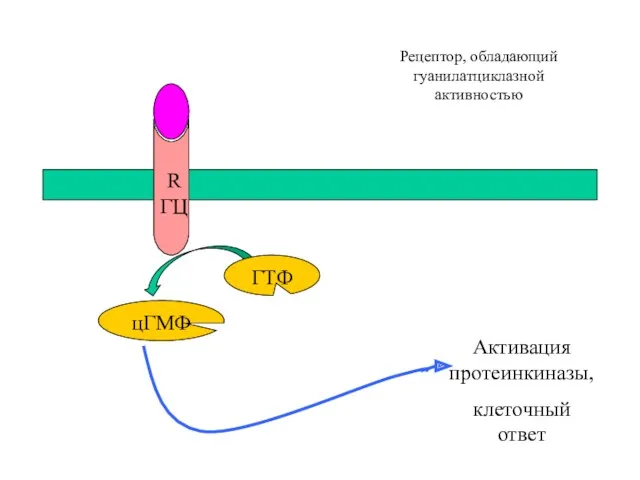
цГМФ-зависимый механизм действия
Рецептор
Гуанилатциклаза (образует цГМФ)
Протеинкиназа G (активируется цГМФ)
Фосфодиэстераза (разрушает цГМФ)
Фосфатаза (дефосфорилирует
цГМФ-зависимый механизм действия
Рецептор
Гуанилатциклаза (образует цГМФ)
Протеинкиназа G (активируется цГМФ)
Фосфодиэстераза (разрушает цГМФ)
Фосфатаза (дефосфорилирует

Тирозинкиназный механизм действия
Димеризация рецепторов после взаимодействия с сигнальной молекулой;
Аутофосфорилирование остатков тирозина
Тирозинкиназный механизм действия
Димеризация рецепторов после взаимодействия с сигнальной молекулой;
Аутофосфорилирование остатков тирозина
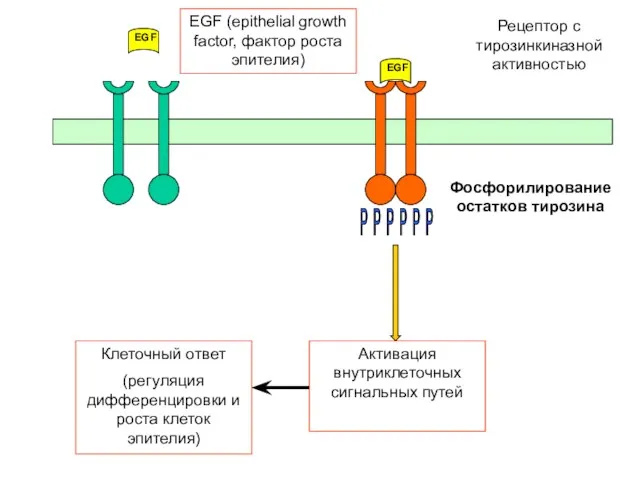
Активация внутриклеточных сигнальных путей
EGF
EGF
Фосфорилирование остатков тирозина
Рецептор с тирозинкиназной активностью
P P P
Активация внутриклеточных сигнальных путей
EGF
EGF
Фосфорилирование остатков тирозина
Рецептор с тирозинкиназной активностью
P P P
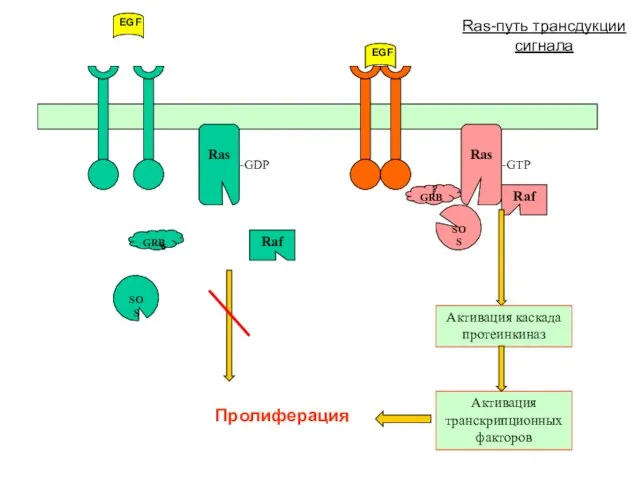
Ras-путь трансдукции сигнала
Ras-путь трансдукции сигнала
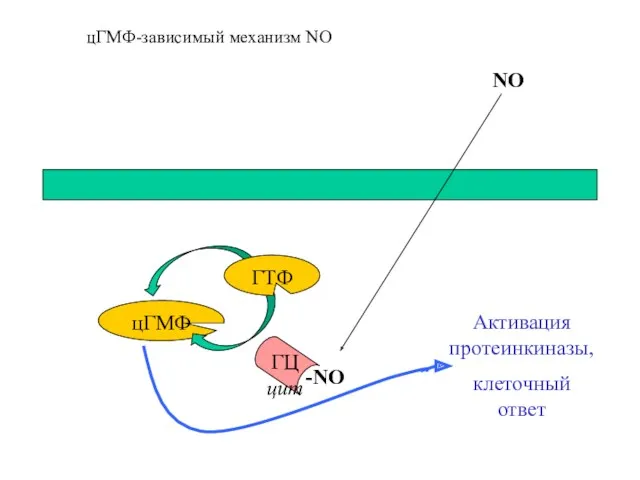
цГМФ
ГТФ
-NO
Активация протеинкиназы,
клеточный ответ
ГЦ цит
NO
цГМФ-зависимый механизм NO
цГМФ
ГТФ
-NO
Активация протеинкиназы,
клеточный ответ
ГЦ цит
NO
цГМФ-зависимый механизм NO

ГОРМОНЫ
- это сигнальные молекулы, которые синтезируются в клетках эндокринных желез, выделяются
ГОРМОНЫ
- это сигнальные молекулы, которые синтезируются в клетках эндокринных желез, выделяются

Классификация гормонов
По химическому строению
пептидные (белковые), производные аминокислот, стероидные
По локализации синтеза
Гормоны центральных
Классификация гормонов
По химическому строению
пептидные (белковые), производные аминокислот, стероидные
По локализации синтеза
Гормоны центральных

ГОРМОНЫ ГИПОТАЛАМУСА
ГИПОТАЛАМУС является компонентом и своеобразным «выходным каналом» лимбической системы.
Это
ГОРМОНЫ ГИПОТАЛАМУСА
ГИПОТАЛАМУС является компонентом и своеобразным «выходным каналом» лимбической системы.
Это


ГОРМОНЫ ГИПОФИЗА
В гипофизе выделяют переднюю (аденогипофиз) и заднюю доли (нейрогипофиз).
Гормоны аденогипофиза
ГОРМОНЫ ГИПОФИЗА
В гипофизе выделяют переднюю (аденогипофиз) и заднюю доли (нейрогипофиз).
Гормоны аденогипофиза

Простые белки
Гормон роста (соматотропный гормон, ГР, СТГ) является полипептидом, состоящим из
Простые белки
Гормон роста (соматотропный гормон, ГР, СТГ) является полипептидом, состоящим из

Метаболические эффекты гормона роста:
Влияние на белковый обмен:
увеличение транспорта аминокислот в клетки-мишени;
стимуляция
Метаболические эффекты гормона роста:
Влияние на белковый обмен:
увеличение транспорта аминокислот в клетки-мишени;
стимуляция

Влияние гормона роста на рост скелета и мягких тканей осуществляется через
Влияние гормона роста на рост скелета и мягких тканей осуществляется через

Гипосекреция гормона роста в детском возрасте проявляется гипофизарным нанизмом (карликовость)
Гиперсекреция у
Гипосекреция гормона роста в детском возрасте проявляется гипофизарным нанизмом (карликовость)
Гиперсекреция у

Пролактин
Синтезируется лактотрофными клетками аденогипофиза, состоит из 199 аминокислотных остатков.
Основная физиологическая
Пролактин
Синтезируется лактотрофными клетками аденогипофиза, состоит из 199 аминокислотных остатков.
Основная физиологическая

ГЛИКОПРОТЕИНЫ
тиреотропный гормон (ТТГ),
фолликулостимулирующий гормон (ФСГ),
лютеинизирующий гормон (ЛГ)
ГЛИКОПРОТЕИНЫ
тиреотропный гормон (ТТГ),
фолликулостимулирующий гормон (ФСГ),
лютеинизирующий гормон (ЛГ)

Тиреотропный гормон (ТТГ)
Клетки-мишени - тиреоциты.
Функции:
- повышает синтез трийодтиронина и тироксина;
- повышает
Тиреотропный гормон (ТТГ)
Клетки-мишени - тиреоциты.
Функции:
- повышает синтез трийодтиронина и тироксина;
- повышает

Фолликулостимулирующий гормон (ФСГ):
В женском организме клетки-мишени - фолликулярные клетки,
В мужском организме
Фолликулостимулирующий гормон (ФСГ):
В женском организме клетки-мишени - фолликулярные клетки,
В мужском организме

Лютеинизирующий гормон (ЛГ)
В женском организме клетки-мишени - клетки желтых тел,
В мужском
Лютеинизирующий гормон (ЛГ)
В женском организме клетки-мишени - клетки желтых тел,
В мужском

ПЕПТИДЫ СЕМЕЙСТВА ПРООПИОМЕЛАНОКОРТИНА
Проопиомеланокортин – это белок, состоящий из 265 аминокислотных остатков.
ПЕПТИДЫ СЕМЕЙСТВА ПРООПИОМЕЛАНОКОРТИНА
Проопиомеланокортин – это белок, состоящий из 265 аминокислотных остатков.
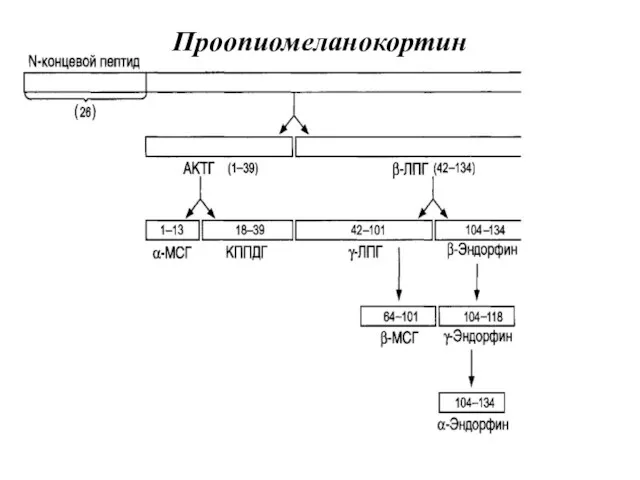
Проопиомеланокортин
Проопиомеланокортин

Адренокортикотропный гормон
– полипептид, состоящий из 39 аминокислотных остатков.
Мишенью для него
Адренокортикотропный гормон
– полипептид, состоящий из 39 аминокислотных остатков.
Мишенью для него

β- эндорфины
находятся в гипофизе в ацетилированном состоянии и неактивны, но активны
β- эндорфины
находятся в гипофизе в ацетилированном состоянии и неактивны, но активны

β-липотропин стимулирует липолиз.
Меланоцитстимулирующие гормоны стимулируют меланогенез (образование пигмента - меланина).
β-липотропин стимулирует липолиз.
Меланоцитстимулирующие гормоны стимулируют меланогенез (образование пигмента - меланина).

ГОРМОНЫ ЗАДНЕЙ ДОЛИ ГИПОФИЗА
Вазопрессин (антидиуретический гормон, АДГ) и окситоцин.
Они образуются
ГОРМОНЫ ЗАДНЕЙ ДОЛИ ГИПОФИЗА
Вазопрессин (антидиуретический гормон, АДГ) и окситоцин.
Они образуются

Эти гормоны – нонапептиды со сходной первичной структурой.
Эти гормоны – нонапептиды со сходной первичной структурой.

Антидиуретический гормон
Клетками-мишенями для АДГ являются клетки гладкой мускулатуры сосудов и клетки
Антидиуретический гормон
Клетками-мишенями для АДГ являются клетки гладкой мускулатуры сосудов и клетки

Окситоцин
стимулирует сокращение гладкой мускулатуры матки (стимуляция родовой деятельности)
стимулирует сокращение миоэпителиальных
Окситоцин
стимулирует сокращение гладкой мускулатуры матки (стимуляция родовой деятельности)
стимулирует сокращение миоэпителиальных

ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
В щитовидной железе синтезируются гормоны, которые являются йодированными производными
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
В щитовидной железе синтезируются гормоны, которые являются йодированными производными
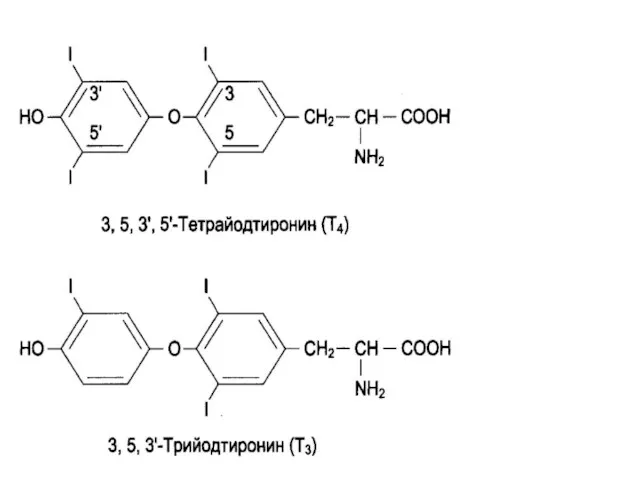

Биосинтез йодтиронинов
Синтез йодтиронинов происходит в составе белка – тиреоглобулина, который находится
Биосинтез йодтиронинов
Синтез йодтиронинов происходит в составе белка – тиреоглобулина, который находится

Йод поступает в организм с пищей и водой. Суточная потребность в
Йод поступает в организм с пищей и водой. Суточная потребность в

Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина также под
Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина также под

Эти гормоны поступают в кровь и связываются с тироксинсвязывающим глобулином,
Эти гормоны поступают в кровь и связываются с тироксинсвязывающим глобулином,

Влияние йодтиронинов на метаболические процессы
Обмен белков
В физиологической концентрации йодтиронины усиливают биосинтез
Влияние йодтиронинов на метаболические процессы
Обмен белков
В физиологической концентрации йодтиронины усиливают биосинтез

Нарушения секреции тиреоидных гормонов
Гипотиреоз развивается вследствие дефицита йодтиронинов при недостаточности функции
Нарушения секреции тиреоидных гормонов
Гипотиреоз развивается вследствие дефицита йодтиронинов при недостаточности функции

Эндемический зоб (нетоксический зоб) часто встречается у людей, живущих в районах,
Эндемический зоб (нетоксический зоб) часто встречается у людей, живущих в районах,

Гипертиреоз возникает при повышенной продукции йодтиронинов вследствие развития опухоли, тиреоидите, избыточном
Гипертиреоз возникает при повышенной продукции йодтиронинов вследствие развития опухоли, тиреоидите, избыточном

ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ
Паратгормон синтезируется в паращитовидных железах и состоит из 84
ГОРМОНЫ ПАРАЩИТОВИДНЫХ ЖЕЛЕЗ
Паратгормон синтезируется в паращитовидных железах и состоит из 84

При связывании ПТГ с рецепторами клеток костной ткани их активность увеличивается:
При связывании ПТГ с рецепторами клеток костной ткани их активность увеличивается:

Гиперсекреция ПТГ
Избыточная секреция ПТГ приводит к повышенному выходу кальция и фосфатов
Гиперсекреция ПТГ
Избыточная секреция ПТГ приводит к повышенному выходу кальция и фосфатов

Кальцитонин – полипептид, состоящий из 32 аминокислот. Образуется в С-клетках паращитовидных
Кальцитонин – полипептид, состоящий из 32 аминокислот. Образуется в С-клетках паращитовидных

ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
смешанная железа
Эндокринная функция - островки ЛАНГЕРГАНСА
4 гормона -
ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
смешанная железа
Эндокринная функция - островки ЛАНГЕРГАНСА
4 гормона -

Инсулин - это пептид,
состоящий из 51 аминокислотного остатка в
Инсулин - это пептид,
состоящий из 51 аминокислотного остатка в

МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА
Клетки-мишени:
клетки печени, миоциты, жировой ткани
Рецепторы находятся на
МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА
Клетки-мишени:
клетки печени, миоциты, жировой ткани
Рецепторы находятся на

ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
Углеводный обмен:
А) снижает уровень ГЛЮКОЗЫ В КРОВИ
Б)повышение
ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
Углеводный обмен:
А) снижает уровень ГЛЮКОЗЫ В КРОВИ
Б)повышение

2. ЛИПИДНЫЙ ОБМЕН:
ингибирует липолиз
- стимулирует липогенез из углеводов
2. ЛИПИДНЫЙ ОБМЕН:
ингибирует липолиз
- стимулирует липогенез из углеводов

3. Обмен белков:
- Стимулирует синтез белков
-Стимулирует транспорт аминокислот
3. Обмен белков:
- Стимулирует синтез белков
-Стимулирует транспорт аминокислот

Стимулирует пролиферацию клеток
за счёт повышения секреции
фактора роста фибробластов (ФРФ),
тромбоцитарного
Стимулирует пролиферацию клеток
за счёт повышения секреции
фактора роста фибробластов (ФРФ),
тромбоцитарного

Дефицит эффектов инсулина проявляется
в виде сахарного диабета.
Сахарный диабет 1 типа
Дефицит эффектов инсулина проявляется
в виде сахарного диабета.
Сахарный диабет 1 типа

Признаки дефицита эффектов инсулина :
1. ГИПЕРГЛИКЕМИЯ
2. ПОЛИФАГИЯ
3. ПОЛИДИПСИЯ
4. ГЛЮКОЗУРИЯ
ПОЛИУРИЯ
Признаки дефицита эффектов инсулина :
1. ГИПЕРГЛИКЕМИЯ
2. ПОЛИФАГИЯ
3. ПОЛИДИПСИЯ
4. ГЛЮКОЗУРИЯ
ПОЛИУРИЯ

ГЛЮКАГОН -полипептид,
состоящий из 29 аминокислот.
МЕХАНИЗМ ДЕЙСТВИЯ.
Клетки-мишени: гепатоциты.
Рецепторы-?на поверхности
мембраны
ГЛЮКАГОН -полипептид,
состоящий из 29 аминокислот.
МЕХАНИЗМ ДЕЙСТВИЯ.
Клетки-мишени: гепатоциты.
Рецепторы-?на поверхности
мембраны

ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
Повышает концентрацию глюкозы
в крови
Усиливает процессы глюконеогенеза
3. Повышает интенсивность
ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
Повышает концентрацию глюкозы
в крови
Усиливает процессы глюконеогенеза
3. Повышает интенсивность

СОМАТОСТАТИН - пептид,
состоящий из 14 аминокислот.
Подавляет образование других
СОМАТОСТАТИН - пептид,
состоящий из 14 аминокислот.
Подавляет образование других

ГОРМОНЫ НАДПОЧЕЧНИКОВ.
Мозговой слой
А) Адреналин
В) Норадреналин
образуются из аминокислоты тирозина
ГОРМОНЫ НАДПОЧЕЧНИКОВ.
Мозговой слой
А) Адреналин
В) Норадреналин
образуются из аминокислоты тирозина
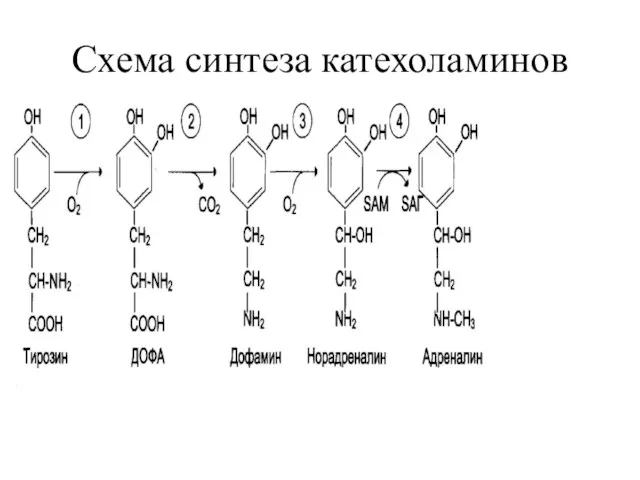
Схема синтеза катехоламинов
Схема синтеза катехоламинов

МЕХАНИЗМ ДЕЙСТВИЯ
Клетки мишени: клетки печени,
скелетных мышц,
сердца, слюнных желез, матки.
Рецепторы
МЕХАНИЗМ ДЕЙСТВИЯ
Клетки мишени: клетки печени,
скелетных мышц,
сердца, слюнных желез, матки.
Рецепторы

ВЛИЯНИЕ НА МЕТАБОЛИЗМ
1. Повышает уровень глюкозы в крови.
2. Адреналин действует не
ВЛИЯНИЕ НА МЕТАБОЛИЗМ
1. Повышает уровень глюкозы в крови.
2. Адреналин действует не

Корковый слой надпочечников
более 30 стероидов---
3 группы: ГЛЮКОКОРТИКОИДЫ
МИНЕРАЛОКОРТИКОИДЫ
ПОЛОВЫЕ
Корковый слой надпочечников
более 30 стероидов---
3 группы: ГЛЮКОКОРТИКОИДЫ
МИНЕРАЛОКОРТИКОИДЫ
ПОЛОВЫЕ

МИНЕРАЛОКОРТИКОИДЫ--
альдостерон
1. ГЛЮКОКОРТИКОИДЫ - кортикостерон,
кортизол, кортизон
В организме человека
МИНЕРАЛОКОРТИКОИДЫ--
альдостерон
1. ГЛЮКОКОРТИКОИДЫ - кортикостерон,
кортизол, кортизон
В организме человека
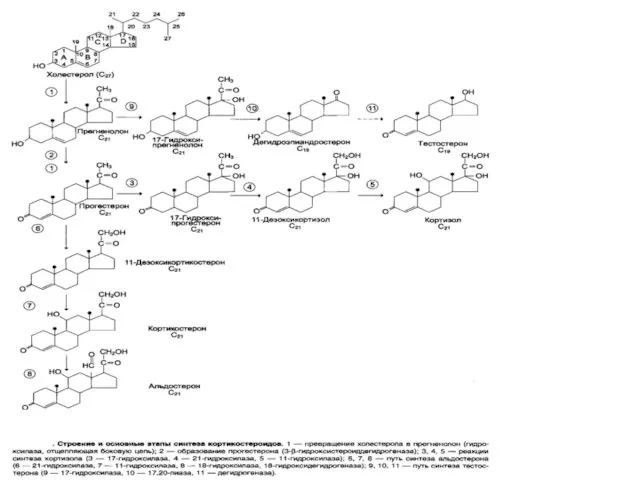

МЕХАНИЗМ ДЕЙСТВИЯ
Рецепторы находятся внутри клетки
Механизм действия липофильный
Гормон-R?ген---?белок
ГЛЮКОКОРТИКОИДЫ:
В
МЕХАНИЗМ ДЕЙСТВИЯ
Рецепторы находятся внутри клетки
Механизм действия липофильный
Гормон-R?ген---?белок
ГЛЮКОКОРТИКОИДЫ:
В

ВЛИЯНИЕ НА МЕТАБОЛИЗМ
- повышают глюконеогенез,
- усиливают липолиз,
- усиливают образование кетоновых
ВЛИЯНИЕ НА МЕТАБОЛИЗМ
- повышают глюконеогенез,
- усиливают липолиз,
- усиливают образование кетоновых

2. Минералокортикоиды:
повышают реабсорбцию натрия из первичной мочи
- Усиливают секрецию калия
2. Минералокортикоиды:
повышают реабсорбцию натрия из первичной мочи
- Усиливают секрецию калия

Изменения метаболизма при гипер- и гипофункции коры надпочечников
Гиперпродукция глюкокортикоидов
повышение уровня АКТГ
Изменения метаболизма при гипер- и гипофункции коры надпочечников
Гиперпродукция глюкокортикоидов
повышение уровня АКТГ

Признаки гиперкортицизма :
Гипергликемия, снижение толерантности к глюкозе (стимуляция глюконеогенеза) «стероидный диабет»;
Усиление
Признаки гиперкортицизма :
Гипергликемия, снижение толерантности к глюкозе (стимуляция глюконеогенеза) «стероидный диабет»;
Усиление

Гипофункция коры надпочечников
Острая надпочечниковая недостаточность:
Нарушения обмена электролитов (потеря ионов натрия, хлора
Гипофункция коры надпочечников
Острая надпочечниковая недостаточность:
Нарушения обмена электролитов (потеря ионов натрия, хлора

Первичная надпочечниковая недостаточность (болезнь Аддисона)
Туберкулезное или аутоиммунное поражение коры надпочечников;
Снижение массы
Первичная надпочечниковая недостаточность (болезнь Аддисона)
Туберкулезное или аутоиммунное поражение коры надпочечников;
Снижение массы

Половые гормоны — гормоны стероидной природы, определяющие у человека и животных
Половые гормоны — гормоны стероидной природы, определяющие у человека и животных

Мужские половые гормоны вырабатываются в основном в мужских половых железах —
Мужские половые гормоны вырабатываются в основном в мужских половых железах —

Превращение прегненолона в тестостерон катализируется пятью микросомальными ферментами и может протекать
Превращение прегненолона в тестостерон катализируется пятью микросомальными ферментами и может протекать

Секреция андрогенов
Суточная секреция тестостерона у мужчин составляет в норме примерно
Секреция андрогенов
Суточная секреция тестостерона у мужчин составляет в норме примерно

Транспорт
Тестостерон транспортируется в крови в основном в комплексе с альбумином (40%)
Транспорт
Тестостерон транспортируется в крови в основном в комплексе с альбумином (40%)

ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Синтез.
1. Превращение ХС в прегненолон катализирует в митохондриях
ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Синтез.
1. Превращение ХС в прегненолон катализирует в митохондриях

Прогестерон выделяется главным образом жёлтым телом в лютеиновую фазу и фетоплацентарным
Прогестерон выделяется главным образом жёлтым телом в лютеиновую фазу и фетоплацентарным

Секреция
Регуляция секреции
1. Из гипоталамуса импульсно секретируется гонадотропин-рилизинг-гормон (декапептид).
2. Гонадолиберин
Секреция
Регуляция секреции
1. Из гипоталамуса импульсно секретируется гонадотропин-рилизинг-гормон (декапептид).
2. Гонадолиберин

В начале менструального цикла ФСГ и ЛГ вызывают развитие первичных фолликулов.
В начале менструального цикла ФСГ и ЛГ вызывают развитие первичных фолликулов.

При наступлении беременности, жёлтое тело продолжает функционировать и секретировать прогестерон, однако
При наступлении беременности, жёлтое тело продолжает функционировать и секретировать прогестерон, однако

Транспорт. Примерно 95% циркулирующих в крови эстрогенов связано с транспортными белками
Транспорт. Примерно 95% циркулирующих в крови эстрогенов связано с транспортными белками

Эстрогены через ядерные рецепторы регулируют транскрипцию свыше 50 структурных генов.
Эстрогены:
стимулируют развитие
Эстрогены через ядерные рецепторы регулируют транскрипцию свыше 50 структурных генов.
Эстрогены:
стимулируют развитие

Прогестерон:
влияет в основном на репродуктивную функцию организма;
увеличивает базальную температуру тела на
Прогестерон:
влияет в основном на репродуктивную функцию организма;
увеличивает базальную температуру тела на
 Эволюция растений
Эволюция растений презентация Самые высокие горы
презентация Самые высокие горы Оценка качества почвы с помощью растения – биоиндикатора
Оценка качества почвы с помощью растения – биоиндикатора Мінеральне та повітряне живлення рослин
Мінеральне та повітряне живлення рослин Биология оқыту үрдісінде тәрбиелеу Ептіліктердің және білімдердің қалыптастыру
Биология оқыту үрдісінде тәрбиелеу Ептіліктердің және білімдердің қалыптастыру Генетические основы селекции.
Генетические основы селекции. Поджелудочная железа
Поджелудочная железа Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия)
Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия) Лекарственные растения
Лекарственные растения Презентация к уроку биологии на тему Витамины.
Презентация к уроку биологии на тему Витамины. Gardis. Хвойные декоративные растения (каталог)
Gardis. Хвойные декоративные растения (каталог) Семейство Rosacea. Розоцветные
Семейство Rosacea. Розоцветные Эволюция нервной, кровеносной и выделительной систем органов
Эволюция нервной, кровеносной и выделительной систем органов Общая характеристика типа Моллюски
Общая характеристика типа Моллюски Биопестициды
Биопестициды В гостях у природы
В гостях у природы Тканевый обмен веществ
Тканевый обмен веществ Морфологія, будова та хімічний склад бактеріальної клітини
Морфологія, будова та хімічний склад бактеріальної клітини Функциональное состояние в структуре поведения
Функциональное состояние в структуре поведения Болезни эмбрионов с/х птицы
Болезни эмбрионов с/х птицы Иван Владимирович Мичурин (1855-1935)
Иван Владимирович Мичурин (1855-1935) Основні закономірності незалежного та зчепленого успадкування
Основні закономірності незалежного та зчепленого успадкування Дигибридное скрещивание. Закон независимого наследования признаков
Дигибридное скрещивание. Закон независимого наследования признаков Презентация по биологии Класс Птицы для 7 класса.
Презентация по биологии Класс Птицы для 7 класса. Свободные жирные кислоты и кальциевый гомеостаз скелетно-мышечных клеток
Свободные жирные кислоты и кальциевый гомеостаз скелетно-мышечных клеток Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2)
Основные понятия о системе движений. Задача биомеханики двигательных действий. (Лекция 2) Бесполое размножение клетки
Бесполое размножение клетки Биотехнологии. Генная инженерия
Биотехнологии. Генная инженерия