- Рецепторы, проявляющие ферментативную активность

Содержание
- 2. СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ N-концевой домен Участки гликозилирования С-концевой домен
- 3. ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ АДРЕНАЛИН, НОРАДРЕНАЛИН АЦЕТИЛХОЛИН СЕРОТОНИН ГИСТАМИН ОПИОИДЫ, КАННАБИНОИДЫ ГОРМОНЫ
- 4. ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ α1АДРЕНОРЕЦЕПТОР β2 АДРЕНОРЕЦЕПТОР
- 5. Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
- 6. Калиевый канал
- 7. СТРУКТУРА G-БЕЛКА
- 8. МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
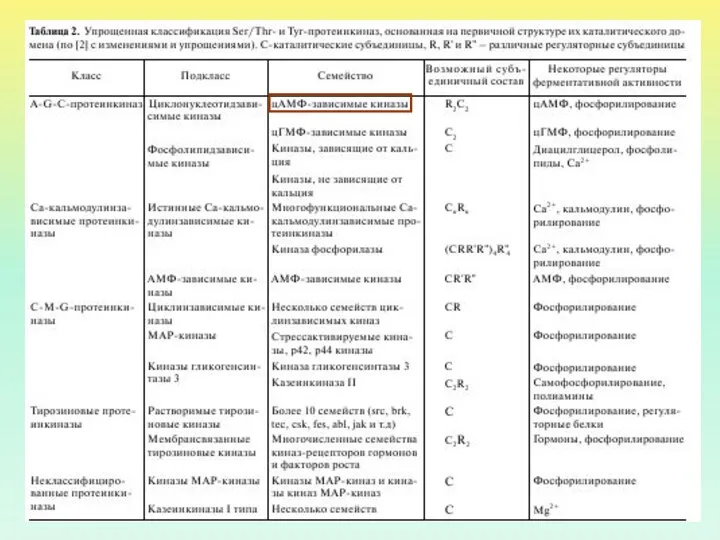
- 10. ВИДЫ G-БЕЛКОВ ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ GS И GI БЕЛКИ ДЕЙСТВУЮТ НА АДЕНИЛАТЦИКЛАЗУ GO ИНГИБИРУЕТ
- 11. ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ стимулирующий ингибирующий Холерный токсин
- 13. Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу. АДФрибозилирование α-субъединицы GS белка приводит к ее
- 14. ВИДЫ G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ
- 15. ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
- 16. цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином. Циклический аденозинмонофосфат (цАМФ) – первое соединение, которое
- 17. Фермент, который образует цАМФ, - аденилатциклаза Фермент, который разрушает цАМФ (переводит в нециклическую форму) – фосфодиэстераза
- 18. ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада этого соединения
- 19. АТФ АМФ цАМФ АЦ ФДЭ РЕСИНТЕЗ АТФ
- 20. МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ β-адренорецепторы α2 адренорецептор
- 21. ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной ткани В ответ на
- 22. ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
- 23. ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ СТРУКТУР цАМФ-зависимые
- 24. МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
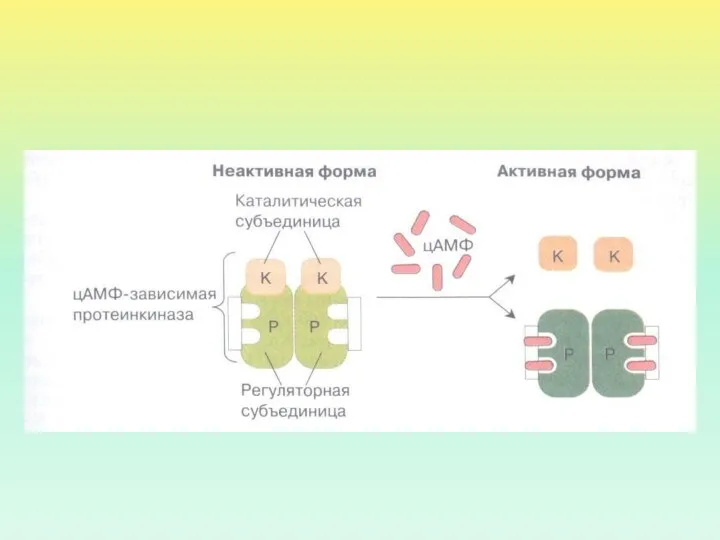
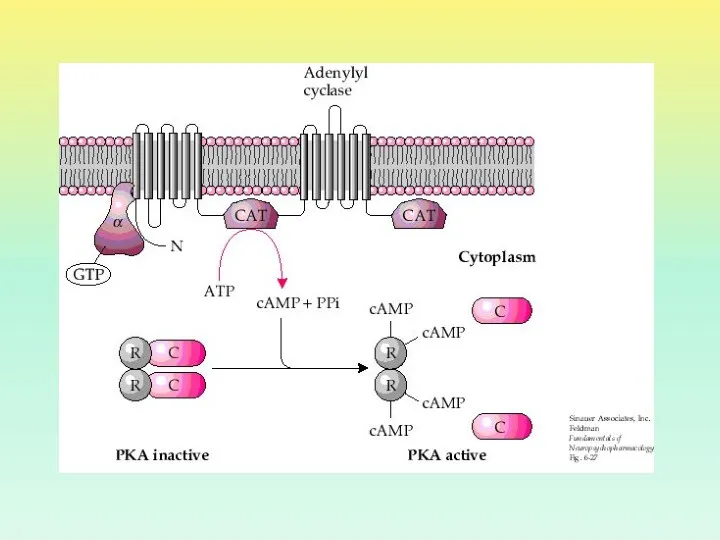
- 26. ПРОТЕИНКИНАЗА А Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и 2 каталитических субъединиц. После
- 29. БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А БЕЛКИ ИОННЫХ КАНАЛОВ ИОНООБМЕННИКИ ТРАНСПОРТНЫЕ АТФазы БЕЛКИ ЦИТОСКЕЛЕТА ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
- 30. цГМФ ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
- 31. ОБРАЗОВАНИЕ И РАСПАД цГМФ ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА ФЕРМЕНТ, КОТОРЫЙ ЕГО РАЗРУШАЕТ - ФОСФОДИЭСТЕРАЗА
- 32. цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток. Повышение концентрации цГМФ приводит к
- 33. ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению насоса. Результат: ослабление реабсорбции ионов
- 34. ПУТИ ОБРАЗОВАНИЯ цГМФ ОПОСРЕДОВАННЫЙ БАВ Например, натрийуретическим пептидом ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
- 35. ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ Мембранная гуанилатциклаза Растворимая форма гуанилатциклазы активируется оксидом азота ПРОТЕИНКИНАЗА G
- 36. МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА NaУП + R → ↑ГЦ → ↑цГМФ → ↑ПК G
- 37. ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ ОТСУТСТВУЮТ G-БЕЛКИ СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ ГУАНИЛАТЦИКЛАЗЫ
- 38. РАСТВОРИМАЯ ФОРМА ГЦ ГЕТЕРОДИМЕР СОДЕРЖИТ SH-группы СОДЕРЖИТ ГЕМ АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА ШИРОКО РАСПРОСТРАНЕНА
- 39. ПРОТЕИНКИНАЗА G Протеинкиназа G в неактивной форме состоит из 2 субъединиц, имеет 2 регуляторных центра, с
- 40. РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
- 41. РЕЦЕПТОРНЫЕ ТИРОЗИНКИНАЗЫ Агонисты: факторы роста инсулин
- 42. Факторы роста (GF) или митогены стимулируют рост клетки и ее прохождение через митоз. GF - полипептиды,
- 43. РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА 1 – внеклеточный N-концевой участок, связывающий агонисты; 2 – трансмембранный участок; 3 – внутриклеточный
- 45. МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ (РТК) ДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ ТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ ФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ БЕЛКОВ-СУБСТРАТОВ, СОДЕРЖАЩИХ
- 46. Ras-БЕЛОК – МОНОМЕРНАЯ ГТФаза В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ С ГТФ, В НЕАКТИВНОМ – С ГДФ
- 47. МЕХАНИЗМ АКТИВАЦИИ Ras-БЕЛКА ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ (КИНАЗНЫЙ КАСКАД) ЭФФЕКТ –
- 49. МЕМБРАННОСВЯЗАННАЯ ГУАНИЛАТЦИКЛАЗА АГОНИСТЫ: Na-УРЕТИЧЕСКИЙ ПЕПТИД ПЕПТИДЫ, СЕКРЕТИРУЕМЫЕ ЯИЧНИКАМИ И СТИМУЛИРУЮЩИЕ ПОДВИЖНОСТЬ СПЕРМАТОЗОИДОВ
- 50. РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ
- 51. Эти рецепторы при активации связываются с цитоплазматическими тирозинкиназами и образуют сигнальный комплекс. Рецепторы цитокинов Рецепторы антигенов
- 52. АГОНИСТЫ – ЦИТОКИНЫ Интерлейкины (IL) Интерфероны (IFNs) Колониестимулирующие факторы (CSFs ) Фактор некроза опухоли (TNF)
- 53. Цитокины — группа гормоноподобных белков и пептидов синтезируются и секретируются клетками иммунной системы и другими типами
- 54. ЦИТОКИНОВЫЙ РЕЦЕПТОР Эти рецепторы не имеют киназного участка в своем составе и фосфорилируются особыми цитоплазматическими киназами
- 55. МЕХАНИЗМ ДЕЙСТВИЯ ЦИТОКИНОВЫХ РЕЦЕПТОРОВ 1 – СВЯЗЫВАНИЕ РЕЦЕПТОРА С ЛИГАНДОМ 2 – СТИМУЛЯЦИЯ ДИМЕРИЗАЦИИ БЕЛКА-ПЕРЕНОСЧИКА СИГНАЛА.
- 56. РЕЦЕПТОРЫ КЛЕТОЧНОЙ АДГЕЗИИ ИНТЕГРИНЫ КАДГЕРИНЫ СЕЛЕКТИНЫ
- 57. ИНТЕГРИНЫ взаимодействуют как с внеклеточными молекулами (коллаген, фибронектин и др.), так и с внутриклеточными белками цитоскелета.
- 58. Примеры интегринов: рецепторы для белков внеклеточного матрикса. Они связываются с гликопротеиновыми компонентами внеклеточного матрикса, в частности
- 59. Кадгерины – кальций-зависимые адгезивные молекулы. Три типа: нейрональные (N), плацентарные (Р), эпителиальные (Е). Участвуют в структурной
- 60. Молекулы адгезии в межклеточной коммуникации Взаимодействие трансмембранных молекул адгезии (кадгеринов) обеспечивает узнавание клеточных партнёров и их
- 61. L-селектин Р-селектин Е-селектин Внеклеточная часть селектинов состоит из 3 доменов: 1 представлен 2-9 блоками повторяющихся аминокислотных
- 63. Скачать презентацию
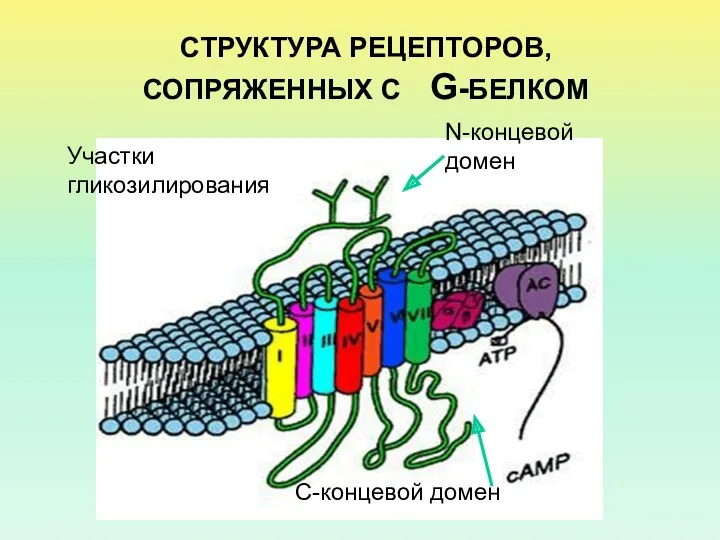
СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ
N-концевой домен
Участки гликозилирования
С-концевой домен
СТРУКТУРА РЕЦЕПТОРОВ,СОПРЯЖЕННЫХ С G-БЕЛКОМ
N-концевой домен
Участки гликозилирования
С-концевой домен

ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
АДРЕНАЛИН, НОРАДРЕНАЛИН
АЦЕТИЛХОЛИН
СЕРОТОНИН
ГИСТАМИН
ОПИОИДЫ, КАННАБИНОИДЫ
ГОРМОНЫ ПЕПТИДНОЙ
ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ
АДРЕНАЛИН, НОРАДРЕНАЛИН
АЦЕТИЛХОЛИН
СЕРОТОНИН
ГИСТАМИН
ОПИОИДЫ, КАННАБИНОИДЫ
ГОРМОНЫ ПЕПТИДНОЙ
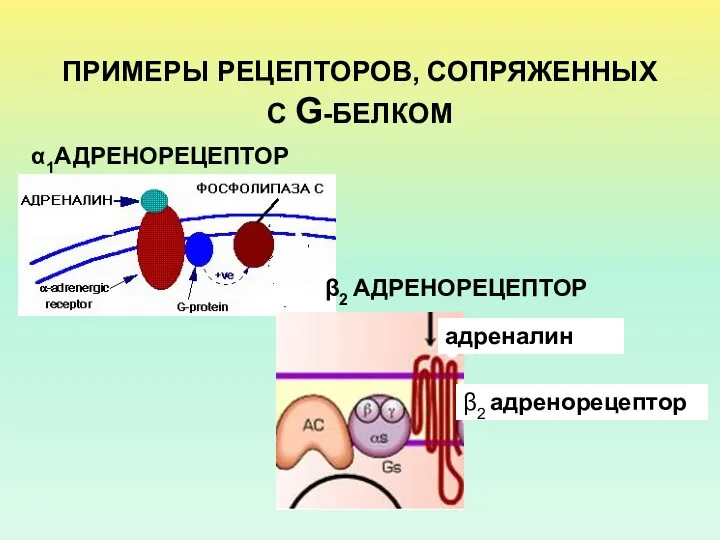
ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
α1АДРЕНОРЕЦЕПТОР
β2 АДРЕНОРЕЦЕПТОР
ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
α1АДРЕНОРЕЦЕПТОР
β2 АДРЕНОРЕЦЕПТОР
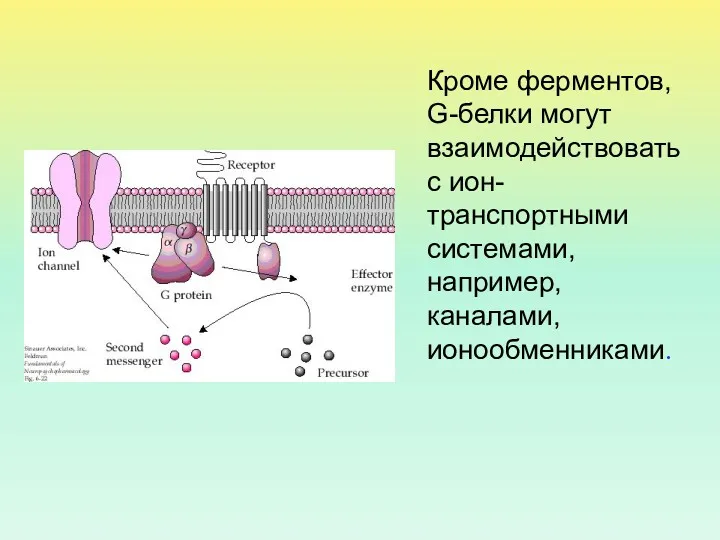
Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
Кроме ферментов, G-белки могут взаимодействовать с ион-транспортными системами, например, каналами, ионообменниками.
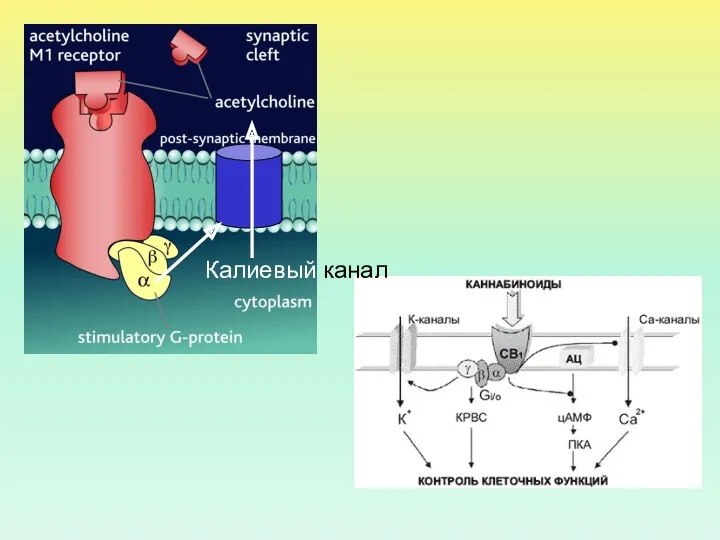
Калиевый канал
Калиевый канал
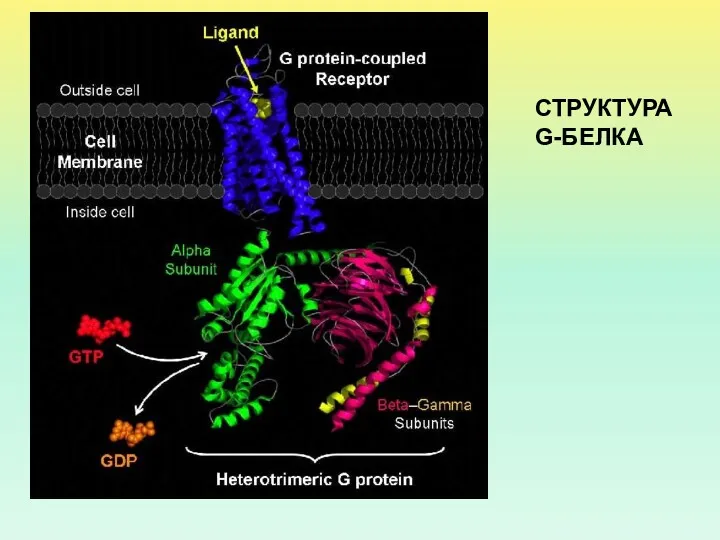
СТРУКТУРА G-БЕЛКА
СТРУКТУРА G-БЕЛКА
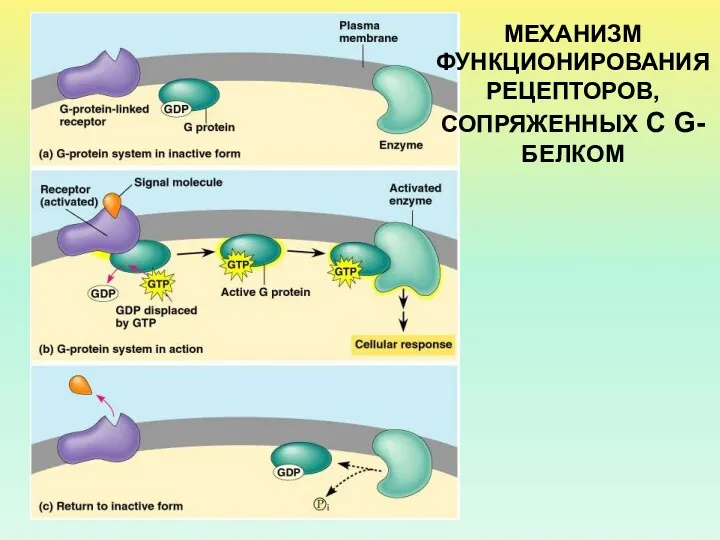
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ
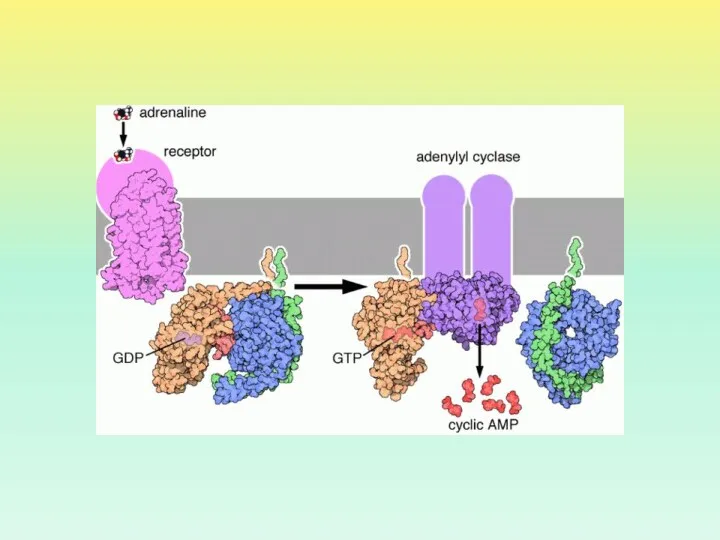

ВИДЫ G-БЕЛКОВ
ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ
GS И GI БЕЛКИ ДЕЙСТВУЮТ НА
ВИДЫ G-БЕЛКОВ
ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ
GS И GI БЕЛКИ ДЕЙСТВУЮТ НА
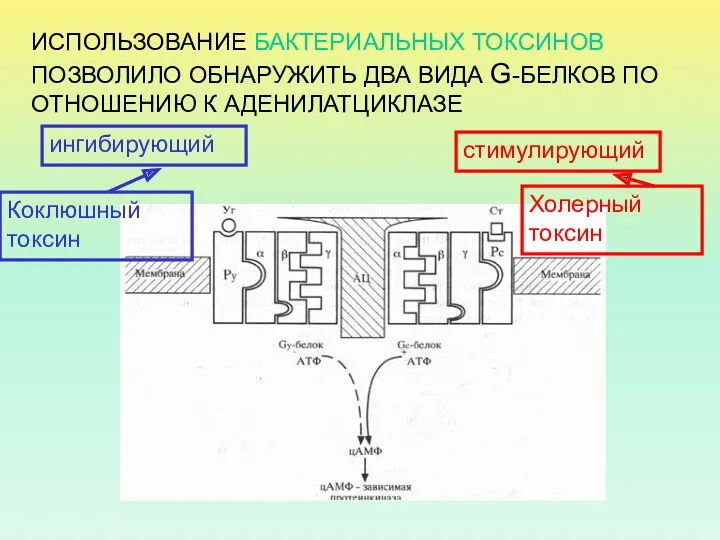
ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К
ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К
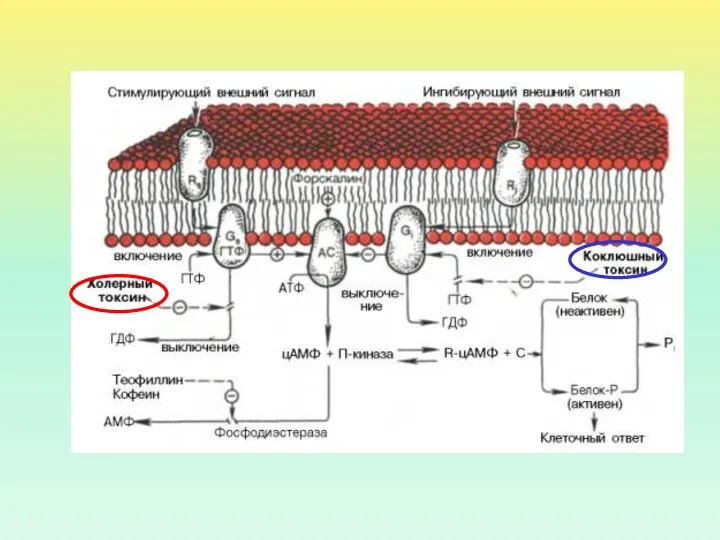

Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу.
АДФрибозилирование α-субъединицы
Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу.
АДФрибозилирование α-субъединицы
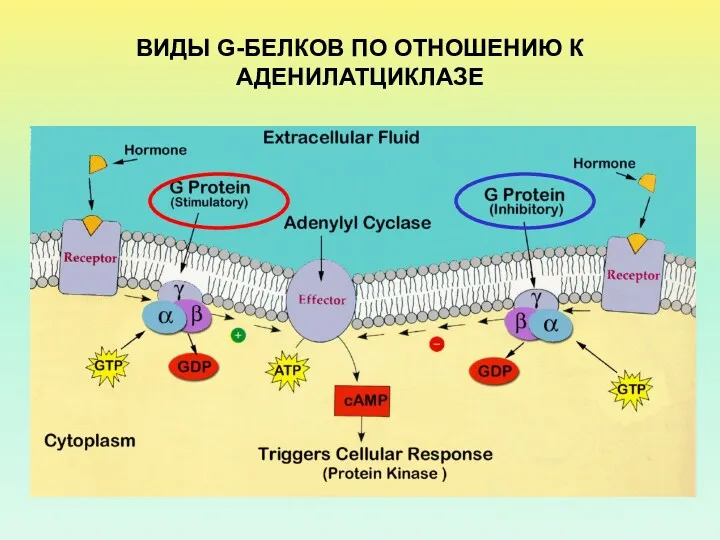
ВИДЫ G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ
ВИДЫ G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ

ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
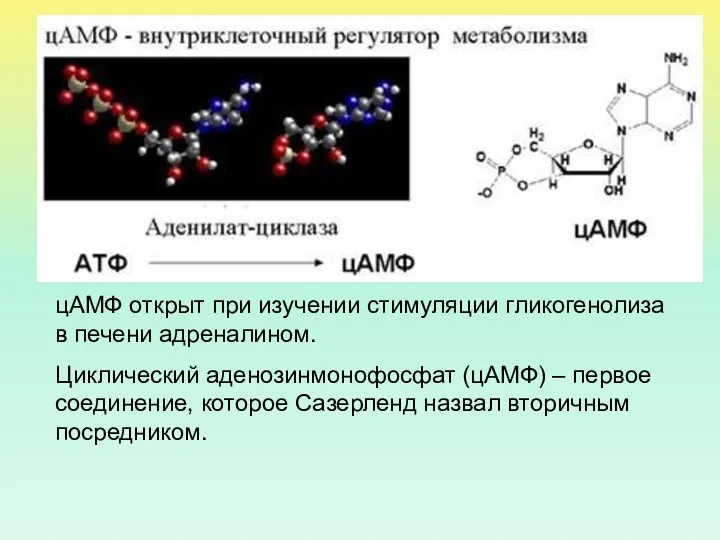
цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином.
Циклический аденозинмонофосфат
цАМФ открыт при изучении стимуляции гликогенолиза в печени адреналином.
Циклический аденозинмонофосфат
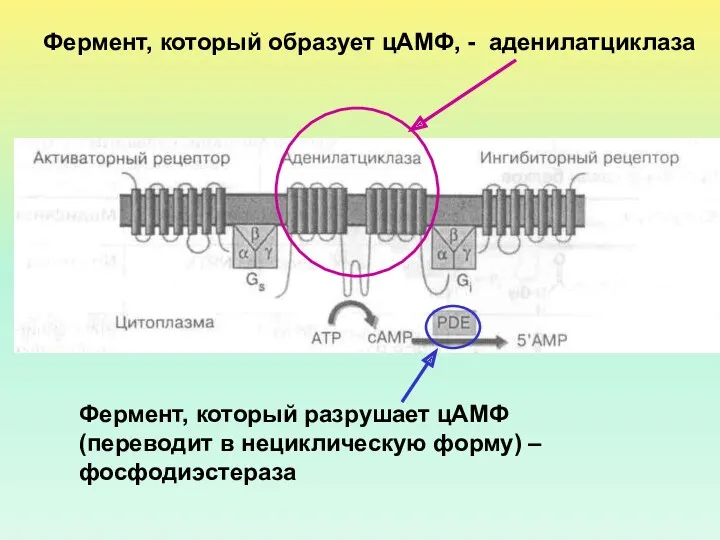
Фермент, который образует цАМФ, - аденилатциклаза
Фермент, который разрушает цАМФ (переводит
Фермент, который образует цАМФ, - аденилатциклаза
Фермент, который разрушает цАМФ (переводит
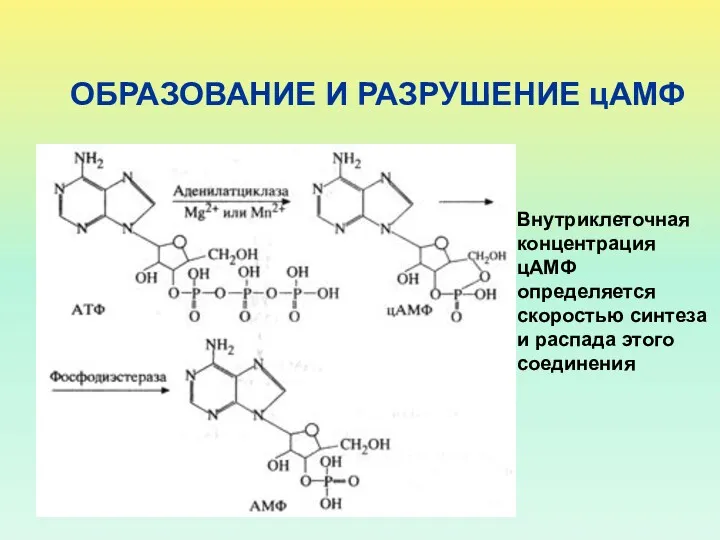
ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ
Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада
ОБРАЗОВАНИЕ И РАЗРУШЕНИЕ цАМФ
Внутриклеточная концентрация цАМФ определяется скоростью синтеза и распада
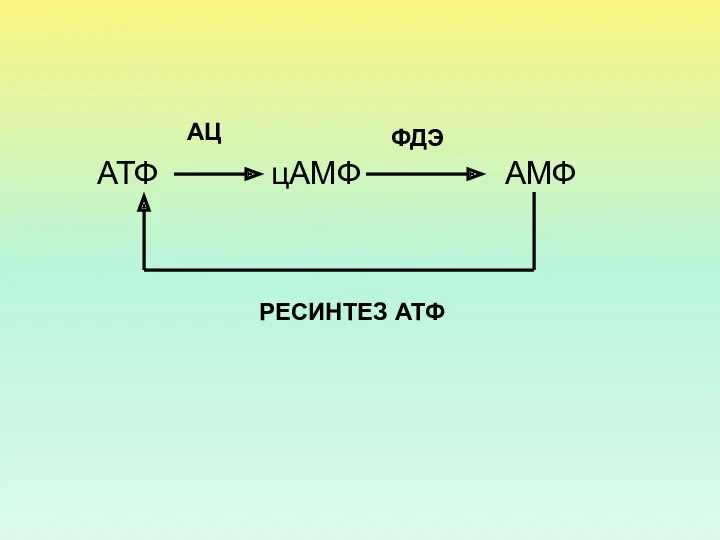
АТФ
АМФ
цАМФ
АЦ
ФДЭ
РЕСИНТЕЗ АТФ
АТФ
АМФ
цАМФ
АЦ
ФДЭ
РЕСИНТЕЗ АТФ
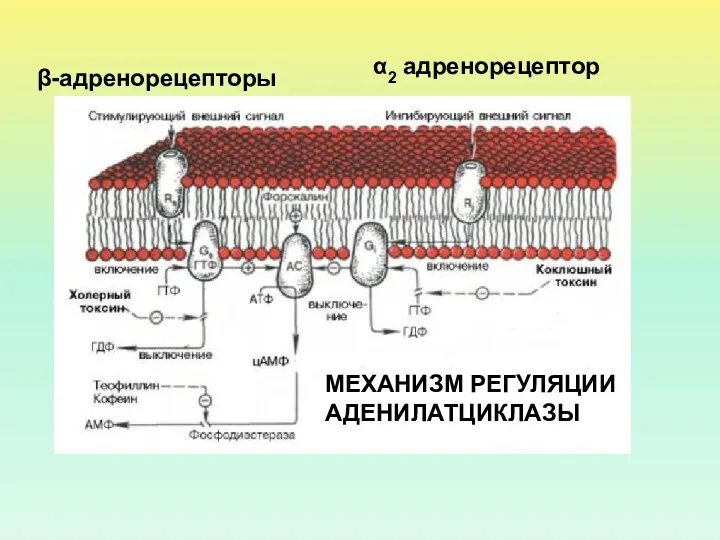
МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ
β-адренорецепторы
α2 адренорецептор
МЕХАНИЗМ РЕГУЛЯЦИИ АДЕНИЛАТЦИКЛАЗЫ
β-адренорецепторы
α2 адренорецептор
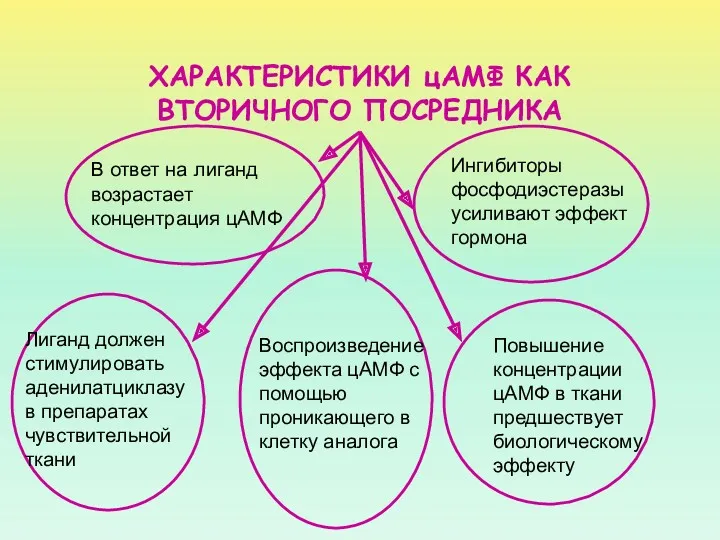
ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА
Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной
ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА
Лиганд должен стимулировать аденилатциклазу в препаратах чувствительной

ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
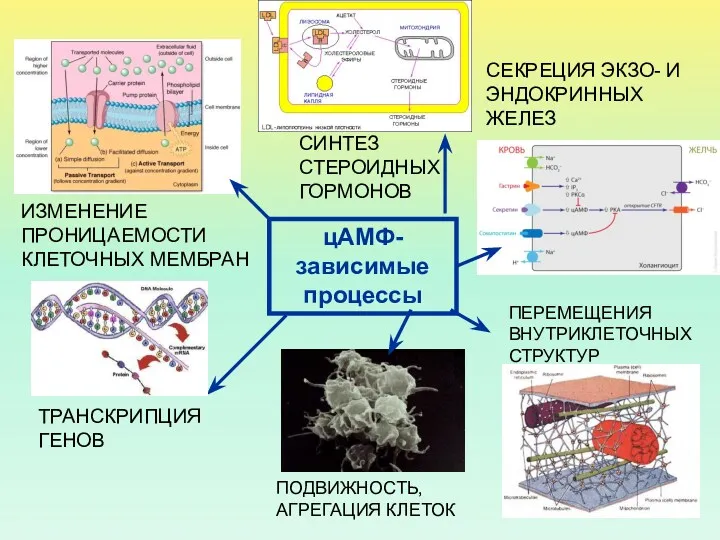
ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН
СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ
СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ
ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ
ИЗМЕНЕНИЕ ПРОНИЦАЕМОСТИ КЛЕТОЧНЫХ МЕМБРАН
СИНТЕЗ СТЕРОИДНЫХ ГОРМОНОВ
СЕКРЕЦИЯ ЭКЗО- И ЭНДОКРИННЫХ ЖЕЛЕЗ
ПЕРЕМЕЩЕНИЯ ВНУТРИКЛЕТОЧНЫХ
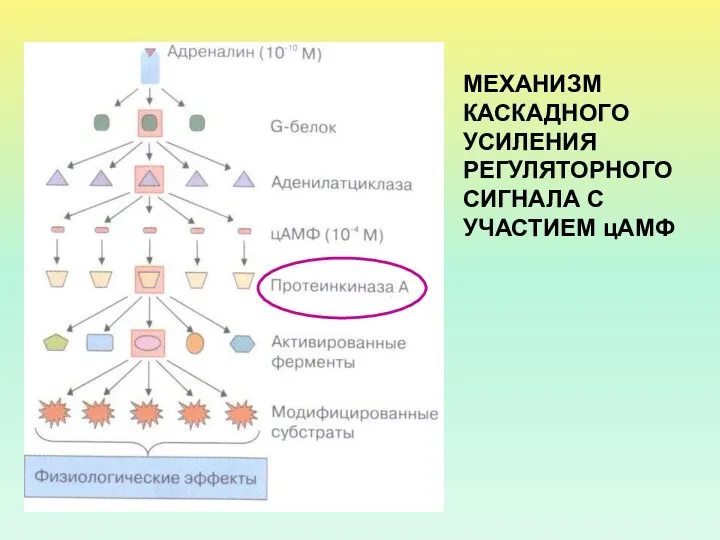
МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
МЕХАНИЗМ КАСКАДНОГО УСИЛЕНИЯ РЕГУЛЯТОРНОГО СИГНАЛА С УЧАСТИЕМ цАМФ
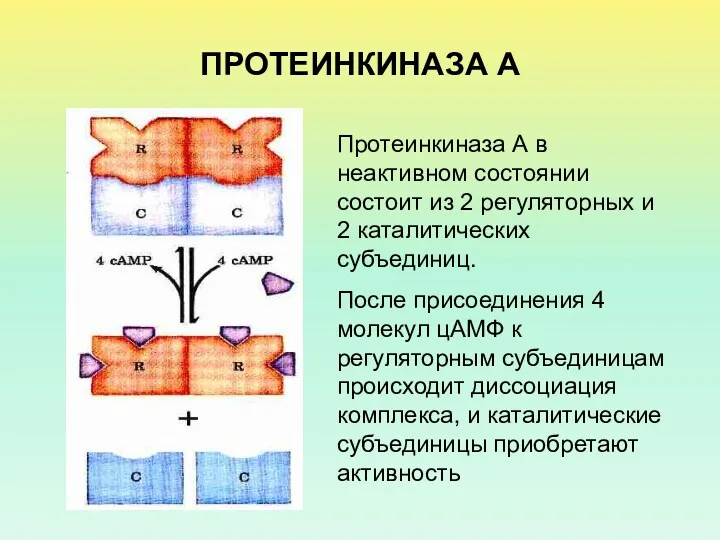
ПРОТЕИНКИНАЗА А
Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и
ПРОТЕИНКИНАЗА А
Протеинкиназа А в неактивном состоянии состоит из 2 регуляторных и
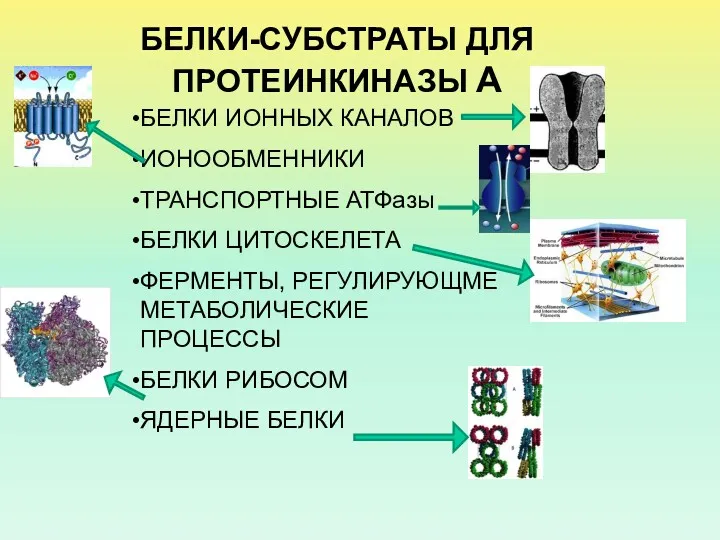
БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А
БЕЛКИ ИОННЫХ КАНАЛОВ
ИОНООБМЕННИКИ
ТРАНСПОРТНЫЕ АТФазы
БЕЛКИ ЦИТОСКЕЛЕТА
ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
БЕЛКИ
БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А
БЕЛКИ ИОННЫХ КАНАЛОВ
ИОНООБМЕННИКИ
ТРАНСПОРТНЫЕ АТФазы
БЕЛКИ ЦИТОСКЕЛЕТА
ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ МЕТАБОЛИЧЕСКИЕ ПРОЦЕССЫ
БЕЛКИ
цГМФ
ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
цГМФ
ОКАЗАЛОСЬ, ЧТО И цГМФ ЯВЛЯЕТСЯ ВТОРИЧНЫМ ПОСРЕДНИКОМ
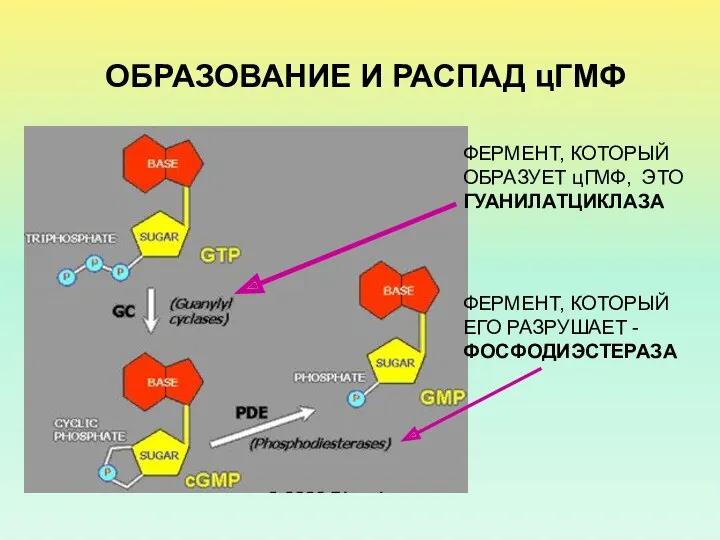
ОБРАЗОВАНИЕ И РАСПАД цГМФ
ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА
ФЕРМЕНТ, КОТОРЫЙ ЕГО
ОБРАЗОВАНИЕ И РАСПАД цГМФ
ФЕРМЕНТ, КОТОРЫЙ ОБРАЗУЕТ цГМФ, ЭТО ГУАНИЛАТЦИКЛАЗА
ФЕРМЕНТ, КОТОРЫЙ ЕГО
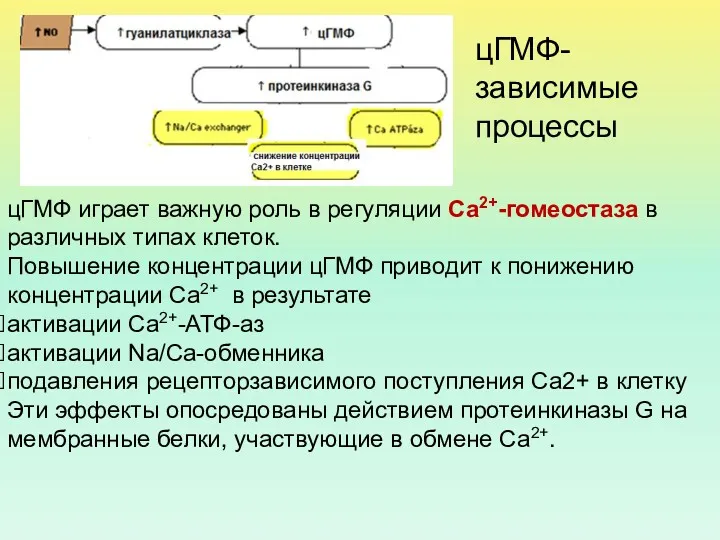
цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток.
цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток.

ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению
ПК G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению
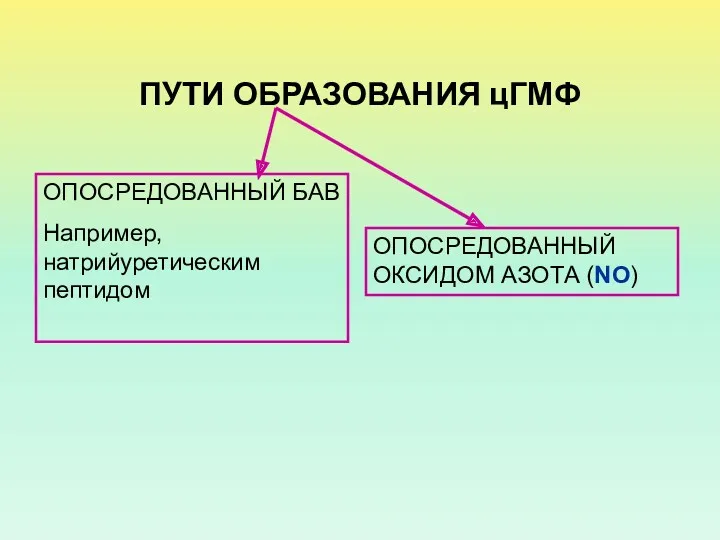
ПУТИ ОБРАЗОВАНИЯ цГМФ
ОПОСРЕДОВАННЫЙ БАВ
Например, натрийуретическим пептидом
ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
ПУТИ ОБРАЗОВАНИЯ цГМФ
ОПОСРЕДОВАННЫЙ БАВ
Например, натрийуретическим пептидом
ОПОСРЕДОВАННЫЙ ОКСИДОМ АЗОТА (NO)
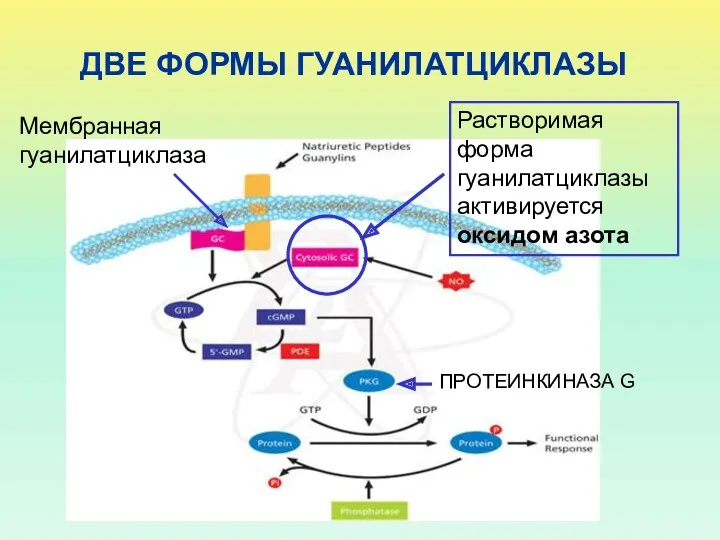
ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ
Мембранная гуанилатциклаза
Растворимая форма гуанилатциклазы активируется оксидом азота
ПРОТЕИНКИНАЗА G
ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ
Мембранная гуанилатциклаза
Растворимая форма гуанилатциклазы активируется оксидом азота
ПРОТЕИНКИНАЗА G

МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА
NaУП + R → ↑ГЦ → ↑цГМФ →
МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА
NaУП + R → ↑ГЦ → ↑цГМФ →

ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ
ОТСУТСТВУЮТ G-БЕЛКИ
СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ
ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ РЕГУЛЯЦИИ
ОТСУТСТВУЮТ G-БЕЛКИ
СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ
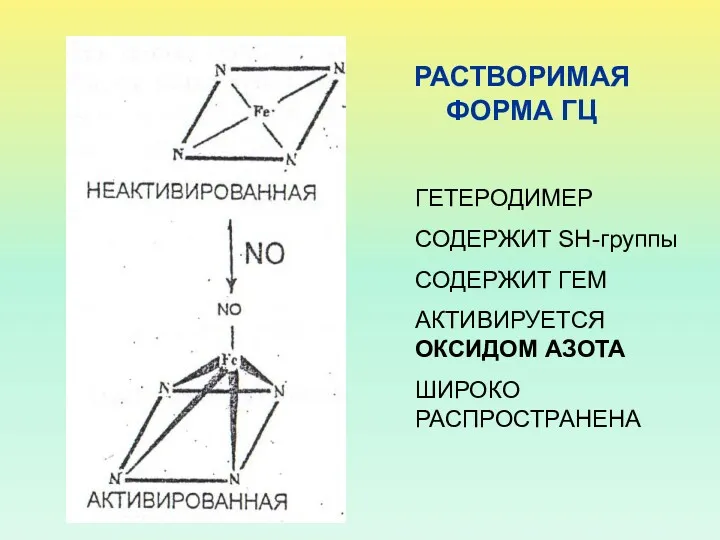
РАСТВОРИМАЯ ФОРМА ГЦ
ГЕТЕРОДИМЕР
СОДЕРЖИТ SH-группы
СОДЕРЖИТ ГЕМ
АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА
ШИРОКО РАСПРОСТРАНЕНА
РАСТВОРИМАЯ ФОРМА ГЦ
ГЕТЕРОДИМЕР
СОДЕРЖИТ SH-группы
СОДЕРЖИТ ГЕМ
АКТИВИРУЕТСЯ ОКСИДОМ АЗОТА
ШИРОКО РАСПРОСТРАНЕНА
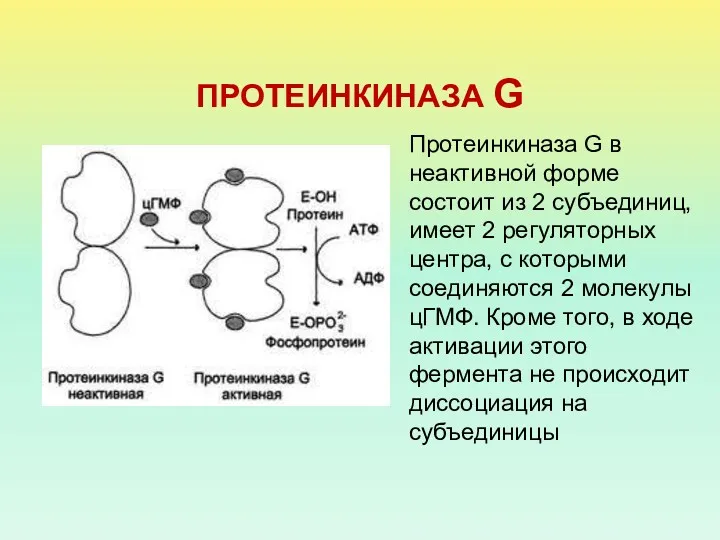
ПРОТЕИНКИНАЗА G
Протеинкиназа G в неактивной форме состоит из 2 субъединиц, имеет
ПРОТЕИНКИНАЗА G
Протеинкиназа G в неактивной форме состоит из 2 субъединиц, имеет

РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ

РЕЦЕПТОРНЫЕ ТИРОЗИНКИНАЗЫ
Агонисты:
факторы роста
инсулин
РЕЦЕПТОРНЫЕ ТИРОЗИНКИНАЗЫ
Агонисты:
факторы роста
инсулин

Факторы роста (GF) или митогены
стимулируют рост клетки и ее прохождение
Факторы роста (GF) или митогены
стимулируют рост клетки и ее прохождение
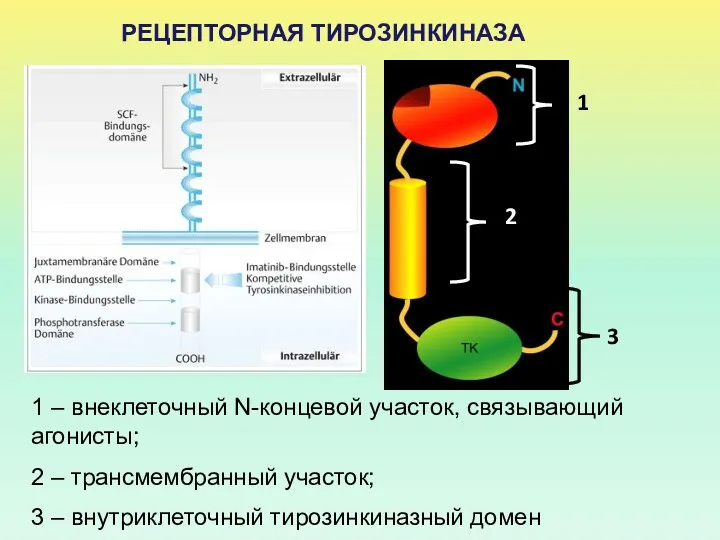
РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА
1 – внеклеточный N-концевой участок, связывающий агонисты;
2 – трансмембранный участок;
3
РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА
1 – внеклеточный N-концевой участок, связывающий агонисты;
2 – трансмембранный участок;
3
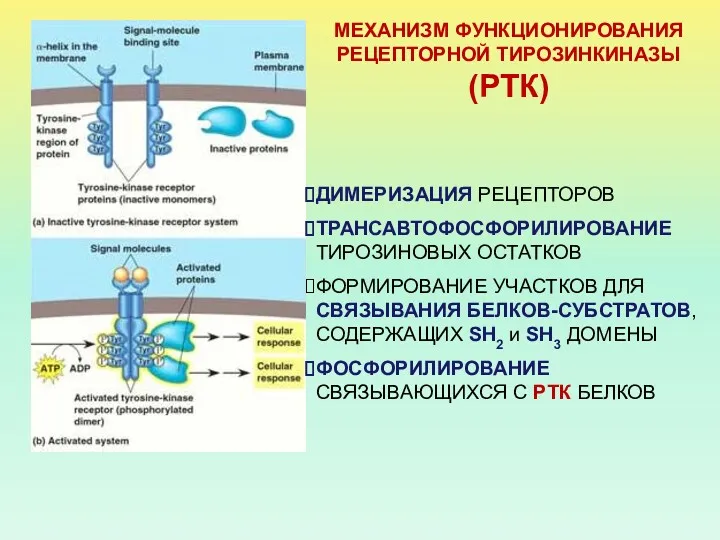
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ (РТК)
ДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ
ТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ
ФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ (РТК)
ДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ
ТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ
ФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ

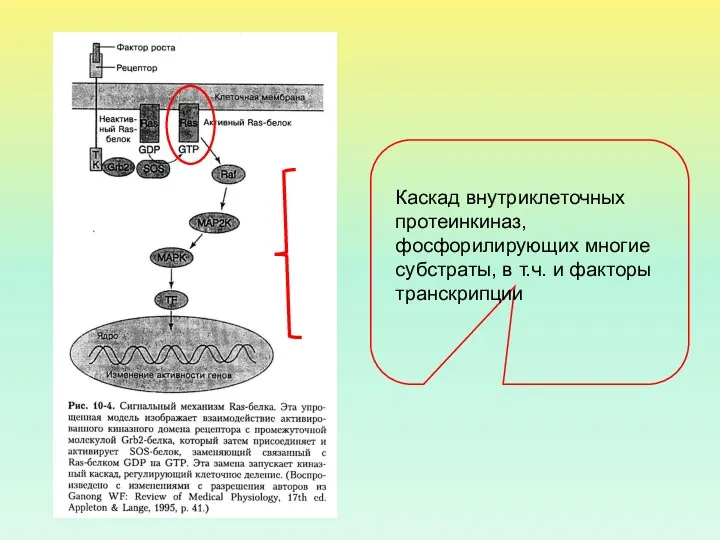
Ras-БЕЛОК – МОНОМЕРНАЯ ГТФаза
В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ С ГТФ, В
Ras-БЕЛОК – МОНОМЕРНАЯ ГТФаза
В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ С ГТФ, В
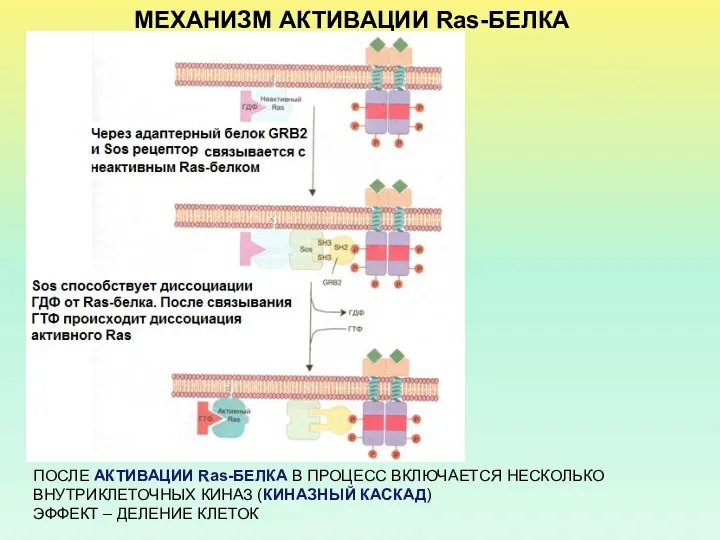
МЕХАНИЗМ АКТИВАЦИИ Ras-БЕЛКА
ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ
МЕХАНИЗМ АКТИВАЦИИ Ras-БЕЛКА
ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ
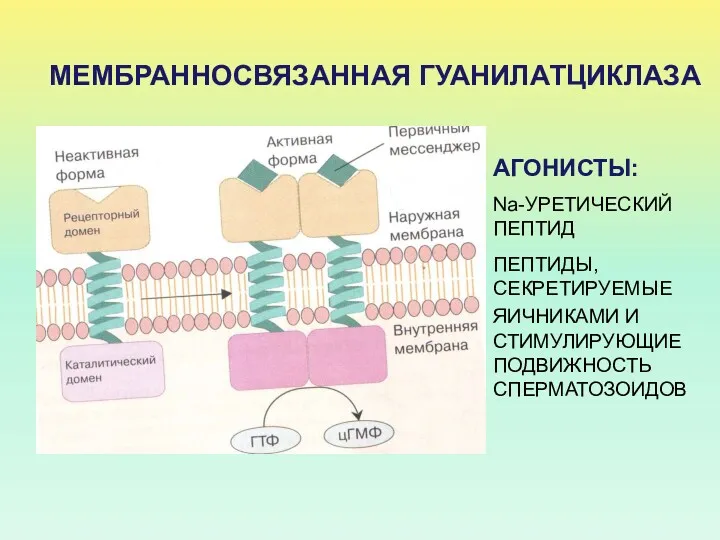
МЕМБРАННОСВЯЗАННАЯ ГУАНИЛАТЦИКЛАЗА
АГОНИСТЫ:
Na-УРЕТИЧЕСКИЙ ПЕПТИД
ПЕПТИДЫ, СЕКРЕТИРУЕМЫЕ ЯИЧНИКАМИ И СТИМУЛИРУЮЩИЕ ПОДВИЖНОСТЬ СПЕРМАТОЗОИДОВ
МЕМБРАННОСВЯЗАННАЯ ГУАНИЛАТЦИКЛАЗА
АГОНИСТЫ:
Na-УРЕТИЧЕСКИЙ ПЕПТИД
ПЕПТИДЫ, СЕКРЕТИРУЕМЫЕ ЯИЧНИКАМИ И СТИМУЛИРУЮЩИЕ ПОДВИЖНОСТЬ СПЕРМАТОЗОИДОВ

РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ
РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ

Эти рецепторы при активации связываются с цитоплазматическими тирозинкиназами и
образуют сигнальный
Эти рецепторы при активации связываются с цитоплазматическими тирозинкиназами и
образуют сигнальный
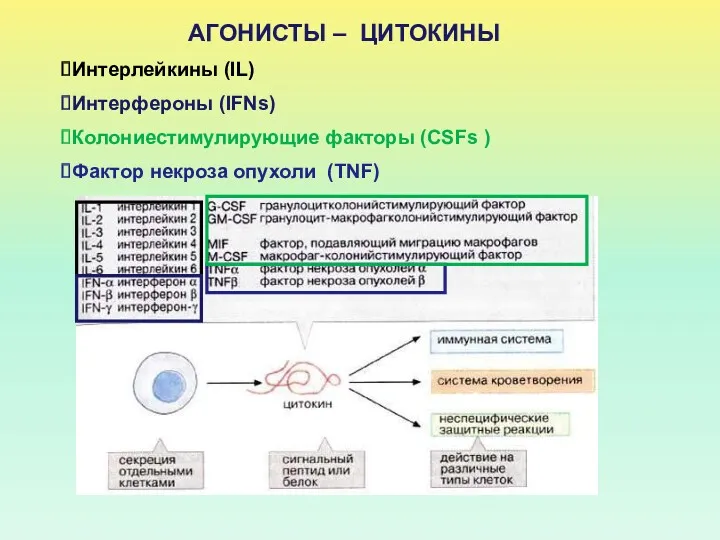
АГОНИСТЫ – ЦИТОКИНЫ
Интерлейкины (IL)
Интерфероны (IFNs)
Колониестимулирующие факторы (CSFs )
Фактор некроза опухоли (TNF)
АГОНИСТЫ – ЦИТОКИНЫ
Интерлейкины (IL)
Интерфероны (IFNs)
Колониестимулирующие факторы (CSFs )
Фактор некроза опухоли (TNF)

Цитокины — группа гормоноподобных белков и пептидов
синтезируются и секретируются клетками
Цитокины — группа гормоноподобных белков и пептидов
синтезируются и секретируются клетками

ЦИТОКИНОВЫЙ РЕЦЕПТОР
Эти рецепторы не имеют киназного участка в своем составе и
ЦИТОКИНОВЫЙ РЕЦЕПТОР
Эти рецепторы не имеют киназного участка в своем составе и
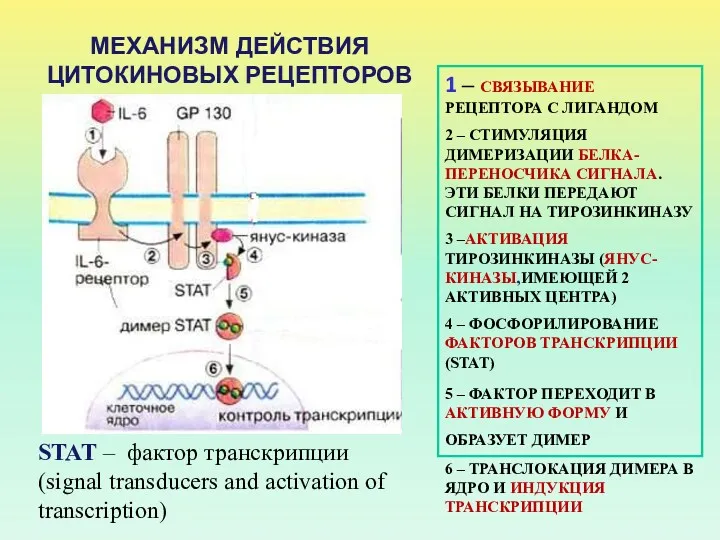
МЕХАНИЗМ ДЕЙСТВИЯ ЦИТОКИНОВЫХ РЕЦЕПТОРОВ
1 – СВЯЗЫВАНИЕ РЕЦЕПТОРА С ЛИГАНДОМ
2 – СТИМУЛЯЦИЯ
МЕХАНИЗМ ДЕЙСТВИЯ ЦИТОКИНОВЫХ РЕЦЕПТОРОВ
1 – СВЯЗЫВАНИЕ РЕЦЕПТОРА С ЛИГАНДОМ
2 – СТИМУЛЯЦИЯ

РЕЦЕПТОРЫ КЛЕТОЧНОЙ АДГЕЗИИ
ИНТЕГРИНЫ
КАДГЕРИНЫ
СЕЛЕКТИНЫ
РЕЦЕПТОРЫ КЛЕТОЧНОЙ АДГЕЗИИ
ИНТЕГРИНЫ
КАДГЕРИНЫ
СЕЛЕКТИНЫ
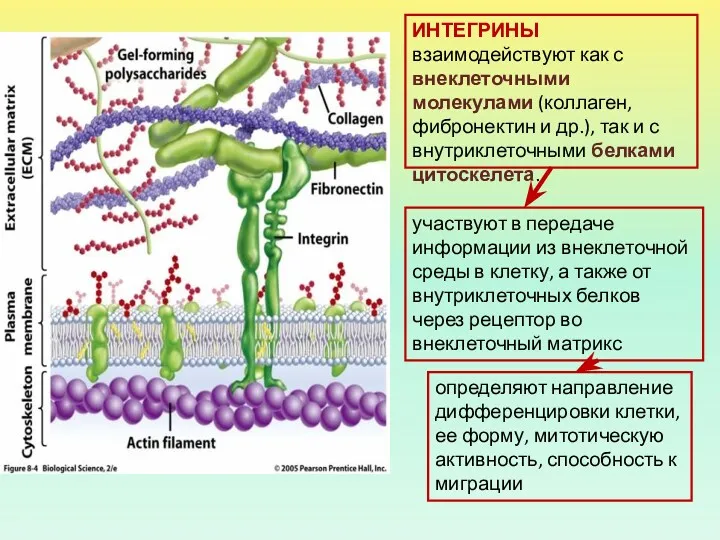
ИНТЕГРИНЫ взаимодействуют как с внеклеточными молекулами (коллаген, фибронектин и др.), так
ИНТЕГРИНЫ взаимодействуют как с внеклеточными молекулами (коллаген, фибронектин и др.), так

Примеры интегринов:
рецепторы для белков внеклеточного матрикса. Они связываются с гликопротеиновыми компонентами
рецепторы для белков внеклеточного матрикса. Они связываются с гликопротеиновыми компонентами

Кадгерины – кальций-зависимые адгезивные молекулы.
Три типа:
нейрональные (N),
плацентарные (Р),
Кадгерины – кальций-зависимые адгезивные молекулы.
Три типа:
нейрональные (N),
плацентарные (Р),
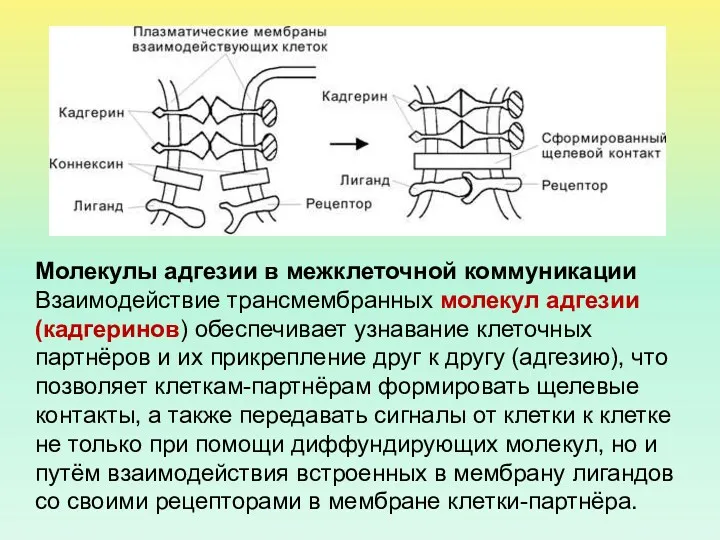
Молекулы адгезии в межклеточной коммуникации Взаимодействие трансмембранных молекул адгезии (кадгеринов) обеспечивает
Молекулы адгезии в межклеточной коммуникации Взаимодействие трансмембранных молекул адгезии (кадгеринов) обеспечивает
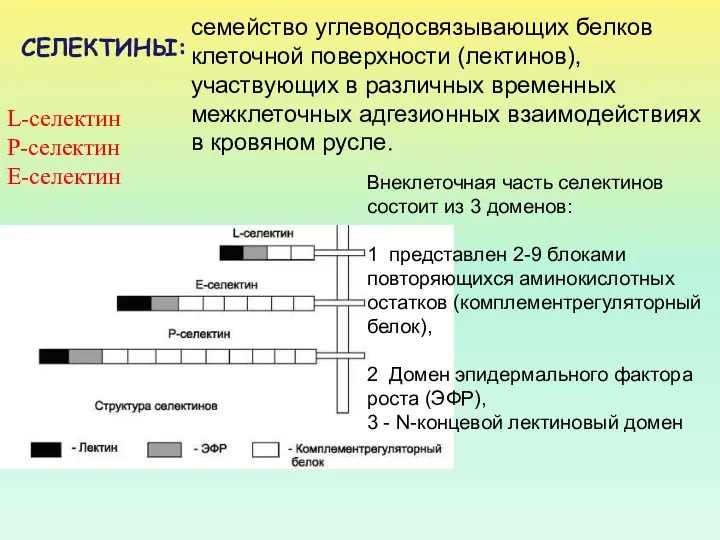
L-селектин
Р-селектин
Е-селектин
Внеклеточная часть селектинов состоит из 3 доменов:
1 представлен 2-9
L-селектин
Р-селектин
Е-селектин
Внеклеточная часть селектинов состоит из 3 доменов:
1 представлен 2-9
 Фотосинтез. Фазы фотосинтеза
Фотосинтез. Фазы фотосинтеза Ботаника. Салалары
Ботаника. Салалары Бактериялар
Бактериялар У Казахстані на основі гібридизації тонкорунних овець з диким гірським бараном Архар створена нова порода тонкорунних
У Казахстані на основі гібридизації тонкорунних овець з диким гірським бараном Архар створена нова порода тонкорунних Значение двигательной активности для организма человека
Значение двигательной активности для организма человека Дыхательная система
Дыхательная система Урок биологии в 10 классе Дигибридное скрещивание
Урок биологии в 10 классе Дигибридное скрещивание Порода собак такса
Порода собак такса Возрастная анатомия, физиология и гигиена
Возрастная анатомия, физиология и гигиена Группа ископаемых высших приматов австралопитеки
Группа ископаемых высших приматов австралопитеки Индивидуальные параметры тела
Индивидуальные параметры тела Клетка. Структура клетки. Органоиды клетки
Клетка. Структура клетки. Органоиды клетки Сравнительный анализ проращивания семян горчицы в разных образцах талой воды и дистиллированной воды
Сравнительный анализ проращивания семян горчицы в разных образцах талой воды и дистиллированной воды Слюнные железы
Слюнные железы Medical Academy Named after S. I. Georgievsky of Crimea
Medical Academy Named after S. I. Georgievsky of Crimea Тройничный нерв
Тройничный нерв Физиология высшей нервной нервной деятельности. Условные и безусловные рефлексы
Физиология высшей нервной нервной деятельности. Условные и безусловные рефлексы Тип Членистоногие
Тип Членистоногие Ученые - биологи во время Великой Отечественной войны
Ученые - биологи во время Великой Отечественной войны Человек. Общий обзор организма. (ОГЭ. Тест 1)
Человек. Общий обзор организма. (ОГЭ. Тест 1) Анатомия дыхательной системы
Анатомия дыхательной системы Своя игра
Своя игра Физиология лактации коров
Физиология лактации коров Урок ДОТ
Урок ДОТ Организм и среда. Наземно-воздушная среда жизни
Организм и среда. Наземно-воздушная среда жизни Основные понятия генетики
Основные понятия генетики Пшеница – важнейшая зерновая культура
Пшеница – важнейшая зерновая культура Виды рыб
Виды рыб