- Соединительная ткань

Содержание
- 2. Специализированной частью межклеточного матрикса являются базальные мембраны, из которых "растут" все клетки организма, кроме клеток соединительной
- 3. Базальные мембраны с элементами соединительной ткани имеются между эпидермальным и дермальными слоями кожи, под эпителием, выстилающим
- 4. Функции соединительной ткани -опорная - она соединяет клетки и ткани, образует каркас организма, -трофическая - формируя
- 5. Функции соединительной ткани структурно-образовательная - эта функция связана с регулирующим влиянием коллагена и глюкозамингликанов на рост
- 6. Соединительную ткань отличает от других тканей большие промежутки между клетками и соответственно большое количество межклеточного вещества.
- 7. Межклеточное вещество в нормальных условиях представляет собой гель и состоит из "тканевых структурных биоколлоидов“ - белков:
- 8. В зависимости от типа соединительной ткани фибриллярные элементы представлены тремя типами волокон: коллагеновые, эластиновые и ретикулярные
- 9. Склеропротеины соединительной ткани - коллаген, эластин и кератины. Важнейшими склеропротеинами являются : Коллаген, который представляет главную
- 10. Коллаген – самый распространенный в животном мире белок. На его долю приходится 30% от всего количества
- 11. Коллаген как гликопротеин: содержит галактозильные и лактозильные остатки, соединенные с гидроксильными группами оксилизина. Каждая из трех
- 12. Синтез коллагена Вначале из трех спирализованных полипептидных цепей, синтезируемых фибробластами, формируется молекула проколлагена, где полипептидные цепи
- 13. В процессе созревания полипептидных цепей проколлагена под воздействием специфических гидроксилаз и в присутствии витамина С происходит
- 14. Созревание проколлагена -NH - С=О -NH - С=О | | СН-NH СН-NH | | (СН2)2 (СН2)2
- 15. Гидроксильные группы лизина подвергаются гликолизированию и галактозированию – в результате у проколлагена наращиваются углеводные фрагменты. Суперспирализованные
- 16. Тропоколлаген Молекула тропоколлагена (мол. масса 120 тыс. дальтон) является структурной единицей коллагена Отдельные молекулы тропоколлагена соединяясь
- 17. Затем, за счет альдегидных групп отдельные молекулы тропоколлагена соединяются друг с другом различного типа ковалентными связями
- 18. Укладка коллагеновых фибрил Поперечные ковалентные связи между мономерами тропоколлагена расположены не беспорядочно, а распределены в определенных
- 19. В молекулах проколлагена, тропоколлагена и коллагена, построенных из трех полипептидных цепей, свёрнутых в тройную спираль, имеются
- 20. Разновидности коллагена. Коллаген I - характерен для большинства типов соединительной ткани (коллаген костей, кожи и сухожилий)
- 21. Коллаген IV содержит три L1(IV) субъединицы. [ L1(IV) ]3 Коллаген как внеклеточный белок определяет формирование структуры
- 22. При недостатке в организме витамина С нарушается гидроксилирование пролина и лизина, снижается активность лизилоксидазы, что приводит
- 23. Ключевую роль в катаболизма коллагена играет специфический фермент коллагеназа, разрушающий пептидную связь в молекуле коллагена, образованную
- 24. Распад коллагена и увеличение экскреции оксипролина с мочой наблюдается: при гиперпаратиреоидизме, болезни Педжета. Синтез коллагена усиливается
- 25. Эластин. Структура эластиновых волокон соединительной ткани формируется из молекул белка эластина, молекула которого построена из четырех
- 26. Синтезируется эластин фибробластами в виде проэластина, который дозревает в тропоэластин. Молекулы тропоэластина не имеют поперечных связей,
- 27. Дегидратации и окисления трех альдегидных групп лизина с аминогруппой четвертой молекулы лизина, что приводит к образованию
- 28. Эластин резиноподобный белок - основной компонент эластических волокон соединительной ткани. Он содержится в больших количествах в
- 29. Кератины Кератины являются белками, участвующими в построении эпидермиса и его производных, как волосы, ногти и у
- 30. Молекула кератинов содержит большое количество нейтральных и основных аминокислот. Много цистина, серина и треонина. Совсем отсутствует
- 31. Установлено, что главный компонент кератиновых волокон представлен кератином- белком с "низким содержанием серы", имеющий альфа-спиральную конфигурацию
- 32. Развернутые (растянутые) цепи - бета-кератин. Бета-кератин может быть произведен из альфа-типа мягких кератинов. Например, при воздействии
- 33. Гликозамингликаны соединительной ткани. Глюкозамингликаны в соединении с белками межклеточного матрикса формируют, так называемые, протеогликаны соединительной ткани.
- 34. Гиалуронидаза разрушая гиалуроновую кислоту повышает межклеточную проницаемость. Некоторые бактерии секретируют гиалуронидазу, что позволяет им проникать из
- 35. Хондроитинсульфаты. Мономер этих полисахаридов состоит из глюкуроновой кислоты и N-ацетилгалактозамином, у которого в зависимости от типа
- 36. Кератансульфаты состоят из мономеров образованных бета-галактозой и N-ацетилглюкозамин-6 сульфата. Кератансульфаты находятся в роговице глаза. Гликозамингликаны несут
- 37. Протеогликаны соединительной ткани Протеогликаны различаются набором гликозамингликанов, размером молекулы и содержанием белка. Встречаются соединения образованные небольшими
- 38. Единый протеогликановый комплекс межклеточного матрикса образуется путем присоединения таких субъединиц к гигантской молекуле гиалуроновой кислоты. Одна
- 39. Вследствие отталкивания одноименно заряженных сульфатированных цепей гликозамингликанов, протеогликаны в водной среде " распушены" и хорошо гидратированы.
- 40. Но поскольку цепи одноименно заряжен, сопротивление сдавливанию нарастает по мере сжатия молекул. Если механическое сдавливание прекратить,
- 41. Межклеточный матрикс Межклеточный матрикс формируется из комплексов образованных гликозамингликанами, протеогликанами и коллагеновыми волокнами. Объединяясь между собой
- 42. Один из них фибронектин, который состоит из двух полипептидных идентичных цепей, соединенных вблизи С-конца двумя дисульфидными
- 43. Фибронектин также присоединяется к коллагену, к гиалуроновой кислоте и сульфированным гликозамингликанам протеогликанов. Фибронектин связывает фермент трансглутаминазу.
- 44. В результате этих процессов формируется структура межклеточного матрикса фиксируемая прочными ковалентными связями. В разных тканях и
- 45. Базальная мембрана Базальная мембрана высокоспециализированная часть межклеточного матрикса имеет особое строение. Она одной стороны контактирует с
- 46. Базальная мембрана имеет отрицательный заряд за счет сульфатированных гликозамингликанов и непроницаема для многих белков плазмы крови.
- 47. Чтобы достигнуть клеток, транспортируемые с кровью питательные вещества (углеводы, аминокислоты, минеральные соли и др.), должны последовательно
- 48. Изменения соединительной ткани при старении С возрастом в соединительной ткани происходят значительные изменения качественного состава и
- 49. С возрастом в костях вместо коллагена I накапливается коллаген III, что приводит к повышенной ломкости костей.
- 50. Изменения соединительной ткани при коллагенозах Группа заболеваний связанных с изменениями структуры соединительной ткани, нарушениями синтеза коллагена
- 51. Известны наследственные патологии, связанные с нарушениями синтеза коллагена. Так при синдроме Элерса-Данлоса имеет место деформация суставов
- 52. Наследственно обусловленная хрупкость костей связана с содержанием в костях наряду с коллагеном I, коллагена типа III.
- 53. При употреблении в пищу большого количества сладкого зеленого горошка в котором имеется ингибитор лизилоксидазы - бетацианаланин
- 54. Гормоны и соединительная ткань Формирование соединительной ткани контролируется гормонами. Глюкокортикоиды угнетают рост и развитие фибробластов и
- 55. Эстрогены ускоряют синтез кислых гликозамингликанов, Тироксин подавляет образование коллагеновых фибрилл и основного вещества. При недостатке тироксина
- 57. Скачать презентацию

Специализированной частью межклеточного матрикса являются базальные мембраны, из которых "растут" все
Специализированной частью межклеточного матрикса являются базальные мембраны, из которых "растут" все

Базальные мембраны с элементами соединительной ткани имеются между
эпидермальным и дермальными
Базальные мембраны с элементами соединительной ткани имеются между
эпидермальным и дермальными

Функции соединительной ткани
-опорная - она соединяет клетки и ткани, образует
Функции соединительной ткани
-опорная - она соединяет клетки и ткани, образует

Функции соединительной ткани
структурно-образовательная - эта функция связана с регулирующим влиянием
Функции соединительной ткани
структурно-образовательная - эта функция связана с регулирующим влиянием

Соединительную ткань отличает от других тканей большие промежутки между клетками и
Соединительную ткань отличает от других тканей большие промежутки между клетками и

Межклеточное вещество в нормальных условиях представляет собой гель и состоит из

В зависимости от типа соединительной ткани фибриллярные элементы представлены тремя типами
В зависимости от типа соединительной ткани фибриллярные элементы представлены тремя типами

Склеропротеины соединительной ткани - коллаген, эластин и кератины.
Важнейшими склеропротеинами являются
Склеропротеины соединительной ткани - коллаген, эластин и кератины.
Важнейшими склеропротеинами являются

Коллаген –
самый распространенный в животном мире белок. На его
Коллаген –
самый распространенный в животном мире белок. На его

Коллаген как гликопротеин:
содержит галактозильные и лактозильные остатки, соединенные с гидроксильными
Коллаген как гликопротеин:
содержит галактозильные и лактозильные остатки, соединенные с гидроксильными

Синтез коллагена
Вначале из трех спирализованных полипептидных цепей, синтезируемых фибробластами, формируется молекула
Синтез коллагена
Вначале из трех спирализованных полипептидных цепей, синтезируемых фибробластами, формируется молекула
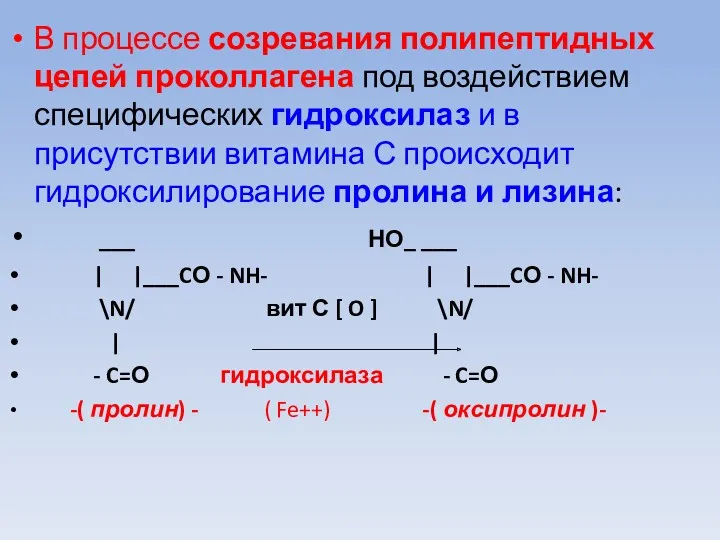
В процессе созревания полипептидных цепей проколлагена под воздействием специфических гидроксилаз и
В процессе созревания полипептидных цепей проколлагена под воздействием специфических гидроксилаз и
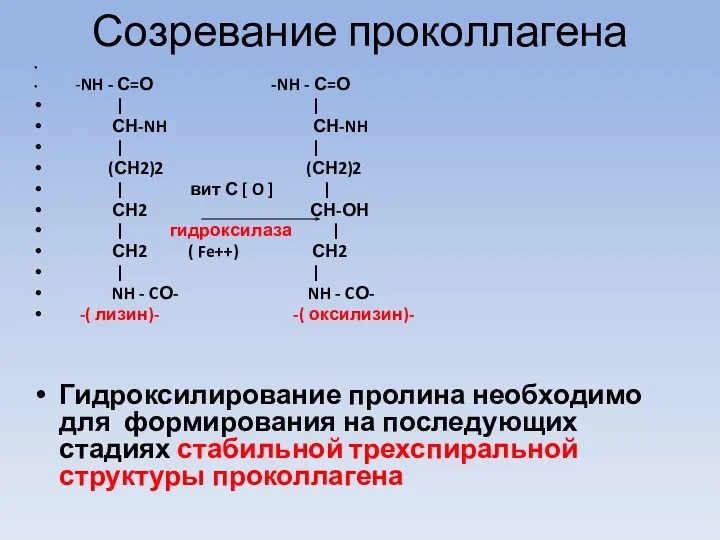
Созревание проколлагена
-NH - С=О -NH - С=О
| |
Созревание проколлагена
-NH - С=О -NH - С=О
| |

Гидроксильные группы лизина подвергаются гликолизированию и галактозированию –
в результате у
Гидроксильные группы лизина подвергаются гликолизированию и галактозированию –
в результате у
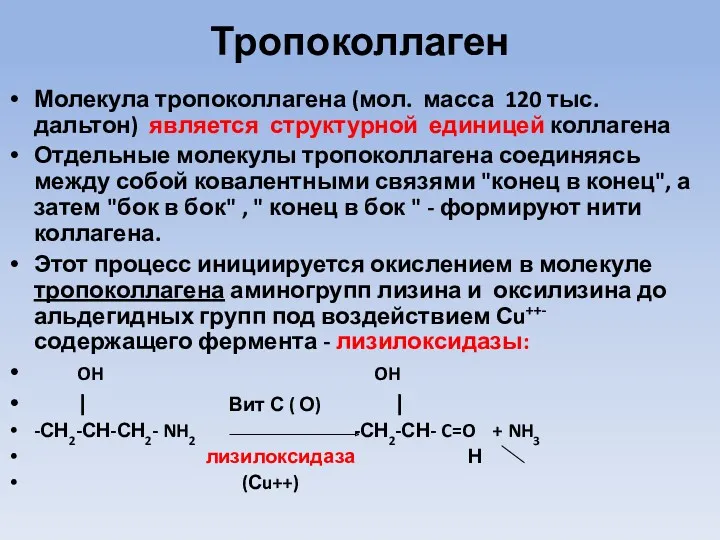
Тропоколлаген
Молекула тропоколлагена (мол. масса 120 тыс. дальтон) является структурной единицей коллагена
Тропоколлаген
Молекула тропоколлагена (мол. масса 120 тыс. дальтон) является структурной единицей коллагена

Затем, за счет альдегидных групп отдельные молекулы тропоколлагена соединяются друг с

Укладка коллагеновых фибрил
Поперечные ковалентные связи между мономерами тропоколлагена расположены не беспорядочно,
Укладка коллагеновых фибрил
Поперечные ковалентные связи между мономерами тропоколлагена расположены не беспорядочно,

В молекулах проколлагена, тропоколлагена и коллагена, построенных из трех полипептидных цепей,

Разновидности коллагена.
Коллаген I - характерен для большинства типов соединительной ткани (коллаген
Разновидности коллагена.
Коллаген I - характерен для большинства типов соединительной ткани (коллаген
![Коллаген IV содержит три L1(IV) субъединицы. [ L1(IV) ]3 Коллаген](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/251092/slide-20.jpg)
Коллаген IV содержит три L1(IV) субъединицы. [ L1(IV) ]3
Коллаген как
Коллаген как

При недостатке в организме витамина С нарушается гидроксилирование пролина и лизина,
При недостатке в организме витамина С нарушается гидроксилирование пролина и лизина,

Ключевую роль в катаболизма коллагена играет специфический фермент коллагеназа,
разрушающий пептидную
разрушающий пептидную

Распад коллагена и увеличение экскреции оксипролина с мочой наблюдается:
при гиперпаратиреоидизме,
при гиперпаратиреоидизме,

Эластин.
Структура эластиновых волокон соединительной ткани формируется из молекул белка эластина,
молекула
Эластин.
Структура эластиновых волокон соединительной ткани формируется из молекул белка эластина,
молекула

Синтезируется эластин фибробластами в виде проэластина, который дозревает в тропоэластин.
Молекулы
Синтезируется эластин фибробластами в виде проэластина, который дозревает в тропоэластин.
Молекулы

Дегидратации и окисления трех альдегидных групп лизина с аминогруппой четвертой молекулы

Эластин резиноподобный белок - основной компонент эластических волокон соединительной ткани.
Он
Эластин резиноподобный белок - основной компонент эластических волокон соединительной ткани.
Он

Кератины
Кератины являются белками, участвующими в построении эпидермиса и его производных, как
Кератины
Кератины являются белками, участвующими в построении эпидермиса и его производных, как

Молекула кератинов содержит большое количество нейтральных и основных аминокислот.
Много цистина,
Молекула кератинов содержит большое количество нейтральных и основных аминокислот.
Много цистина,

Установлено, что главный компонент кератиновых волокон представлен кератином- белком с "низким
Установлено, что главный компонент кератиновых волокон представлен кератином- белком с "низким

Развернутые (растянутые) цепи - бета-кератин.
Бета-кератин может быть произведен из альфа-типа
Развернутые (растянутые) цепи - бета-кератин.
Бета-кератин может быть произведен из альфа-типа

Гликозамингликаны соединительной ткани.
Глюкозамингликаны в соединении с белками межклеточного матрикса формируют, так
Гликозамингликаны соединительной ткани.
Глюкозамингликаны в соединении с белками межклеточного матрикса формируют, так

Гиалуронидаза разрушая гиалуроновую кислоту повышает межклеточную проницаемость.
Некоторые бактерии секретируют гиалуронидазу,
Гиалуронидаза разрушая гиалуроновую кислоту повышает межклеточную проницаемость.
Некоторые бактерии секретируют гиалуронидазу,

Хондроитинсульфаты.
Мономер этих полисахаридов состоит из глюкуроновой кислоты и N-ацетилгалактозамином, у
Хондроитинсульфаты.
Мономер этих полисахаридов состоит из глюкуроновой кислоты и N-ацетилгалактозамином, у

Кератансульфаты состоят из мономеров образованных бета-галактозой и N-ацетилглюкозамин-6 сульфата. Кератансульфаты находятся
Кератансульфаты состоят из мономеров образованных бета-галактозой и N-ацетилглюкозамин-6 сульфата. Кератансульфаты находятся

Протеогликаны соединительной ткани
Протеогликаны различаются набором гликозамингликанов, размером молекулы и содержанием белка.
Протеогликаны соединительной ткани
Протеогликаны различаются набором гликозамингликанов, размером молекулы и содержанием белка.

Единый протеогликановый комплекс межклеточного матрикса образуется путем присоединения таких субъединиц к
Единый протеогликановый комплекс межклеточного матрикса образуется путем присоединения таких субъединиц к

Вследствие отталкивания одноименно заряженных сульфатированных цепей гликозамингликанов, протеогликаны в водной среде
Вследствие отталкивания одноименно заряженных сульфатированных цепей гликозамингликанов, протеогликаны в водной среде

Но поскольку цепи одноименно заряжен, сопротивление сдавливанию нарастает по мере сжатия
Но поскольку цепи одноименно заряжен, сопротивление сдавливанию нарастает по мере сжатия

Межклеточный матрикс
Межклеточный матрикс формируется из комплексов образованных гликозамингликанами, протеогликанами и коллагеновыми
Межклеточный матрикс
Межклеточный матрикс формируется из комплексов образованных гликозамингликанами, протеогликанами и коллагеновыми

Один из них фибронектин, который состоит из двух полипептидных идентичных цепей,
Один из них фибронектин, который состоит из двух полипептидных идентичных цепей,

Фибронектин также присоединяется к коллагену, к гиалуроновой кислоте и сульфированным гликозамингликанам

В результате этих процессов формируется структура межклеточного матрикса фиксируемая прочными ковалентными
В результате этих процессов формируется структура межклеточного матрикса фиксируемая прочными ковалентными

Базальная мембрана
Базальная мембрана высокоспециализированная часть межклеточного матрикса имеет особое строение. Она
Базальная мембрана
Базальная мембрана высокоспециализированная часть межклеточного матрикса имеет особое строение. Она

Базальная мембрана имеет отрицательный заряд за счет сульфатированных гликозамингликанов и непроницаема
Базальная мембрана имеет отрицательный заряд за счет сульфатированных гликозамингликанов и непроницаема

Чтобы достигнуть клеток, транспортируемые с кровью питательные вещества (углеводы, аминокислоты, минеральные

Изменения соединительной ткани при старении
С возрастом в соединительной ткани происходят значительные
Изменения соединительной ткани при старении
С возрастом в соединительной ткани происходят значительные

С возрастом в костях вместо коллагена I накапливается коллаген III, что
С возрастом в костях вместо коллагена I накапливается коллаген III, что

Изменения соединительной ткани при коллагенозах
Группа заболеваний связанных с изменениями структуры соединительной
Изменения соединительной ткани при коллагенозах
Группа заболеваний связанных с изменениями структуры соединительной

Известны наследственные патологии, связанные с нарушениями синтеза коллагена.
Так при синдроме
Известны наследственные патологии, связанные с нарушениями синтеза коллагена.
Так при синдроме

Наследственно обусловленная хрупкость костей связана с содержанием в костях наряду с
Наследственно обусловленная хрупкость костей связана с содержанием в костях наряду с

При употреблении в пищу большого количества сладкого зеленого горошка
в котором
При употреблении в пищу большого количества сладкого зеленого горошка
в котором

Гормоны и соединительная ткань
Формирование соединительной ткани контролируется гормонами.
Глюкокортикоиды угнетают рост
Гормоны и соединительная ткань
Формирование соединительной ткани контролируется гормонами.
Глюкокортикоиды угнетают рост

Эстрогены ускоряют синтез кислых гликозамингликанов,
Тироксин подавляет образование коллагеновых фибрилл и
Эстрогены ускоряют синтез кислых гликозамингликанов,
Тироксин подавляет образование коллагеновых фибрилл и
 Насекомые. Подготовительная группа
Насекомые. Подготовительная группа Введение в зоологию
Введение в зоологию Декоративные устройства для оформления объектов. Устройство и содержание цветников, вертикальное озеленение, каменистые участки
Декоративные устройства для оформления объектов. Устройство и содержание цветников, вертикальное озеленение, каменистые участки Тип: хордовые. Подтипы: бесчерепные и черепные, позвоночные
Тип: хордовые. Подтипы: бесчерепные и черепные, позвоночные The Human-Animal Bond
The Human-Animal Bond Топ 10 новых пород собак
Топ 10 новых пород собак В гостях у природы
В гостях у природы Парадигмы классической генетики
Парадигмы классической генетики Физиология размножения животных
Физиология размножения животных Биосинтез белка
Биосинтез белка Функции белков
Функции белков Наземно-воздушная среда обитания
Наземно-воздушная среда обитания дикий интерактив конец
дикий интерактив конец Лекарственные растения
Лекарственные растения Комнатные растения
Комнатные растения Строение и работа сердца. Круги кровообращения. Движение крови и лимфы
Строение и работа сердца. Круги кровообращения. Движение крови и лимфы Водоросли. Особенности строения, питания, размножения
Водоросли. Особенности строения, питания, размножения Регуляція експресії генів. (Лекція 2)
Регуляція експресії генів. (Лекція 2) Как появился человек на Земле
Как появился человек на Земле Презентация, конспект урока, карточки - задания к уроку биологии для 6 класса на тему Фотосинтез.
Презентация, конспект урока, карточки - задания к уроку биологии для 6 класса на тему Фотосинтез. Растительный и животный мир различных природных зон
Растительный и животный мир различных природных зон Плесневые грибы и дрожжи
Плесневые грибы и дрожжи Презентация для урока биологии. Мендель. Жизненный путь
Презентация для урока биологии. Мендель. Жизненный путь Еңбектенудің физиологиялық негізі
Еңбектенудің физиологиялық негізі Азотсодержащие гетероциклические соединения. Нуклеиновые кислоты
Азотсодержащие гетероциклические соединения. Нуклеиновые кислоты Органы чувств. Анализаторы
Органы чувств. Анализаторы Хромосомы. Набор хромосом
Хромосомы. Набор хромосом Лишайники 6 класс
Лишайники 6 класс