- Специфика энергетических систем растительной клетки

Содержание
- 2. Самый древний энергетический путь: гликолиз, общая схема У растений гликолиз идет в двух компартментах – в
- 3. Варианты образования глюкозы и фруктозы из сахарозы… Сахарозосинтаза: синтез и распад. Инвертаза – только гидролиз… Ивертаза
- 4. Варианты образования фруктозо-6-фосфата – «стартовой» молекулы гликолиза Пул гексоз в растительной клетке
- 5. Основные стадии гликолиза и их «шунты» в растительной клетке АТФ-зависимая фосфофоруктокиназа Альдолаза Глицеральдегид- 3-Ф-дегидрогеназа Фософоглицераткиназа Пируваткиназа
- 6. Два пути образования фруктозо-1,6-бисфосфата у растений Два изозима – цитозольный и пластидный
- 7. Еще один сюрприз растительной клетки… Одна из догм биохимии: дифосфат (пирофосфат) не накапливается в клетке, поскольку
- 8. Положение второго фосфата кардинально изменяет функции молекулы
- 9. Дифосфат-зивисимая фосфофруктокиназа. Только у растений… АТФ-зависимая фосфофоруктокиназа
- 10. Образование триоз. В целом - типично…
- 11. Основные реакции гликолиза – окисление глицеральдегид-фосфата
- 12. Основные стадии гликолиза и их «шунты» в растительной клетке АТФ-зависимая фосфофоруктокиназа Альдолаза Глицеральдегид- 3-Ф-дегидрогеназа Фософоглицераткиназа Пируваткиназа
- 13. Гликолиз в растительной клетке можно повернуть вспять. Или - как из жира сделать сахар… Глюконеогенез и
- 14. Глиоксилатный цикл чуть подробнее…
- 15. Регулирование гликолиза у животных и растений АДФ АТФ Р Помним, что у растений, помимо прочего, еще
- 16. Общая схема энергетической системы клетки - дыхание Особенности ЦТК у растений
- 17. Общая схема ЦТК (цикла Кребса)
- 18. Окисление пирувата – пируватдегидрогеназный комплекс E0’ пируват/ацетат + СО2 = - 0,70v. Надо раскулачивать… ΔЕ ~
- 19. Еще несколько важных метаболических «шунтов» в дыхании растений: конечный продукт гликолиза не только пируват, но и
- 20. Транспорт веществ через мембрану митохондрии
- 21. Цикл Кребса в растительных митохондриях Особенности ЦТК растений 1. Два «входа» в цикл – пируват и
- 22. «Синтетические» функции митохондрий растительной клетки.
- 23. Общая схема энергетической системы клетки Особенности растительной ЭТЦ : шунты, регулирование
- 24. Особенности ЭТЦ дыхания растений: дополнительные НАД(Ф)Н-дегидрогеназы И зачем они нужны? Шунты? Для одной из них –
- 25. Цианидрезистентное дыхание Термогенез ароидных растений Термогенез в цветках некоторых растений (Arum maculatum, Symplocarpus foetidis, Sauromatum guttatum,
- 26. Sauromatum guttatum
- 27. Symplocarpus foetidus
- 28. Helicodeceros muscivorus
- 29. Особенности ЭТЦ-дыхания растений: альтернативная оксидаза (цианидрезистентное дыхание) Гомодимер, 37 kDa, ~ 350 а-к. Передает е- от
- 30. Зачем нужна альтернативная оксидаза и как она включается? Гипотезы: 1. Для шунтирования ЭТЦ – модели «перелива»
- 31. Регулирование синтеза и активности альтернативной оксидазы
- 32. Итак, ЭТЦ дыхания растений Похоже, у растений обнаружили еще и разобщающий белок, как и у животных…
- 33. Транспорт интермедиатов дыхания через мембрану митохондрий. Трансгидрогеназа НАДН → НАД+ НАДФН → НАДФ+ В растениях трансгидрогеназа
- 34. Митохондрии растений играют важную роль во многих процессах и в кооперации с разыми органеллами – фотодыхание,
- 35. Челночные механизмы транспорта для обмена метаболитами и восстановленными эквивалентами В растительной клетке эти механизмы работают и
- 36. И еще раз - общая схема энергетической системы клетки Особенности дыхания растений: «глобальные» шунты (альтернативные пути),
- 37. Альтернативные пути дыхания растений Пул гексоз
- 38. Окислительная стадия пентозофосфатного цикла А где же цикл? Как и гликолиз, окислительный ПФШ у растений идет
- 39. Окислительный пентозофосфатный цикл (шунт) - ПФШ С6Н12О6 + 12НАДФ+ + 6Н2О → 6СО2 + 12НАДФН +
- 40. Фаза регенерации пентозофосфатного цикла
- 41. «Начало» гликолиза – составная часть пентозофосфатного цикла В норме на долю гликолиза приходится до 70 -90%
- 42. NB - гликолиз и ПФШ имеют общие пулы гексоз и триоз. пируват, малат пируват, малат Получается,
- 43. Альтернативные пути дыхания растений Пул гексоз Г-3Ф
- 44. Цитоплазматические электрон-транспортные цепи растений
- 45. Тиоредоксин и глутатион – компоненты многих регуляторных Red-Ox реакций Глутатион: трипептид: L-γ-глутамил-L-цистенил-глицин. 2GSH ↔ GS–SG +
- 46. Аскорбатоксидаза Медь- содержащий фермент. 8 атомов Cu – 6Cu2+, 2Cu+. М.в. 140-170 kDa Работает в паре
- 47. Дифенолоксидазы (полифенолоксидазы) о-дифенолоксидаза: Медь-содержащий фермент, М.в. 32 kDa. Крезолазная и катехолазная активность п-дифенолоксидаза: Медь-содержащий фермент, 4
- 48. Структура полифенолоксидазы и механизм реакции
- 49. Оксигеназы растений Пример: Липоксигеназы: Мономерные белки 94 – 97 kDa, Много изоформ: цитозоль (семена) хлоропласты (листья)
- 50. Структура липоксигеназы соевых бобов Мономерные белки 94 – 97 kDa Небольшой N-домен (фиолетовый) и значительный С-домен
- 51. Пероксидаза и каталаза Пероксидазы: гем-содержащие белки, М.в. ~44 kDa. Окисляют субстраты за счет перекиси водорода. Е-Fe-OH
- 52. Пероксидаза и ее активный центр Пероксидазы окисляют субстраты за счет перекиси водорода. У них обязателено присутсвует
- 53. Структура каталазы и ее защитные функции Каталаза – тетрамер, 60 или 75 kDa, каждая из субъединиц
- 54. Оксидазы митохондрий и ЭР Альтернативная оксидаза Цит а + а3 Цит Р-450 х
- 55. Оксидазы цитозоля Аскорбат оксидаза Полифенол оксидаза Флавопротеин-оксидаза
- 57. Скачать презентацию
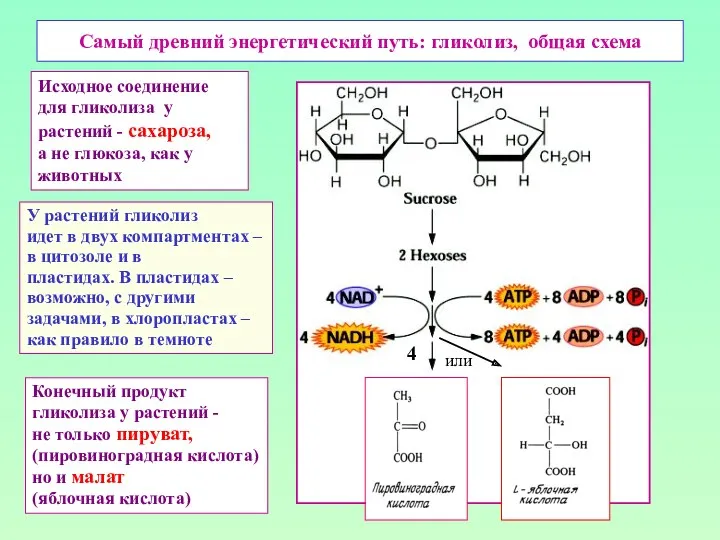
Самый древний энергетический путь: гликолиз, общая схема
У растений гликолиз
идет в двух
Самый древний энергетический путь: гликолиз, общая схема
У растений гликолиз
идет в двух
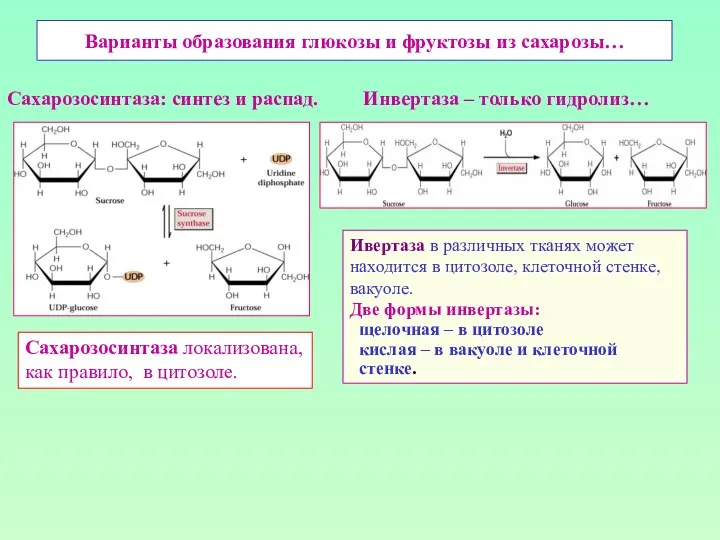
Варианты образования глюкозы и фруктозы из сахарозы…
Сахарозосинтаза: синтез и распад. Инвертаза
Варианты образования глюкозы и фруктозы из сахарозы…
Сахарозосинтаза: синтез и распад. Инвертаза
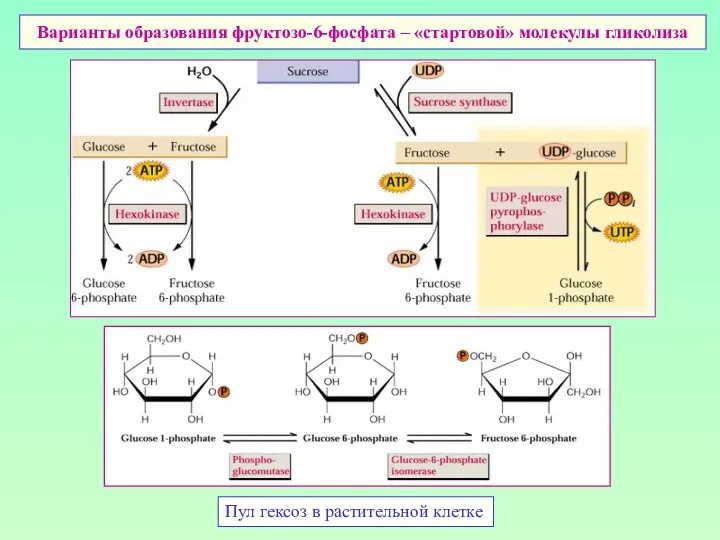
Варианты образования фруктозо-6-фосфата – «стартовой» молекулы гликолиза
Пул гексоз в растительной клетке
Варианты образования фруктозо-6-фосфата – «стартовой» молекулы гликолиза
Пул гексоз в растительной клетке
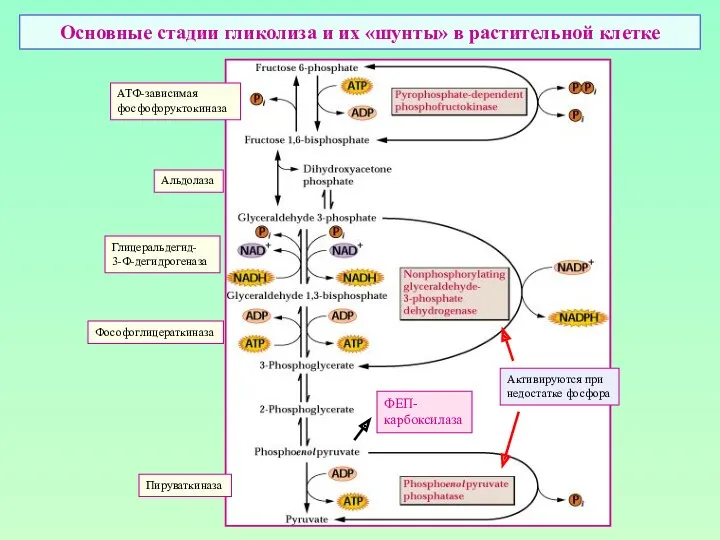
Основные стадии гликолиза и их «шунты» в растительной клетке
АТФ-зависимая
фосфофоруктокиназа
Альдолаза
Глицеральдегид-
3-Ф-дегидрогеназа
Фософоглицераткиназа
Пируваткиназа
Активируются при
недостатке фосфора
ФЕП-
карбоксилаза
Основные стадии гликолиза и их «шунты» в растительной клетке
АТФ-зависимая
фосфофоруктокиназа
Альдолаза
Глицеральдегид-
3-Ф-дегидрогеназа
Фософоглицераткиназа
Пируваткиназа
Активируются при
недостатке фосфора
ФЕП-
карбоксилаза
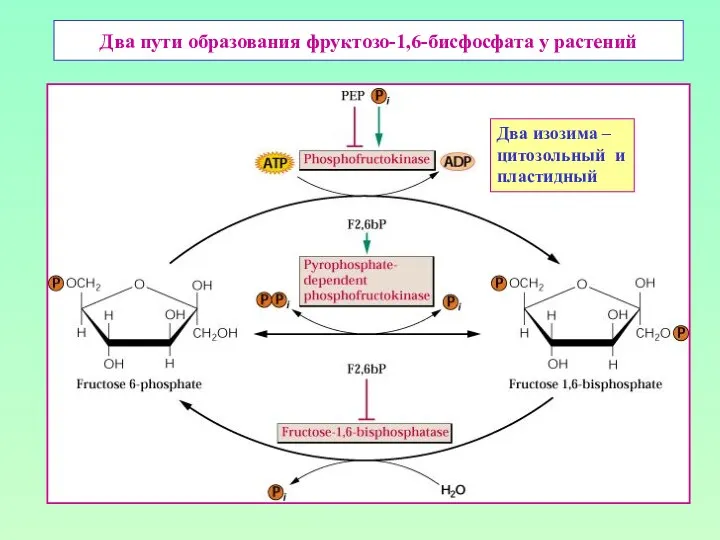
Два пути образования фруктозо-1,6-бисфосфата у растений
Два изозима –
цитозольный и
пластидный
Два пути образования фруктозо-1,6-бисфосфата у растений
Два изозима –
цитозольный и
пластидный
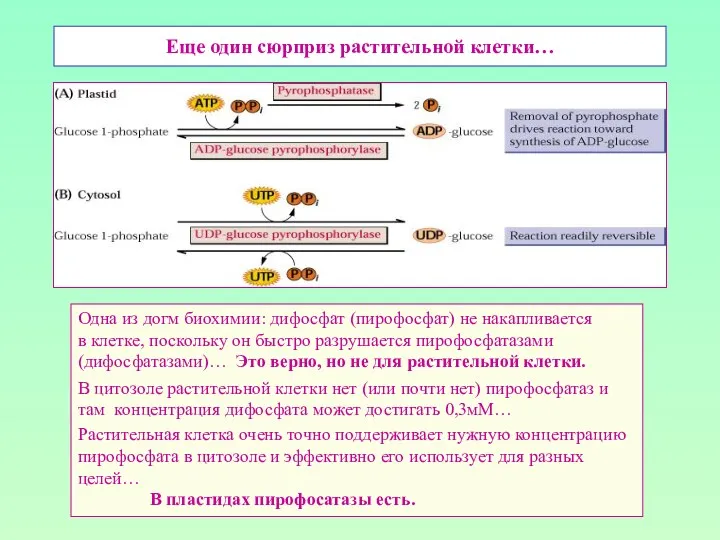
Еще один сюрприз растительной клетки…
Одна из догм биохимии: дифосфат (пирофосфат) не
Еще один сюрприз растительной клетки…
Одна из догм биохимии: дифосфат (пирофосфат) не
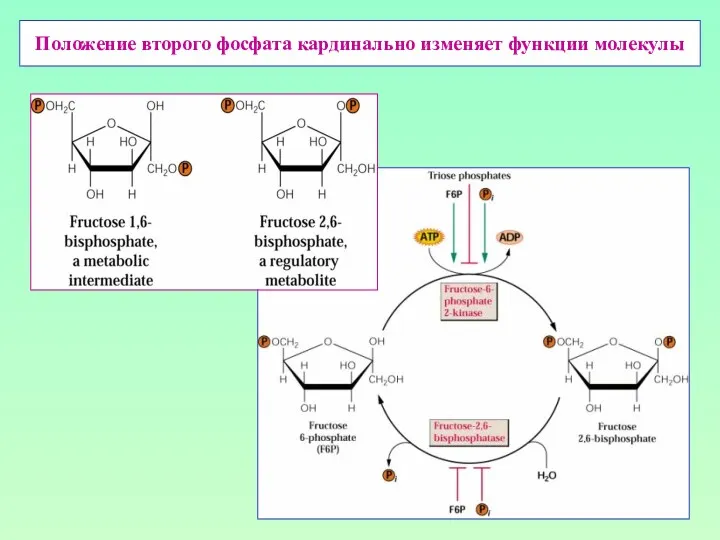
Положение второго фосфата кардинально изменяет функции молекулы
Положение второго фосфата кардинально изменяет функции молекулы
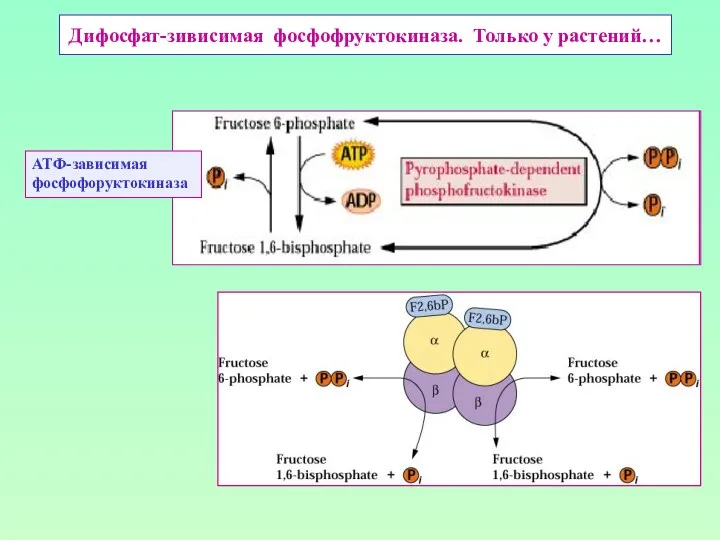
Дифосфат-зивисимая фосфофруктокиназа. Только у растений…
АТФ-зависимая
фосфофоруктокиназа
Дифосфат-зивисимая фосфофруктокиназа. Только у растений…
АТФ-зависимая
фосфофоруктокиназа
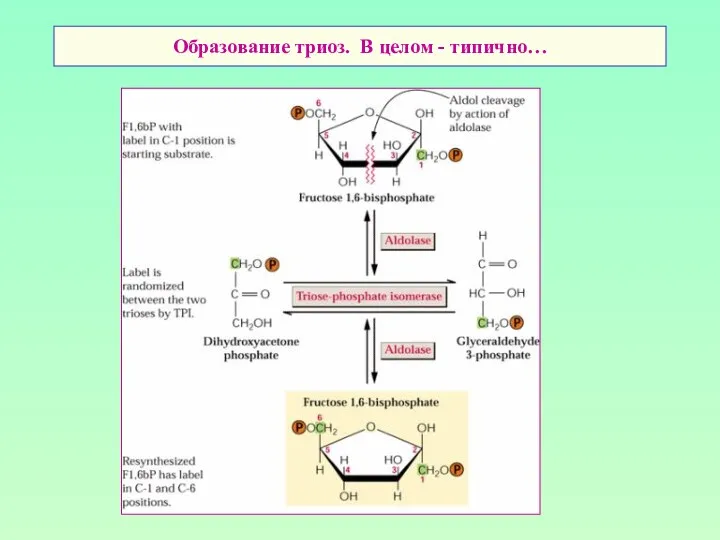
Образование триоз. В целом - типично…
Образование триоз. В целом - типично…
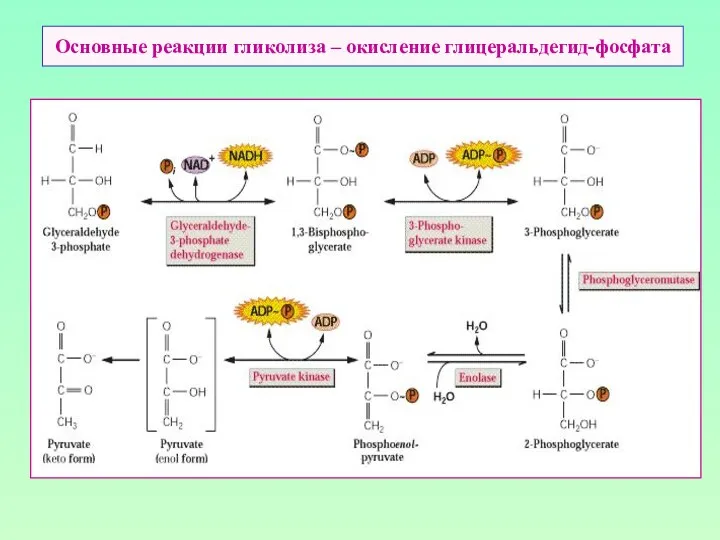
Основные реакции гликолиза – окисление глицеральдегид-фосфата
Основные реакции гликолиза – окисление глицеральдегид-фосфата
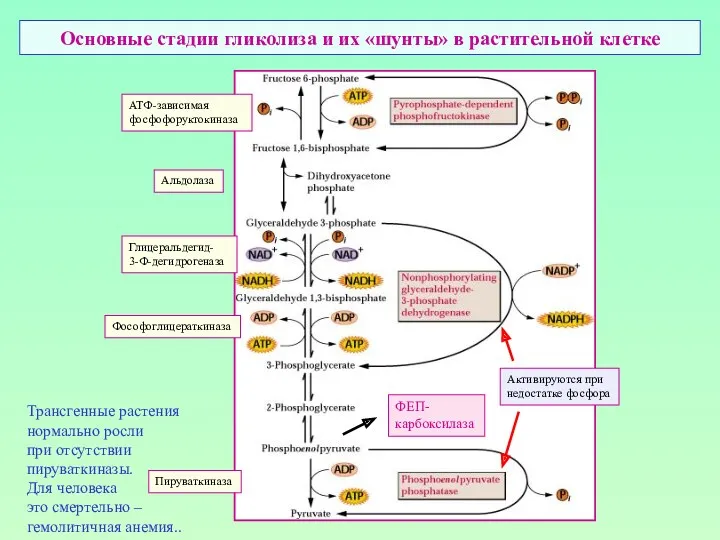
Основные стадии гликолиза и их «шунты» в растительной клетке
АТФ-зависимая
фосфофоруктокиназа
Альдолаза
Глицеральдегид-
3-Ф-дегидрогеназа
Фософоглицераткиназа
Пируваткиназа
Активируются при
недостатке фосфора
Трансгенные
Основные стадии гликолиза и их «шунты» в растительной клетке
АТФ-зависимая
фосфофоруктокиназа
Альдолаза
Глицеральдегид-
3-Ф-дегидрогеназа
Фософоглицераткиназа
Пируваткиназа
Активируются при
недостатке фосфора
Трансгенные
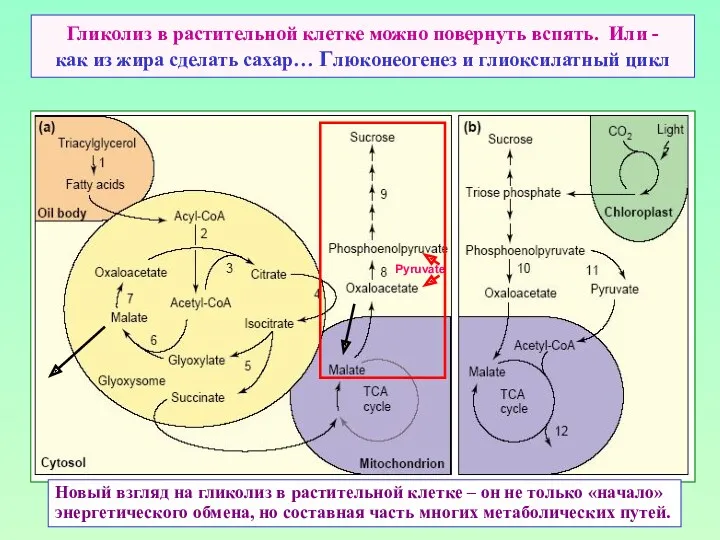
Гликолиз в растительной клетке можно повернуть вспять. Или -
как из жира
Гликолиз в растительной клетке можно повернуть вспять. Или - как из жира
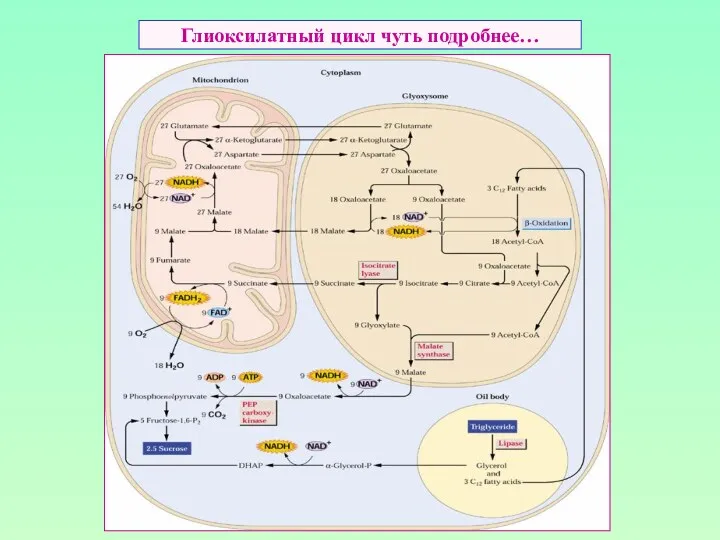
Глиоксилатный цикл чуть подробнее…
Глиоксилатный цикл чуть подробнее…
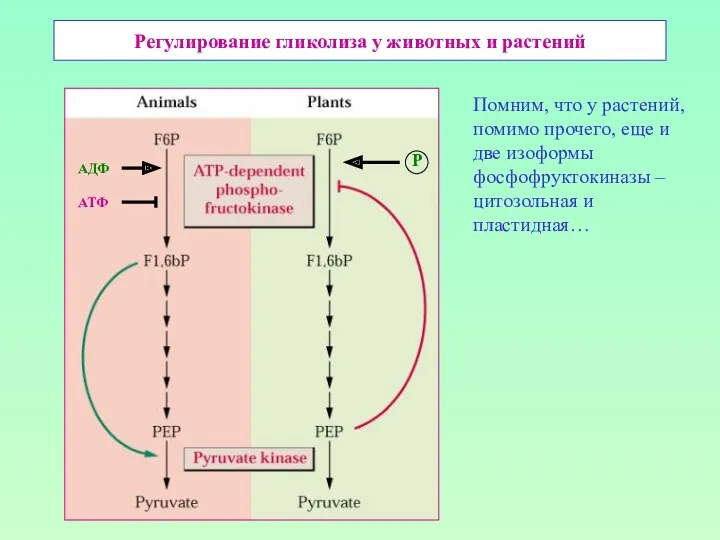
Регулирование гликолиза у животных и растений
АДФ
АТФ
Р
Помним, что у растений, помимо прочего,
Регулирование гликолиза у животных и растений
АДФ
АТФ
Р
Помним, что у растений, помимо прочего,
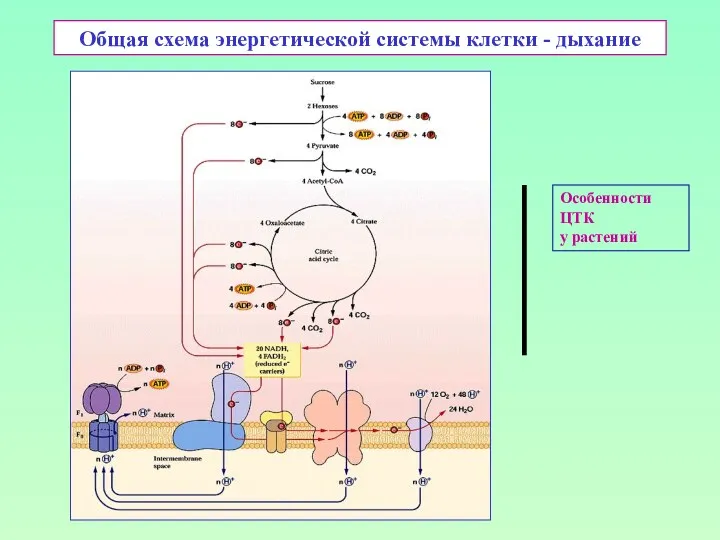
Общая схема энергетической системы клетки - дыхание
Особенности
ЦТК
у растений
Общая схема энергетической системы клетки - дыхание
Особенности
ЦТК
у растений
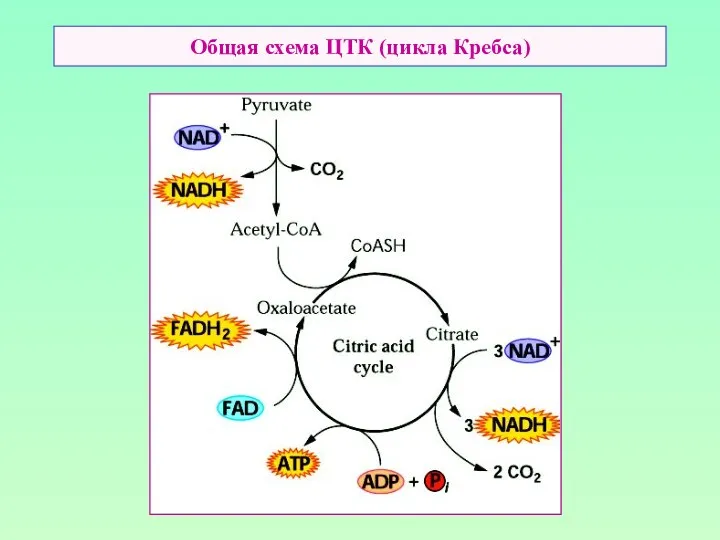
Общая схема ЦТК (цикла Кребса)
Общая схема ЦТК (цикла Кребса)
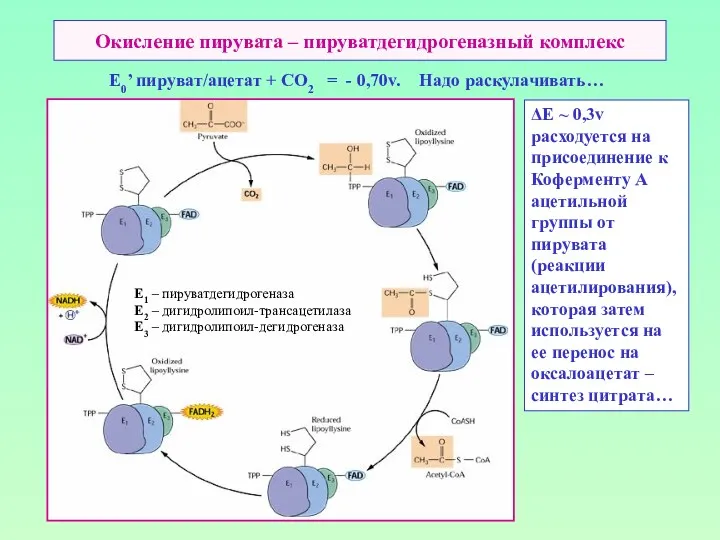
Окисление пирувата – пируватдегидрогеназный комплекс
E0’ пируват/ацетат + СО2 = -
Окисление пирувата – пируватдегидрогеназный комплекс
E0’ пируват/ацетат + СО2 = -
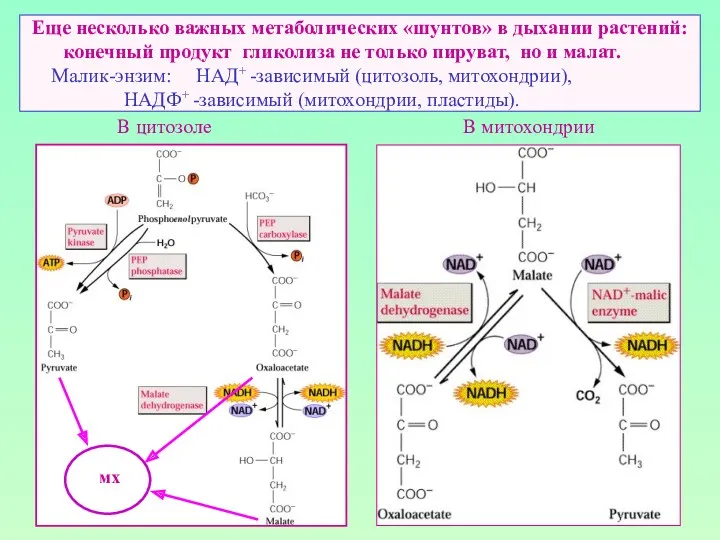
Еще несколько важных метаболических «шунтов» в дыхании растений: конечный продукт
Еще несколько важных метаболических «шунтов» в дыхании растений: конечный продукт
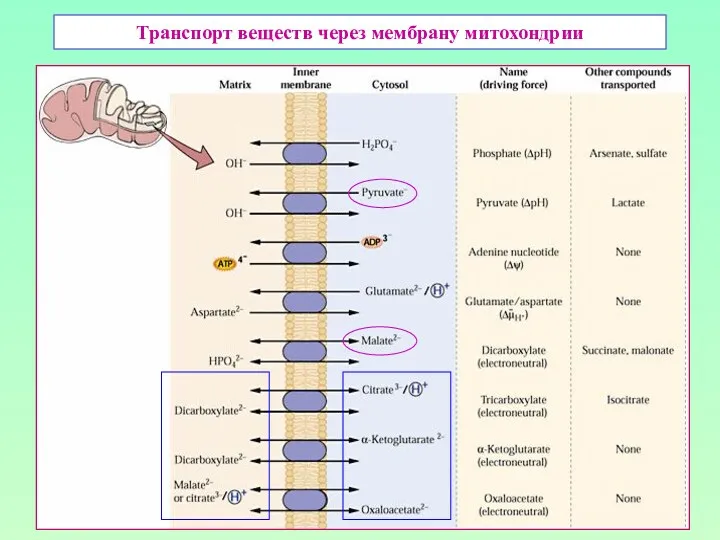
Транспорт веществ через мембрану митохондрии
Транспорт веществ через мембрану митохондрии
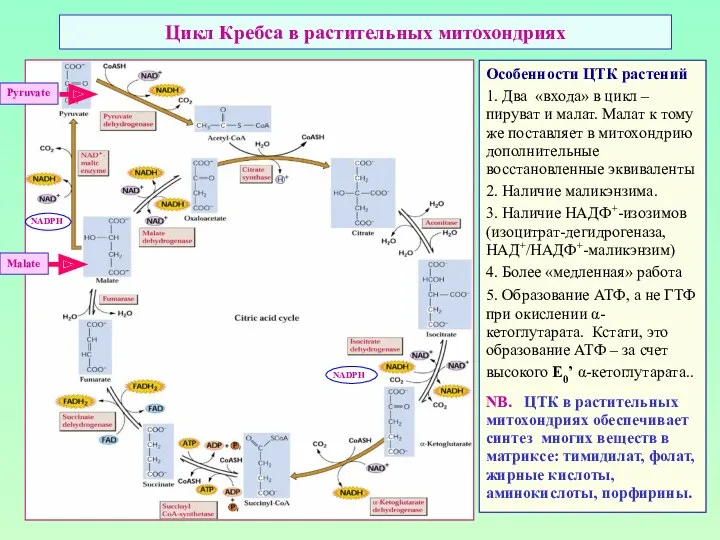
Цикл Кребса в растительных митохондриях
Особенности ЦТК растений
1. Два «входа» в цикл
Цикл Кребса в растительных митохондриях
Особенности ЦТК растений
1. Два «входа» в цикл
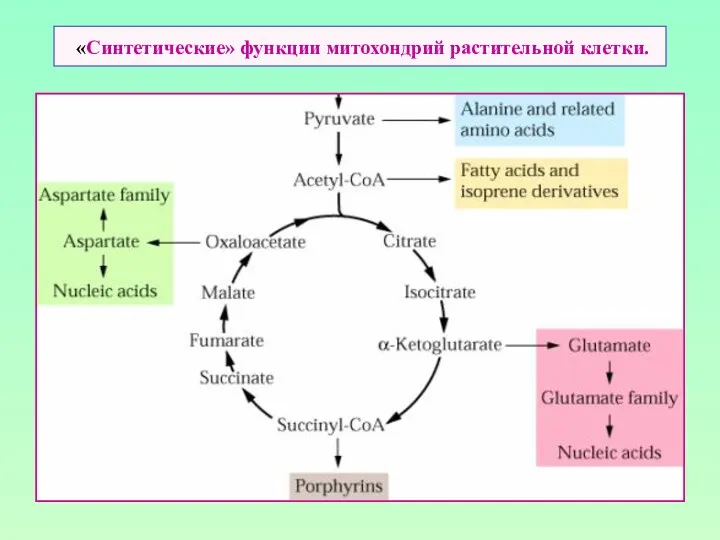
«Синтетические» функции митохондрий растительной клетки.
«Синтетические» функции митохондрий растительной клетки.
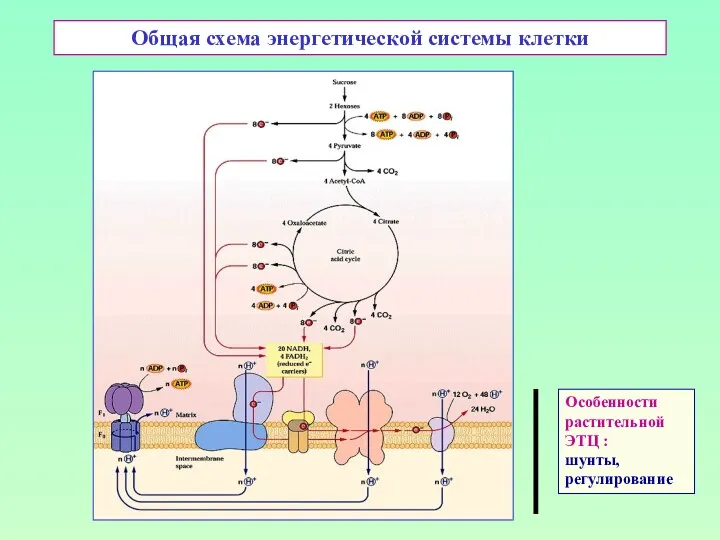
Общая схема энергетической системы клетки
Особенности
растительной
ЭТЦ :
шунты,
регулирование
Общая схема энергетической системы клетки
Особенности
растительной
ЭТЦ :
шунты,
регулирование
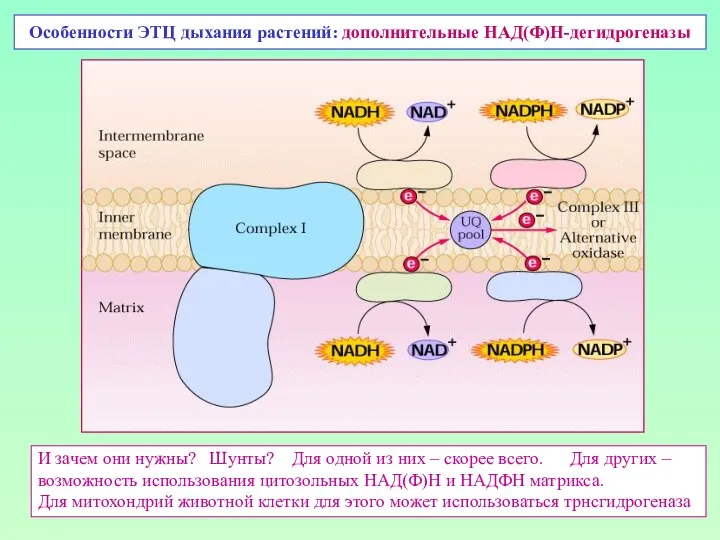
Особенности ЭТЦ дыхания растений: дополнительные НАД(Ф)Н-дегидрогеназы
И зачем они нужны? Шунты? Для
Особенности ЭТЦ дыхания растений: дополнительные НАД(Ф)Н-дегидрогеназы
И зачем они нужны? Шунты? Для
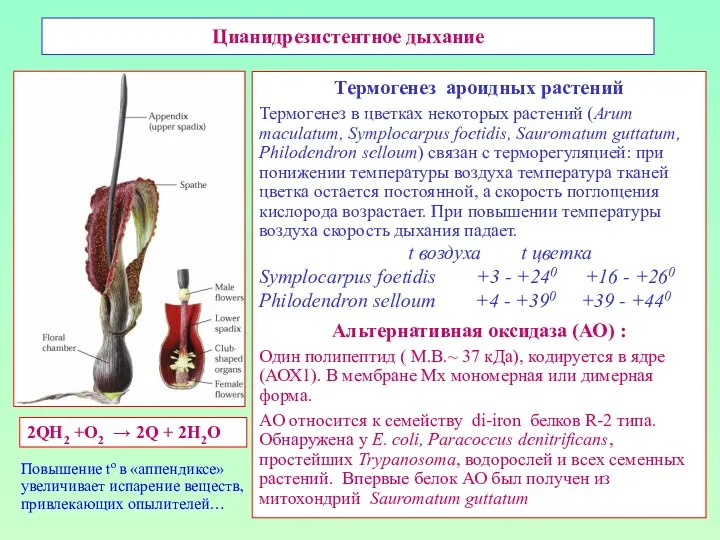
Цианидрезистентное дыхание
Термогенез ароидных растений
Термогенез в цветках некоторых растений (Arum maculatum,
Цианидрезистентное дыхание
Термогенез ароидных растений
Термогенез в цветках некоторых растений (Arum maculatum,

Sauromatum guttatum
Sauromatum guttatum

Symplocarpus foetidus
Symplocarpus foetidus

Helicodeceros muscivorus
Helicodeceros muscivorus
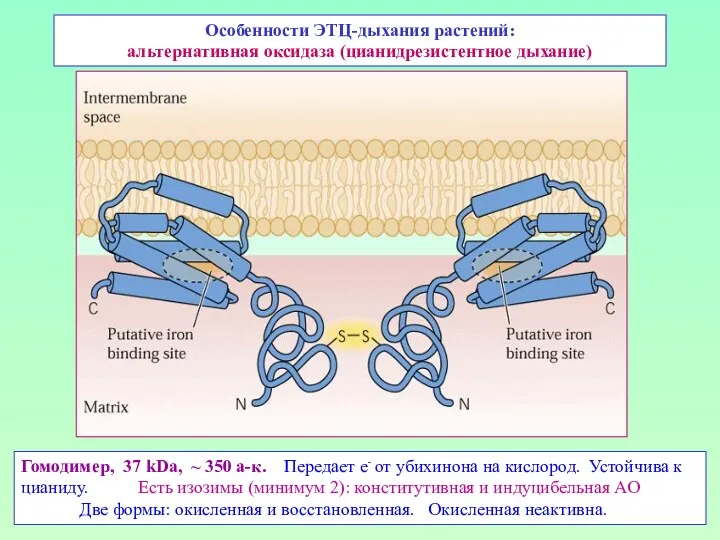
Особенности ЭТЦ-дыхания растений:
альтернативная оксидаза (цианидрезистентное дыхание)
Гомодимер, 37 kDa, ~ 350 а-к.
Особенности ЭТЦ-дыхания растений:
альтернативная оксидаза (цианидрезистентное дыхание)
Гомодимер, 37 kDa, ~ 350 а-к.
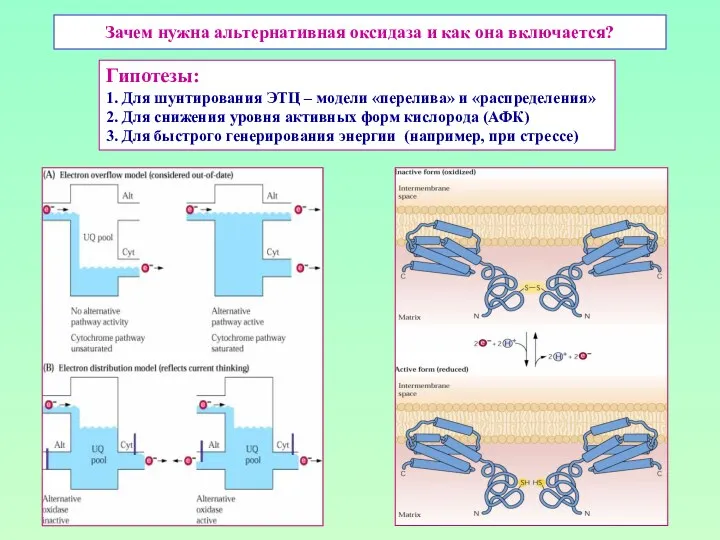
Зачем нужна альтернативная оксидаза и как она включается?
Гипотезы:
1. Для шунтирования ЭТЦ
Зачем нужна альтернативная оксидаза и как она включается?
Гипотезы:
1. Для шунтирования ЭТЦ
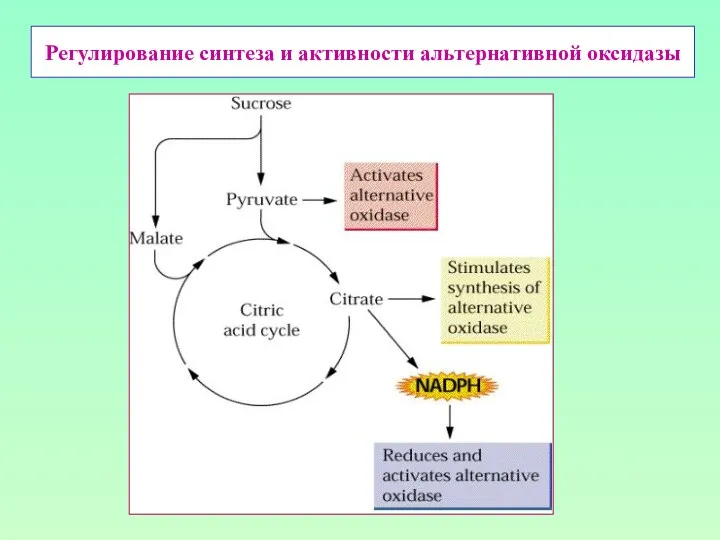
Регулирование синтеза и активности альтернативной оксидазы
Регулирование синтеза и активности альтернативной оксидазы
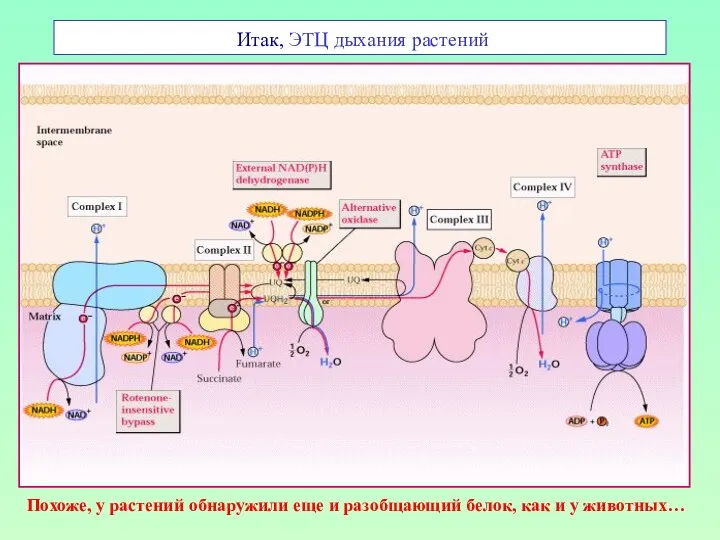
Итак, ЭТЦ дыхания растений
Похоже, у растений обнаружили еще и
Итак, ЭТЦ дыхания растений
Похоже, у растений обнаружили еще и
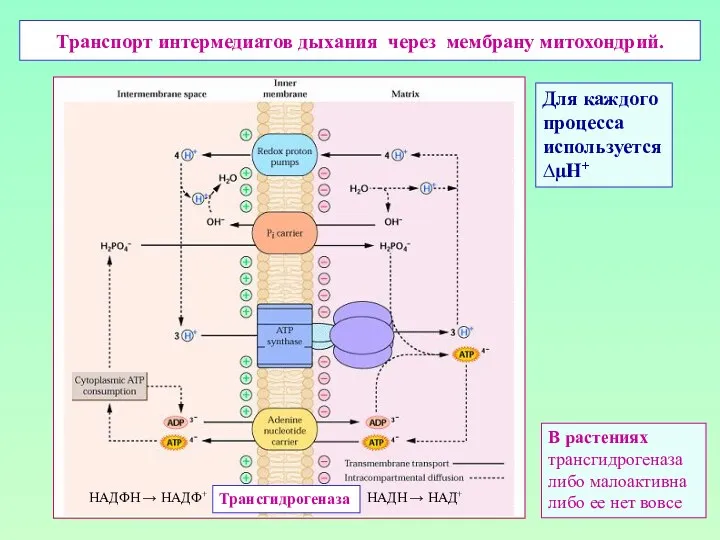
Транспорт интермедиатов дыхания через мембрану митохондрий.
Трансгидрогеназа
НАДН → НАД+
НАДФН → НАДФ+
В растениях
трансгидрогеназа
либо
Транспорт интермедиатов дыхания через мембрану митохондрий.
Трансгидрогеназа
НАДН → НАД+
НАДФН → НАДФ+
В растениях
трансгидрогеназа
либо
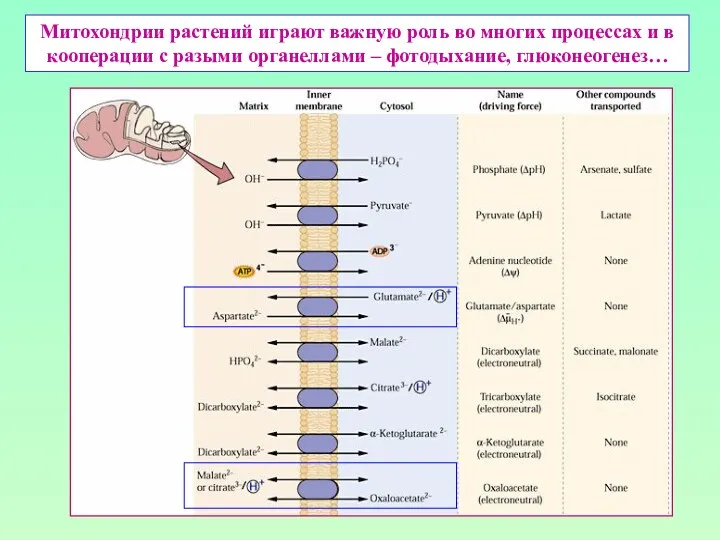
Митохондрии растений играют важную роль во многих процессах и в кооперации
Митохондрии растений играют важную роль во многих процессах и в кооперации
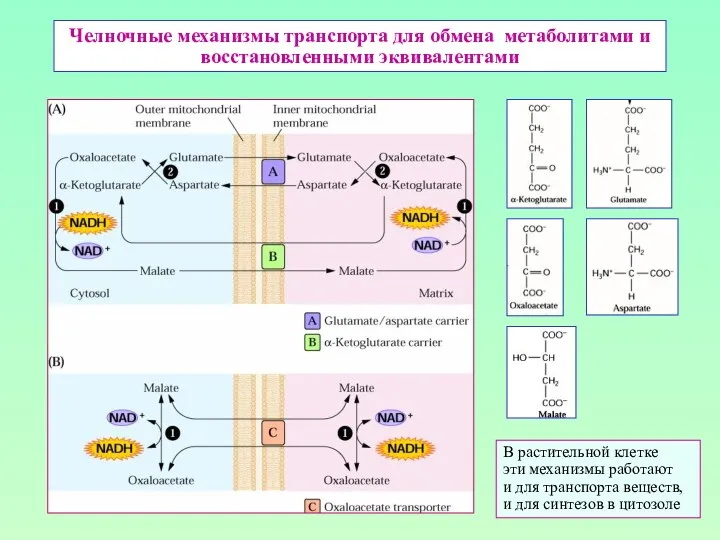
Челночные механизмы транспорта для обмена метаболитами и восстановленными эквивалентами
В растительной клетке
эти
Челночные механизмы транспорта для обмена метаболитами и восстановленными эквивалентами
В растительной клетке
эти
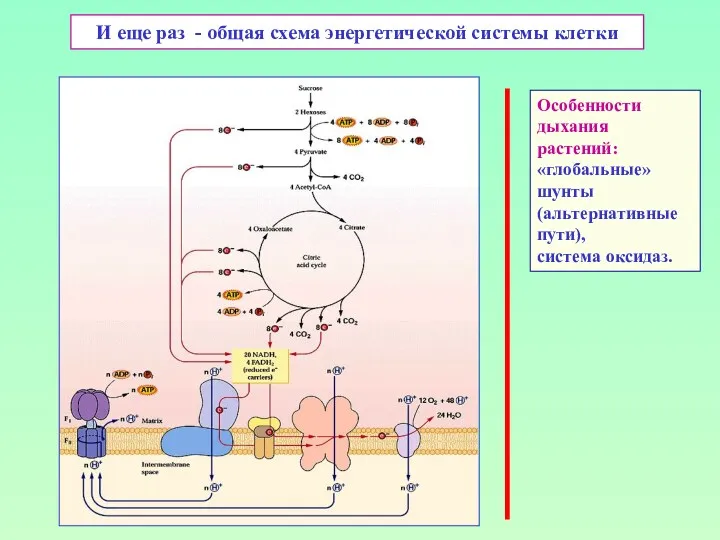
И еще раз - общая схема энергетической системы клетки
Особенности
дыхания растений:
«глобальные»
шунты
(альтернативные
пути),
система
И еще раз - общая схема энергетической системы клетки
Особенности
дыхания растений:
«глобальные»
шунты
(альтернативные
пути),
система
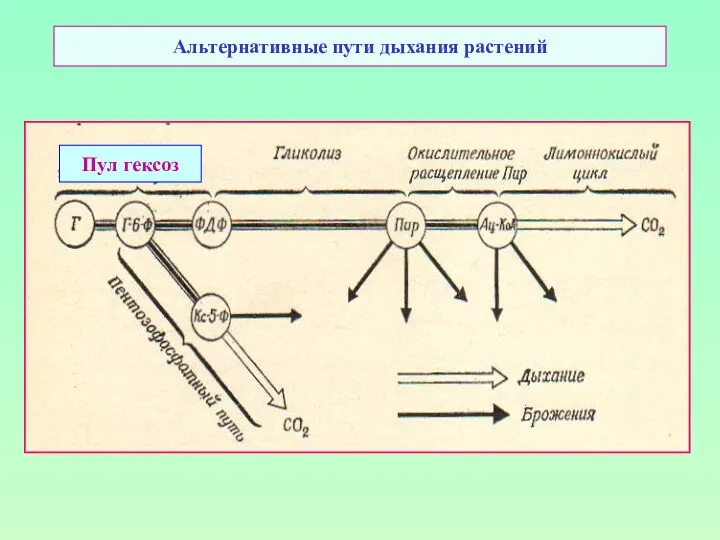
Альтернативные пути дыхания растений
Пул гексоз
Альтернативные пути дыхания растений
Пул гексоз
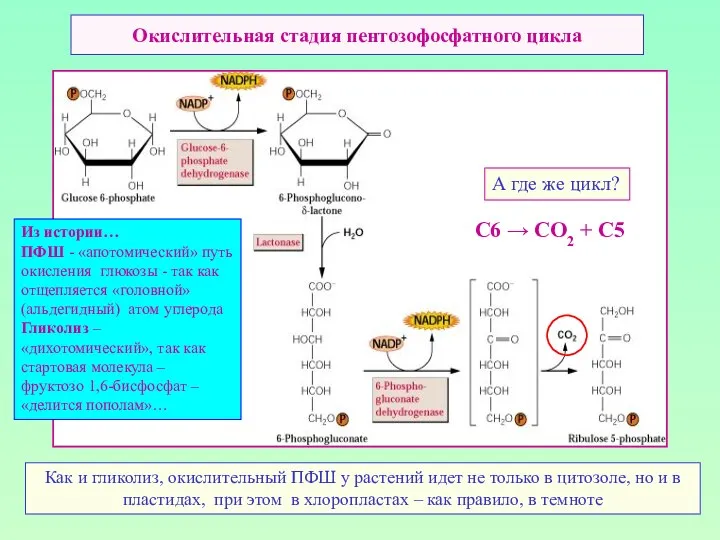
Окислительная стадия пентозофосфатного цикла
А где же цикл?
Как и гликолиз, окислительный ПФШ
Окислительная стадия пентозофосфатного цикла
А где же цикл?
Как и гликолиз, окислительный ПФШ
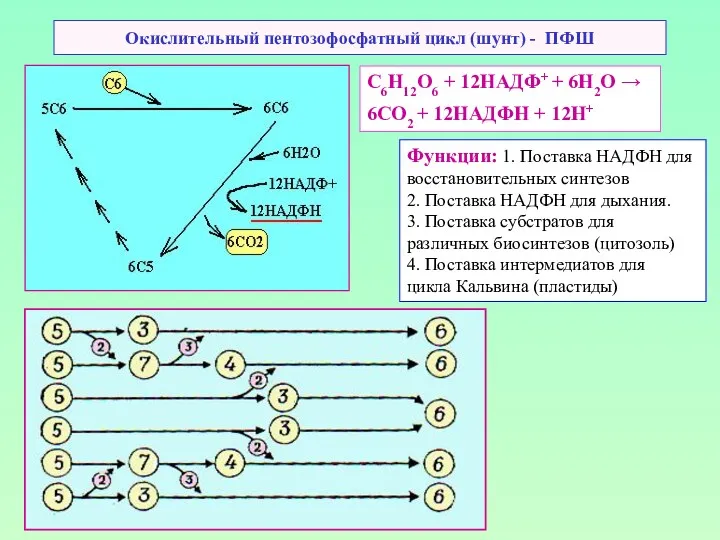
Окислительный пентозофосфатный цикл (шунт) - ПФШ
С6Н12О6 + 12НАДФ+ + 6Н2О
Окислительный пентозофосфатный цикл (шунт) - ПФШ
С6Н12О6 + 12НАДФ+ + 6Н2О
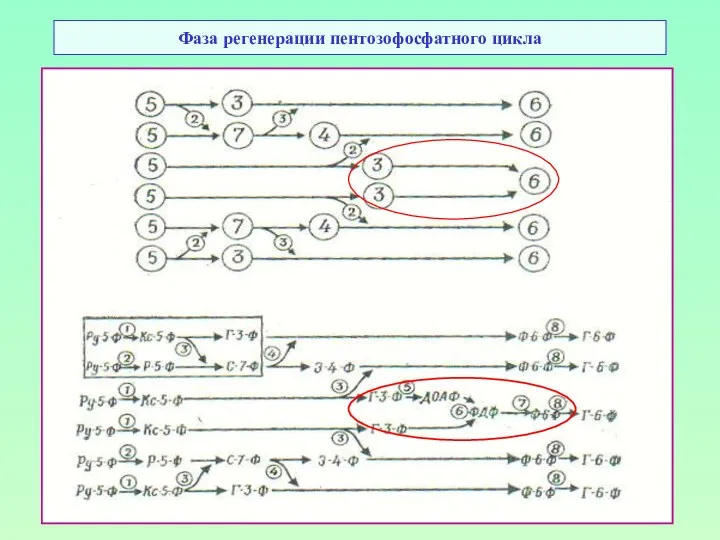
Фаза регенерации пентозофосфатного цикла
Фаза регенерации пентозофосфатного цикла
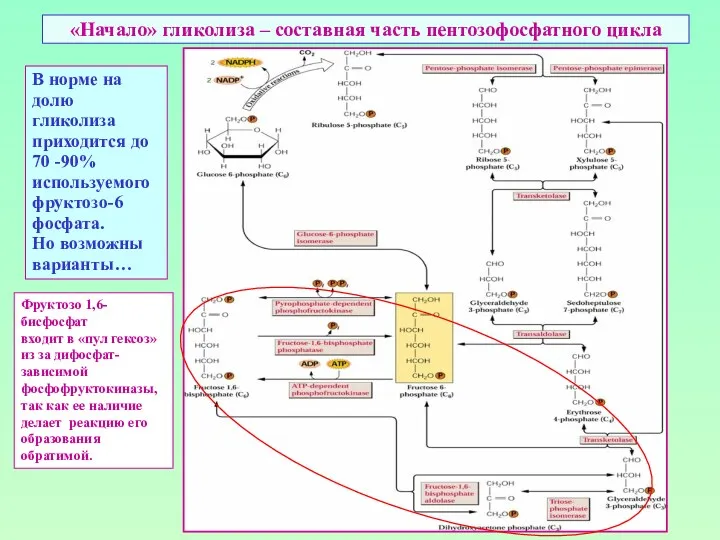
«Начало» гликолиза – составная часть пентозофосфатного цикла
В норме на долю
гликолиза
«Начало» гликолиза – составная часть пентозофосфатного цикла
В норме на долю
гликолиза
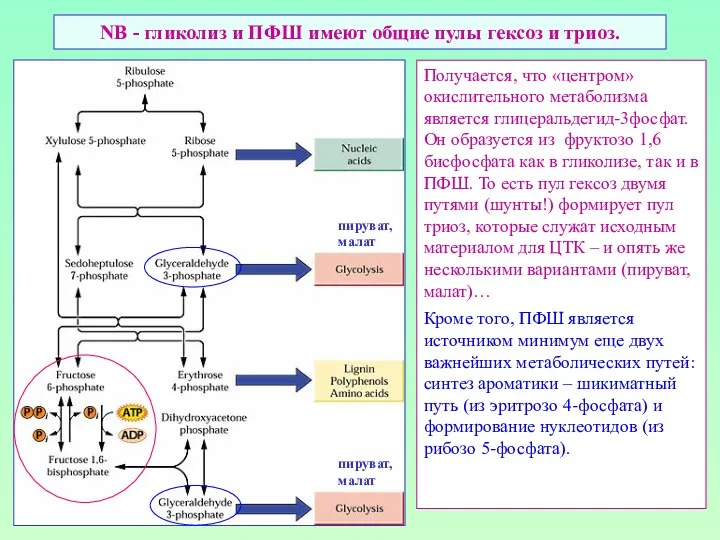
NB - гликолиз и ПФШ имеют общие пулы гексоз и триоз.
NB - гликолиз и ПФШ имеют общие пулы гексоз и триоз.
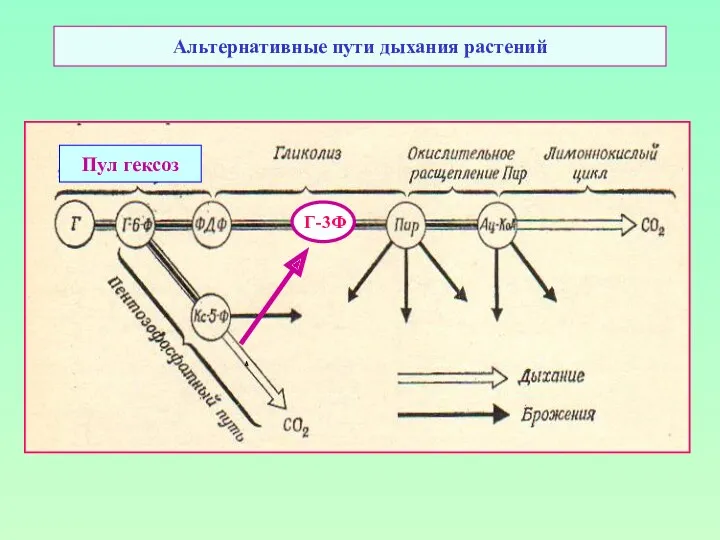
Альтернативные пути дыхания растений
Пул гексоз
Г-3Ф
Альтернативные пути дыхания растений
Пул гексоз
Г-3Ф

Цитоплазматические электрон-транспортные цепи растений
Цитоплазматические электрон-транспортные цепи растений

Тиоредоксин и глутатион – компоненты многих регуляторных Red-Ox реакций
Глутатион: трипептид:
L-γ-глутамил-L-цистенил-глицин.
2GSH ↔
Тиоредоксин и глутатион – компоненты многих регуляторных Red-Ox реакций
Глутатион: трипептид:
L-γ-глутамил-L-цистенил-глицин.
2GSH ↔
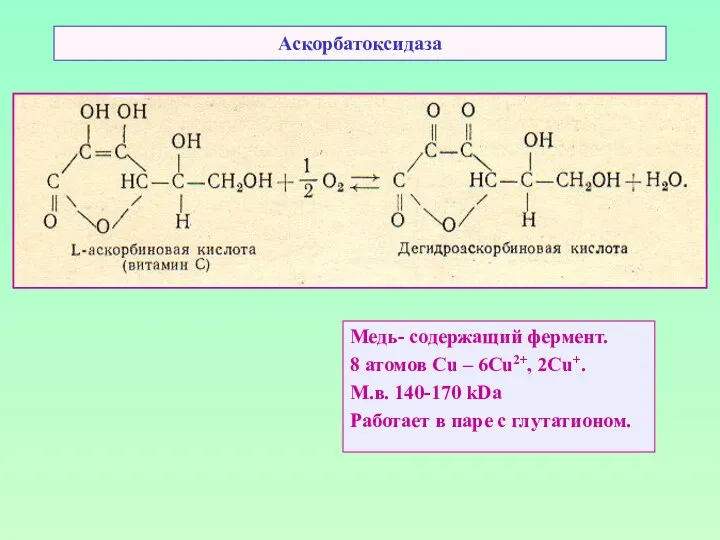
Аскорбатоксидаза
Медь- содержащий фермент.
8 атомов Cu – 6Cu2+, 2Cu+.
М.в. 140-170 kDa
Работает в
Аскорбатоксидаза
Медь- содержащий фермент.
8 атомов Cu – 6Cu2+, 2Cu+.
М.в. 140-170 kDa
Работает в
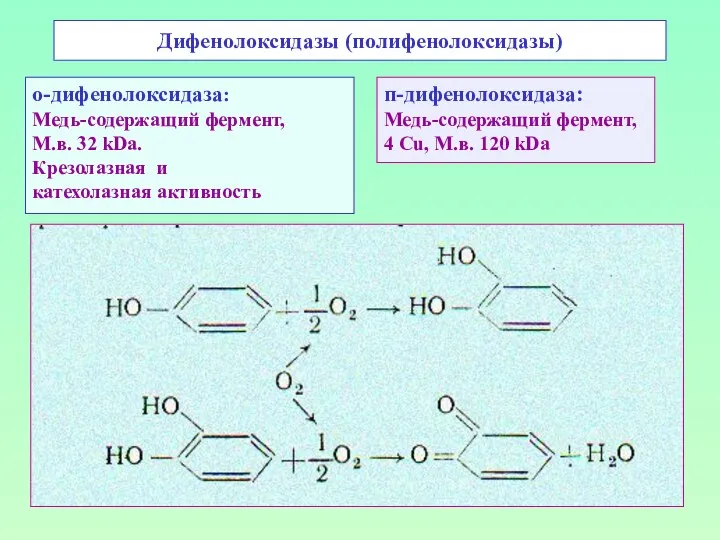
Дифенолоксидазы (полифенолоксидазы)
о-дифенолоксидаза:
Медь-содержащий фермент,
М.в. 32 kDa.
Крезолазная и
катехолазная активность
п-дифенолоксидаза:
Медь-содержащий фермент,
4 Cu, М.в.
Дифенолоксидазы (полифенолоксидазы)
о-дифенолоксидаза:
Медь-содержащий фермент,
М.в. 32 kDa.
Крезолазная и
катехолазная активность
п-дифенолоксидаза:
Медь-содержащий фермент,
4 Cu, М.в.
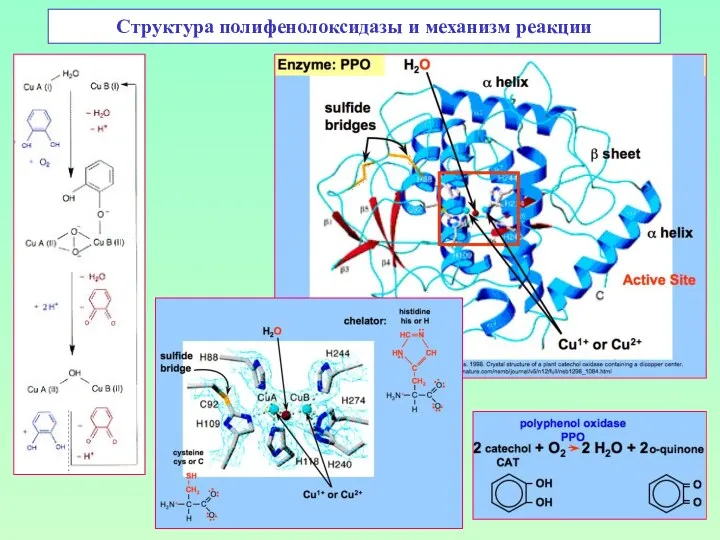
Структура полифенолоксидазы и механизм реакции
Структура полифенолоксидазы и механизм реакции

Оксигеназы растений
Пример:
Липоксигеназы:
Мономерные белки
94 – 97 kDa,
Много изоформ:
цитозоль (семена)
хлоропласты (листья)
вакуоль (корни)
Оксигеназы растений
Пример:
Липоксигеназы:
Мономерные белки
94 – 97 kDa,
Много изоформ:
цитозоль (семена)
хлоропласты (листья)
вакуоль (корни)
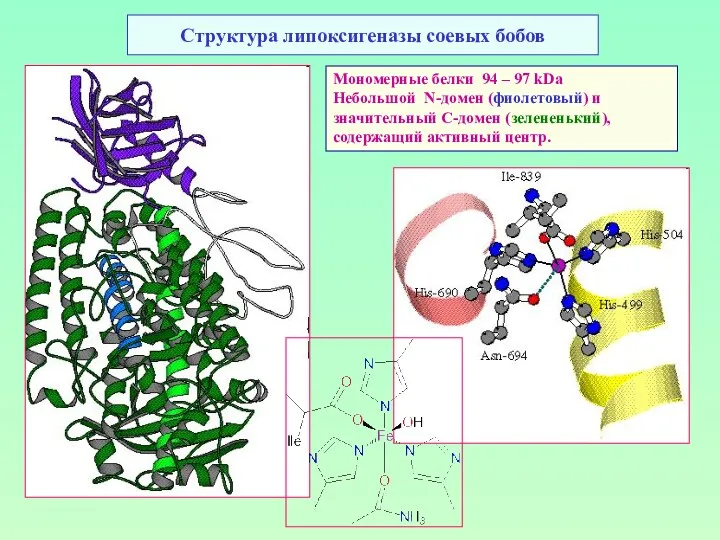
Структура липоксигеназы соевых бобов
Мономерные белки 94 – 97 kDa
Небольшой N-домен
Структура липоксигеназы соевых бобов
Мономерные белки 94 – 97 kDa
Небольшой N-домен

Пероксидаза и каталаза
Пероксидазы: гем-содержащие белки, М.в. ~44 kDa.
Окисляют субстраты за
Пероксидаза и каталаза
Пероксидазы: гем-содержащие белки, М.в. ~44 kDa.
Окисляют субстраты за

Пероксидаза и ее активный центр
Пероксидазы окисляют субстраты за счет перекиси водорода.
Пероксидаза и ее активный центр
Пероксидазы окисляют субстраты за счет перекиси водорода.
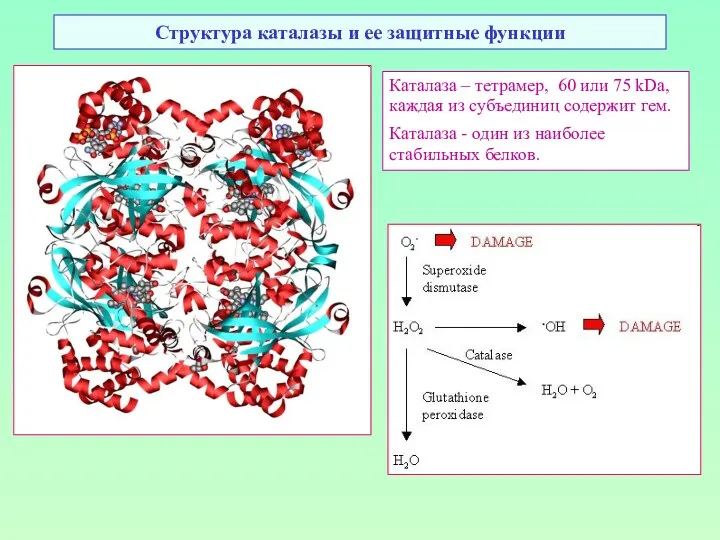
Структура каталазы и ее защитные функции
Каталаза – тетрамер, 60 или 75
Структура каталазы и ее защитные функции
Каталаза – тетрамер, 60 или 75
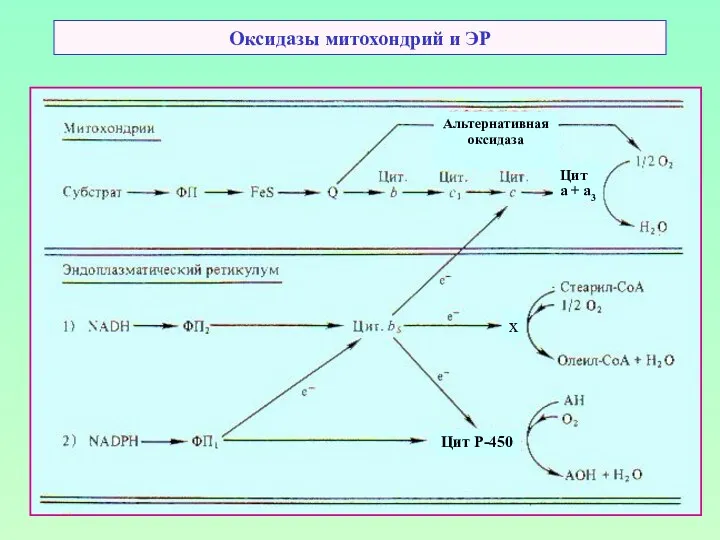
Оксидазы митохондрий и ЭР
Альтернативная
оксидаза
Цит
а + а3
Цит Р-450
х
Оксидазы митохондрий и ЭР
Альтернативная
оксидаза
Цит
а + а3
Цит Р-450
х
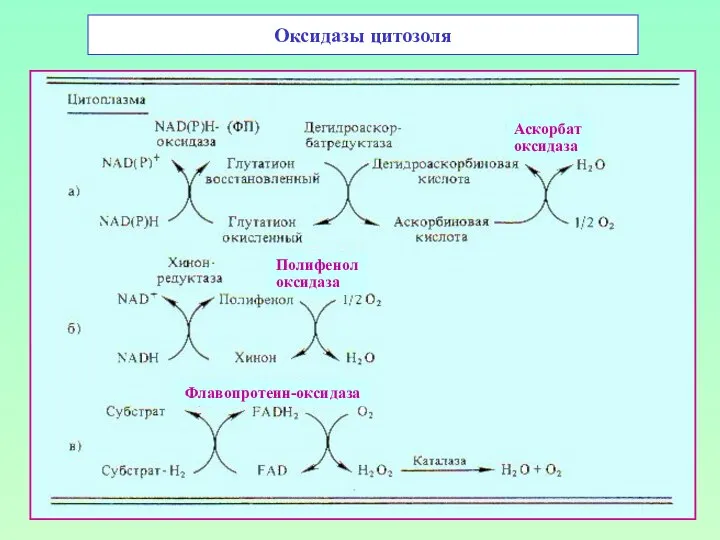
Оксидазы цитозоля
Аскорбат
оксидаза
Полифенол
оксидаза
Флавопротеин-оксидаза
Оксидазы цитозоля
Аскорбат
оксидаза
Полифенол
оксидаза
Флавопротеин-оксидаза
 Витамины в жизни человека. Понятия авитаминоз, гиповитаминоз
Витамины в жизни человека. Понятия авитаминоз, гиповитаминоз Какие бывают животные
Какие бывают животные Гомеостаздың физиологиялық механизмдері
Гомеостаздың физиологиялық механизмдері Изменения в жизни домашних животных весной. Труд людей весной (человек и мир, 1 класс)
Изменения в жизни домашних животных весной. Труд людей весной (человек и мир, 1 класс) Степная лисица
Степная лисица Невидимые нити в весеннем лесу. Окружающий мир, 2 класс
Невидимые нити в весеннем лесу. Окружающий мир, 2 класс Шмель Тип: Членистоногие Класс: Насекомые Отряд: Перепончатокрылые Семейство: Пчёлы настоящие
Шмель Тип: Членистоногие Класс: Насекомые Отряд: Перепончатокрылые Семейство: Пчёлы настоящие О Динозаврах. Иллюстрированный атлас мира
О Динозаврах. Иллюстрированный атлас мира Моллюски осьминоги
Моллюски осьминоги Презентация Основные понятия и законы генетики
Презентация Основные понятия и законы генетики многообразие простейших
многообразие простейших Сады средневековья
Сады средневековья Механизмы эволюционного процесса. Главные направления эволюции
Механизмы эволюционного процесса. Главные направления эволюции Древние сосудистые растения
Древние сосудистые растения Проект Использование возможностей ЭОР и сетевых сервисов с целью реализации требований ФГОС
Проект Использование возможностей ЭОР и сетевых сервисов с целью реализации требований ФГОС Красная книга. Редкие животные, растения и грибы Московской области
Красная книга. Редкие животные, растения и грибы Московской области Экология растений. Группы растений по отношению к экологическим факторам
Экология растений. Группы растений по отношению к экологическим факторам Кожа и ее производные. Наружная оболочка тела животного
Кожа и ее производные. Наружная оболочка тела животного Среда обитания организмов. Тухватуллина Азалия
Среда обитания организмов. Тухватуллина Азалия Фотосинтез. 9 класс
Фотосинтез. 9 класс Топография и инервация жевательных и мимических мышц
Топография и инервация жевательных и мимических мышц Слуховой анализатор. Гигиена слуха
Слуховой анализатор. Гигиена слуха певчие птицы россии
певчие птицы россии Болезни мелких животных
Болезни мелких животных Царство Грибы. Общая характеристика грибов
Царство Грибы. Общая характеристика грибов Рептилии или пресмыкающиеся. Черепахи
Рептилии или пресмыкающиеся. Черепахи Сердечно-сосудистая система анатомия и физиология
Сердечно-сосудистая система анатомия и физиология Лабораторная работа Явление плазмолиза и деплазмолиза в клетках кожицы чешуи лука
Лабораторная работа Явление плазмолиза и деплазмолиза в клетках кожицы чешуи лука