- Строение и функция клеточного ядра

Содержание
- 2. Ядро Термин «ядро» впервые был применен Робертом Брауном в 1833 г. Он открыл его в клетках
- 3. Функции клеточного ядра Сохранение генетической информации в неизменном виде: - ХРАНЕНИЕ генетической информации в структуре ДНК.
- 4. Опыты Геммерлинга Доказательства роли ядра в передаче наследственной информации: одноклеточная водоросль (Acetabularia), имеющая форму гриба (шляпка,
- 5. Опыты Астаурова с тутовым шелкопрядом Астауров Борис Львович Объект: два подвида тутового шелкопряда. У одного подвида
- 6. Прямые и косвенные доказательства функции ядра хромосом: Прямыми доказательствами роли ядра являются наследственные болезни, связанные с
- 7. Строение ядра Ядро имеет вид округлой или удлиненной структуры. В ядре различают: Ядерная оболочка (кариолема), Кариоплазма
- 8. Ядерная оболочка Ядерная оболочка состоит из двух мембран: внешней и внутренней, между которыми располагается перинуклеарное пространство.
- 9. Функция ядерной оболочки: Защитная Барьерная Регуляторная Транспортная Фиксирующая
- 10. Ядерная оболочка К наружной ядерной мембране прикреплены полирибосомы, что говорит о тесной связи с ЭПС. С
- 11. Ядерные поры Поры представляют собой восьмиугольный поровый комплекс, состоящий более чем из 100 белков (нуклеопоринов). Ядерный
- 12. Ядерные поры Благодаря порам, осуществляется обмен веществ между ядром и цитоплазмой. Молекулы до 9 нм свободно
- 13. Функция ядерной поры: Барьерная, Регуляторная, Транспортная, Фиксирующая (для хроматина). В то же время ядерные поры осуществляют
- 14. Ядерный сок Ядерный сок (кариоплазма) - внутренняя среда ядра, представляющая собой коллоидное (гелеобразное) вязкое вещество, в
- 15. Ядерный матрикс Ядерный матрикс заполняет пространство между хроматином и ядрышками. Он состоит из белков, метаболитов и
- 16. Ядерная ламина Ламина представляет собой тонкий фиброзный слой, подстилающий внутреннюю мембрану ядерной оболочки. В ее состав
- 17. Ламина сформирована промежуточными филаментами. Она поддерживает ядерную мембрану и контактирует с хроматином и ядерными РНК. Ламины
- 18. Итак, ламины представлены тремя белками (ламины A, B, C). Два из них, ламины A и C,
- 19. Функции ламины Поддерживает форму ядра. Участвует в формировании порового комплекса. Отвечает за упорядоченное расположение хроматина в
- 20. 12 марта 2019 г., 21:02 Учёные лишили геном плодовой мушки каркаса и посмотрели, что с ним
- 21. Диаметр клеточного ядра не превышает двух сотых долей миллиметра, а длина молекулы ДНК достигает почти двух
- 22. «Хотя сегодня накоплен большой объём наблюдений о роли ядерной ламины в регуляции работы генетического аппарата клетки,
- 23. Исследователи выяснили, что в клетках с разрушенной ядерной ламиной ЛАДы перемещаются от оболочки ядра в сторону
- 24. Швейцарские ученые применили метод криоэлектронной томографии в изучении ядерных структур клетки. Ламина ядра клетки - слой,
- 25. Фибриллярная сеть ламины имеет толщину около 14 нм и состоит из более или менее уплотненных зон.
- 26. Прогерия Хатчинсона-Гилфорда - что мы знаем о ней Детская прогерия Как можно прочитать на этом сайте
- 27. Прогерия у взрослых У взрослых синдром прогерии протекает по тем же признакам чрезвычайно быстрого и преждевременного
- 28. Строение и функция хроматина: эу- и гетерохроматин. Хромосомный цикл. Морфология митотических хромосом. Кариотип вида. Уровни компактизации
- 29. Хроматин Хроматин представляет собой хромосомы различной степени раскручивания. Различают два типа хроматина: Гетерохроматин – обладает высокой
- 30. Гетерохоматин Конституционный (постоянный) гетерохроматин - постоянно конденсированные участки хромосом в интерфазных ядрах Конститутивный гетерохроматин генетически не
- 31. Гетерохоматин Факультативный (непостоянный) гетерохроматин - может менять степень своей компактизации в зависимости от функциональной активности, он
- 32. Хроматин – комплекс ДНК и белков (гистонов и негистонов) Хроматин Эухроматин (слабо конденсированный, активный) Гетерохроматин (сильно
- 33. Хромосомная теория наследственности Гены лежат в хромосомах в линейном порядке Каждый ген занимает определенное место –
- 34. Томас Морган Томас Хант Морган (англ. Thomas Hunt Morgan, 25 сентября 1866), Лексингтон — 4 декабря
- 35. Хромосомы к клетке в зависимости от фазы клеточного цикла бывают: Интерфазные, активные Митотические, неактивные интерфаза митоз
- 36. Хромосомный цикл Половые клетки (сперматозоиды и ооциты) с одинарным набором хромосом называют гаплоидными. Плоидность (от греч.
- 38. Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции и др.) Вид интерфазного ядра
- 39. Изучение митотических хромосом – цитогенетика.
- 40. Митотические хромосомы – подобны упакованным для переезда вещам Метафазная хромосома видна в микроскоп и неактивна
- 41. Метафазные хромосомы
- 42. Морфология митотических хромосом Необходимо помнить, что метафазная хромосома – это хромосома в нерабочем состоянии, упакованная для
- 43. Морфология митотических хромосом В области первичной перетяжки (центромеры) расположен кинетохор - пластинчатая структура, имеющая форму диска
- 44. Типы хромосом Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины –
- 45. Размеры и число хромосом Размеры хромосом у разных организмов варьируют в широких пределах от 0,2 до
- 46. ВИДЫ ХРОМОСОМ: ГИГАНТСКИЕ ХРОМОСОМЫ Видны в некоторых клетках на определенных стадиях клеточного цикла. Например, в клетках
- 47. ВИДЫ ХРОМОСОМ: ПОЛИТЕННЫЕ ХРОМОСОМЫ Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster. Впервые обнаружены Е.Г. Бальбиани
- 48. Схема строения политенных хромосом а — нить интерфазной хромосомы; б — две нити после редупликации; в
- 49. ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК Обнаружены Рюккертом в 1892 году. По длине превышают политенные хромосомы, наблюдаются в
- 50. ВИДЫ ХРОМОСОМ: ПОЛИТЕННЫЕ ХРОМОСОМЫ Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster. Впервые обнаружены Е.Г. Бальбиани
- 51. Схема строения политенных хромосом а — нить интерфазной хромосомы; б — две нити после редупликации; в
- 52. ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК Обнаружены Рюккертом в 1892 году. По длине превышают политенные хромосомы, наблюдаются в
- 53. Кариотип домашней кошки Felis catus ( Брайен С. и др. Генетика кошки, 1993). КАРИОТИП Это совокупность
- 54. Белки хроматина Белки в составе хроматина очень разнообразны, но их можно разделить на две группы: гистоны
- 55. Уровни компактизации ДНК Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46
- 56. Уровни компактизации ДНК Двойная спираль ДНК - отрицательно заряженная молекула диаметром 2 нм и длиной несколько
- 57. Уровни компактизации ДНК Соленоидный уровень организации хромосом характеризуется скручиванием нуклеосомной нити и образованием из нее более
- 58. Уровни компактизации ДНК Петлевой уровень обеспечивается негистоновыми сайт-специфическими ДНК-связывающими белками, которые распознают определенные последовательности ДНК и
- 59. Уровни компактизации ДНК Доменный уровень организации хромосом изучен недостаточно. На данном уровне отмечается образование петлевых доменов
- 60. Уровни компактизации ДНК На хромосомном уровне происходит конденсация профазной хромосомы в метафазную с уплотнением петельных доменов
- 61. Канадский ученый Барр (1908 – 1995) и его студент Бертрам открыли в 1948 году в ядрах
- 62. Тельце Барра – пример факультативного гетерохроматина, можно видеть в соматических клетках женского организма млекопитающих
- 63. Исследование полового хроматина – тельца Барра У пациента берется соскоб эпителия ротовой полости Помещается на предметное
- 64. Ген, отвечающий за инактивацию Х-хромосомы, на ней же и лежит. Которая из Х-хромосом будет инактивирована, по-видимому,
- 65. Этапы цитогенетического исследования
- 66. Кровь (или другой материал) Отделение лейкоцитов Добавление стимулятора митоза – ФГА (фитогемагглютинина) 72 часа Добавление колхицина
- 67. Добавление гипотонического раствора – клетки разбухают Х ХХ х х при раскапывании от удара о стекло
- 68. Виды окраски хромосом Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация поделила все хромосомы человека
- 69. Виды метафазных хромосом согласно Денверской классификации Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая, со спутником (первичная перетяжка) вторичная перетяжка
- 70. Парижская классификация основана на дифференциальной окраске (чаще всего G-окраска)
- 71. Плечи делят на районы (бенды) и суббенды
- 72. FISH -метод – Fluorescent in situ hybridization дал еще больше возможностей
- 73. Не для зарисовки!
- 75. FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска
- 76. Исследование кариотипа (кариотипирование = цитогенетическое исследование позволяет диагностировать хромосомные и геномные мутации Хромосомные мутации – изменение
- 77. Хромосомные мутации ЧАСТО ЯВЛЯЮТСЯ РЕЗУЛЬТАТОМ НАРУШЕНИЯ КРОССИНГОВЕРА – ОБМЕНА УЧАСТКАМИ МЕЖДУ ХРОМОСОМАМИ, ПРОИСХОДЯЩЕГО В ПРОФАЗЕ 1
- 78. Внутрихромосомные перестройки Делеция Дупликация Инверсия Кольцевая хромосомам Изохромосома
- 79. Делеция(del) Дупликация(dup) Инверсия(inv)
- 80. Варианты инверсий перицентрическая парацентрическая
- 81. Возникновение изохромосомы (i) при неправильном разделении хроматид p q p q p q p q Нормальное
- 82. Образование кольцевой хромосомы (r) кольцо ацентрические фрагменты
- 83. Межхромосомные перестройки - транслокации Взаимные Невзаимные Робертсоновские
- 84. Транслокация (t):1 – невзаимная (инсерция), 2 – взаимная (реципрокная)
- 85. Робертсоновская транслокация (rob) (центрическое слияние) Между разными акроцентриками – у человека 13,14,15,21,22
- 86. В результате транслокаций могут возникать дицентрические (dic) хромосомы и парные ацентрические фрагменты
- 87. Кроме того, хромосомные мутации бывают: Спонтанные/индуцированные Соматические /генеративные Вредные/полезные/нейтральные
- 88. С клинической точки зрения хромосомные мутации удобнее делить на Сбалансированные (нет потери или добавления генов) Например,
- 89. Значение хромосомных мутаций Материал для эволюции, способствует появлению новых видов Патология у человека
- 90. Сравнение хромосомных наборов человека (слева) и шимпанзе (справа). Видно, что наша хромосома 2 – результат Робертсоновской
- 91. При лейкозах выявляются множественные хромосомные перестройки
- 92. Самый известный пример: Филадельфийская хромосома – транслокация между 22 и 9 хромосомами – пример соматической мутации
- 93. Различные случаи делеций -- Синдром кошачьего крика -- Синдром Вольфа-Хиршхорна
- 94. Делеция короткого плеча хромосомы 5 – синдром кошачьего крика, cri du chat
- 95. Хромосомные карты Генетические – где лежит какой ген Цитологические – по окраске Физические – основаны на
- 96. Карта хромосомы 9
- 97. Карта хромосомы 21 и митохондриального генома
- 98. 24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС,
- 99. 24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б., Карамышева Т. В. Вестн. ВОГиС,
- 100. ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
- 101. Идиограмма Идиограмма схематическое изображение гаплоидного набора хромосом организма, которые располагают в ряд в соответствии с их
- 102. В митотической хромосоме ДНК упакована в 10000 раз
- 103. Половой хроматин Исследования хроматина показали, что у самок имеются определенные скопления гетерохроматина, а у самцов таких
- 104. Структура и функция ядрышек. Гранулярный и фибриллярный компоненты ядрышек. Фибриллярные центры и ядрышковый организатор. Структурные типы
- 105. Впервые ядрышки были описаны Фонтана в 1774 г. В живых клетках они выделяются на фоне диффузной
- 106. По данным цитохимических и биохимических исследований основным компонентом ядрышка является белок: на его долю приходится до
- 107. Встречаются во всех эукариотических клетках. К таким исключениям относятся клетки дробящихся яиц, где ядрышки отсутствуют на
- 108. Число и размеры ядрышек в клетках непостоянны. Локализация ядрышковых организаторов определяется довольно точно на митотических хромосомах
- 109. амплификации генов рРНК амплификация рДНК, происходит в профазе I деления созревания, когда синтез хромосомной ДНК давно
- 110. Биологический смысл появления сверхчисленных экстрахромосомных ядрышек при росте ооцитов совершенно понятен: для синтеза огромного количества запасных
- 111. Структура ядрышка
- 112. Структурные типы ядрышек
- 113. Ядрышко Ядрышко представляет собой сферическую структуру богатую рРНК и белком. При окраске гематоксилином и эозином ядрышко
- 114. Клеточный цикл, Митоз, Мейоз
- 115. Клеточный цикл Клеточный цикл – это период жизнедеятельности клетки от конца одного деления до конца следующего.
- 116. Клеточный цикл включает строго детерминированный ряд последовательных процессов, согласно позиции Hartwellа, 1995. Клетка должна между двумя
- 117. Интерфаза: период G1 (пресинтетический) Пресинтетический период G1 наступает сразу после деления клетки. В этом периоде клетка
- 118. Интерфаза: период S (синтез ДНК) В периоде S происходит репликация (синтез или копирование) ДНК. Точность синтеза
- 119. Кроме репликации ДНК, в периоде S также происходит удвоение центриолей клеточного центра. Материнская центриоль строит в
- 120. Интерфаза: период G2 (постсинтетический) Окончание стадии S является началом стадии G2. Постсинтетический период – это финальная
- 122. Фаза репликации количества ДНК (S –фаза) При подготовке к делению происходит удвоение молекул ДНК, на каждой
- 123. Постсинтетический период (G 2 фаза) В этот период происходит активное накопление энергии и ферментов, необходимых для
- 124. Митоз Митоз (от греч. mitos - нить) - деление ядра, следующее за репликацией хромосом, в результате
- 125. Профаза В начале профазы многочисленные цитоплазматические микротрубочки, входящие в состав цитоскелета, распадаются; при этом образуется большой
- 126. Метафаза Эта стадия митоза часто продолжается длительное время Все хромосомы располагаются таким образом, что их центромеры
- 128. Анафаза Продолжается обычно всего несколько минут. Анафаза начинается внезапным расщеплением каждой хромосомы, которое обусловлено разделением сестринских
- 129. Телофаза В заключительной стадии митоза телофазе разделенные дочерние хроматиды подходят к полюсам, кинетохорные нити исчезают. После
- 130. Цитокинез Процесс деления цитоплазмы – цитокинез, проходит под действием сократимого кольца и начинается обычно в поздней
- 131. Мейоз При половом размножении растений и животных (в том числе и человека) преемственность между поколениями обеспечивается
- 132. При первом делении мейоза каждая дочерняя клетка наследует две копии одного из двух гомологов и поэтому
- 133. Профаза I Эта фаза состоит из пяти основных стадий: Лептотены, Зиготены, Пахитены, Диплотены, Диакинез Схема мейоза:
- 134. Лептотена Каждая хромосома, изменив свою интерфазную конформацию, переходит в конденсированную форму, образуя длинное, тонкое волокно с
- 135. Зиготена Зиготена - стадия спаривания хромосом. Моментом перехода лептотены в зиготену считают начало синапсиса - тесной
- 136. Пахитена Пахитена - стадия профазы мейоза, на которой спаривание гомологов завершено. Хромосомы выглядят более толстыми, чем
- 137. Диплотена Диплотена - стадия мейоза после пахитены и перед диакинезом. Стадия расхождения хромосом. В диплотене гомологичные
- 138. Диакинез Диакинез - заключительная стадия профазы I. В диакинезе биваленты резко укороченные, утолщенные дочерние хроматиды каждой
- 140. Метафаза I. Число бивалентов вдвое меньше от диплоидного набора хромосом. Биваленты значительно менее короткие, чем хромосомы
- 142. Скачать презентацию

Ядро
Термин «ядро» впервые был применен Робертом Брауном в 1833 г.
Он
Ядро
Термин «ядро» впервые был применен Робертом Брауном в 1833 г.
Он

Функции клеточного ядра
Сохранение генетической информации в неизменном виде:
- ХРАНЕНИЕ
Функции клеточного ядра
Сохранение генетической информации в неизменном виде:
- ХРАНЕНИЕ
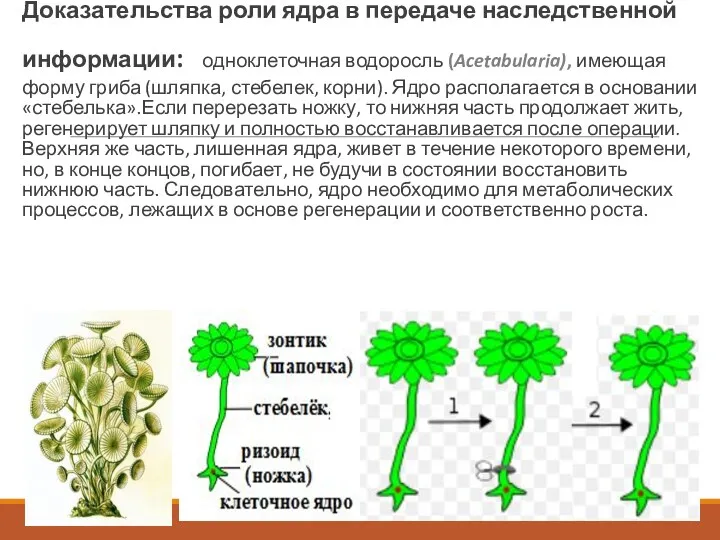
Опыты Геммерлинга
Доказательства роли ядра в передаче наследственной информации: одноклеточная
Опыты Геммерлинга Доказательства роли ядра в передаче наследственной информации: одноклеточная

Опыты Астаурова с тутовым шелкопрядом
Астауров
Борис Львович
Объект: два подвида тутового шелкопряда.
Опыты Астаурова с тутовым шелкопрядом
Астауров
Борис Львович
Объект: два подвида тутового шелкопряда.

Прямые и косвенные доказательства функции ядра хромосом:
Прямыми доказательствами роли ядра являются
Прямые и косвенные доказательства функции ядра хромосом:
Прямыми доказательствами роли ядра являются
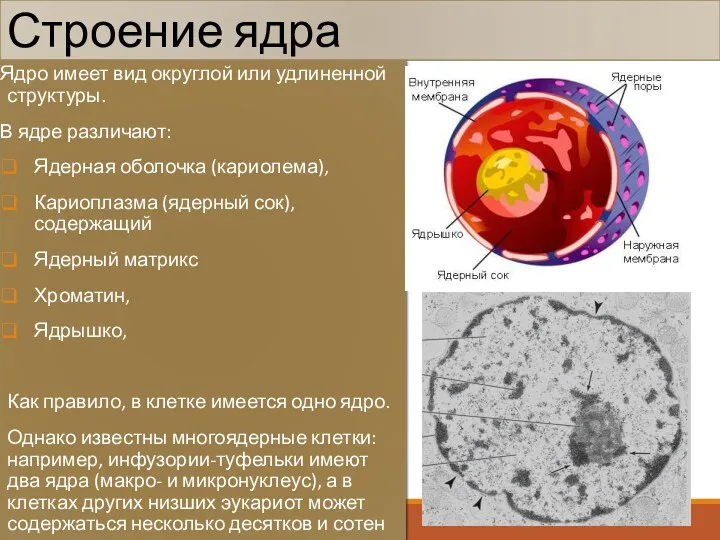
Строение ядра
Ядро имеет вид округлой или удлиненной структуры.
В ядре различают:
Ядерная оболочка
Строение ядра
Ядро имеет вид округлой или удлиненной структуры.
В ядре различают:
Ядерная оболочка
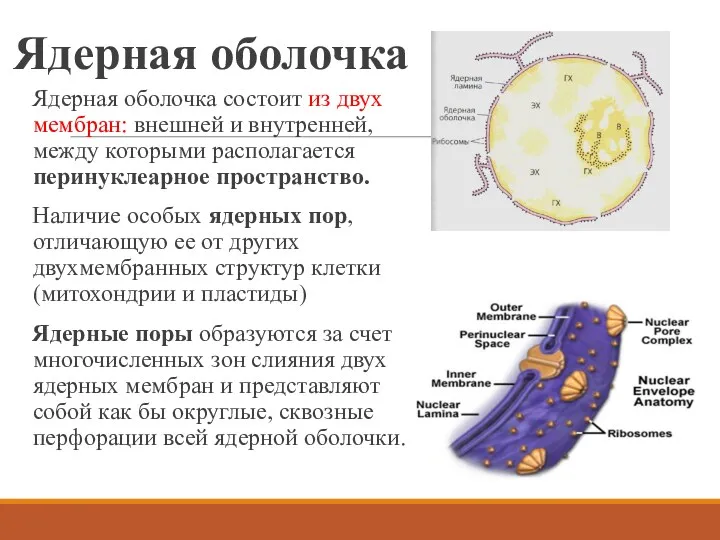
Ядерная оболочка
Ядерная оболочка состоит из двух мембран: внешней и внутренней, между
Ядерная оболочка
Ядерная оболочка состоит из двух мембран: внешней и внутренней, между

Функция ядерной оболочки:
Защитная
Барьерная
Регуляторная
Транспортная
Фиксирующая
Функция ядерной оболочки:
Защитная
Барьерная
Регуляторная
Транспортная
Фиксирующая
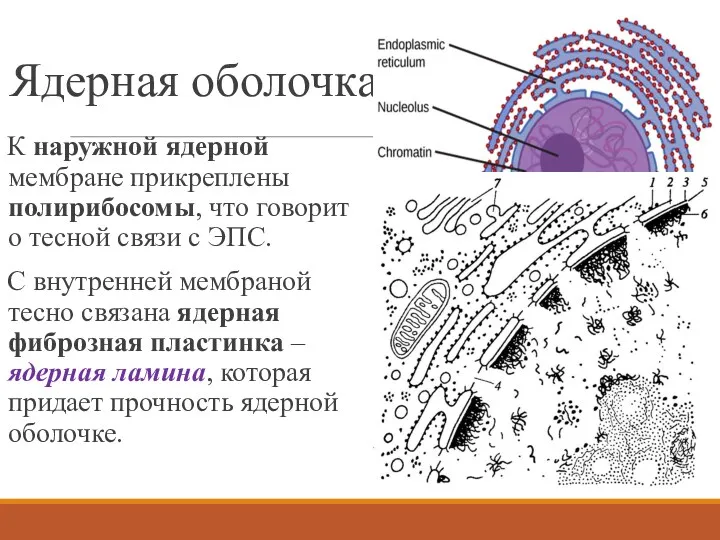
Ядерная оболочка
К наружной ядерной мембране прикреплены полирибосомы, что говорит о тесной
Ядерная оболочка
К наружной ядерной мембране прикреплены полирибосомы, что говорит о тесной
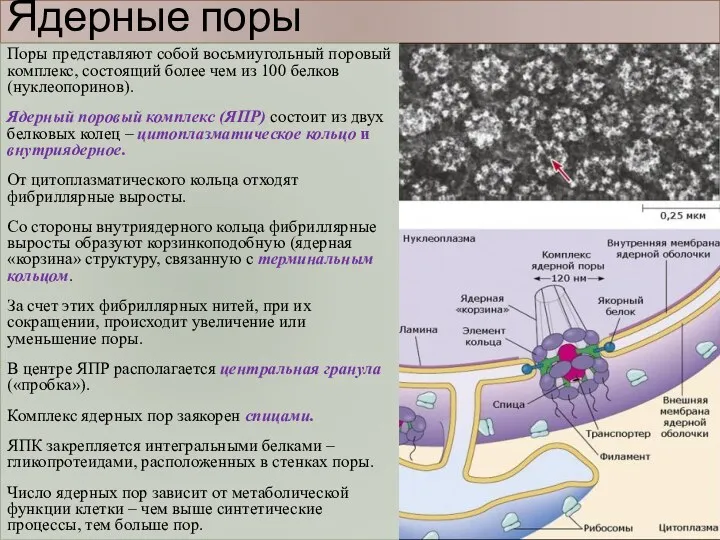
Ядерные поры
Поры представляют собой восьмиугольный поровый комплекс, состоящий более чем из
Ядерные поры
Поры представляют собой восьмиугольный поровый комплекс, состоящий более чем из

Ядерные поры
Благодаря порам, осуществляется обмен веществ между ядром и цитоплазмой.
Молекулы
Ядерные поры
Благодаря порам, осуществляется обмен веществ между ядром и цитоплазмой.
Молекулы
Функция ядерной поры:
Барьерная,
Регуляторная,
Транспортная,
Фиксирующая (для хроматина).
В то
Функция ядерной поры:
Барьерная,
Регуляторная,
Транспортная,
Фиксирующая (для хроматина).
В то

Ядерный сок
Ядерный сок (кариоплазма) - внутренняя среда ядра, представляющая собой коллоидное
Ядерный сок
Ядерный сок (кариоплазма) - внутренняя среда ядра, представляющая собой коллоидное
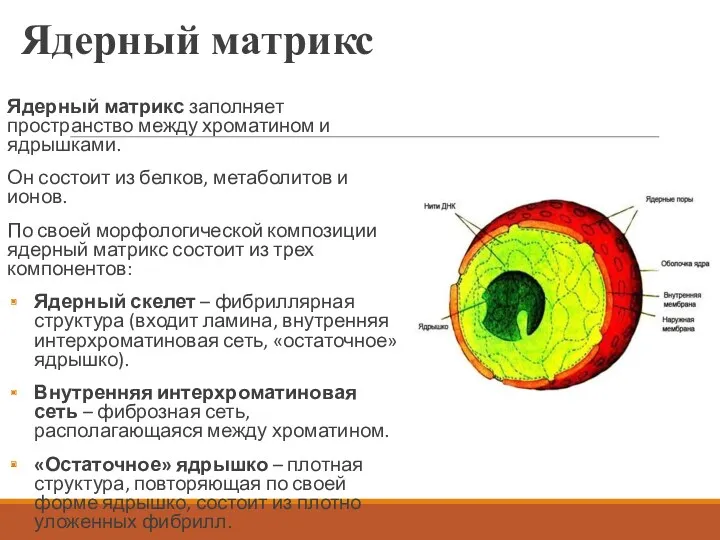
Ядерный матрикс
Ядерный матрикс заполняет пространство между хроматином и ядрышками.
Он состоит из
Ядерный матрикс
Ядерный матрикс заполняет пространство между хроматином и ядрышками.
Он состоит из
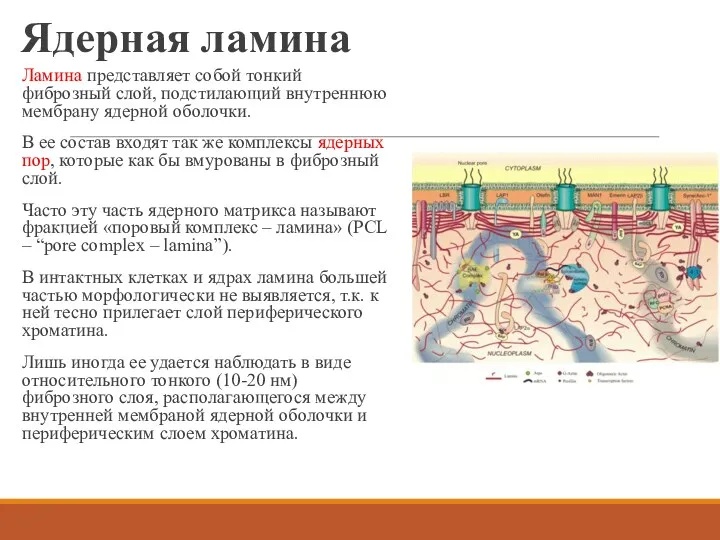
Ядерная ламина
Ламина представляет собой тонкий фиброзный слой, подстилающий внутреннюю мембрану ядерной
Ядерная ламина
Ламина представляет собой тонкий фиброзный слой, подстилающий внутреннюю мембрану ядерной
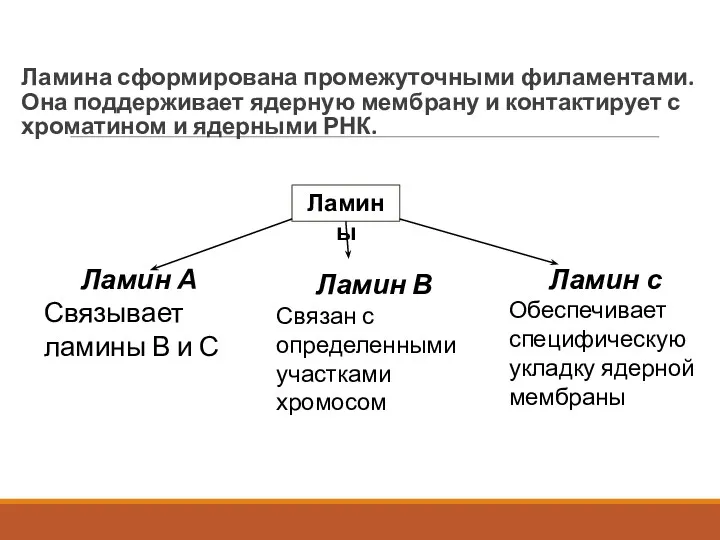
Ламина сформирована промежуточными филаментами. Она поддерживает ядерную мембрану и контактирует с
Ламина сформирована промежуточными филаментами. Она поддерживает ядерную мембрану и контактирует с

Итак, ламины представлены тремя белками (ламины A, B, C).
Два из
Итак, ламины представлены тремя белками (ламины A, B, C).
Два из

Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
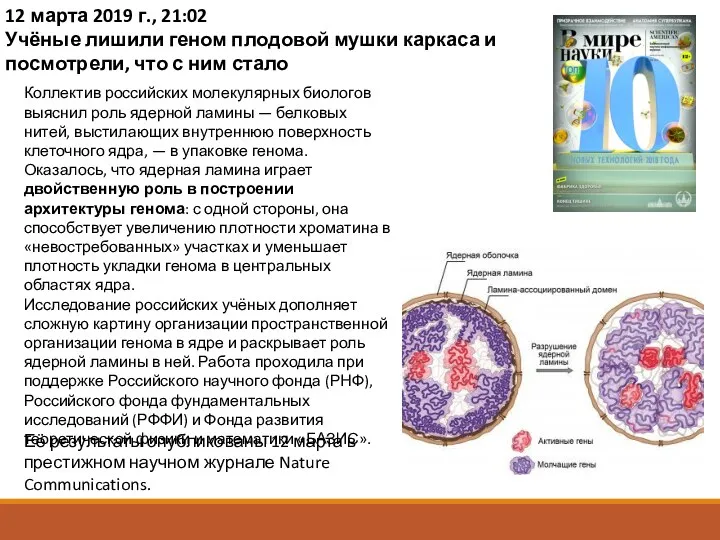
12 марта 2019 г., 21:02
Учёные лишили геном плодовой мушки каркаса и
12 марта 2019 г., 21:02
Учёные лишили геном плодовой мушки каркаса и

Диаметр клеточного ядра не превышает двух сотых долей миллиметра, а длина
Диаметр клеточного ядра не превышает двух сотых долей миллиметра, а длина

«Хотя сегодня накоплен большой объём наблюдений о роли ядерной ламины в
«Хотя сегодня накоплен большой объём наблюдений о роли ядерной ламины в

Исследователи выяснили, что в клетках с разрушенной ядерной ламиной ЛАДы перемещаются
Исследователи выяснили, что в клетках с разрушенной ядерной ламиной ЛАДы перемещаются

Швейцарские ученые применили метод криоэлектронной томографии в изучении ядерных структур клетки.
Ламина
Швейцарские ученые применили метод криоэлектронной томографии в изучении ядерных структур клетки.
Ламина

Фибриллярная сеть ламины имеет толщину около 14 нм и состоит из
Фибриллярная сеть ламины имеет толщину около 14 нм и состоит из

Прогерия Хатчинсона-Гилфорда - что мы знаем о ней
Детская прогерия
Как можно прочитать
Прогерия Хатчинсона-Гилфорда - что мы знаем о ней
Детская прогерия
Как можно прочитать

Прогерия у взрослых
У взрослых синдром прогерии протекает по тем же признакам
Прогерия у взрослых
У взрослых синдром прогерии протекает по тем же признакам

Строение и функция хроматина: эу- и гетерохроматин. Хромосомный цикл. Морфология митотических
Строение и функция хроматина: эу- и гетерохроматин. Хромосомный цикл. Морфология митотических
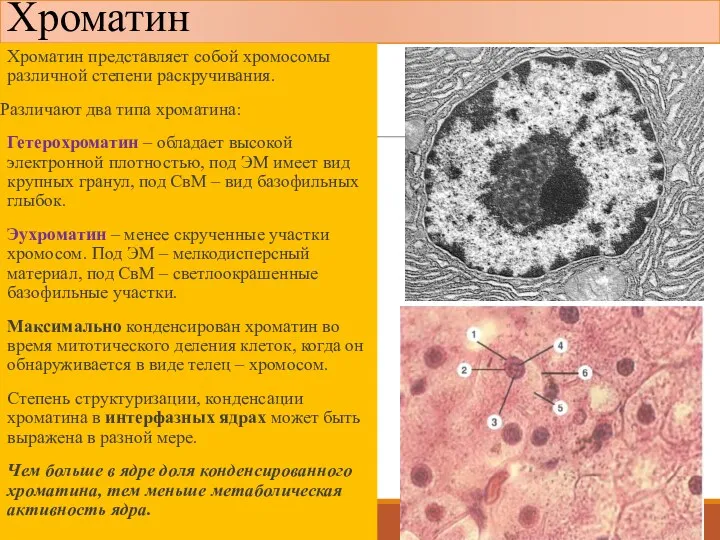
Хроматин
Хроматин представляет собой хромосомы различной степени раскручивания.
Различают два типа хроматина:
Гетерохроматин –
Хроматин
Хроматин представляет собой хромосомы различной степени раскручивания.
Различают два типа хроматина:
Гетерохроматин –
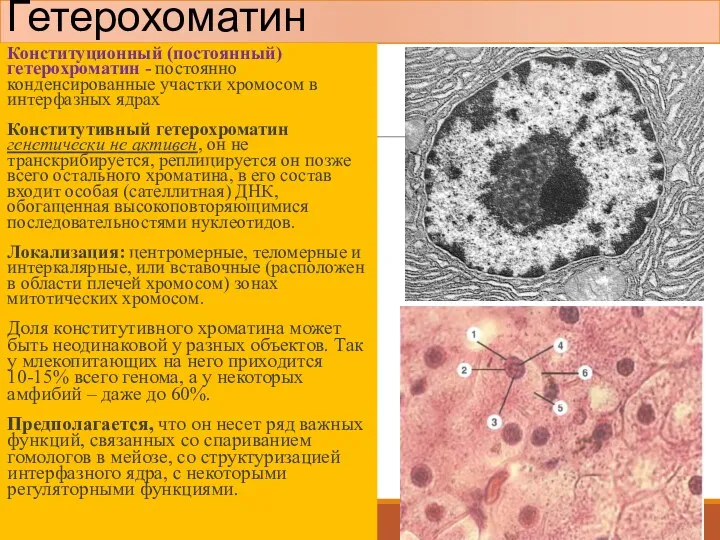
Гетерохоматин
Конституционный (постоянный) гетерохроматин - постоянно конденсированные участки хромосом в интерфазных ядрах
Гетерохоматин
Конституционный (постоянный) гетерохроматин - постоянно конденсированные участки хромосом в интерфазных ядрах
Гетерохоматин
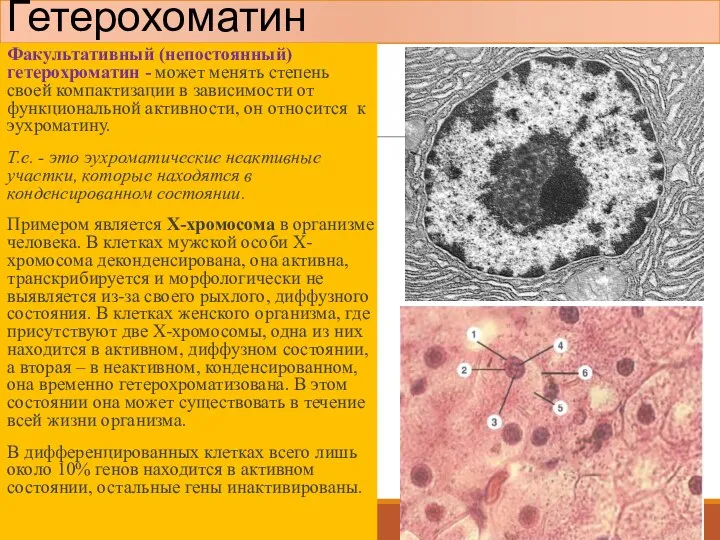
Факультативный (непостоянный) гетерохроматин - может менять степень своей компактизации в зависимости
Гетерохоматин
Факультативный (непостоянный) гетерохроматин - может менять степень своей компактизации в зависимости

Хроматин – комплекс ДНК и белков (гистонов и негистонов)
Хроматин
Эухроматин (слабо
Хроматин – комплекс ДНК и белков (гистонов и негистонов)
Хроматин
Эухроматин (слабо

Хромосомная теория наследственности
Гены лежат в хромосомах в линейном порядке
Каждый ген
Хромосомная теория наследственности
Гены лежат в хромосомах в линейном порядке
Каждый ген

Томас Морган
Томас Хант Морган (англ. Thomas Hunt Morgan, 25 сентября 1866),
Томас Морган
Томас Хант Морган (англ. Thomas Hunt Morgan, 25 сентября 1866),
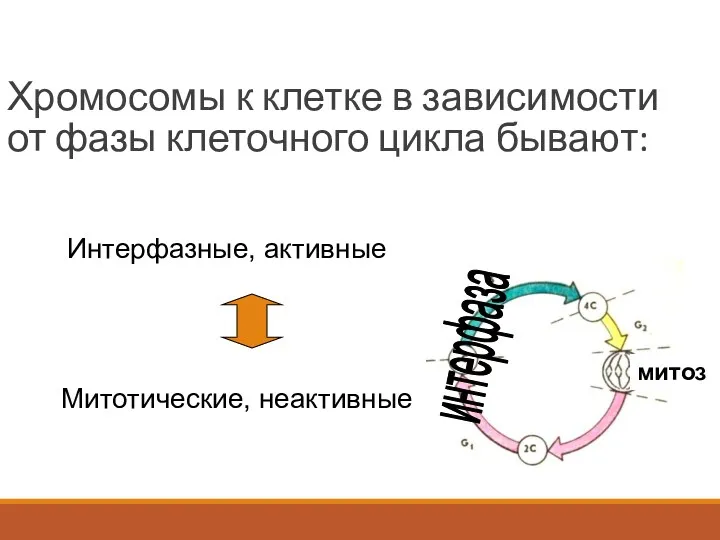
Хромосомы к клетке в зависимости от фазы клеточного цикла бывают:
Интерфазные,
Хромосомы к клетке в зависимости от фазы клеточного цикла бывают:
Интерфазные,

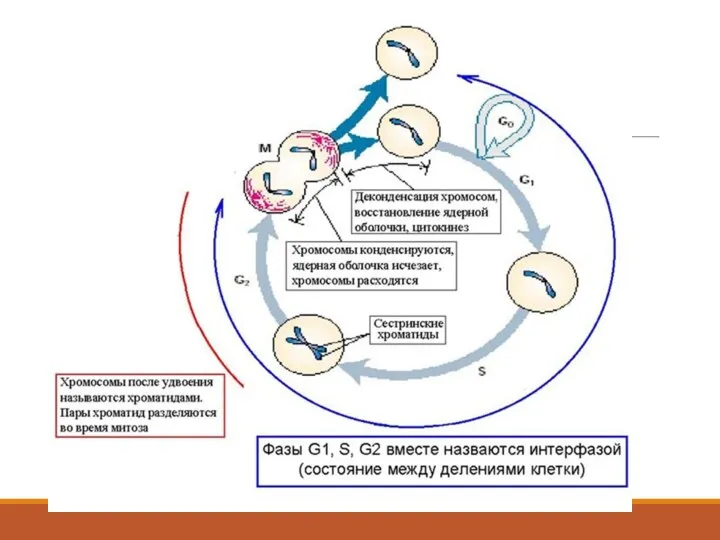
Хромосомный цикл
Половые клетки (сперматозоиды и ооциты) с одинарным набором хромосом называют
Хромосомный цикл
Половые клетки (сперматозоиды и ооциты) с одинарным набором хромосом называют
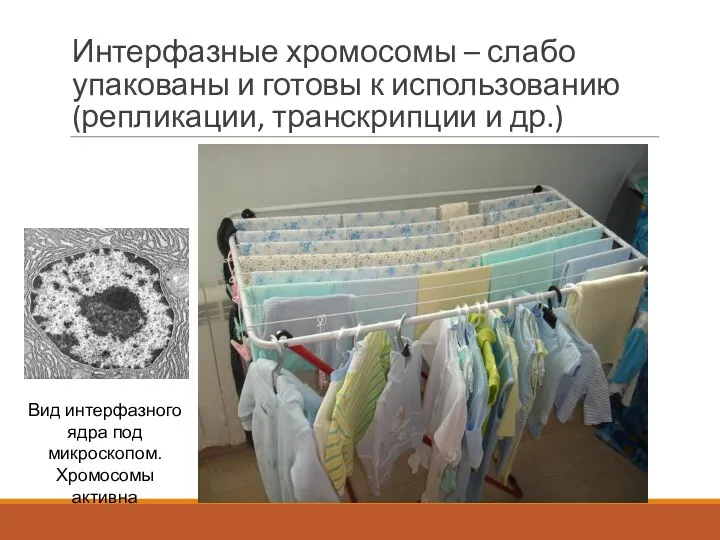
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции

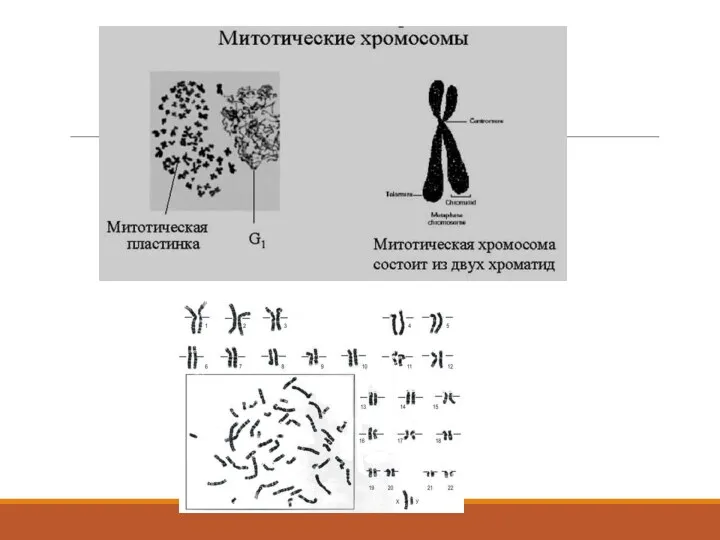
Изучение митотических хромосом – цитогенетика.
Изучение митотических хромосом – цитогенетика.

Митотические хромосомы – подобны упакованным для переезда вещам
Метафазная хромосома видна в
Митотические хромосомы – подобны упакованным для переезда вещам
Метафазная хромосома видна в
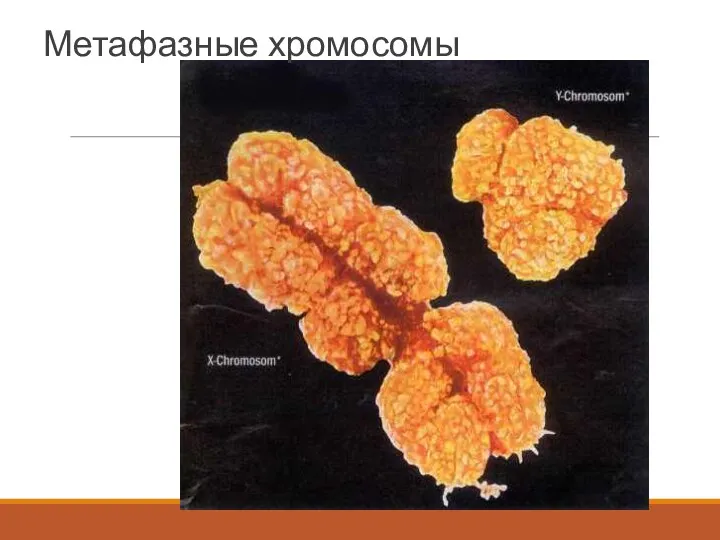
Метафазные хромосомы
Метафазные хромосомы
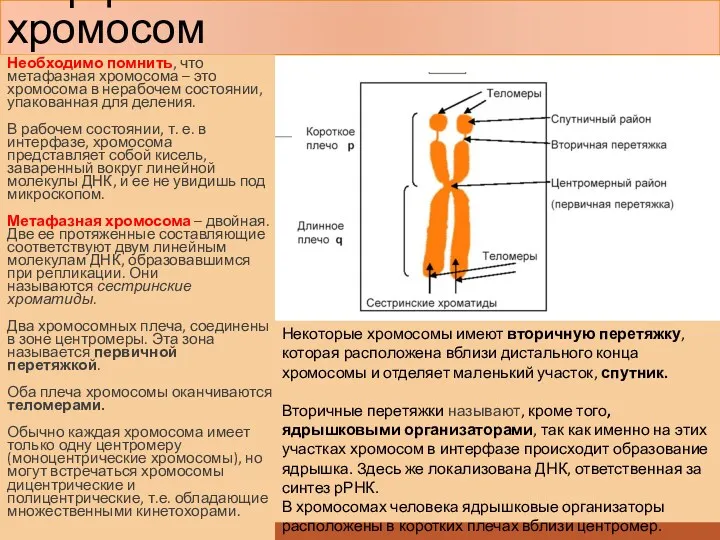
Морфология митотических хромосом
Необходимо помнить, что метафазная хромосома – это хромосома в нерабочем
Морфология митотических хромосом
Необходимо помнить, что метафазная хромосома – это хромосома в нерабочем
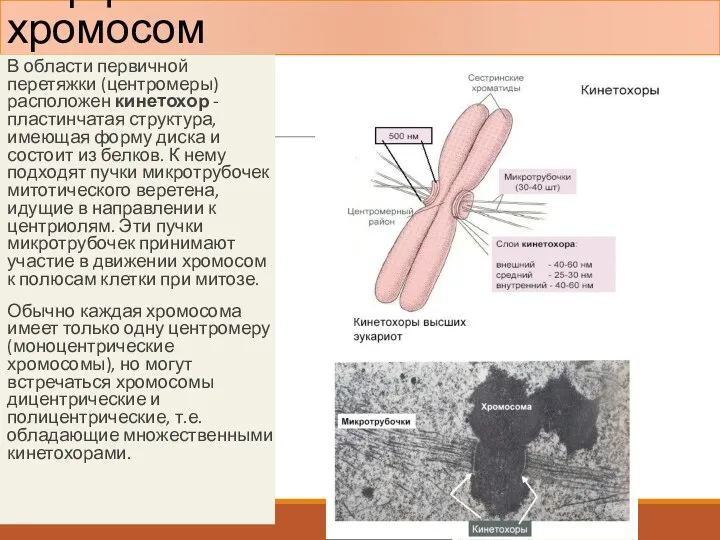
Морфология митотических хромосом
В области первичной перетяжки (центромеры) расположен кинетохор - пластинчатая
Морфология митотических хромосом
В области первичной перетяжки (центромеры) расположен кинетохор - пластинчатая
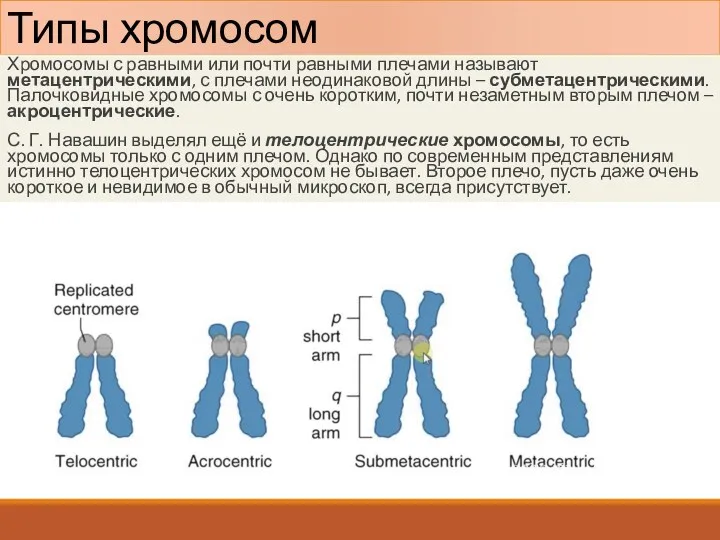
Типы хромосом
Хромосомы с равными или почти равными плечами называют метацентрическими, с
Типы хромосом
Хромосомы с равными или почти равными плечами называют метацентрическими, с

Размеры и число хромосом
Размеры хромосом у разных организмов варьируют в широких
Размеры и число хромосом
Размеры хромосом у разных организмов варьируют в широких

ВИДЫ ХРОМОСОМ:
ГИГАНТСКИЕ ХРОМОСОМЫ
Видны в некоторых клетках на определенных стадиях клеточного цикла.
ВИДЫ ХРОМОСОМ:
ГИГАНТСКИЕ ХРОМОСОМЫ
Видны в некоторых клетках на определенных стадиях клеточного цикла.

ВИДЫ ХРОМОСОМ:
ПОЛИТЕННЫЕ ХРОМОСОМЫ
Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster.
Впервые
ВИДЫ ХРОМОСОМ:
ПОЛИТЕННЫЕ ХРОМОСОМЫ
Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster.
Впервые

Схема строения политенных хромосом
а — нить интерфазной хромосомы; б —
Схема строения политенных хромосом
а — нить интерфазной хромосомы; б —

ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК
Обнаружены Рюккертом в 1892 году.
По длине превышают
ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК
Обнаружены Рюккертом в 1892 году.
По длине превышают
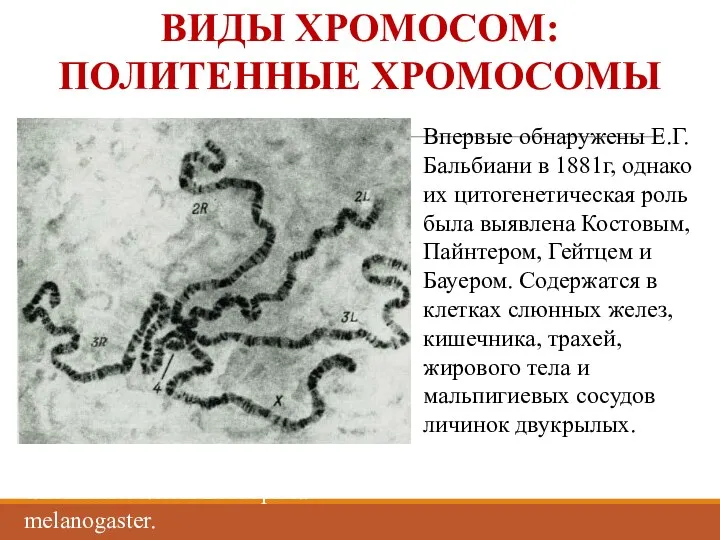
ВИДЫ ХРОМОСОМ:
ПОЛИТЕННЫЕ ХРОМОСОМЫ
Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster.
Впервые
ВИДЫ ХРОМОСОМ:
ПОЛИТЕННЫЕ ХРОМОСОМЫ
Гигантские хромосомы из клеток слюнной железы Drosophila melanogaster.
Впервые

Схема строения политенных хромосом
а — нить интерфазной хромосомы; б —
Схема строения политенных хромосом
а — нить интерфазной хромосомы; б —
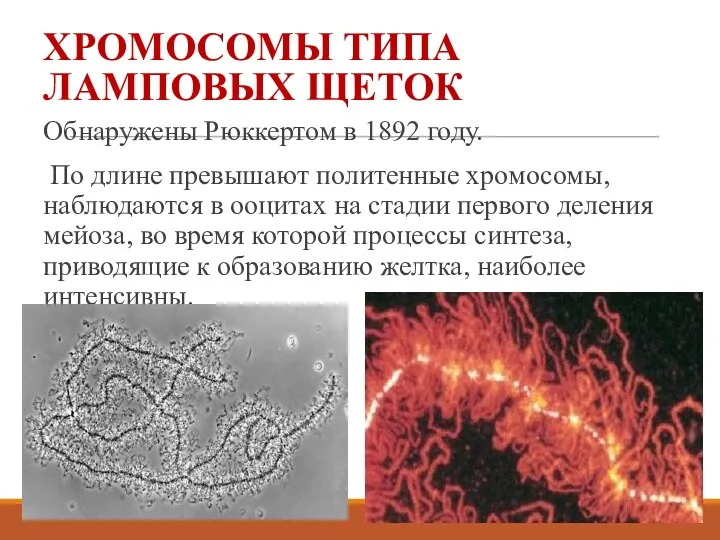
ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК
Обнаружены Рюккертом в 1892 году.
По длине превышают
ХРОМОСОМЫ ТИПА ЛАМПОВЫХ ЩЕТОК
Обнаружены Рюккертом в 1892 году.
По длине превышают

Кариотип домашней кошки Felis catus
( Брайен С. и др. Генетика
Кариотип домашней кошки Felis catus ( Брайен С. и др. Генетика

Белки хроматина
Белки в составе хроматина очень разнообразны, но их можно разделить
Белки хроматина
Белки в составе хроматина очень разнообразны, но их можно разделить

Уровни компактизации ДНК
Каждая молекула ДНК упакована в отдельную хромосому.
В диплоидных
Уровни компактизации ДНК
Каждая молекула ДНК упакована в отдельную хромосому.
В диплоидных
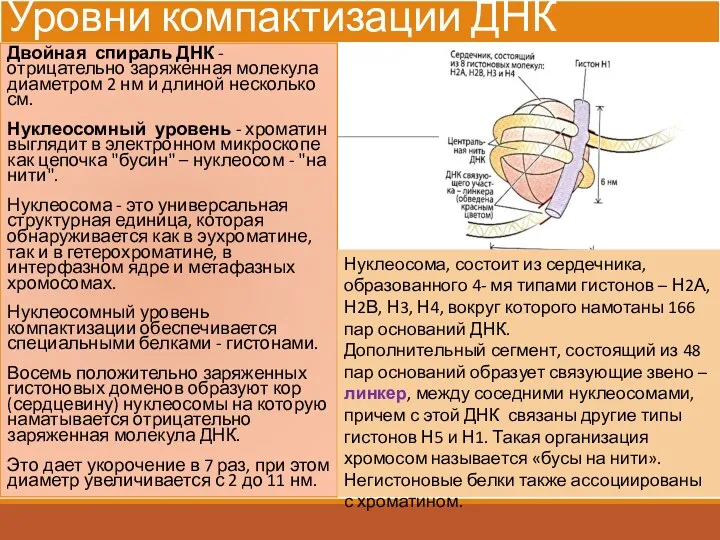
Уровни компактизации ДНК
Двойная спираль ДНК - отрицательно заряженная молекула диаметром 2
Уровни компактизации ДНК
Двойная спираль ДНК - отрицательно заряженная молекула диаметром 2
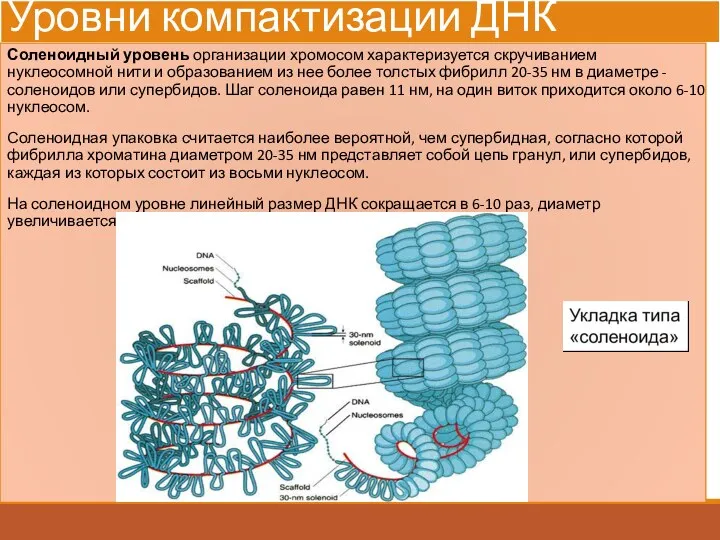
Уровни компактизации ДНК
Соленоидный уровень организации хромосом характеризуется скручиванием нуклеосомной нити и
Уровни компактизации ДНК
Соленоидный уровень организации хромосом характеризуется скручиванием нуклеосомной нити и
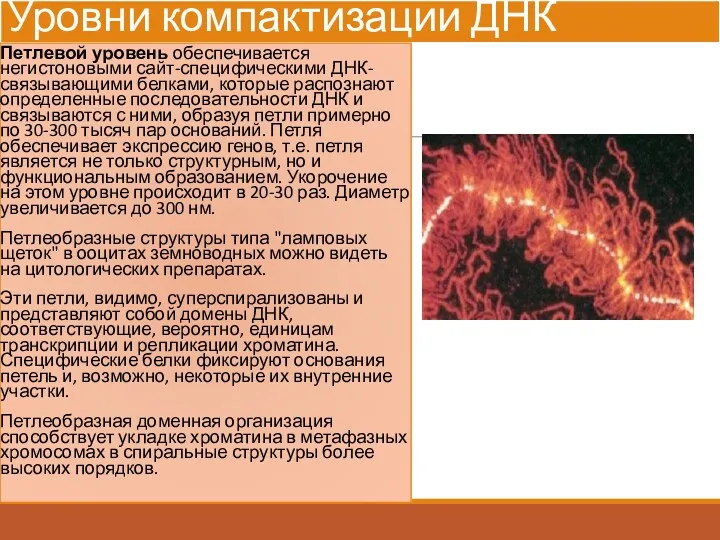
Уровни компактизации ДНК
Петлевой уровень обеспечивается негистоновыми сайт-специфическими ДНК-связывающими белками, которые распознают
Уровни компактизации ДНК
Петлевой уровень обеспечивается негистоновыми сайт-специфическими ДНК-связывающими белками, которые распознают

Уровни компактизации ДНК
Доменный уровень организации хромосом изучен недостаточно. На данном уровне
Уровни компактизации ДНК
Доменный уровень организации хромосом изучен недостаточно. На данном уровне
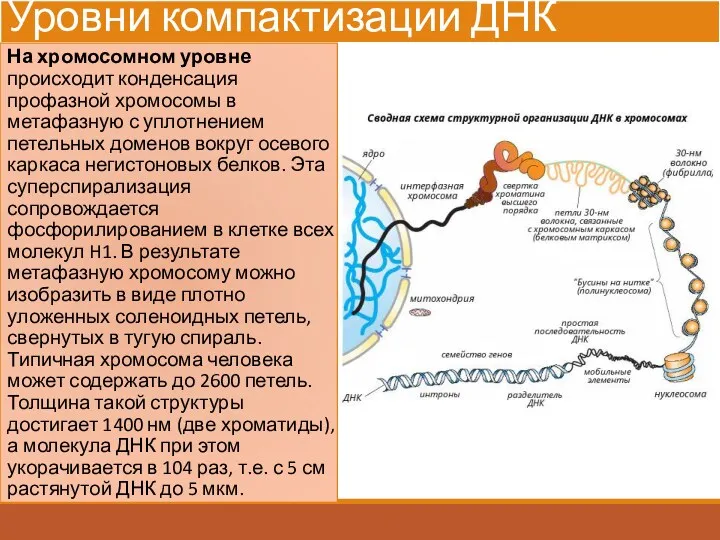
Уровни компактизации ДНК
На хромосомном уровне происходит конденсация профазной хромосомы в метафазную
Уровни компактизации ДНК
На хромосомном уровне происходит конденсация профазной хромосомы в метафазную
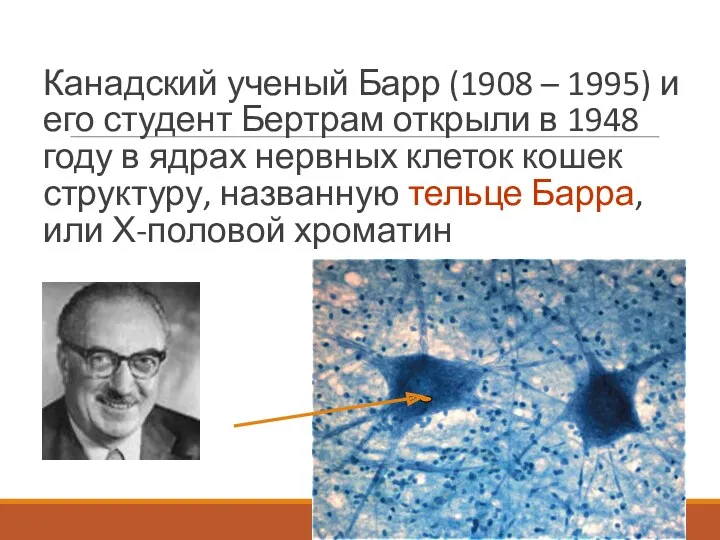
Канадский ученый Барр (1908 – 1995) и его студент Бертрам открыли
Канадский ученый Барр (1908 – 1995) и его студент Бертрам открыли
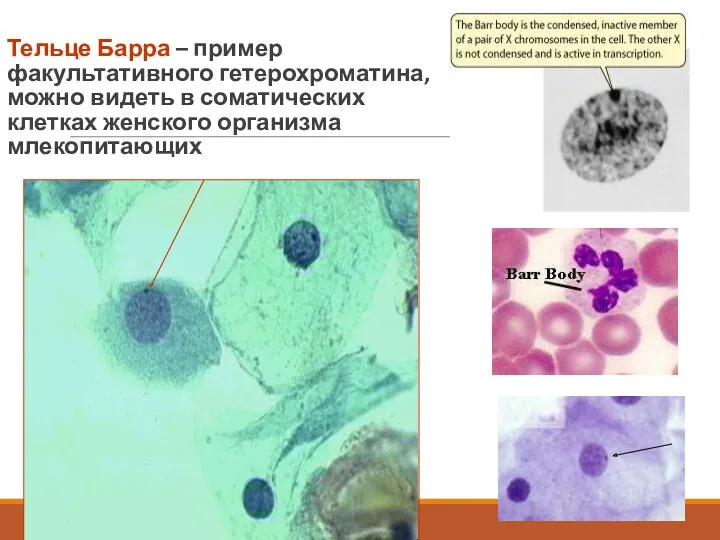
Тельце Барра – пример факультативного гетерохроматина, можно видеть в соматических клетках
Тельце Барра – пример факультативного гетерохроматина, можно видеть в соматических клетках

Исследование полового хроматина – тельца Барра
У пациента берется соскоб эпителия ротовой
Исследование полового хроматина – тельца Барра
У пациента берется соскоб эпителия ротовой

Ген, отвечающий за инактивацию Х-хромосомы, на ней же и лежит. Которая
Ген, отвечающий за инактивацию Х-хромосомы, на ней же и лежит. Которая

Этапы цитогенетического исследования
Этапы цитогенетического исследования
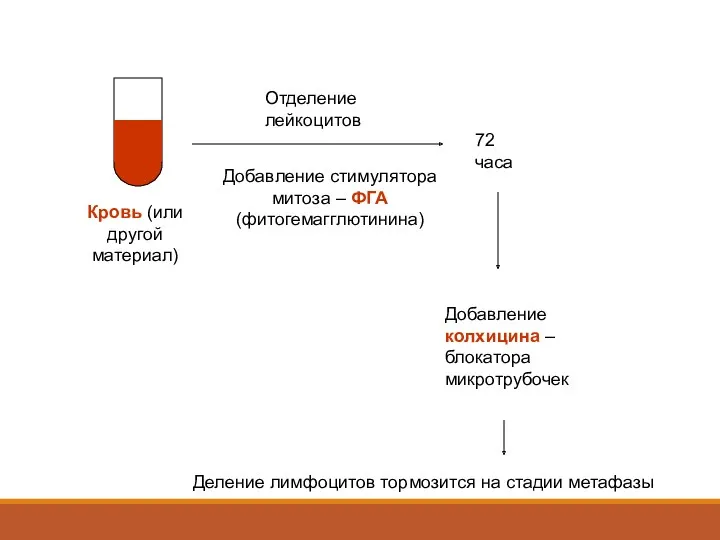
Кровь (или другой материал)
Отделение лейкоцитов
Добавление стимулятора митоза – ФГА (фитогемагглютинина)
72 часа
Добавление
Кровь (или другой материал)
Отделение лейкоцитов
Добавление стимулятора митоза – ФГА (фитогемагглютинина)
72 часа
Добавление
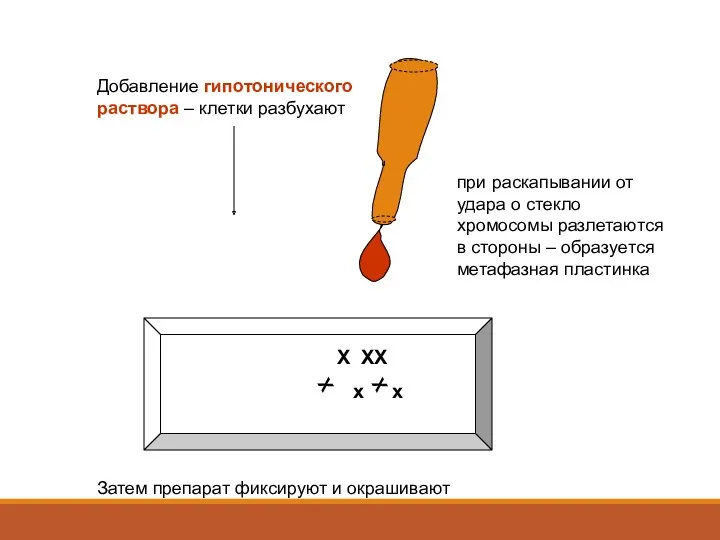
Добавление гипотонического раствора – клетки разбухают
Х ХХ
х х
при раскапывании
Добавление гипотонического раствора – клетки разбухают
Х ХХ
х х
при раскапывании

Виды окраски хромосом
Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация
Виды окраски хромосом
Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация
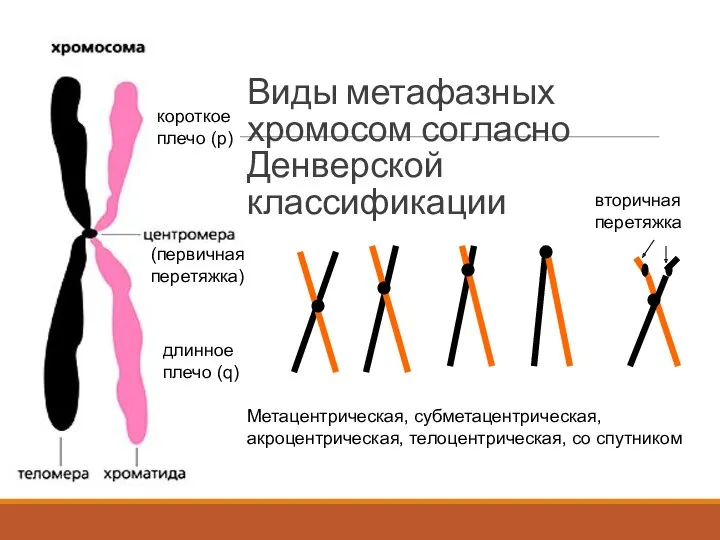
Виды метафазных хромосом согласно Денверской классификации
Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая, со спутником
(первичная
Виды метафазных хромосом согласно Денверской классификации
Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая, со спутником
(первичная

Парижская классификация основана на дифференциальной окраске (чаще всего G-окраска)
Парижская классификация основана на дифференциальной окраске (чаще всего G-окраска)
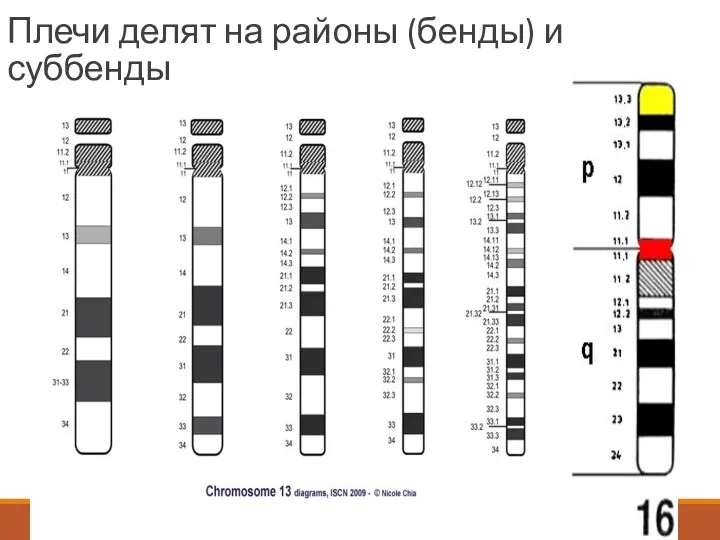
Плечи делят на районы (бенды) и суббенды
Плечи делят на районы (бенды) и суббенды
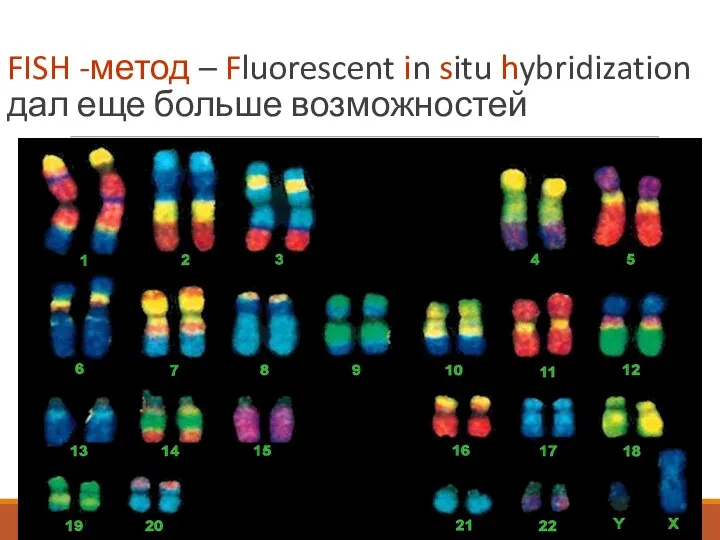
FISH -метод – Fluorescent in situ hybridization дал еще больше возможностей
FISH -метод – Fluorescent in situ hybridization дал еще больше возможностей
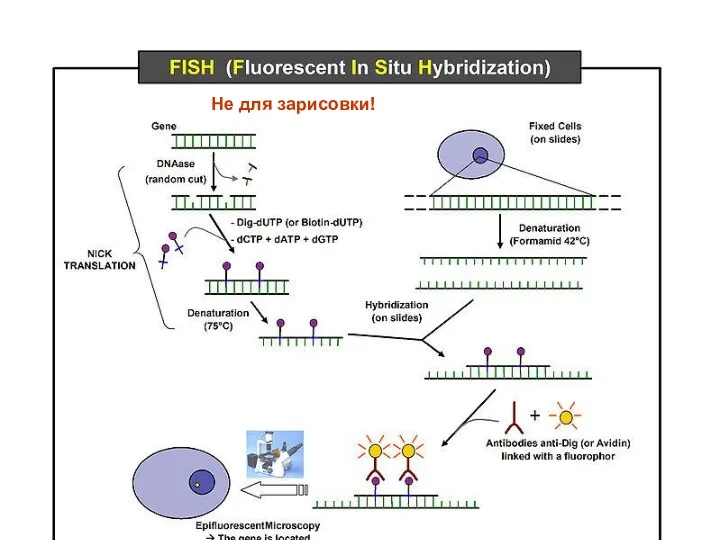
Не для зарисовки!
Не для зарисовки!
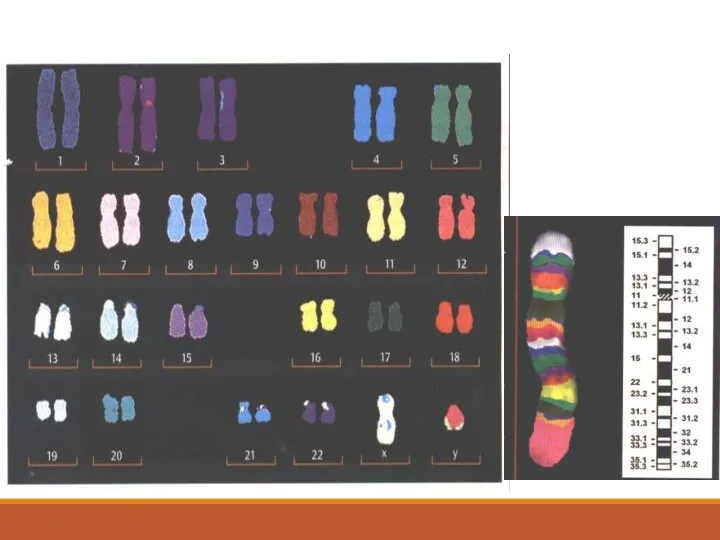
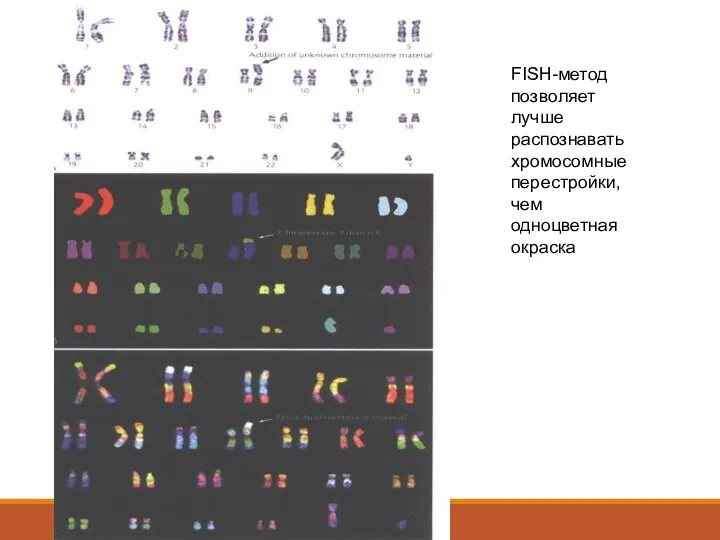
FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска
FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска

Исследование кариотипа (кариотипирование = цитогенетическое исследование позволяет диагностировать хромосомные и геномные
Исследование кариотипа (кариотипирование = цитогенетическое исследование позволяет диагностировать хромосомные и геномные

Хромосомные мутации
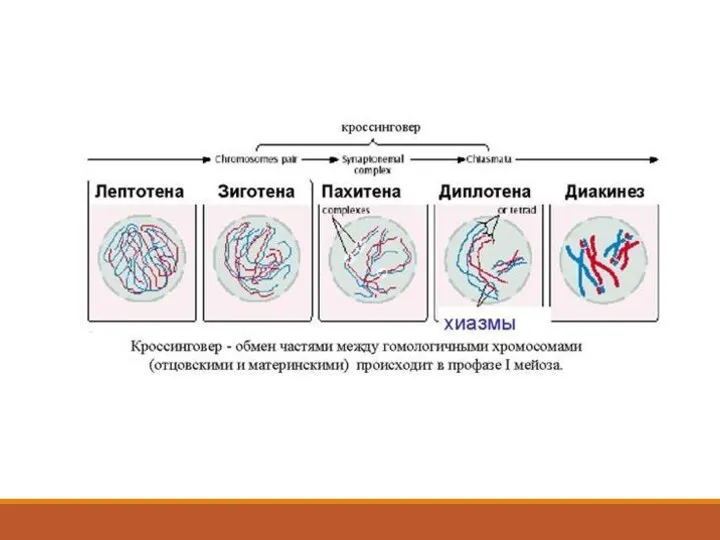
ЧАСТО ЯВЛЯЮТСЯ РЕЗУЛЬТАТОМ НАРУШЕНИЯ КРОССИНГОВЕРА – ОБМЕНА УЧАСТКАМИ МЕЖДУ ХРОМОСОМАМИ,
Хромосомные мутации
ЧАСТО ЯВЛЯЮТСЯ РЕЗУЛЬТАТОМ НАРУШЕНИЯ КРОССИНГОВЕРА – ОБМЕНА УЧАСТКАМИ МЕЖДУ ХРОМОСОМАМИ,

Внутрихромосомные перестройки
Делеция
Дупликация
Инверсия
Кольцевая хромосомам
Изохромосома
Внутрихромосомные перестройки
Делеция
Дупликация
Инверсия
Кольцевая хромосомам
Изохромосома
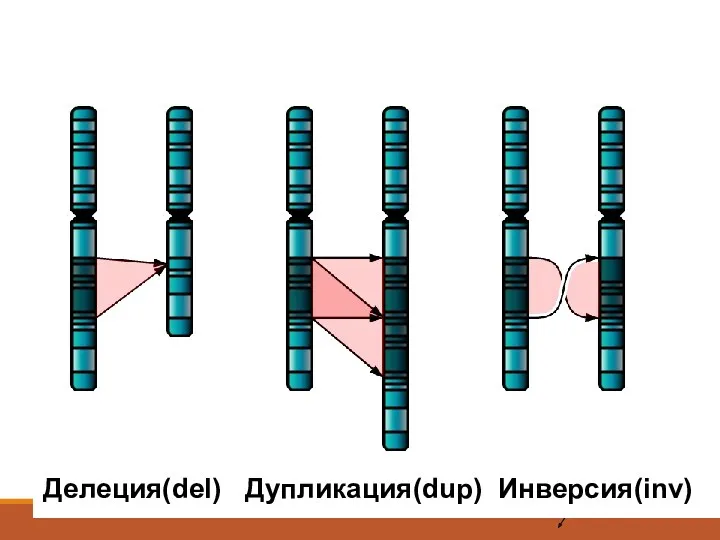
Делеция(del) Дупликация(dup) Инверсия(inv)
Делеция(del) Дупликация(dup) Инверсия(inv)
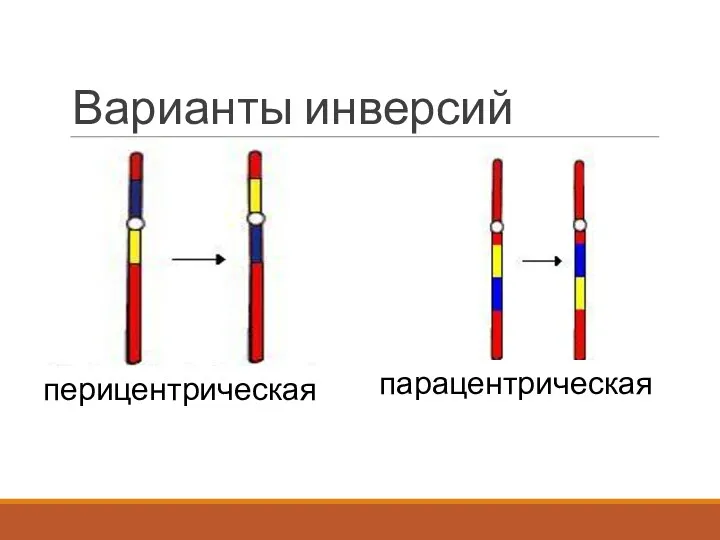
Варианты инверсий
перицентрическая
парацентрическая
Варианты инверсий
перицентрическая
парацентрическая
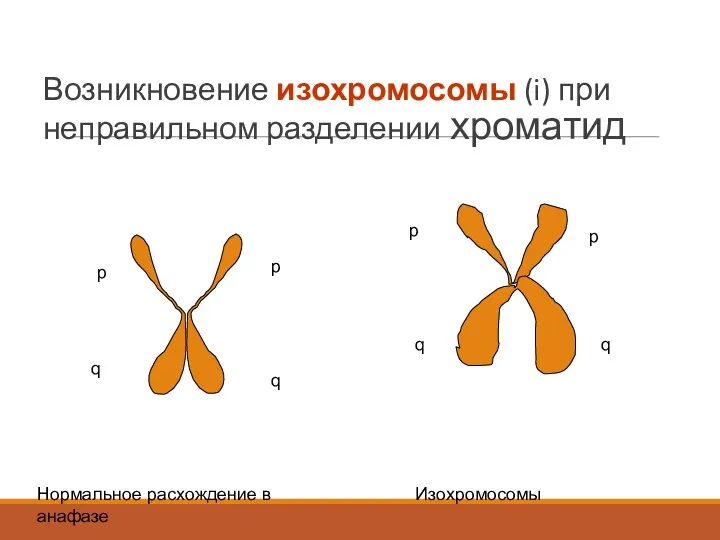
Возникновение изохромосомы (i) при неправильном разделении хроматид
p
q
p
q
p
q
p
q
Нормальное расхождение в анафазе
Изохромосомы
Возникновение изохромосомы (i) при неправильном разделении хроматид
p
q
p
q
p
q
p
q
Нормальное расхождение в анафазе
Изохромосомы
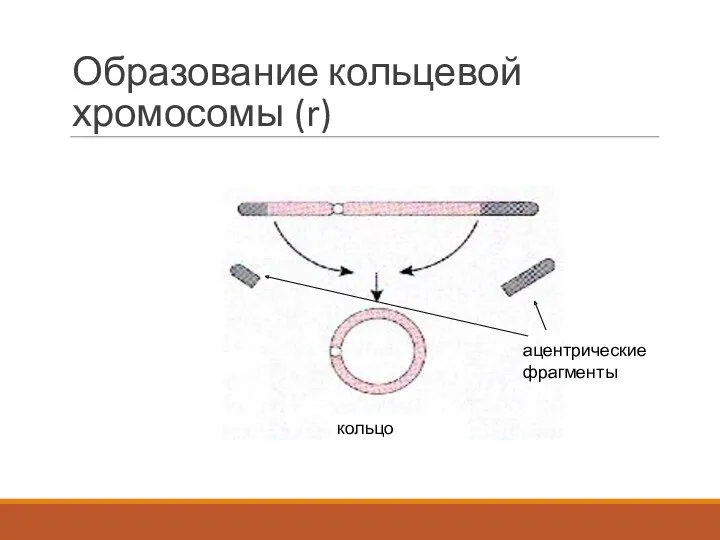
Образование кольцевой хромосомы (r)
кольцо
ацентрические фрагменты
Образование кольцевой хромосомы (r)
кольцо
ацентрические фрагменты

Межхромосомные перестройки - транслокации
Взаимные
Невзаимные
Робертсоновские
Межхромосомные перестройки - транслокации
Взаимные
Невзаимные
Робертсоновские
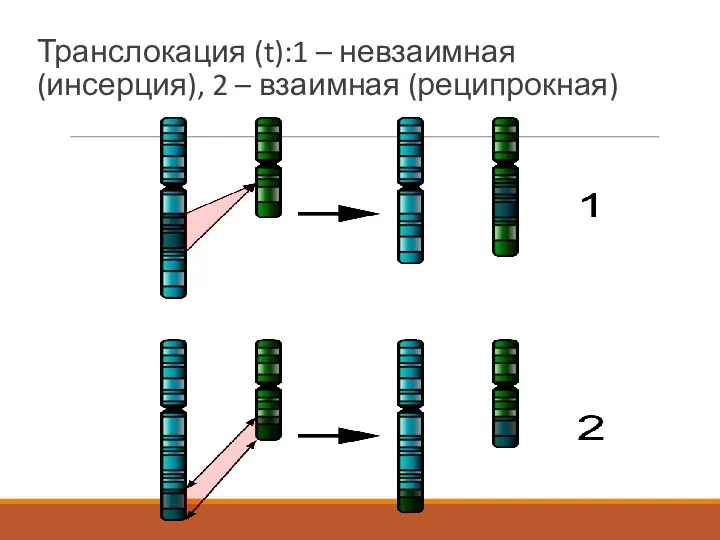
Транслокация (t):1 – невзаимная (инсерция), 2 – взаимная (реципрокная)
Транслокация (t):1 – невзаимная (инсерция), 2 – взаимная (реципрокная)
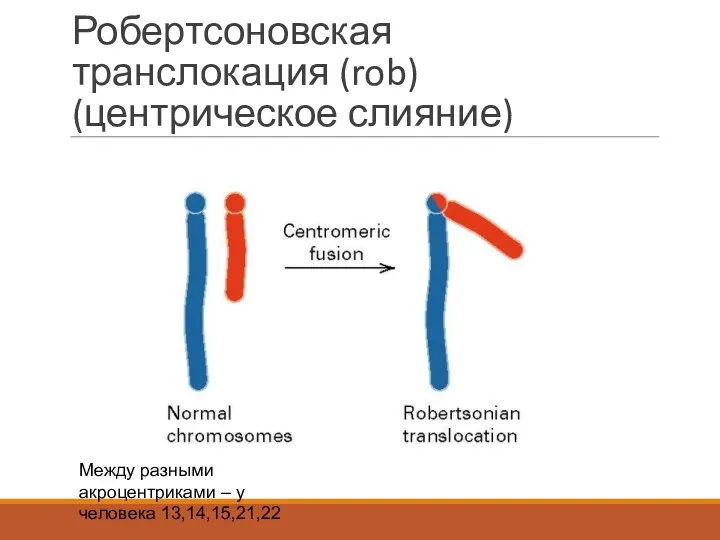
Робертсоновская транслокация (rob) (центрическое слияние)
Между разными акроцентриками – у человека 13,14,15,21,22
Робертсоновская транслокация (rob) (центрическое слияние)
Между разными акроцентриками – у человека 13,14,15,21,22
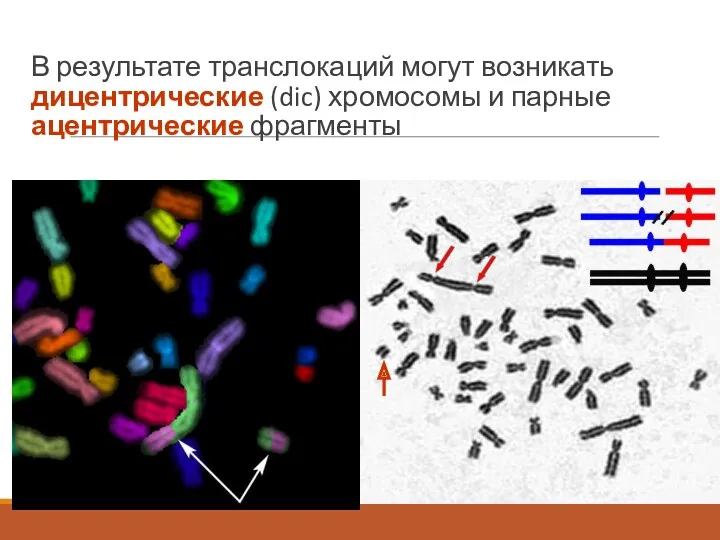
В результате транслокаций могут возникать дицентрические (dic) хромосомы и парные ацентрические
В результате транслокаций могут возникать дицентрические (dic) хромосомы и парные ацентрические

Кроме того, хромосомные мутации бывают:
Спонтанные/индуцированные
Соматические /генеративные
Вредные/полезные/нейтральные
Кроме того, хромосомные мутации бывают:
Спонтанные/индуцированные
Соматические /генеративные
Вредные/полезные/нейтральные

С клинической точки зрения хромосомные мутации удобнее делить на
Сбалансированные (нет потери
С клинической точки зрения хромосомные мутации удобнее делить на
Сбалансированные (нет потери

Значение хромосомных мутаций
Материал для эволюции, способствует появлению новых видов
Патология у человека
Значение хромосомных мутаций
Материал для эволюции, способствует появлению новых видов
Патология у человека

Сравнение хромосомных наборов человека (слева) и шимпанзе (справа). Видно, что наша
Сравнение хромосомных наборов человека (слева) и шимпанзе (справа). Видно, что наша

При лейкозах выявляются множественные хромосомные перестройки
При лейкозах выявляются множественные хромосомные перестройки
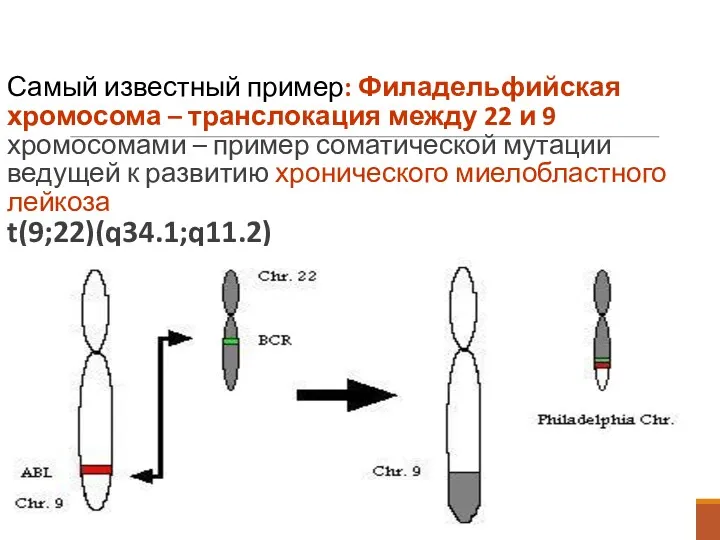
Самый известный пример: Филадельфийская хромосома – транслокация между 22 и 9
Самый известный пример: Филадельфийская хромосома – транслокация между 22 и 9
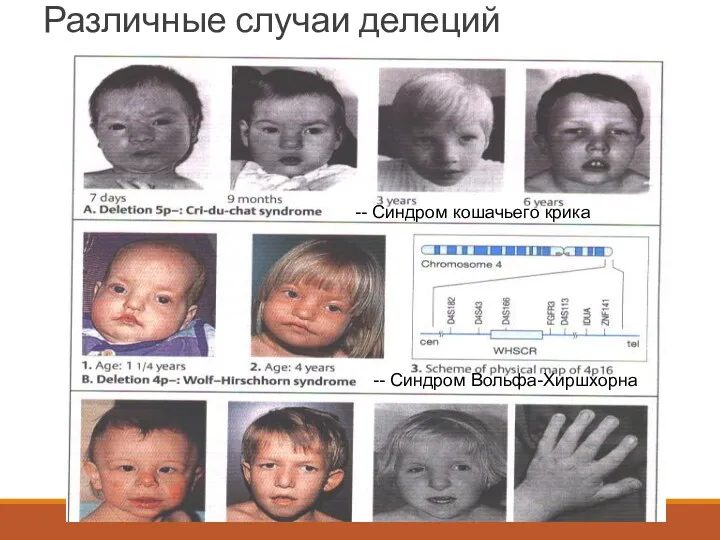
Различные случаи делеций
-- Синдром кошачьего крика
-- Синдром Вольфа-Хиршхорна
Различные случаи делеций
-- Синдром кошачьего крика
-- Синдром Вольфа-Хиршхорна
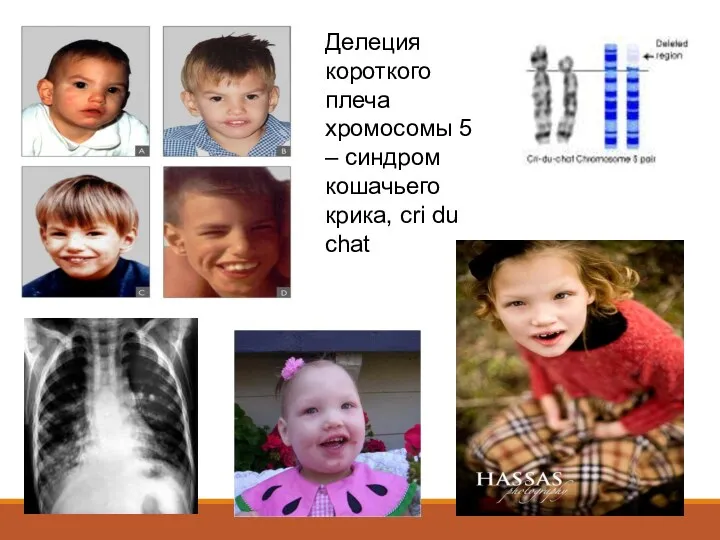
Делеция короткого плеча хромосомы 5 – синдром кошачьего крика, cri du
Делеция короткого плеча хромосомы 5 – синдром кошачьего крика, cri du

Хромосомные карты
Генетические – где лежит какой ген
Цитологические – по окраске
Физические
Хромосомные карты
Генетические – где лежит какой ген
Цитологические – по окраске
Физические

Карта хромосомы 9
Карта хромосомы 9
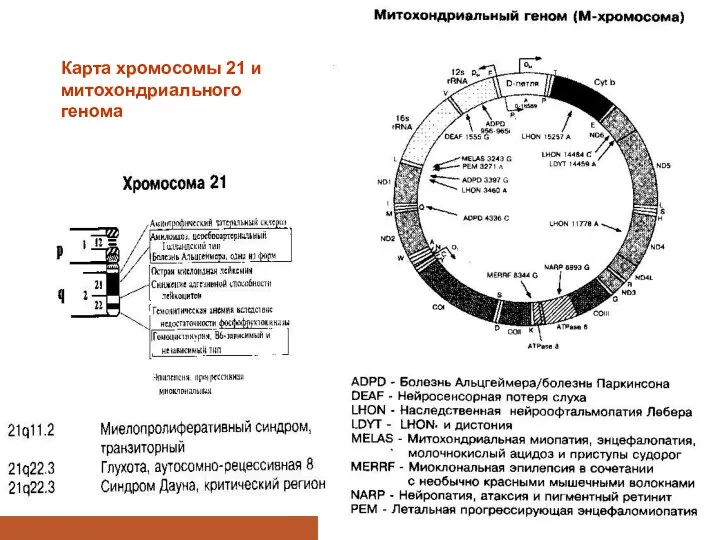
Карта хромосомы 21 и митохондриального генома
Карта хромосомы 21 и митохондриального генома

24-цветная FISH хромосом человека:
a - метафазная пластинка (Рубцов Н. Б.,
24-цветная FISH хромосом человека: a - метафазная пластинка (Рубцов Н. Б.,

24-цветная FISH хромосом человека:
b - pаскладка хромосом.
(Рубцов Н. Б.,
24-цветная FISH хромосом человека: b - pаскладка хромосом. (Рубцов Н. Б.,

ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА

Идиограмма
Идиограмма схематическое изображение гаплоидного набора хромосом организма, которые располагают в
Идиограмма
Идиограмма схематическое изображение гаплоидного набора хромосом организма, которые располагают в

В митотической хромосоме
ДНК упакована
в 10000 раз
В митотической хромосоме
ДНК упакована
в 10000 раз
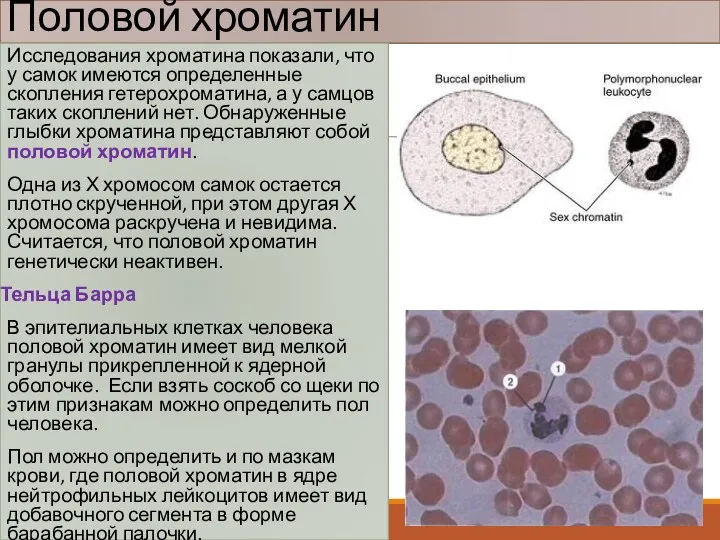
Половой хроматин
Исследования хроматина показали, что у самок имеются определенные скопления гетерохроматина,
Половой хроматин
Исследования хроматина показали, что у самок имеются определенные скопления гетерохроматина,

Структура и функция ядрышек. Гранулярный и фибриллярный компоненты ядрышек. Фибриллярные центры
Структура и функция ядрышек. Гранулярный и фибриллярный компоненты ядрышек. Фибриллярные центры
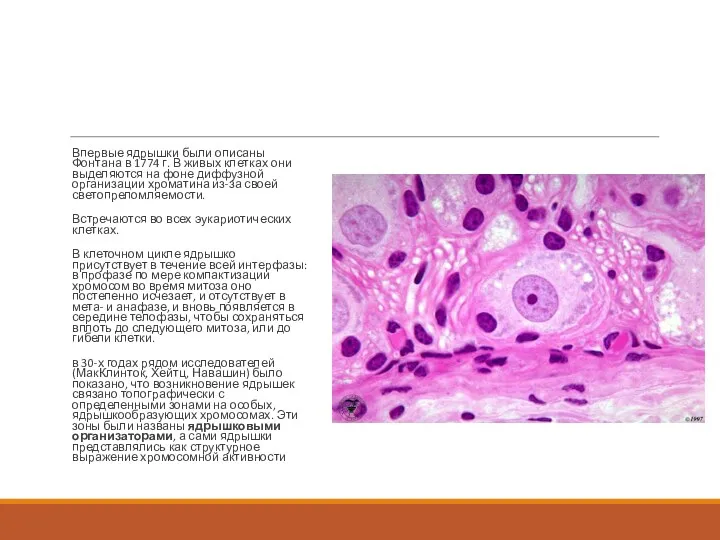
Впервые ядрышки были описаны Фонтана в 1774 г. В живых клетках
Впервые ядрышки были описаны Фонтана в 1774 г. В живых клетках

По данным цитохимических и биохимических исследований основным компонентом ядрышка является белок:
По данным цитохимических и биохимических исследований основным компонентом ядрышка является белок:

Встречаются во всех эукариотических клетках.
К таким исключениям относятся клетки дробящихся яиц,
К таким исключениям относятся клетки дробящихся яиц,

Число и размеры ядрышек в клетках непостоянны.
Локализация ядрышковых организаторов определяется довольно
Число и размеры ядрышек в клетках непостоянны.
Локализация ядрышковых организаторов определяется довольно

амплификации генов рРНК
амплификация рДНК, происходит в профазе I деления созревания, когда
амплификации генов рРНК
амплификация рДНК, происходит в профазе I деления созревания, когда

Биологический смысл появления сверхчисленных экстрахромосомных ядрышек при росте ооцитов совершенно понятен:
Биологический смысл появления сверхчисленных экстрахромосомных ядрышек при росте ооцитов совершенно понятен:

Структура ядрышка
Структура ядрышка

Структурные типы ядрышек
Структурные типы ядрышек
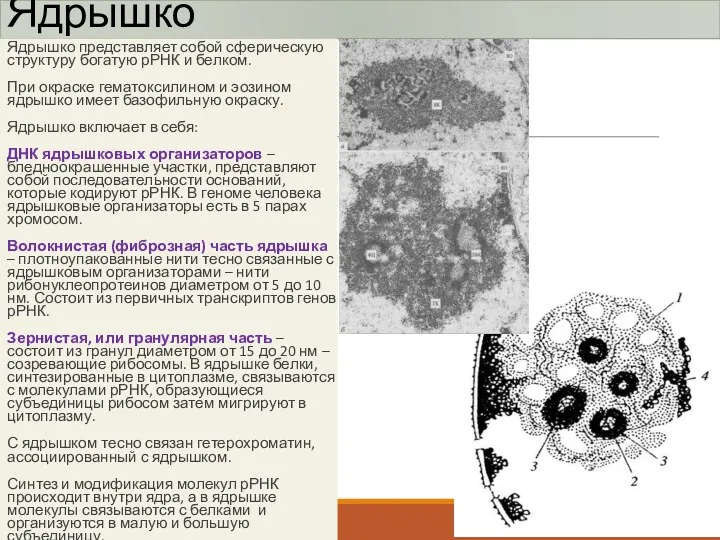
Ядрышко
Ядрышко представляет собой сферическую структуру богатую рРНК и белком.
При окраске гематоксилином
Ядрышко
Ядрышко представляет собой сферическую структуру богатую рРНК и белком.
При окраске гематоксилином

Клеточный цикл, Митоз, Мейоз
Клеточный цикл, Митоз, Мейоз
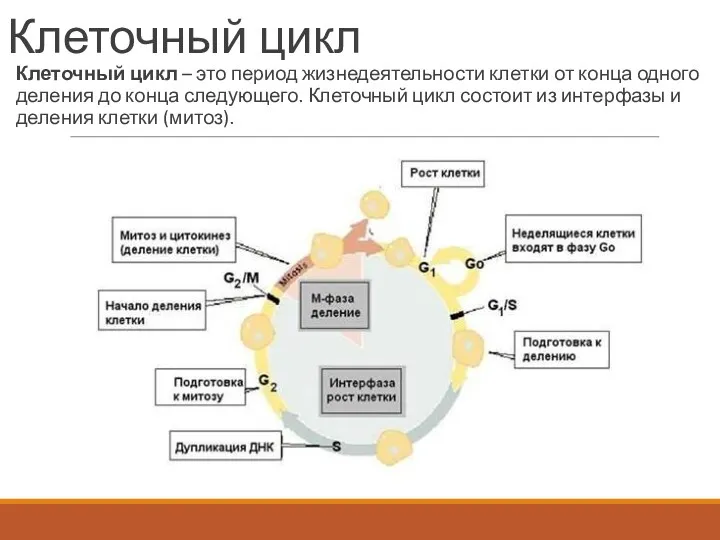
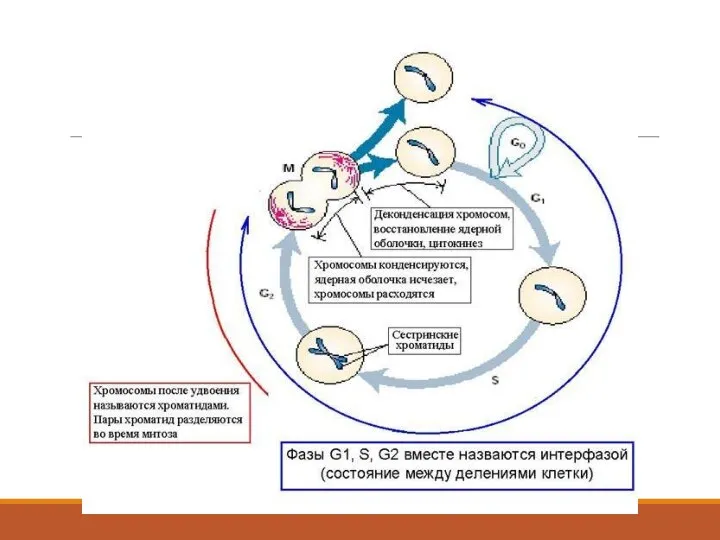
Клеточный цикл
Клеточный цикл – это период жизнедеятельности клетки от конца одного
Клеточный цикл
Клеточный цикл – это период жизнедеятельности клетки от конца одного
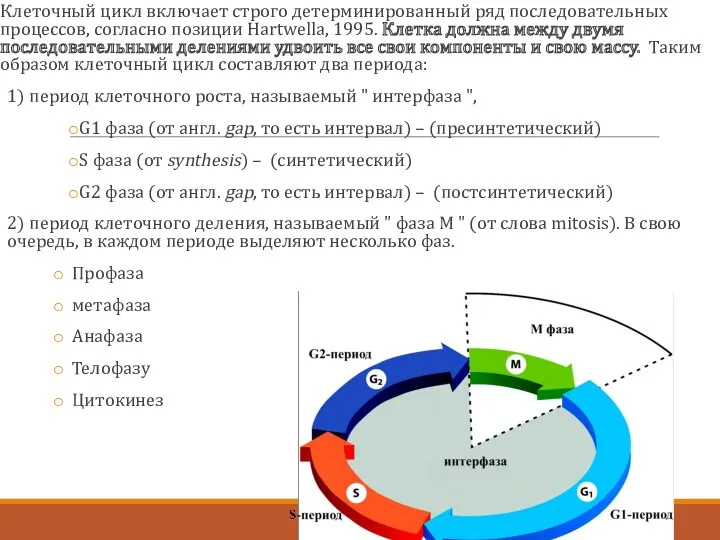
Клеточный цикл включает строго детерминированный ряд последовательных процессов, согласно позиции Hartwellа,
Клеточный цикл включает строго детерминированный ряд последовательных процессов, согласно позиции Hartwellа,

Интерфаза: период G1 (пресинтетический)
Пресинтетический период G1 наступает сразу после деления клетки. В этом
Интерфаза: период G1 (пресинтетический)
Пресинтетический период G1 наступает сразу после деления клетки. В этом
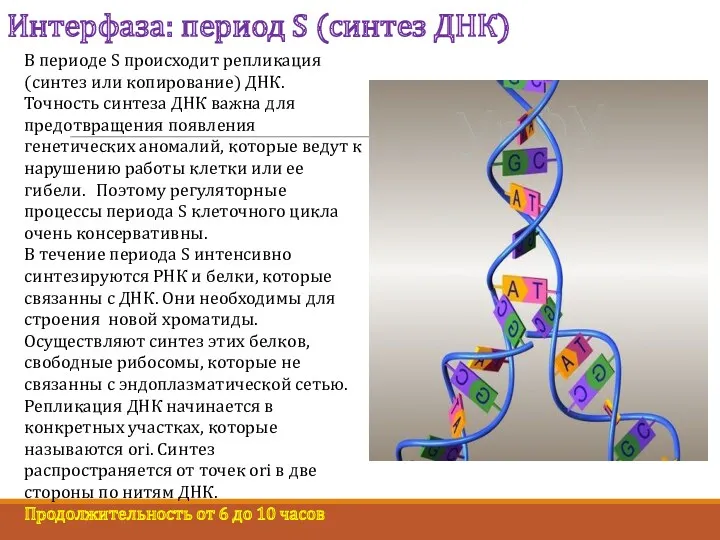
Интерфаза: период S (синтез ДНК)
В периоде S происходит репликация (синтез или
Интерфаза: период S (синтез ДНК)
В периоде S происходит репликация (синтез или
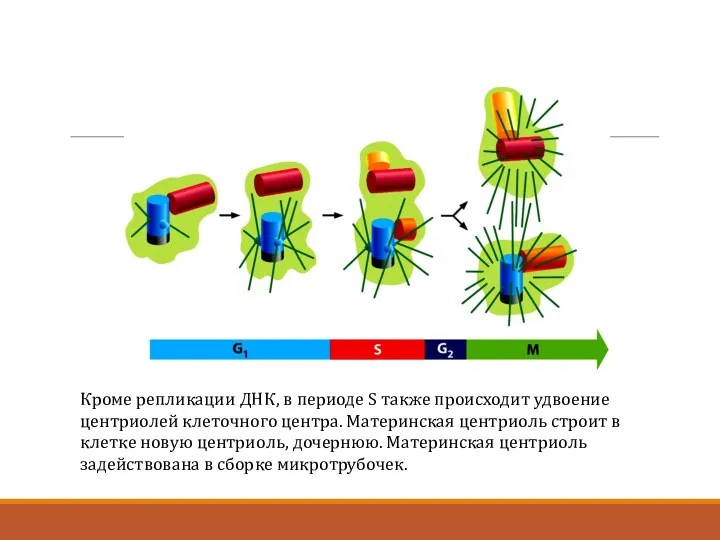
Кроме репликации ДНК, в периоде S также происходит удвоение центриолей клеточного
Кроме репликации ДНК, в периоде S также происходит удвоение центриолей клеточного
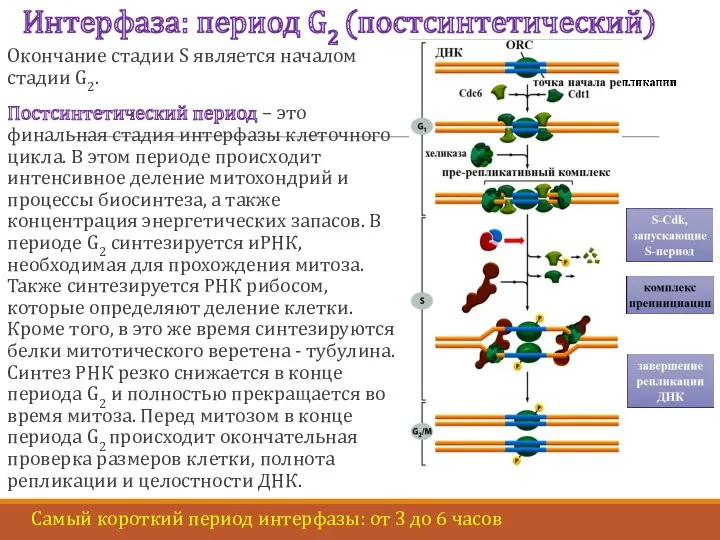
Интерфаза: период G2 (постсинтетический)
Окончание стадии S является началом стадии G2.
Постсинтетический период
Интерфаза: период G2 (постсинтетический)
Окончание стадии S является началом стадии G2.
Постсинтетический период

Фаза репликации количества ДНК
(S –фаза)
При подготовке к делению происходит удвоение
Фаза репликации количества ДНК
(S –фаза)
При подготовке к делению происходит удвоение

Постсинтетический период
(G 2 фаза)
В этот период происходит активное накопление энергии
Постсинтетический период
(G 2 фаза)
В этот период происходит активное накопление энергии

Митоз
Митоз (от греч. mitos - нить) - деление ядра, следующее за
Митоз
Митоз (от греч. mitos - нить) - деление ядра, следующее за

Профаза
В начале профазы многочисленные цитоплазматические микротрубочки, входящие в состав цитоскелета, распадаются;
Профаза
В начале профазы многочисленные цитоплазматические микротрубочки, входящие в состав цитоскелета, распадаются;

Метафаза
Эта стадия митоза часто продолжается длительное время Все хромосомы располагаются таким
Метафаза
Эта стадия митоза часто продолжается длительное время Все хромосомы располагаются таким
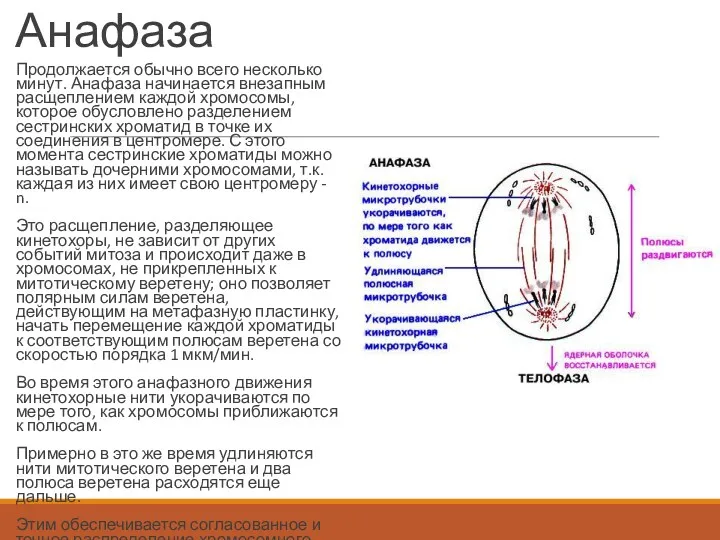
Анафаза
Продолжается обычно всего несколько минут. Анафаза начинается внезапным расщеплением каждой хромосомы,
Анафаза
Продолжается обычно всего несколько минут. Анафаза начинается внезапным расщеплением каждой хромосомы,
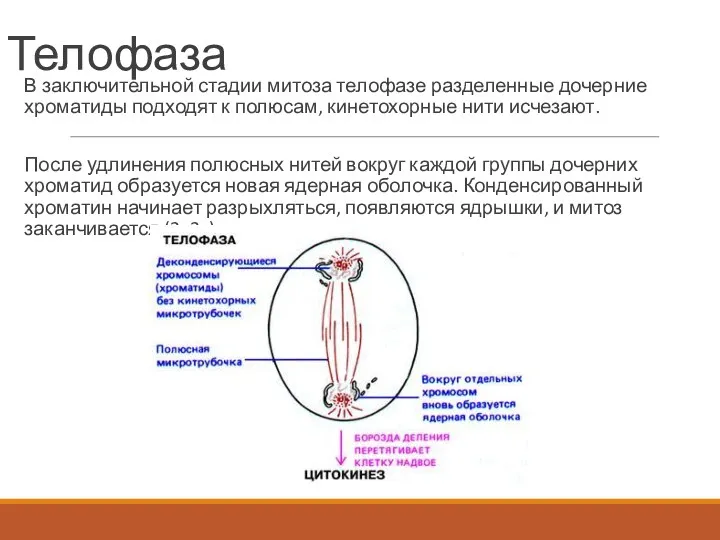
Телофаза
В заключительной стадии митоза телофазе разделенные дочерние хроматиды подходят к полюсам,
Телофаза
В заключительной стадии митоза телофазе разделенные дочерние хроматиды подходят к полюсам,
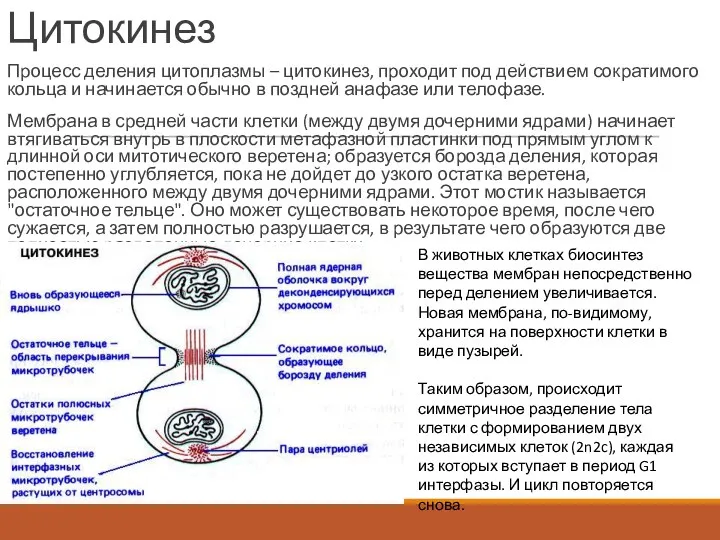
Цитокинез
Процесс деления цитоплазмы – цитокинез, проходит под действием сократимого кольца и
Цитокинез
Процесс деления цитоплазмы – цитокинез, проходит под действием сократимого кольца и

Мейоз
При половом размножении растений и животных (в том числе и человека)
Мейоз
При половом размножении растений и животных (в том числе и человека)

При первом делении мейоза каждая дочерняя клетка наследует две копии одного
При первом делении мейоза каждая дочерняя клетка наследует две копии одного
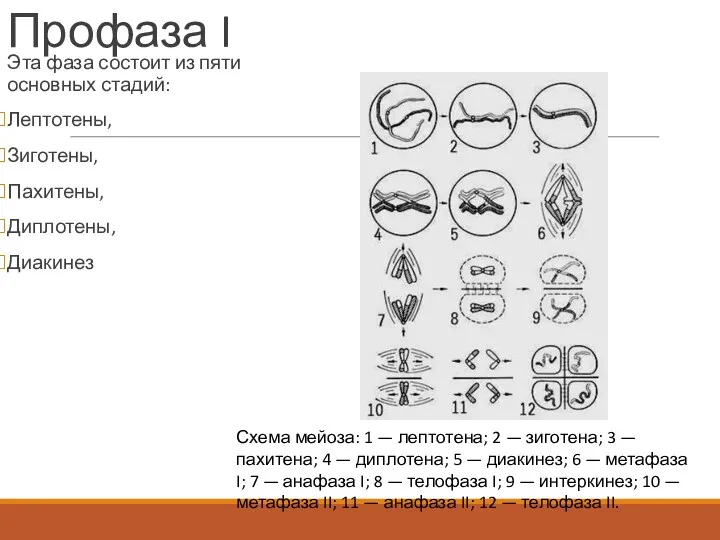
Профаза I
Эта фаза состоит из пяти основных стадий:
Лептотены,
Зиготены,
Пахитены,
Профаза I
Эта фаза состоит из пяти основных стадий:
Лептотены,
Зиготены,
Пахитены,

Лептотена
Каждая хромосома, изменив свою интерфазную конформацию, переходит в конденсированную форму, образуя
Лептотена
Каждая хромосома, изменив свою интерфазную конформацию, переходит в конденсированную форму, образуя

Зиготена
Зиготена - стадия спаривания хромосом. Моментом перехода лептотены в зиготену считают
Зиготена
Зиготена - стадия спаривания хромосом. Моментом перехода лептотены в зиготену считают

Пахитена
Пахитена - стадия профазы мейоза, на которой спаривание гомологов завершено.
Хромосомы
Пахитена
Пахитена - стадия профазы мейоза, на которой спаривание гомологов завершено.
Хромосомы

Диплотена
Диплотена - стадия мейоза после пахитены и перед диакинезом. Стадия расхождения
Диплотена
Диплотена - стадия мейоза после пахитены и перед диакинезом. Стадия расхождения

Диакинез
Диакинез - заключительная стадия профазы I. В диакинезе биваленты резко укороченные,
Диакинез
Диакинез - заключительная стадия профазы I. В диакинезе биваленты резко укороченные,
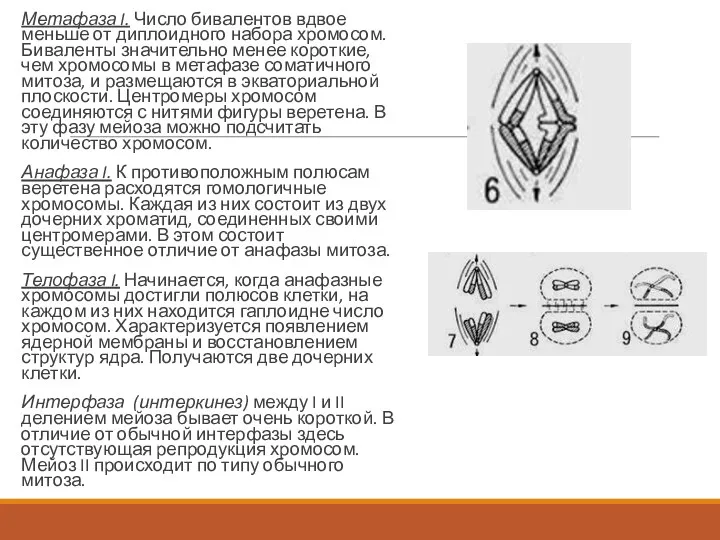
Метафаза I. Число бивалентов вдвое меньше от диплоидного набора хромосом. Биваленты
Метафаза I. Число бивалентов вдвое меньше от диплоидного набора хромосом. Биваленты
 Genetic-test. Биометрическое тестирование способностей человека по типам узоров на подушечках пальцев
Genetic-test. Биометрическое тестирование способностей человека по типам узоров на подушечках пальцев Терморегуляция. Обмен веществ
Терморегуляция. Обмен веществ Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость
Анатомия и физиология ЦНС. Свойства и принципы функционирования нервных центров. Строение СМ. Цереброспинальная жидкость Возникновение жизни на Земле
Возникновение жизни на Земле Лесные профессии
Лесные профессии Игра четвёртый лишний
Игра четвёртый лишний Размножение голосеменных растений
Размножение голосеменных растений Многообразие и значение грибов
Многообразие и значение грибов Парша яблони и меры борьбы с ней в условиях Уйгурского района Алматинской области
Парша яблони и меры борьбы с ней в условиях Уйгурского района Алматинской области Тяжёлые металлы
Тяжёлые металлы Сравнительная характеристика бесполого и полового размножения, их сущность и биологическое значение
Сравнительная характеристика бесполого и полового размножения, их сущность и биологическое значение Заключительная игра по теме: Пресмыкающиеся
Заключительная игра по теме: Пресмыкающиеся Презентация по биологии для учащихся 9 класса по теме Клеточное ядро
Презентация по биологии для учащихся 9 класса по теме Клеточное ядро Физиология и биохимия микроорганизмов
Физиология и биохимия микроорганизмов Шоколад. Плоды какао
Шоколад. Плоды какао Породистые и непородистые кошки
Породистые и непородистые кошки Немецкая овчарка. 3 класс
Немецкая овчарка. 3 класс Профильное обучение биологии
Профильное обучение биологии Түрлер арасындағы қарымқатынастың негізгі түрлері
Түрлер арасындағы қарымқатынастың негізгі түрлері Тип Хордовые (60 000 видов)
Тип Хордовые (60 000 видов) Опале листя - користь чи шкода
Опале листя - користь чи шкода Методы изолирования лекарственных соединений из биологических жидкостей при проведении ХТА с диагностической целью
Методы изолирования лекарственных соединений из биологических жидкостей при проведении ХТА с диагностической целью Царства живой природы
Царства живой природы Школьная клумба
Школьная клумба Тип Моллюски. Класс двустворчатые моллюски
Тип Моллюски. Класс двустворчатые моллюски Методы и условия культивирования тканей и клеток растений
Методы и условия культивирования тканей и клеток растений Исследование: Лечебные свойства лесных ягод (2 класс)
Исследование: Лечебные свойства лесных ягод (2 класс) Высокоэффективная биотехнологическая косметика для ухода за кожей лица и рук
Высокоэффективная биотехнологическая косметика для ухода за кожей лица и рук