- Строение и развитие женских половых клеток. Лекция 2

Содержание
- 2. В зрелом яйце сосредоточены ВСЕ материалы, необходимые для начальных стадий развития зародыша. Ультраструктура яйцеклетки цитоплазма содержит:
- 3. Оболочки яйцеклетки: первичная: производная плазмалеммы яйца. Иногда называется желточной. Присуща всем животным (кроме губок и стрекающих).
- 4. Стадии оогенеза размножение: ППК мигрируют из места возникновения в гонады, где пролиферируют, делятся митозом и называются
- 5. превителлогенез (малый или цитоплазматический рост): пропорциональное увеличение массы ядра и цитоплазмы при подготовке ооцита I порядка
- 6. Приурочены к достижению половой зрелости. Прогестерон фолликулярных клеток связывается с рецепторами плазмалеммы и посредством Gi-белка инактивирует
- 7. примордиальные фолликулы: расположены по периферии. Это ооцит I порядка, покрытый слоем внедряющихся в соединительно-тканную строму яичника
- 8. Граафов пузырек третичный фолликул (Граафов пузырек): фолликул на последней стадии резорбции, стенка которого (зернистая оболочка) состоит
- 9. овуляция: третичный (пузырчатый) фолликул, увеличиваясь, выпячивает поверхность яичника так, что яйценосный бугорок с ооцитом оказывается в
- 10. Схема развития фолликулов в яичнике Развитие фолликулов в яичнике человека (Алмазов, Сутулов, 1978)
- 11. Регуляция полового цикла млекопитающих Рост фолликулов начинается еще в яичниках зародыша. Первичный (малый) рост не зависит
- 12. фолликулиновая (эстрагенная или пролиферативная) фаза: растущие фолликулы секретируют все больше эстрогенов, вызывая пролиферацию эндометрия матки и
- 13. По количеству желтка: алецитальные: безжелтковые (плацентарные млекопитающие, первичнотрахейные); олиголецитальные: маложелтковые (черви, моллюски, иглокожие); мезолецитальные: среднее количество
- 14. Лишь немногие ооциты развиваются исключительно за счет эндогенного желтка. В зависимости от способа доставки к клетке
- 16. Скачать презентацию
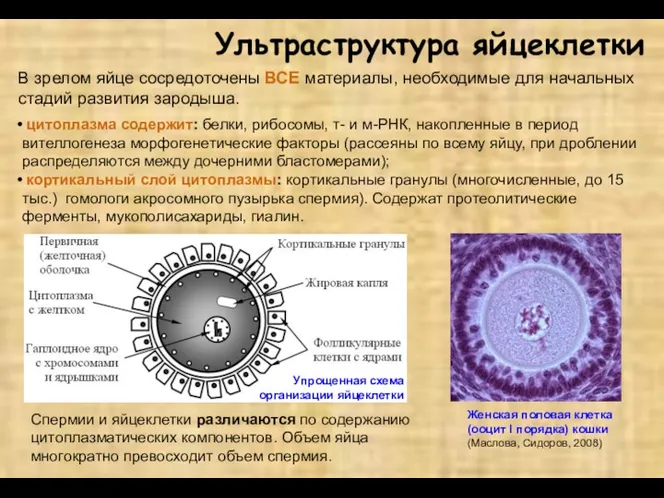
В зрелом яйце сосредоточены ВСЕ материалы, необходимые для начальных стадий развития
В зрелом яйце сосредоточены ВСЕ материалы, необходимые для начальных стадий развития

Оболочки яйцеклетки:
первичная: производная плазмалеммы яйца. Иногда называется желточной. Присуща всем
Оболочки яйцеклетки:
первичная: производная плазмалеммы яйца. Иногда называется желточной. Присуща всем
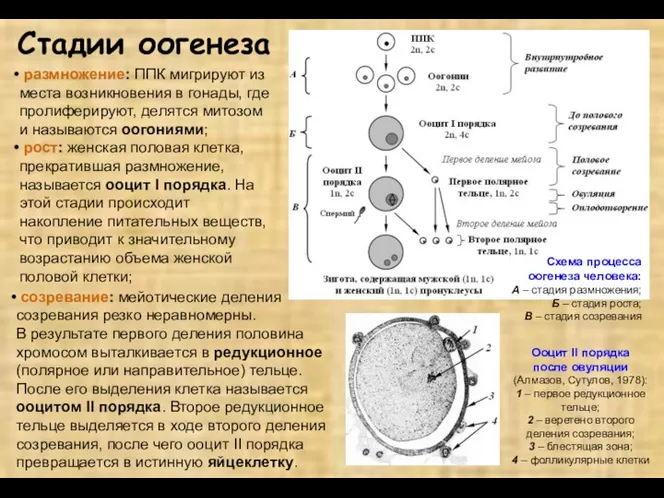
Стадии оогенеза
размножение: ППК мигрируют из места возникновения в гонады, где
Стадии оогенеза
размножение: ППК мигрируют из места возникновения в гонады, где

превителлогенез (малый или цитоплазматический рост): пропорциональное увеличение массы ядра и
превителлогенез (малый или цитоплазматический рост): пропорциональное увеличение массы ядра и

Приурочены к достижению половой зрелости. Прогестерон фолликулярных клеток связывается с
Приурочены к достижению половой зрелости. Прогестерон фолликулярных клеток связывается с
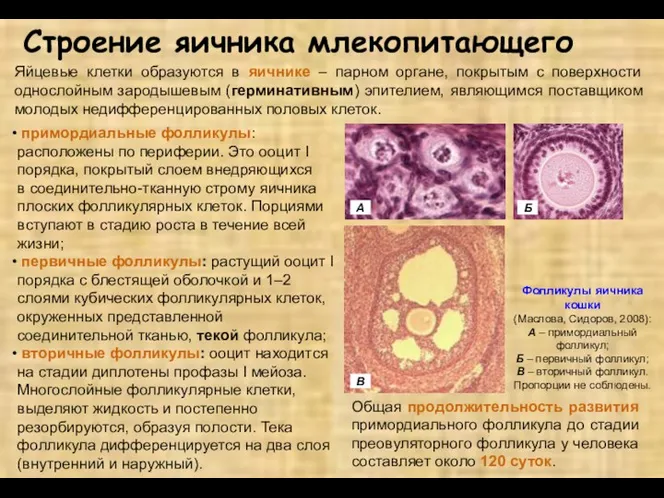
примордиальные фолликулы: расположены по периферии. Это ооцит I порядка, покрытый
примордиальные фолликулы: расположены по периферии. Это ооцит I порядка, покрытый
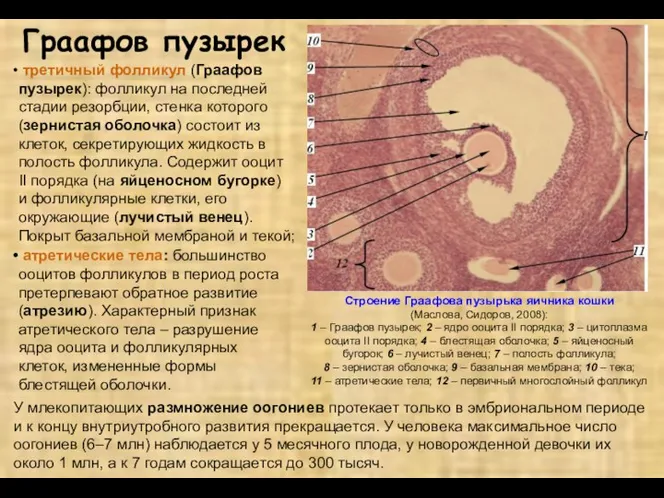
Граафов пузырек
третичный фолликул (Граафов пузырек): фолликул на последней стадии резорбции,
Граафов пузырек
третичный фолликул (Граафов пузырек): фолликул на последней стадии резорбции,

овуляция: третичный (пузырчатый) фолликул, увеличиваясь, выпячивает поверхность яичника так, что
овуляция: третичный (пузырчатый) фолликул, увеличиваясь, выпячивает поверхность яичника так, что
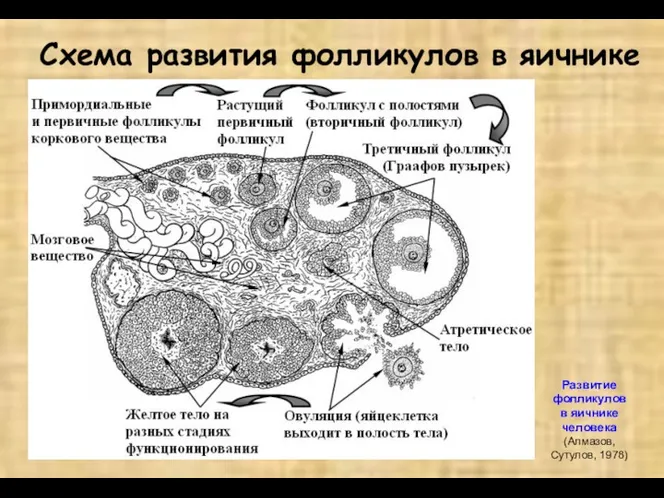
Схема развития фолликулов в яичнике
Развитие фолликулов
в яичнике человека
(Алмазов, Сутулов, 1978)
Схема развития фолликулов в яичнике
Развитие фолликулов
в яичнике человека
(Алмазов, Сутулов, 1978)
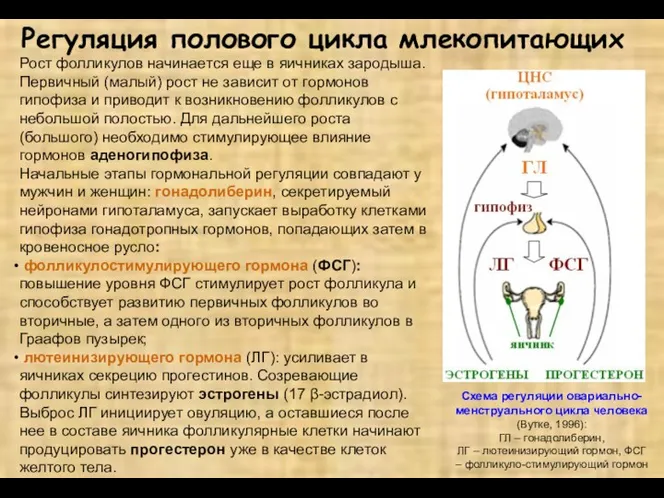
Регуляция полового цикла млекопитающих
Рост фолликулов начинается еще в яичниках зародыша. Первичный
Регуляция полового цикла млекопитающих
Рост фолликулов начинается еще в яичниках зародыша. Первичный
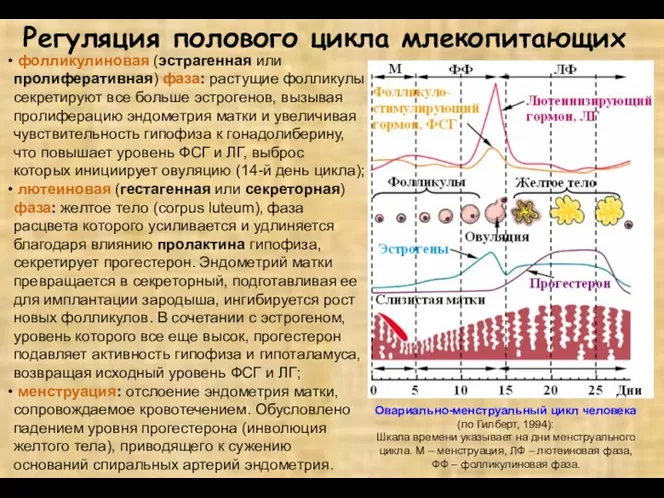
фолликулиновая (эстрагенная или пролиферативная) фаза: растущие фолликулы секретируют все больше
фолликулиновая (эстрагенная или пролиферативная) фаза: растущие фолликулы секретируют все больше

По количеству желтка:
алецитальные: безжелтковые (плацентарные млекопитающие, первичнотрахейные);
олиголецитальные: маложелтковые (черви,
По количеству желтка:
алецитальные: безжелтковые (плацентарные млекопитающие, первичнотрахейные);
олиголецитальные: маложелтковые (черви,

Лишь немногие ооциты развиваются исключительно за счет эндогенного желтка.
В зависимости от
Лишь немногие ооциты развиваются исключительно за счет эндогенного желтка.
В зависимости от
 Энергетический обмен углеводов. 10 класс
Энергетический обмен углеводов. 10 класс Дыхание растений, бактерий и грибов
Дыхание растений, бактерий и грибов Теоретичні основи рослинництва
Теоретичні основи рослинництва Bacillus. General Characteristics of Bacillus
Bacillus. General Characteristics of Bacillus Методи обробки результатів медико-біологічних досліджень у фізичній культурі та спорті
Методи обробки результатів медико-біологічних досліджень у фізичній культурі та спорті Многообразие живых организмов
Многообразие живых организмов Основные систематические группы рыб. 7 класс
Основные систематические группы рыб. 7 класс Насекомые
Насекомые Птицы Ульяновской области
Птицы Ульяновской области Клетки кожицы лука
Клетки кожицы лука Загальна характеристика гормонів
Загальна характеристика гормонів Животные зооуголка: дегу
Животные зооуголка: дегу Внешнее строение листа. Жилкование
Внешнее строение листа. Жилкование Вплив іонізуючого випромінювання на живі підсистеми
Вплив іонізуючого випромінювання на живі підсистеми Клеточное строение листа
Клеточное строение листа Значение фтора для организма человека
Значение фтора для организма человека презентации по теме Органичекие вещества, входящие в состав клетки
презентации по теме Органичекие вещества, входящие в состав клетки Профильное обучение биологии
Профильное обучение биологии Методы выделения чистых культур бактерий. Питание микроорганизмов. Методы стерилизации и дезинфекции
Методы выделения чистых культур бактерий. Питание микроорганизмов. Методы стерилизации и дезинфекции Урок по теме Органы кровообращения
Урок по теме Органы кровообращения Диазины. Характеристика диазинов. (Лекция 8)
Диазины. Характеристика диазинов. (Лекция 8) Молекулярно-генетические механизмы. Постэмбриональный онтогенез
Молекулярно-генетические механизмы. Постэмбриональный онтогенез Правила создания цветников
Правила создания цветников Үлкен Панда
Үлкен Панда Генетика. Спадко́вість
Генетика. Спадко́вість Ёлочка, живи! Природоохранная акция
Ёлочка, живи! Природоохранная акция Endangered species
Endangered species Эволюция. Видообразование. Процесс образования новых видов
Эволюция. Видообразование. Процесс образования новых видов