- Строение коры больших полушарий

Содержание
- 2. Кора больших полушарий представляет собой скопление нейронов и глиальных клеток. Толщина коры составляет от 1,2 до
- 3. Основная часть коры большого мозга (95,9% всей поверхности полушарий) представляет собой неокортекс – новую кору. Филогенетически
- 4. Схема классификации коры
- 5. Древняя и старая кора в филогенезе позвоночных появляются раньше и несут в себе черты относительно примитивного
- 6. Микроскопическое строение новой коры Новая кора. Все области новой коры построены по единому принципу. Исходным типом
- 7. III слой – пирамидный, толщиной около 1 мм, состоит из мелких и средних пирамидных клеток. Типичный
- 8. IV внутренний зернистый. Он состоит из звездчатых клеток имеющих короткие отростки, и малых пирамид. Дендриты клеток
- 9. V слой ганглионарный или внутренний пирамидный. Самые крупные клетки коры расположены именно в этом слое (гигантские
- 10. VI слой коры - полиморфный. Содержит клетки разнообразной формы и размера. Имеет толщину от 0,1 до
- 11. Микроскопическое строение коры больших полушарий: I – молекулярный слой; II – наружный зернистый слой; III –
- 12. Послойное расположение нейронов в коре называется цитоархитектоникой. Цитоахитектонические особенности позволили разделить всю поверхность коры на одиннадцать
- 13. Цитоархитектонические поля коры больших полушарий (по К. Бродману)
- 14. Каждое поле коры выполняет определенную функцию. Часть полей коры является сенсорными, в первичных сенсорных полях заканчиваются
- 15. В медиальной части представлены нижние конечности, а наиболее низко на латеральной части извилины расположены проекции рецепторных
- 16. Представительство различных мышечных групп в коре больших полушарий человека («человечек Пенфилда)
- 17. Поля 17 – 19 расположены в затылочной доле, являются зрительным центром коры; 17 поле, занимающее сам
- 18. Модульная организация коры больших полушарий Изучение нейронной организации коры большого мозга позволило в начале XX века
- 19. Нейро-глио-сосудистый ансамбль (модуль) коры больших полушарий
- 20. С модулем связан определенный набор афферентных волокон, приносящих информацию, которую он подвергает стандартной обработке, а также
- 21. Распределение нейронов по слоям в пределах коркового модуля
- 22. Филогенез коры больших полушарий У низших позвоночных (круглоротые, рыбы) развитие конечного мозга идет по пути утолщения
- 23. Усложнение организации конечного мозга у млекопитающих происходит за счет развития мантии. Мантия покрыта корой. Кроме древней
- 24. Онтогенез коры больших полушарий Передний мозг у человека является закругленным концом нервной трубки; на этом этапе
- 25. Примерно на 8-ой неделе эмбрионального развития растущие аксоны покидают кору, огибая снизу средний мозг, образуя ножки
- 26. Мозолистое тело появляется в начале 4-го месяца развития и растет очень быстро. Пузыри конечного мозга разрастаются:
- 27. Этапы развития головного мозга человека
- 29. Скачать презентацию

Кора больших полушарий представляет собой скопление нейронов и глиальных клеток.
Толщина коры
Кора больших полушарий представляет собой скопление нейронов и глиальных клеток.
Толщина коры

Основная часть коры большого мозга (95,9% всей поверхности полушарий) представляет собой
Основная часть коры большого мозга (95,9% всей поверхности полушарий) представляет собой
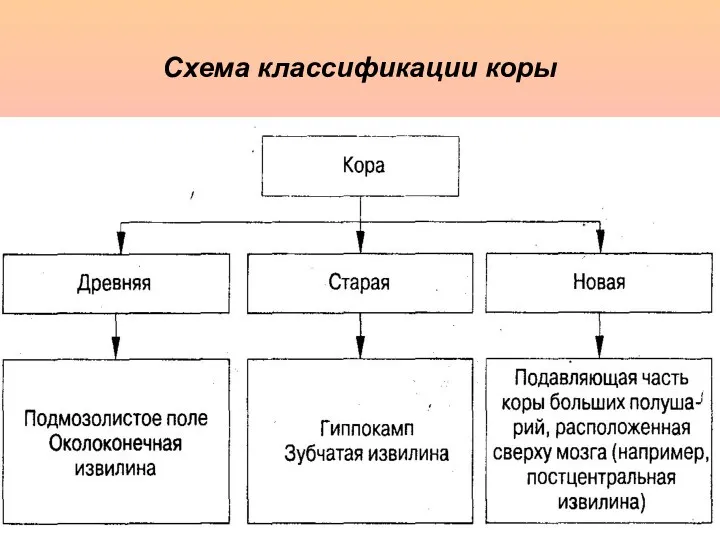
Схема классификации коры
Схема классификации коры

Древняя и старая кора в филогенезе позвоночных появляются раньше и несут
Древняя и старая кора в филогенезе позвоночных появляются раньше и несут

Микроскопическое строение
новой коры
Новая кора. Все области новой коры построены
Микроскопическое строение
новой коры
Новая кора. Все области новой коры построены
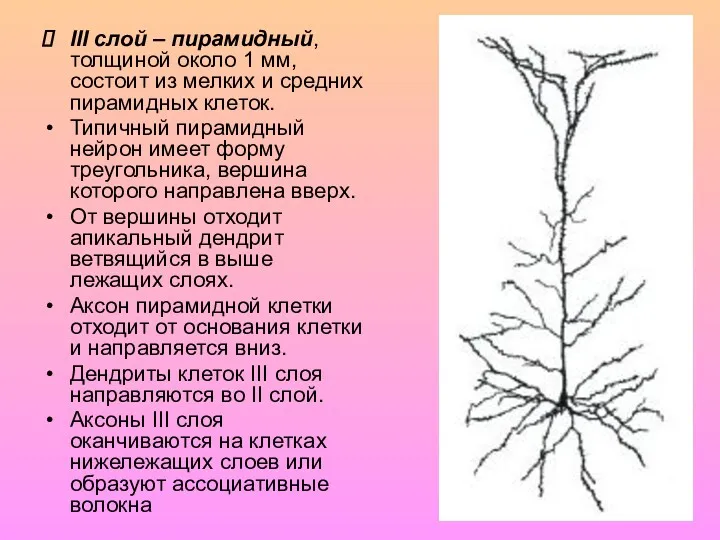
III слой – пирамидный, толщиной около 1 мм, состоит из мелких
III слой – пирамидный, толщиной около 1 мм, состоит из мелких

IV внутренний зернистый.
Он состоит из звездчатых клеток имеющих короткие отростки,
IV внутренний зернистый.
Он состоит из звездчатых клеток имеющих короткие отростки,

V слой ганглионарный или внутренний пирамидный.
Самые крупные клетки коры расположены именно
V слой ганглионарный или внутренний пирамидный.
Самые крупные клетки коры расположены именно

VI слой коры - полиморфный. Содержит клетки разнообразной формы и размера.
VI слой коры - полиморфный. Содержит клетки разнообразной формы и размера.
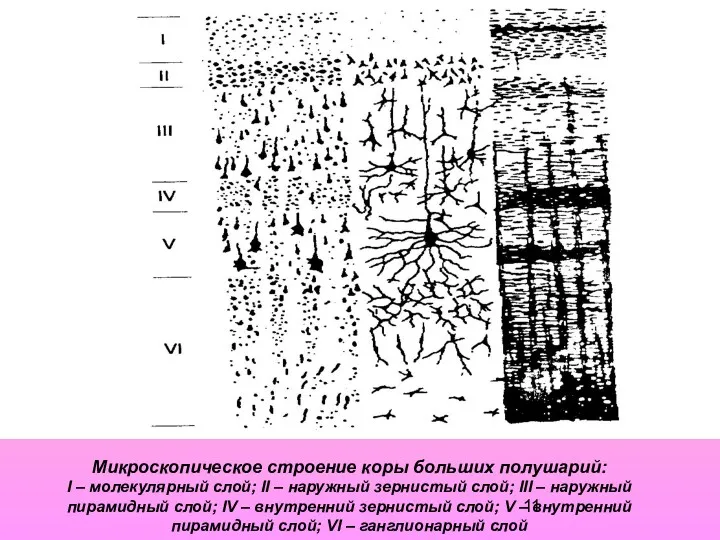
Микроскопическое строение коры больших полушарий:
I – молекулярный слой; II –
Микроскопическое строение коры больших полушарий: I – молекулярный слой; II –

Послойное расположение нейронов в коре называется цитоархитектоникой.
Цитоахитектонические особенности позволили разделить всю
Послойное расположение нейронов в коре называется цитоархитектоникой.
Цитоахитектонические особенности позволили разделить всю
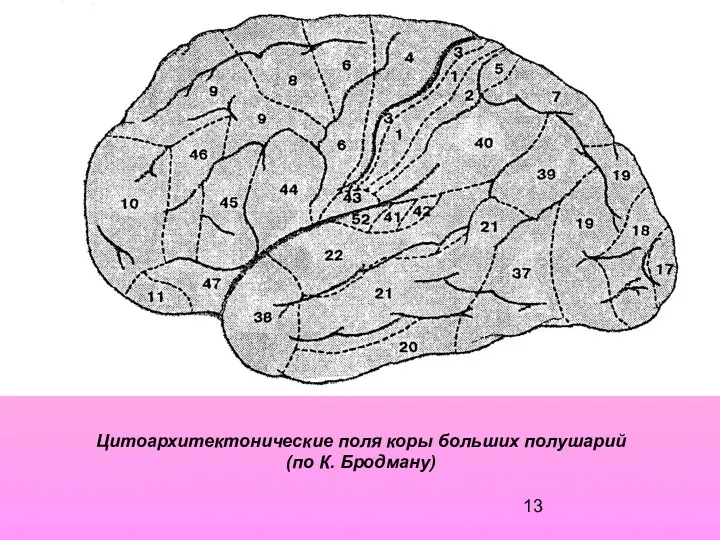
Цитоархитектонические поля коры больших полушарий
(по К. Бродману)
Цитоархитектонические поля коры больших полушарий
(по К. Бродману)

Каждое поле коры выполняет определенную функцию.
Часть полей коры является сенсорными,
Каждое поле коры выполняет определенную функцию.
Часть полей коры является сенсорными,

В медиальной части представлены нижние конечности, а наиболее низко на латеральной
В медиальной части представлены нижние конечности, а наиболее низко на латеральной
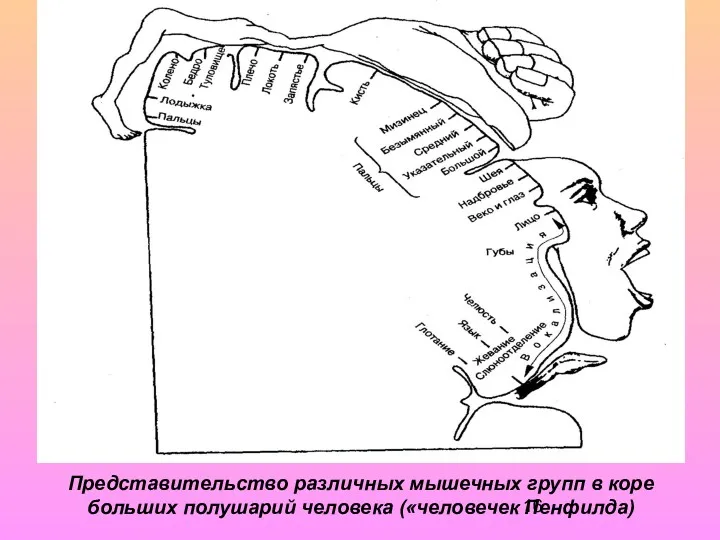
Представительство различных мышечных групп в коре больших полушарий человека («человечек Пенфилда)
Представительство различных мышечных групп в коре больших полушарий человека («человечек Пенфилда)

Поля 17 – 19 расположены в затылочной доле, являются зрительным центром
Поля 17 – 19 расположены в затылочной доле, являются зрительным центром

Модульная организация коры больших полушарий
Изучение нейронной организации коры большого мозга позволило
Модульная организация коры больших полушарий
Изучение нейронной организации коры большого мозга позволило
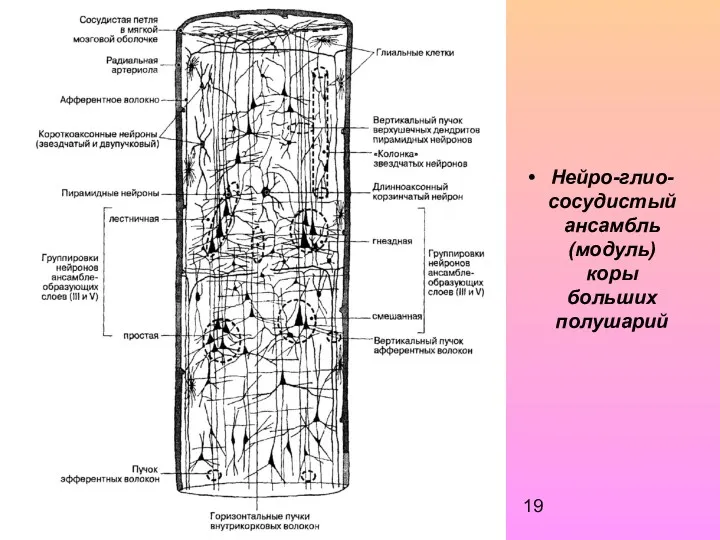
Нейро-глио-сосудистый ансамбль (модуль) коры больших полушарий
Нейро-глио-сосудистый ансамбль (модуль) коры больших полушарий

С модулем связан определенный набор афферентных волокон, приносящих информацию, которую он
С модулем связан определенный набор афферентных волокон, приносящих информацию, которую он
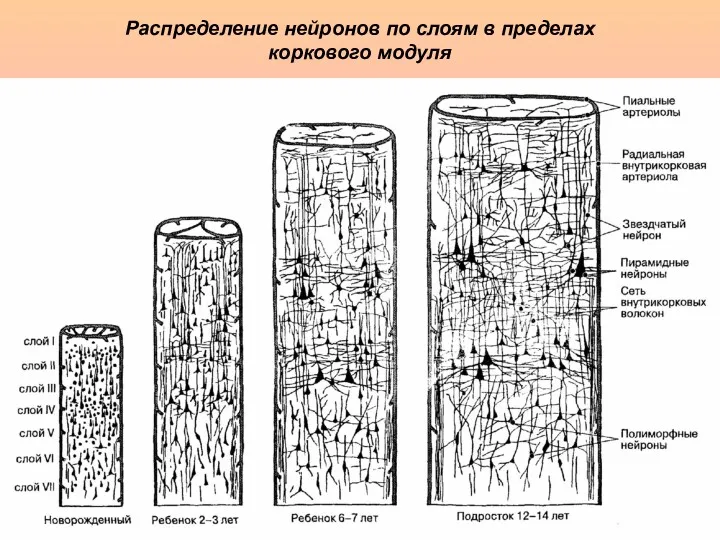
Распределение нейронов по слоям в пределах
коркового модуля
Распределение нейронов по слоям в пределах
коркового модуля

Филогенез коры больших полушарий
У низших позвоночных (круглоротые, рыбы) развитие конечного мозга
Филогенез коры больших полушарий
У низших позвоночных (круглоротые, рыбы) развитие конечного мозга

Усложнение организации конечного мозга у млекопитающих происходит за счет развития мантии.
Мантия
Усложнение организации конечного мозга у млекопитающих происходит за счет развития мантии.
Мантия

Онтогенез коры больших полушарий
Передний мозг у человека является закругленным концом нервной
Онтогенез коры больших полушарий
Передний мозг у человека является закругленным концом нервной

Примерно на 8-ой неделе эмбрионального развития растущие аксоны покидают кору, огибая
Примерно на 8-ой неделе эмбрионального развития растущие аксоны покидают кору, огибая

Мозолистое тело появляется в начале 4-го месяца развития и растет очень
Мозолистое тело появляется в начале 4-го месяца развития и растет очень
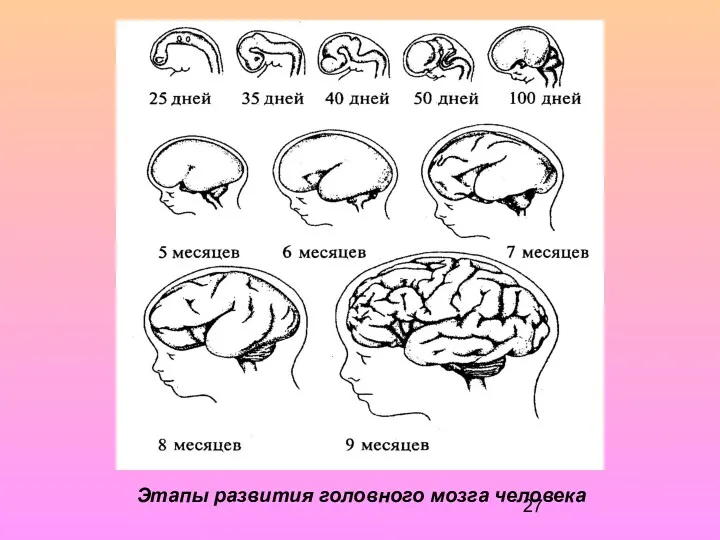
Этапы развития головного мозга человека
Этапы развития головного мозга человека
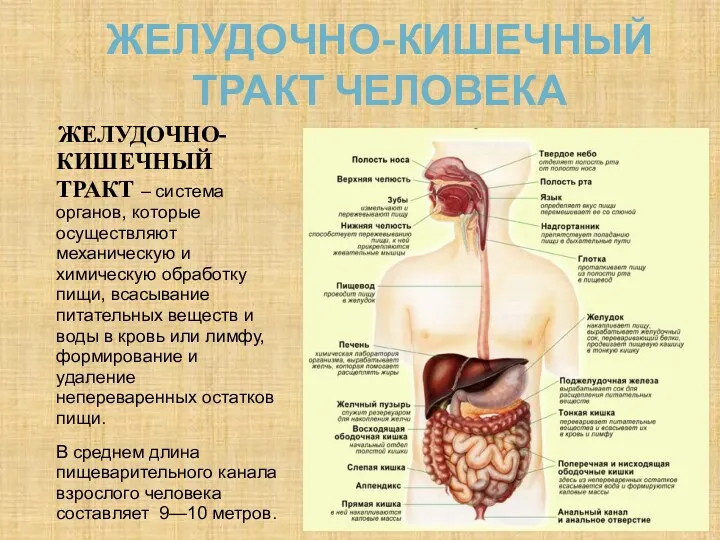 Желудочно-кишечный тракт человека
Желудочно-кишечный тракт человека Размножение. Голосеменные
Размножение. Голосеменные Опорно-двигательный аппарат. Скелет человека
Опорно-двигательный аппарат. Скелет человека Многообразие насекомых, их роль в природе и практическое значение
Многообразие насекомых, их роль в природе и практическое значение Биохимия соединительной ткани
Биохимия соединительной ткани Строение и работа мышц.
Строение и работа мышц. День птиц
День птиц Гербарии растений, содержащих полисахариды
Гербарии растений, содержащих полисахариды Методологія еволюційної систематики та історія макросистем
Методологія еволюційної систематики та історія макросистем Дихальна система
Дихальна система Hedera helix
Hedera helix Молекулярная диагностика. Молекулярно-генетические методы в медицинской практике
Молекулярная диагностика. Молекулярно-генетические методы в медицинской практике Орган вкуса
Орган вкуса Ядро, строение и функции
Ядро, строение и функции Вітаміни у нашому житті
Вітаміни у нашому житті Развитие насекомых
Развитие насекомых Растения и животные лугов и водоемов
Растения и животные лугов и водоемов Высшая нервная деятельность
Высшая нервная деятельность Тип кольчатые черви (16 000 видов)
Тип кольчатые черви (16 000 видов) Микроэкология тела человека. Дисбактериоз и методы исследования
Микроэкология тела человека. Дисбактериоз и методы исследования Методы селекции
Методы селекции разработка урока в 5 классе по теме: Грибы
разработка урока в 5 классе по теме: Грибы Презентация Организм растения - единое целое
Презентация Организм растения - единое целое Популяция как форма существования вида
Популяция как форма существования вида 3 обмен веÑеÑÑв
3 обмен веÑеÑÑв Вид, его критерии и структура
Вид, его критерии и структура Фото для души. Природа
Фото для души. Природа Фізіологія дихальної системи. Зовнішнє дихання
Фізіологія дихальної системи. Зовнішнє дихання