- Структура и функции надорганизменных генетических систем. (Тема 4)

Содержание
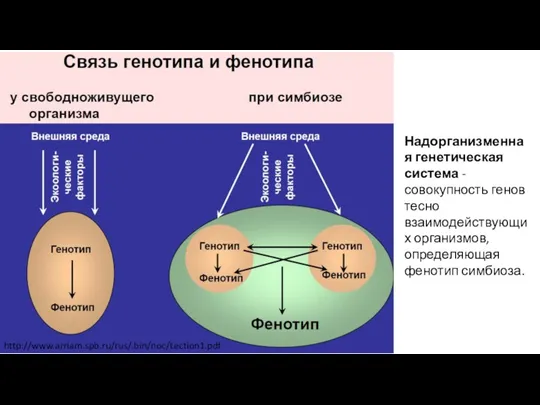
- 2. http://www.arriam.spb.ru/rus/.bin/noc/Lection1.pdf Надорганизменная генетическая система - совокупность генов тесно взаимодействующих организмов, определяющая фенотип симбиоза.
- 3. Единица наследственности при симбиозе состоит как минимум из 2 генов (хозяина и симбионта).
- 4. Программа развития симбиотических систем Узнавание Инфекционный процесс Морфогенез Регуляция Метаболическая интеграция Поддержание Выход партнеров в свободноживущее
- 5. Основные функции надорганизменных генетических систем Сигнальные взаимодействия партнеров Развитие симбиотических структур Метаболическая интеграция партнеров
- 6. Развитие симбиотических структур Функции: 1) вместилище для симбионта; 2) структурная основа для обмена партнерами метаболитов и
- 7. Симбиотические структуры в разных группах организмов
- 8. Морфогенетическая реакция на паразитов у растений Чернильный орешек дубовой орехотворки Cynips quercusfolii Галлы от орехотворки Andricus
- 9. Морфогенетическая реакция на паразитов у растений Галловая нематода – паразит корней высших растений А – здоровая
- 10. Азотфиксирующие клубеньки на корнях гороха («дикий тип»)
- 11. Корончатые галлы на стеблях растений
- 12. Морфогенетическая реакция у животных Моллюск янтарка Succinea с метацеркариями Leucochloridium paradoxum в рожках
- 13. Защитные функции симбиотических структур Строение покровов тела Кутикула нематод Синцитий плоских червей Тегумент скребней Использование экологических
- 14. Токсоплазмоз Хозяева: кошки, собаки, мыши, крысы, домашние животные, человек Источники заражения: сырое мясо, фекалии домашних животных,
- 15. Защитные функции симбиотических структур Индуцирование образования хозяином соединительнотканых капсул Нейтрализация пищеварительных ферментов Отпугивание иммунокомпетентных клеток Препятствование
- 16. Формирование трофосомы Морфогенез личинки вестиментифер после проникновения симбиотических бактерий (показаны розовыми точками). Вверху — продольные разрезы
- 17. Компоненты симбиотического сверхорганизма: тля (вверху) и бактерии Buchnera, живущие в специализированных клетках хозяина — бактериоцитах. Симбионты
- 18. Строение желудка жвачных животных Специализированный отдел - рубец
- 19. ПОСТЕПЕННОЕ ФОРМИРОВАНИЕ СОСТАВА МИКРОФЛОРЫ РУБЦА ЖВАЧНЫХ
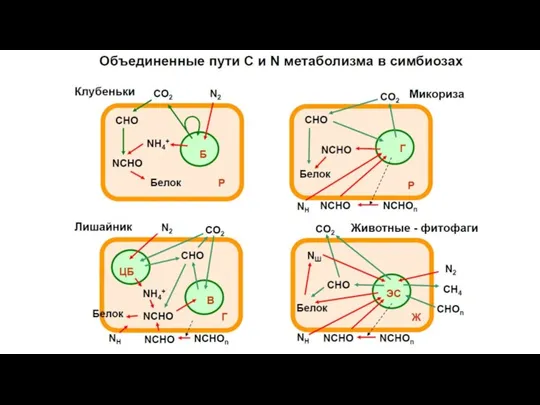
- 20. Метаболическая интеграция партнеров Установление тесных трофических связей Приобретение новых метаболических функций Комплементация биохимических функций симбионтов Донор
- 21. Приобретение новых биохимических функций при симбиозе
- 22. Симбиозы, основанные на биохимической интеграции 1) азотфиксирующие симбиозы специальные ферменты растений, работающие только в клубеньках, снижают
- 23. Биохимическая комплементация в системе «человек-симбионтная микрофлора» В кишечнике взрослого человека присутствует > 1 кг микроорганизмов, относящихся
- 25. Биохимическая кооперация в смешанных культурах микроорганизмов Рис. Схема облигатного мутуализма пекарских дрожжей Saccharomyces cerevisiae и водорослей
- 26. Биохимическая комплементация – преадаптация к формированию мутуалистического симбиоза Рис. Гифы Neurospora crassa (а) и Aspergillus nidulans
- 27. Расширение экологической ниши при биохимической кооперации Гамма-протеобактерии, геном которых удалось ренконструировать, являются хемоавтотрофами, необходимую энергию они
- 28. Закономерности в развитии симбиотических систем, основанных на «биохимической кооперации» Относительность грани между мутуалистическими и антагонистическими отношениями;
- 29. Генетическая комплементация партнеров симбиоза Генетическая комплементация - это взаимодополняющее действие двух (или нескольких) генетических структур, приводящее
- 30. Межгеномная комплементация при внутриклеточном симбиозе В 2001 году открыт «матрёшечный» вид симбиоза - в клетках насекомого
- 31. Эволюция геномов эндосимбионтов Геном бактерии Carsonella, внутриклеточного симбионта насекомых листоблошек содержит всего 159 662 п.о. и
- 32. По Н.А. Проворову (2001), стратегия эволюции симбиотических микроорганизмов включает две составляющие упрощение генома при облигатном симбиозе
- 33. Основные механизмы интеграции партнеров симбиоза Дифференциальная экспрессия генов Направленные модификации генетического материала: полиплоидизация амплификация генетические перестройки
- 34. Мофофизиологическая целостность симбиоза Рис. Повышение целостности бобово-ризобиального симбиоза в процессе макроэволюции: 1 - бесклубеньковые симбиозы, когда
- 35. Физиологическая целостность симбиоза Рис. Системная регуляция бобово-ризобиального симбиоза: LysM RLKs — рецептор-подобная киназа с доменами, связывающими
- 37. Скачать презентацию
http://www.arriam.spb.ru/rus/.bin/noc/Lection1.pdf
Надорганизменная генетическая система - совокупность генов тесно взаимодействующих организмов, определяющая фенотип
http://www.arriam.spb.ru/rus/.bin/noc/Lection1.pdf
Надорганизменная генетическая система - совокупность генов тесно взаимодействующих организмов, определяющая фенотип
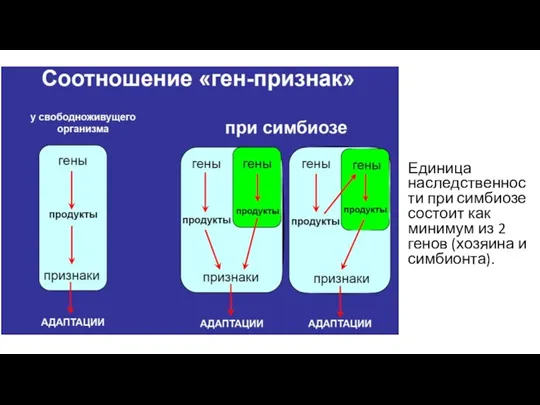
Единица наследственности при симбиозе состоит как минимум из 2 генов (хозяина
Единица наследственности при симбиозе состоит как минимум из 2 генов (хозяина
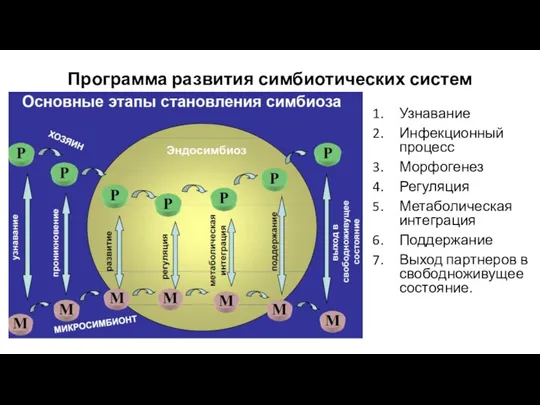
Программа развития симбиотических систем
Узнавание
Инфекционный процесс
Морфогенез
Регуляция
Метаболическая интеграция
Поддержание
Программа развития симбиотических систем
Узнавание
Инфекционный процесс
Морфогенез
Регуляция
Метаболическая интеграция
Поддержание
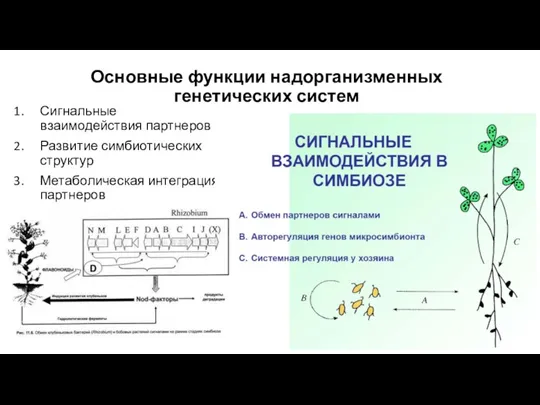
Основные функции надорганизменных генетических систем
Сигнальные взаимодействия партнеров
Развитие симбиотических структур
Метаболическая
Основные функции надорганизменных генетических систем
Сигнальные взаимодействия партнеров
Развитие симбиотических структур
Метаболическая

Развитие симбиотических структур
Функции:
1) вместилище для симбионта;
2) структурная основа для
Развитие симбиотических структур
Функции:
1) вместилище для симбионта;
2) структурная основа для
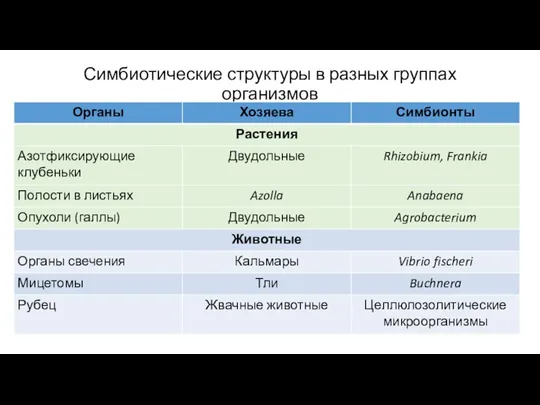
Симбиотические структуры в разных группах организмов
Симбиотические структуры в разных группах организмов

Морфогенетическая реакция на паразитов у растений
Чернильный орешек дубовой
орехотворки Cynips quercusfolii
Галлы
Морфогенетическая реакция на паразитов у растений
Чернильный орешек дубовой
орехотворки Cynips quercusfolii
Галлы

Морфогенетическая реакция на паразитов у растений
Галловая нематода –
паразит корней высших
Морфогенетическая реакция на паразитов у растений
Галловая нематода –
паразит корней высших
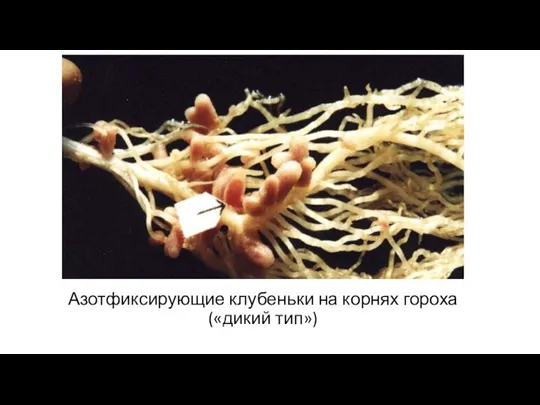
Азотфиксирующие клубеньки на корнях гороха
(«дикий тип»)
Азотфиксирующие клубеньки на корнях гороха
(«дикий тип»)

Корончатые галлы на стеблях растений
Корончатые галлы на стеблях растений

Морфогенетическая реакция у животных
Моллюск янтарка Succinea с метацеркариями Leucochloridium paradoxum в
Морфогенетическая реакция у животных
Моллюск янтарка Succinea с метацеркариями Leucochloridium paradoxum в

Защитные функции симбиотических структур
Строение покровов тела
Кутикула нематод
Синцитий плоских червей
Тегумент скребней
Использование экологических
Защитные функции симбиотических структур
Строение покровов тела
Кутикула нематод
Синцитий плоских червей
Тегумент скребней
Использование экологических
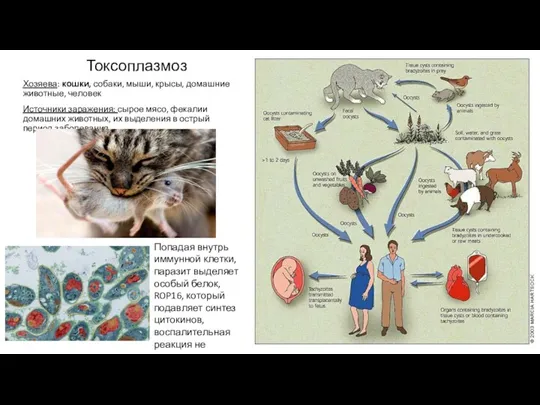
Токсоплазмоз
Хозяева: кошки, собаки, мыши, крысы, домашние животные, человек
Источники заражения: сырое мясо,
Токсоплазмоз
Хозяева: кошки, собаки, мыши, крысы, домашние животные, человек
Источники заражения: сырое мясо,

Защитные функции симбиотических структур
Индуцирование образования хозяином соединительнотканых капсул
Нейтрализация пищеварительных ферментов
Отпугивание
Защитные функции симбиотических структур
Индуцирование образования хозяином соединительнотканых капсул
Нейтрализация пищеварительных ферментов
Отпугивание

Формирование трофосомы
Морфогенез личинки вестиментифер после проникновения симбиотических бактерий (показаны розовыми
Формирование трофосомы
Морфогенез личинки вестиментифер после проникновения симбиотических бактерий (показаны розовыми
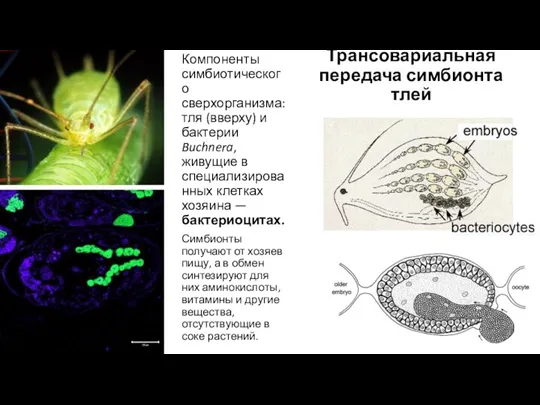
Компоненты симбиотического сверхорганизма: тля (вверху) и бактерии Buchnera, живущие в специализированных
Компоненты симбиотического сверхорганизма: тля (вверху) и бактерии Buchnera, живущие в специализированных
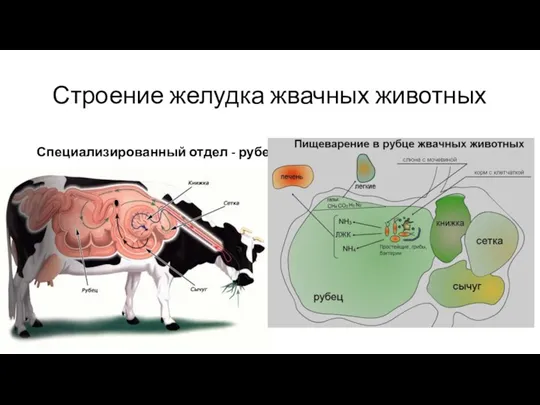
Строение желудка жвачных животных
Специализированный отдел - рубец
Строение желудка жвачных животных
Специализированный отдел - рубец
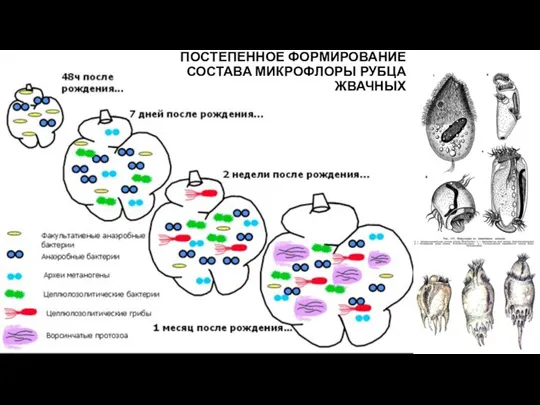
ПОСТЕПЕННОЕ ФОРМИРОВАНИЕ СОСТАВА МИКРОФЛОРЫ РУБЦА ЖВАЧНЫХ
ПОСТЕПЕННОЕ ФОРМИРОВАНИЕ СОСТАВА МИКРОФЛОРЫ РУБЦА ЖВАЧНЫХ
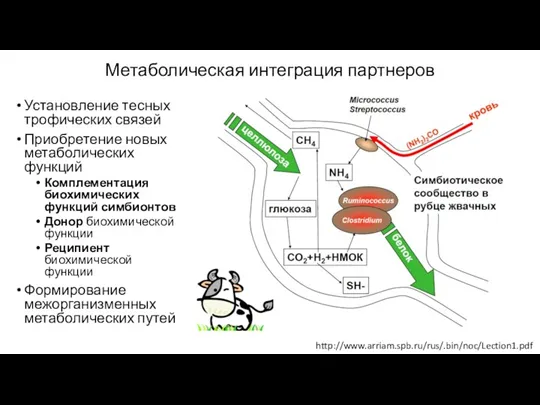
Метаболическая интеграция партнеров
Установление тесных трофических связей
Приобретение новых метаболических функций
Комплементация биохимических функций
Метаболическая интеграция партнеров
Установление тесных трофических связей
Приобретение новых метаболических функций
Комплементация биохимических функций
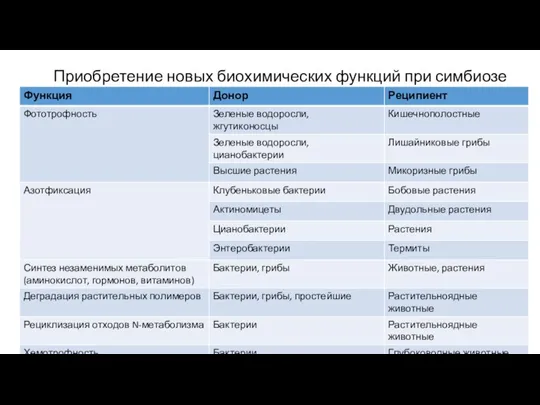
Приобретение новых биохимических функций при симбиозе
Приобретение новых биохимических функций при симбиозе

Симбиозы, основанные на биохимической интеграции
1) азотфиксирующие симбиозы
специальные ферменты растений, работающие только
Симбиозы, основанные на биохимической интеграции
1) азотфиксирующие симбиозы
специальные ферменты растений, работающие только

Биохимическая комплементация в системе
«человек-симбионтная микрофлора»
В кишечнике взрослого человека присутствует >
Биохимическая комплементация в системе
«человек-симбионтная микрофлора»
В кишечнике взрослого человека присутствует >
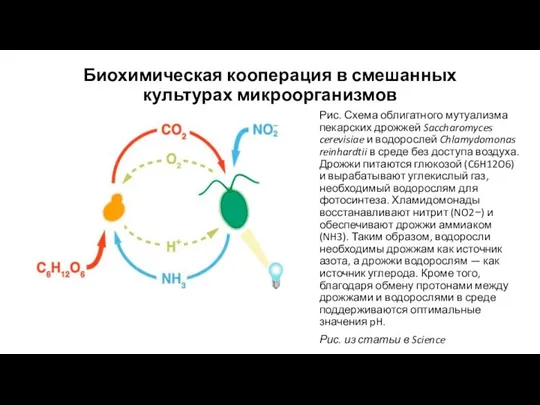
Биохимическая кооперация в смешанных культурах микроорганизмов
Рис. Схема облигатного мутуализма пекарских дрожжей
Биохимическая кооперация в смешанных культурах микроорганизмов
Рис. Схема облигатного мутуализма пекарских дрожжей

Биохимическая комплементация – преадаптация к формированию мутуалистического симбиоза
Рис. Гифы Neurospora crassa
Биохимическая комплементация – преадаптация к формированию мутуалистического симбиоза
Рис. Гифы Neurospora crassa
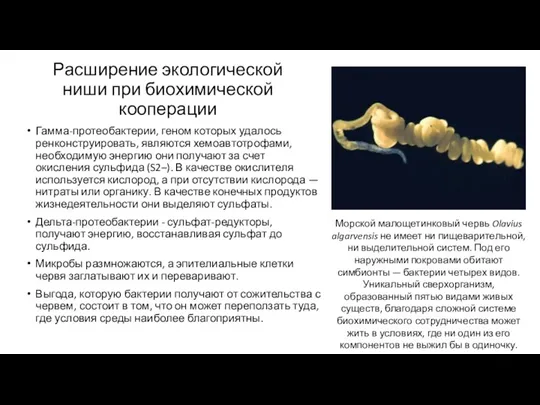
Расширение экологической ниши при биохимической кооперации
Гамма-протеобактерии, геном которых удалось ренконструировать, являются
Расширение экологической ниши при биохимической кооперации
Гамма-протеобактерии, геном которых удалось ренконструировать, являются

Закономерности в развитии симбиотических систем, основанных на «биохимической кооперации»
Относительность грани между
Закономерности в развитии симбиотических систем, основанных на «биохимической кооперации»
Относительность грани между

Генетическая комплементация партнеров симбиоза
Генетическая комплементация - это взаимодополняющее действие двух (или
Генетическая комплементация партнеров симбиоза
Генетическая комплементация - это взаимодополняющее действие двух (или

Межгеномная комплементация при внутриклеточном симбиозе
В 2001 году открыт «матрёшечный» вид симбиоза
Межгеномная комплементация при внутриклеточном симбиозе
В 2001 году открыт «матрёшечный» вид симбиоза
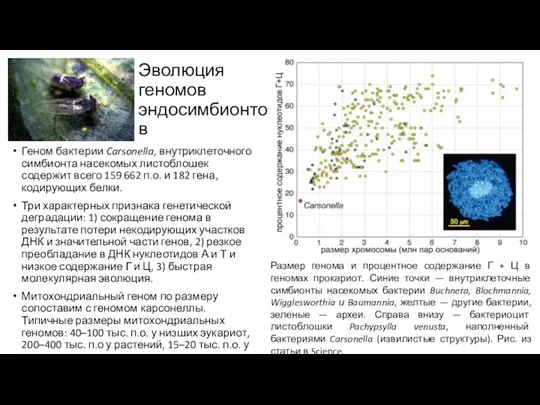
Эволюция геномов эндосимбионтов
Геном бактерии Carsonella, внутриклеточного симбионта насекомых листоблошек содержит всего
Эволюция геномов эндосимбионтов
Геном бактерии Carsonella, внутриклеточного симбионта насекомых листоблошек содержит всего

По Н.А. Проворову (2001), стратегия эволюции симбиотических микроорганизмов включает две составляющие
По Н.А. Проворову (2001), стратегия эволюции симбиотических микроорганизмов включает две составляющие

Основные механизмы интеграции партнеров симбиоза
Дифференциальная экспрессия генов
Направленные модификации генетического материала:
Основные механизмы интеграции партнеров симбиоза
Дифференциальная экспрессия генов
Направленные модификации генетического материала:

Мофофизиологическая целостность симбиоза
Рис. Повышение целостности бобово-ризобиального симбиоза в процессе макроэволюции:
1
Мофофизиологическая целостность симбиоза
Рис. Повышение целостности бобово-ризобиального симбиоза в процессе макроэволюции:
1

Физиологическая целостность симбиоза
Рис. Системная регуляция бобово-ризобиального симбиоза: LysM RLKs — рецептор-подобная
Физиологическая целостность симбиоза
Рис. Системная регуляция бобово-ризобиального симбиоза: LysM RLKs — рецептор-подобная
 Презентация по биологии Многообразие пресмыкающихся
Презентация по биологии Многообразие пресмыкающихся Запилення квіток
Запилення квіток Витаминные, ферментные, антиферментные препараты. Понятие о биологически активных добавках к пище
Витаминные, ферментные, антиферментные препараты. Понятие о биологически активных добавках к пище Надходження речовин у клітину. Біологія. 6 клас
Надходження речовин у клітину. Біологія. 6 клас Звук в природе и технике
Звук в природе и технике Глотка. Пищевод. Желудок
Глотка. Пищевод. Желудок презентация к уроку биологии 7 класс
презентация к уроку биологии 7 класс Водоросли
Водоросли Движение живых организмов
Движение живых организмов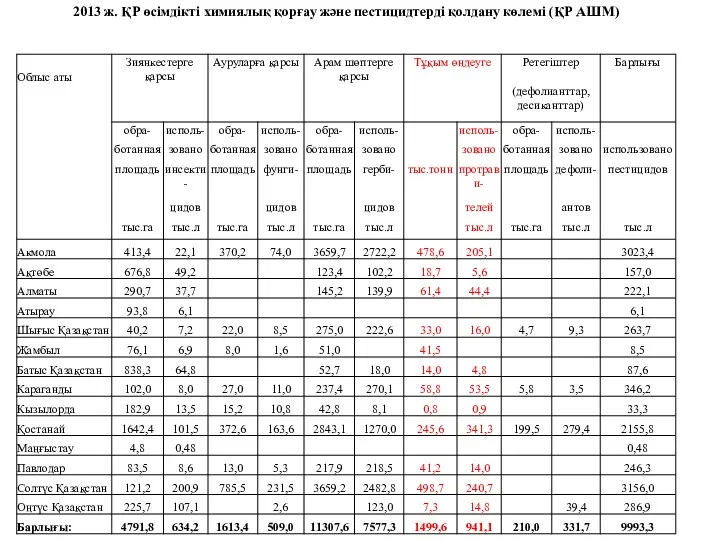 ҚР өсімдікті химиялық қорғау және пестицидтерді қолдану көлемі (ҚР АШМ)
ҚР өсімдікті химиялық қорғау және пестицидтерді қолдану көлемі (ҚР АШМ) Опале листя. Шкода чи користь
Опале листя. Шкода чи користь Транспорт веществ через биологические мембраны
Транспорт веществ через биологические мембраны Видоизменения побегов
Видоизменения побегов Биология – комплекс наук о живой природе. Введение в предмет
Биология – комплекс наук о живой природе. Введение в предмет Тварини рекордсмени
Тварини рекордсмени Трихинелла (Trichinella spiralis)
Трихинелла (Trichinella spiralis) Мастер-класс Наследование групп крови
Мастер-класс Наследование групп крови Микробиология, как наука
Микробиология, как наука Земноводные, или амфибии
Земноводные, или амфибии Биотехнологические источники ЛРС для получения лекарственных средств
Биотехнологические источники ЛРС для получения лекарственных средств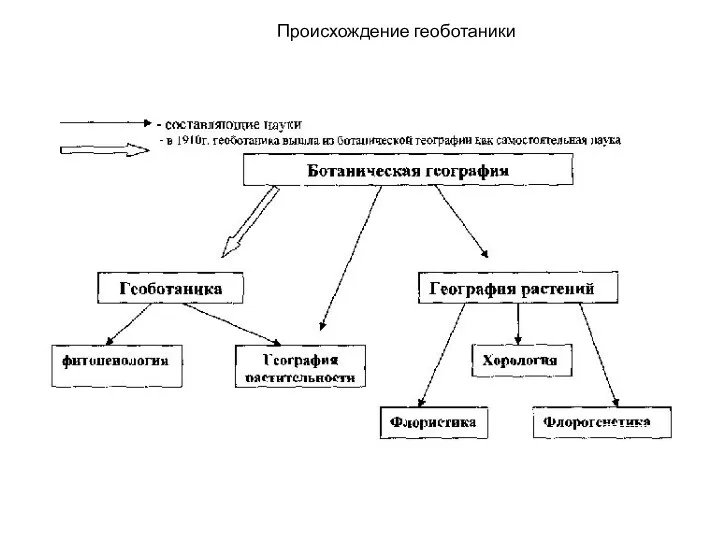 Происхождение геоботаники
Происхождение геоботаники Мейоз
Мейоз Экскурсии по биологии как форма обучения
Экскурсии по биологии как форма обучения Наследственность и изменчивость организмов
Наследственность и изменчивость организмов Мыщцы торса. Поверхностная группа мыщц, формирующая рельеф груди, живота, спины
Мыщцы торса. Поверхностная группа мыщц, формирующая рельеф груди, живота, спины Растительная клетка. Лекция 1
Растительная клетка. Лекция 1 Становлення еволюційних поглядів
Становлення еволюційних поглядів Гепард — хищное млекопитающее семейства кошачьих
Гепард — хищное млекопитающее семейства кошачьих