- Транскрипция и трансляция

Содержание
- 2. Генетический код Характеристики: Записывается в линейной форме, в качестве букв выступают рибонуклеотиды РНК, последовательность которых комплементарна
- 3. Генетический код В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной РНК=РНК посредника. Триплетность кода:
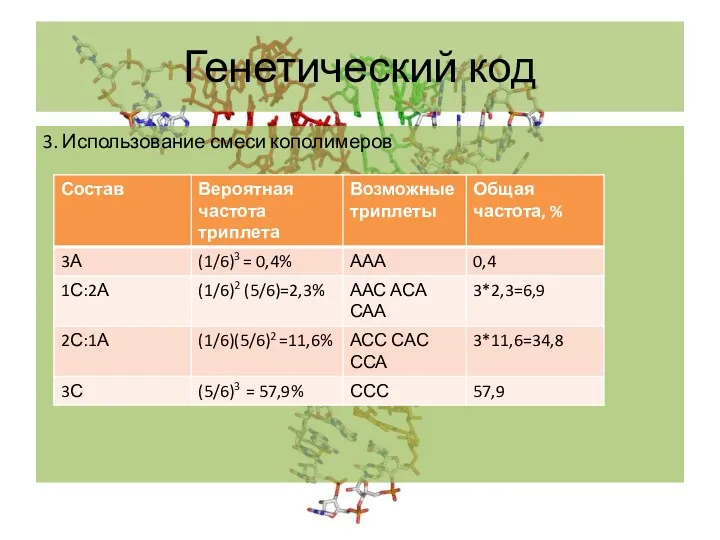
- 4. Генетический код 3. Использование смеси кополимеров
- 5. Генетический код Метод связывания триплетов В 1964 г. Ниренберг и Ледер разработали данный метод для установления
- 6. Генетический код 5.Использование повторяющихся кополимеров Гобинд Корана синтезировал протяженные молекулы РНК с заданной последовательностью, многократно повторяющейся.
- 7. Генетический код Кодовый словарь AUG старт кодон UAA UAG UGA стоп кодоны
- 8. Генетический код В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis). Предположил, что для комплементации с
- 9. Транскрипция Синтез РНК на ДНК-матрице называется транскрипцией. Транскрипция – начало информационного потока в клетке РНК посредник
- 10. Транскрипция Связывание РНК-полимеразы с матрицей происходит в сайтах – промоторах. Локализованы в 5` области, левее точки
- 11. Транскрипция у эукариот Различия: Участвуют три разные формы РНК-полимеразы, процесс происходит в ядре. Кроме промотеров находятся
- 12. Транскрипция у эукариот: инициация 3 формы РНК-полимеразы состоят из: 2 больших субъединицы и 10-15 малых. РНК-полимераза
- 13. Транскрипция эукариот: процессинг Шаг 1: первичная посттранскрипционная модификация: присоединение к 5`- концу молекулы 7-метилгуанозина (кэп) Шаг
- 14. Транскрипция у эукариот: сплайсинг В зависимости от специфичности механихма сплайсинга, интроны подразделяются на группы: Интроны, которые
- 15. Транскрипция у эукариот: эдитинг Эдитинг-редактирование РНК В процессе эдитинга последовательность зрелой РНК отличается от последовательности, кодируемой
- 16. Трансляция Трансляция мРНК- биополимеризация аминокислот в полипептидную цепь. Структура тРНК: Роберт Холли в 1965 г. Расшифровал
- 17. Трансляция Стадии: Инициация трансляции: образование комплекса+ инициирующий кодон: AUG+ последовательность Шайна-Дельгарно Образованный комплекс инициации ассоциирует с
- 18. Трансляция
- 19. Трансляция у эукариот Особенности инициации: Наличие кэпа на 5`-конце увеличивает эффективность трансляции Кодон AUG в эукариотической
- 20. Посттрансляционная модификация белков Модификация N и C концов аминокислот Модификация отдельных аминокислотных остатков Присоединение боковых цепей
- 22. Скачать презентацию
Генетический код
Характеристики:
Записывается в линейной форме, в качестве букв выступают рибонуклеотиды РНК,
Генетический код
Характеристики:
Записывается в линейной форме, в качестве букв выступают рибонуклеотиды РНК,
Генетический код
В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной
Генетический код
В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной
Генетический код
3. Использование смеси кополимеров
Генетический код
3. Использование смеси кополимеров
Генетический код
Метод связывания триплетов
В 1964 г. Ниренберг и Ледер разработали данный
Генетический код
Метод связывания триплетов
В 1964 г. Ниренберг и Ледер разработали данный
Генетический код
5.Использование повторяющихся кополимеров
Гобинд Корана синтезировал протяженные молекулы РНК с заданной
Генетический код
5.Использование повторяющихся кополимеров
Гобинд Корана синтезировал протяженные молекулы РНК с заданной
Генетический код
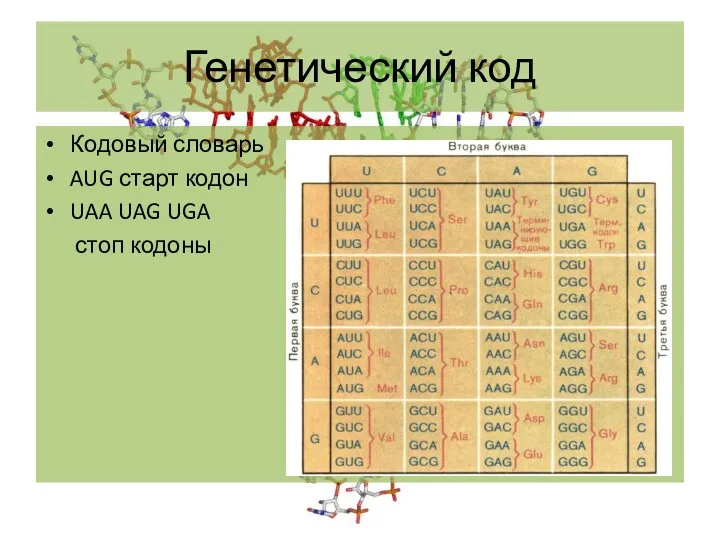
Кодовый словарь
AUG старт кодон
UAA UAG UGA
стоп кодоны
Генетический код
Кодовый словарь
AUG старт кодон
UAA UAG UGA
стоп кодоны
Генетический код
В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis).
Предположил, что
Генетический код
В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis).
Предположил, что

Транскрипция
Синтез РНК на ДНК-матрице называется транскрипцией.
Транскрипция – начало информационного потока в
Транскрипция
Синтез РНК на ДНК-матрице называется транскрипцией.
Транскрипция – начало информационного потока в

Транскрипция
Связывание РНК-полимеразы с матрицей происходит в сайтах – промоторах.
Локализованы в 5`
Транскрипция
Связывание РНК-полимеразы с матрицей происходит в сайтах – промоторах.
Локализованы в 5`

Транскрипция у эукариот
Различия:
Участвуют три разные формы РНК-полимеразы, процесс происходит в ядре.
Кроме
Транскрипция у эукариот
Различия:
Участвуют три разные формы РНК-полимеразы, процесс происходит в ядре.
Кроме

Транскрипция у эукариот: инициация
3 формы РНК-полимеразы состоят из: 2 больших
Транскрипция у эукариот: инициация
3 формы РНК-полимеразы состоят из: 2 больших

Транскрипция эукариот: процессинг
Шаг 1: первичная посттранскрипционная модификация: присоединение к 5`- концу
Транскрипция эукариот: процессинг
Шаг 1: первичная посттранскрипционная модификация: присоединение к 5`- концу

Транскрипция у эукариот: сплайсинг
В зависимости от специфичности механихма сплайсинга, интроны подразделяются
Транскрипция у эукариот: сплайсинг
В зависимости от специфичности механихма сплайсинга, интроны подразделяются
Транскрипция у эукариот: эдитинг
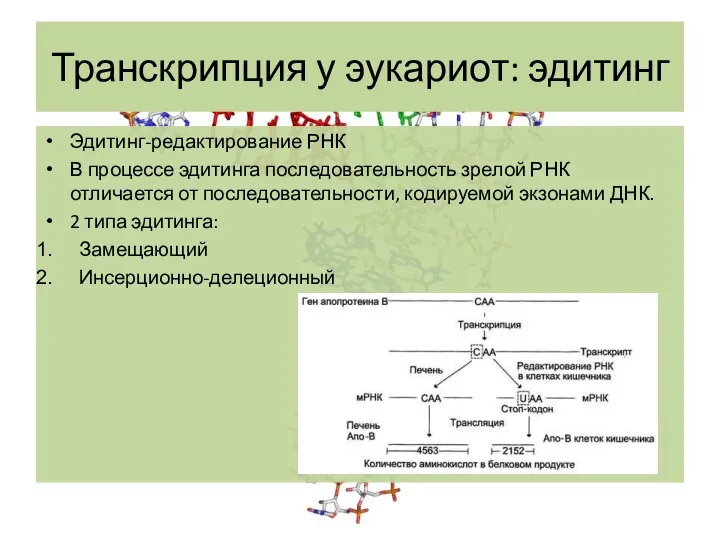
Эдитинг-редактирование РНК
В процессе эдитинга последовательность зрелой РНК отличается
Транскрипция у эукариот: эдитинг
Эдитинг-редактирование РНК
В процессе эдитинга последовательность зрелой РНК отличается

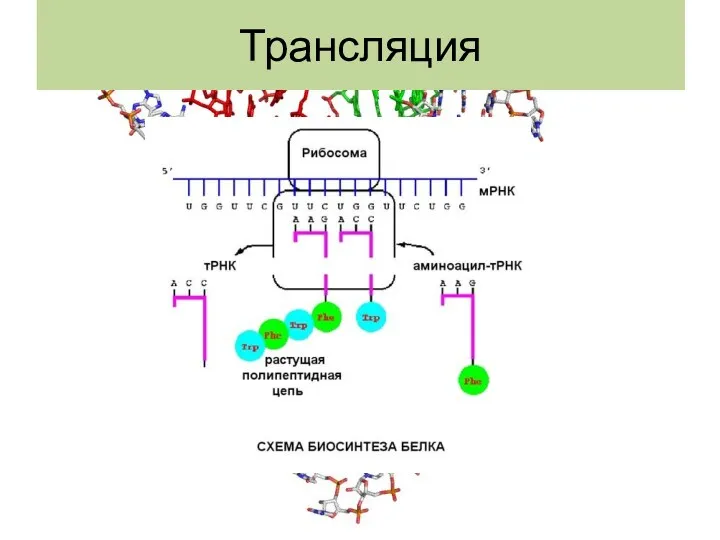
Трансляция
Трансляция мРНК- биополимеризация аминокислот в полипептидную цепь.
Структура тРНК: Роберт Холли в
Трансляция
Трансляция мРНК- биополимеризация аминокислот в полипептидную цепь.
Структура тРНК: Роберт Холли в
Трансляция
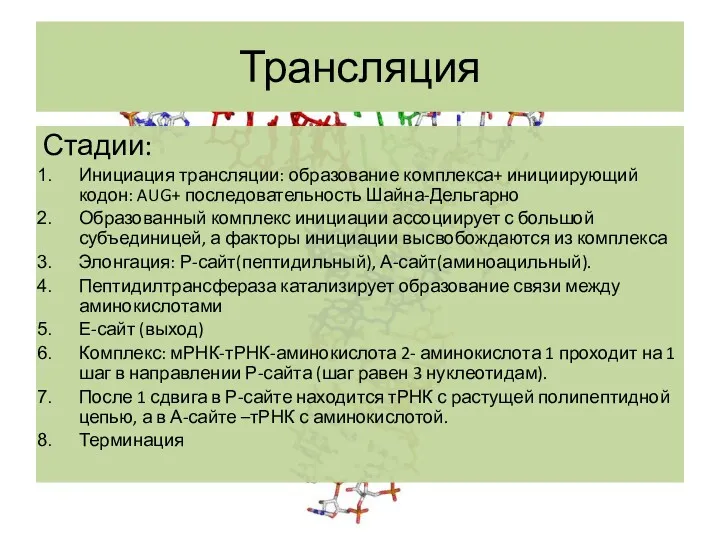
Стадии:
Инициация трансляции: образование комплекса+ инициирующий кодон: AUG+ последовательность Шайна-Дельгарно
Образованный комплекс инициации
Трансляция
Стадии:
Инициация трансляции: образование комплекса+ инициирующий кодон: AUG+ последовательность Шайна-Дельгарно
Образованный комплекс инициации
Трансляция
Трансляция

Трансляция у эукариот
Особенности инициации:
Наличие кэпа на 5`-конце увеличивает эффективность трансляции
Кодон
Трансляция у эукариот
Особенности инициации:
Наличие кэпа на 5`-конце увеличивает эффективность трансляции
Кодон
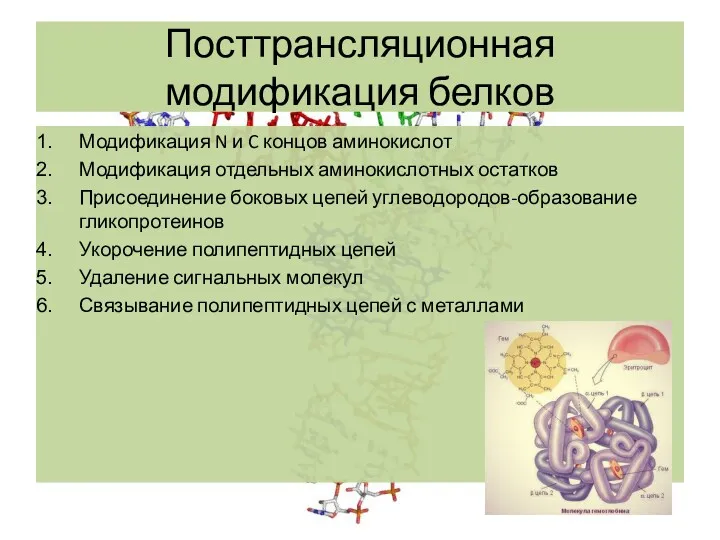
Посттрансляционная модификация белков
Модификация N и C концов аминокислот
Модификация отдельных аминокислотных остатков
Присоединение
Посттрансляционная модификация белков
Модификация N и C концов аминокислот
Модификация отдельных аминокислотных остатков
Присоединение
 Характеристика отдела водорослей Сryptophyta. (Лекция 3)
Характеристика отдела водорослей Сryptophyta. (Лекция 3) Растения и животные разных материков
Растения и животные разных материков Обмен веществ и превращение энергии. Раздел 2
Обмен веществ и превращение энергии. Раздел 2 Теория зародышевых листков
Теория зародышевых листков Кроссинговер
Кроссинговер Метаболизм – основа существования живых организмов. Автотрофный тип обмена веществ. Фотосинтез. Хемосинтез
Метаболизм – основа существования живых организмов. Автотрофный тип обмена веществ. Фотосинтез. Хемосинтез С какого дерева листок?
С какого дерева листок? Vitamin A
Vitamin A Нервная ткань
Нервная ткань Клеточное строение организмов как доказательство их родства, единства живой природы. Тренажер
Клеточное строение организмов как доказательство их родства, единства живой природы. Тренажер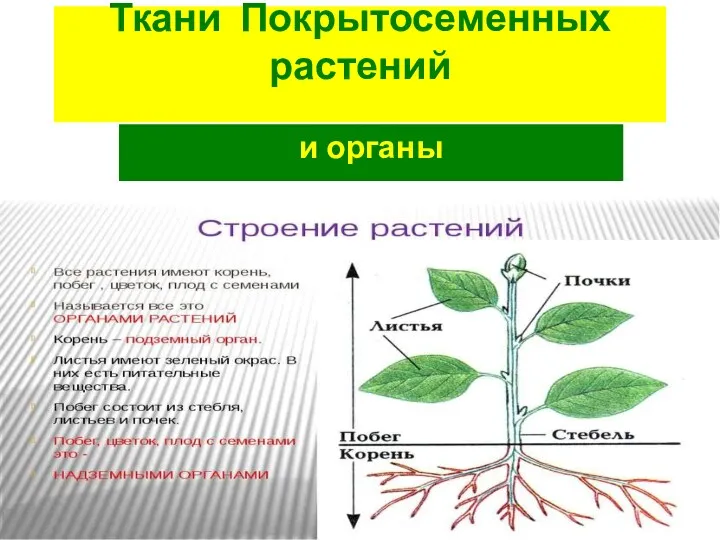 Ткани и органы покрытосеменных растений
Ткани и органы покрытосеменных растений Закономерности изменчивости
Закономерности изменчивости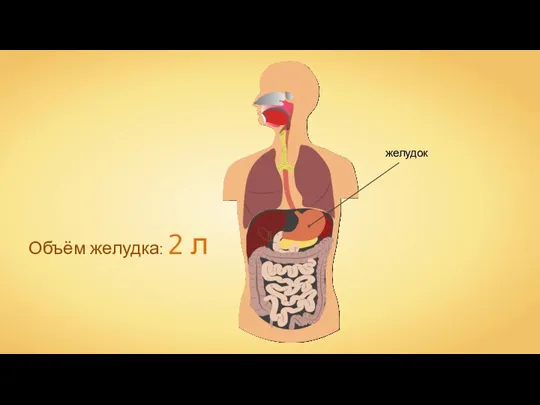 Пищеварение в желудке и кишечнике
Пищеварение в желудке и кишечнике Викторина Животные
Викторина Животные Химия и переваривание углеводов
Химия и переваривание углеводов Як бачать тварини
Як бачать тварини Дикие и домашние животные
Дикие и домашние животные Royal Botanic Gardens — Kew World Heritage in London
Royal Botanic Gardens — Kew World Heritage in London История развития генетики
История развития генетики Класс Птицы
Класс Птицы Биоэкологические особенности вредной черепашки и совершенствование мер борьбы с ней в Центральном Предкавказье
Биоэкологические особенности вредной черепашки и совершенствование мер борьбы с ней в Центральном Предкавказье Луговые растения
Луговые растения Этапы развития науки генетики
Этапы развития науки генетики Подцарство Одноклеточные, или Простейшие
Подцарство Одноклеточные, или Простейшие Возникновение и эволюция биосферы
Возникновение и эволюция биосферы Экспериментальная терапия клеточными культурами нарушений спермато- и овогенеза 2
Экспериментальная терапия клеточными культурами нарушений спермато- и овогенеза 2 Большой год 2019 областной фотоконкурс
Большой год 2019 областной фотоконкурс Работы Никитина Никиты уч-ся 7 класса
Работы Никитина Никиты уч-ся 7 класса