- Углеводы и обмен углеводов

Содержание
- 2. Углеводы и обмен углеводов. Гликоген. Строение. Значение. Синтез гликогена. Ферменты. Мобилизация гликогена. Фосфоролиз. Ферменты. Связь гликогенолиза
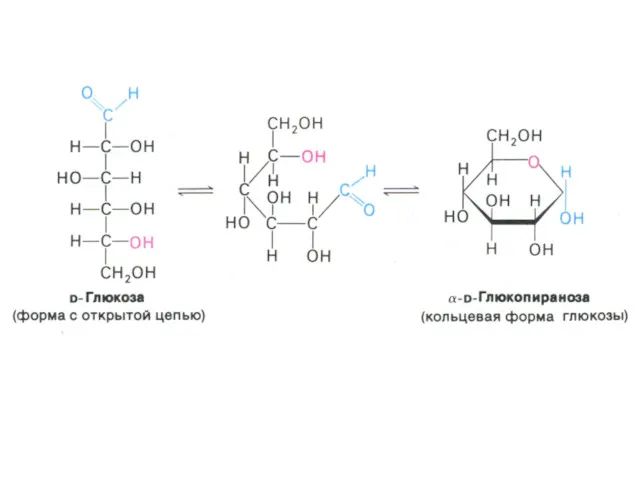
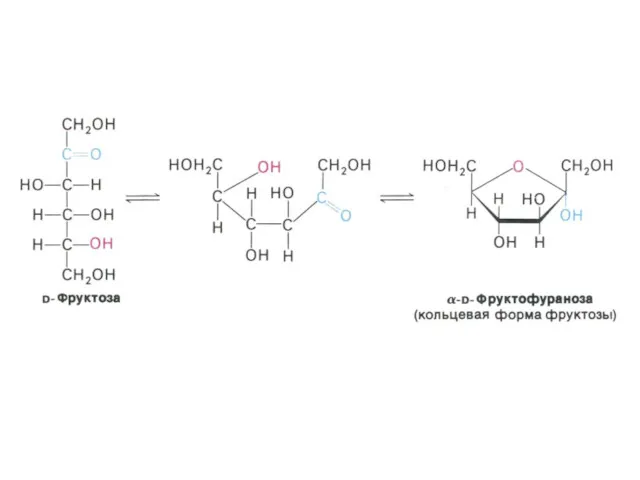
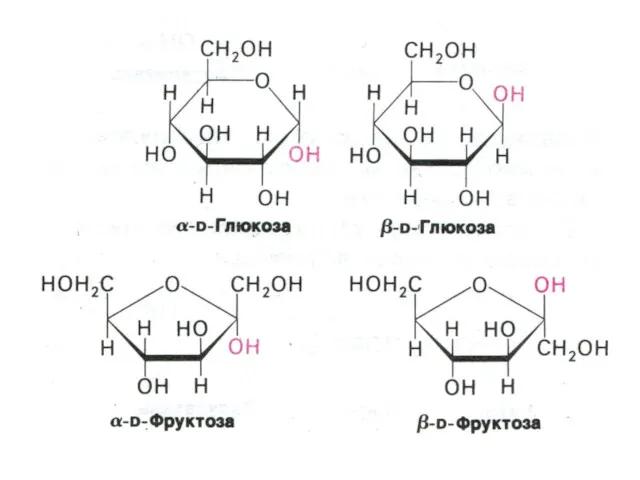
- 3. Углеводы и обмен углеводов Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные и кислые); Ацетилированные, аминированные,
- 9. Протеогликановый агрегат из эпифизарного хряща
- 10. Функции углеводов 1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза. Преимущество окисления углеводов в анаэробных условиях.
- 11. Функции углеводов Модификация белков – гликозилирование (норма и патология); Углеводы – как лекарственные препараты (глюкоза, гепарин,
- 12. Внешний обмен углеводов Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ? декстрины, мальтотриозы, мальтоза. Гидролизуют
- 13. Промежуточный обмен углеводов Катаболизм: Гликолиз и пентозофосфатный шунт Окисление пирувата и ЦТК Гликогенолиз (расщепление гликогена) Анаболизм:
- 14. Особенность промежуточного обмена углеводов Сахара метаболизируют внутри клетки только в фосфорилированном виде. Фосфорилирование осуществляется гексокиназами, дефосфорилирование
- 15. Взаимопревращения сахаров При потреблении различных сахаров, в крови содержание глюкозы в сотни раз превышает таковое для
- 16. Расходование глюкозы клеткой Гликолиз и тканевое дыхание (65%) Запасание гликогена (5%) Синтез липидов при наличии избытка
- 17. ГЛИКОЛИЗ Центральный путь метаболизма глюкозы Самостоятельный способ получения энергии клеткой (в анаэробных условиях) и первый этап
- 30. Гликолиз, конечные продукты Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение). В различных клетках и
- 32. Особенности гликолиза в разных тканях Наиболее распространенный шунт окисления глюкозы – пентозофосфатные превращения в печени, жировой
- 33. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы Три ферментные системы, локализованные в цитоплазме: Окислительно-декарбоксилирующая (образование пентозы – рибулозо-5-фосфата и
- 41. Шунт Раппопорта в эритроцитах Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося в условиях гипоксии) 2,3-
- 43. Аэробный метаболизм глюкозы Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает с помощь переносчика (пируват-
- 50. Окисление пирувата Необратимая реакция. Продукт окисления и декарбоксилирования – ацетил-КоА. Протон фиксируется в НАДН. 2-х углеродный
- 51. Структура и функции КоА Низкомолекулярный термостабильный (не связан с белком) фактор – активатор и переносчик ацетильных
- 52. Ацетил-КоА Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 – в цикле лимонной кислоты
- 53. Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса. 8 реакций, замкнутых в цикл («катализатором» является оксалоацетат), в
- 58. α –кетоглутаратдегидрогеназный комплекс Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному комплексу. Содержит 3 фермента
- 59. Субстратное фосфорилирование Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования АДФ (ГДФ) до АТФ
- 62. Функции цикла Кребса Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса электронов в составе 3
- 63. Глюконеогенез Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК). 4 адаптивных фермента, «обращающих» лимитирующие
- 68. Перемещение оксалоацетата из митохондрий в цитозоль Пируват, проникнув в митохондрии, карбоксилируется до оксалоацетата (с участием биотина).
- 69. Ферменты глюконеогенеза Активны в печени, мозге, почках. Нет в скелетных мышцах. ФЭП- карбоксикиназа; фруктозо-1,6- дифосфатаза; глюкозо-6
- 72. Метаболизм гликогена Запасы гликогена в клетках разных тканей. Значение. Структура гликогена. Синтез гликогена. Условия. Локализация. Затравка.
- 76. Фосфоролиз гликогена В отличие от внешнего обмена (ЖКТ) , в клетке гликоген распадается путем фосфоролиза с
- 80. Синтез гликогена Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1 – глюкоза. Гл ?
- 84. Синтез гликогена Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6 –гликозилтрансферазой Гликогенсинтаза образует 1,4
- 85. Регуляция метаболизма гликогена Синтез и распад гликогена регулируются по одним и тем же механизмам (фосфорилирование-дефосфорилирование ключевых
- 86. Регуляция метаболизма гликогена Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется дефосфорилированием при участии протеинфосфатазы. В
- 87. Регуляция метаболизма гликогена В мышцах: В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ; При слабых и умеренных
- 91. Скачать презентацию

Углеводы и обмен углеводов.
Гликоген. Строение. Значение.
Синтез гликогена. Ферменты.
Мобилизация гликогена.
Углеводы и обмен углеводов.
Гликоген. Строение. Значение.
Синтез гликогена. Ферменты.
Мобилизация гликогена.

Углеводы и обмен углеводов
Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные
Углеводы и обмен углеводов
Классификация углеводов (моно-, дисахариды, олигосахариды, полисахариды - нейтральные
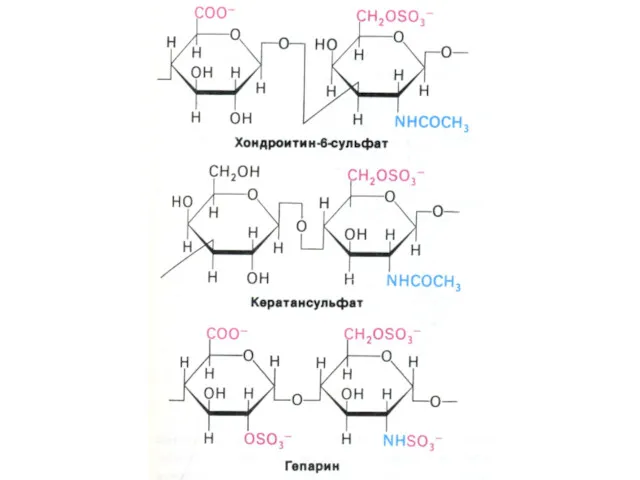

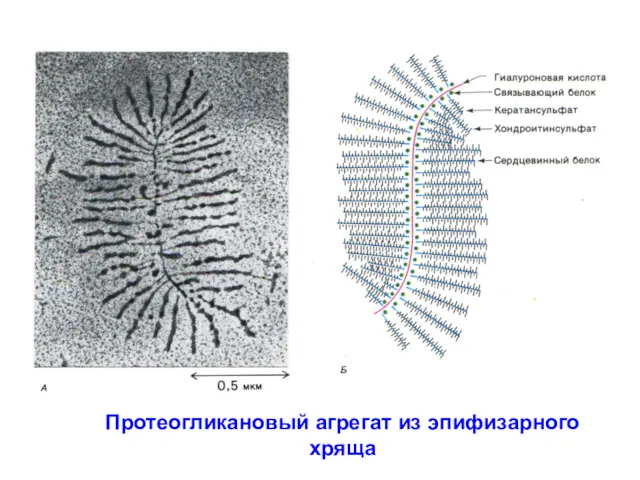
Протеогликановый агрегат из эпифизарного хряща
Протеогликановый агрегат из эпифизарного хряща

Функции углеводов
1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза.
Преимущество
Функции углеводов
1.Энергетическая (1г углеводов – 4,1 ккал) – глюкоза.
Преимущество

Функции углеводов
Модификация белков – гликозилирование (норма и патология);
Углеводы – как лекарственные
Функции углеводов
Модификация белков – гликозилирование (норма и патология);
Углеводы – как лекарственные

Внешний обмен углеводов
Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ?
Внешний обмен углеводов
Эндоамилаза слюны и панкреатического сока: крахмал и гликоген ?

Промежуточный обмен углеводов
Катаболизм:
Гликолиз и пентозофосфатный шунт
Окисление пирувата и ЦТК
Гликогенолиз (расщепление
Промежуточный обмен углеводов
Катаболизм:
Гликолиз и пентозофосфатный шунт
Окисление пирувата и ЦТК
Гликогенолиз (расщепление

Особенность промежуточного обмена углеводов
Сахара метаболизируют внутри клетки только в фосфорилированном виде.
Фосфорилирование
Особенность промежуточного обмена углеводов
Сахара метаболизируют внутри клетки только в фосфорилированном виде.
Фосфорилирование

Взаимопревращения сахаров
При потреблении различных сахаров, в крови содержание глюкозы в сотни
Взаимопревращения сахаров
При потреблении различных сахаров, в крови содержание глюкозы в сотни

Расходование глюкозы клеткой
Гликолиз и тканевое дыхание (65%)
Запасание гликогена (5%)
Синтез липидов при
Расходование глюкозы клеткой
Гликолиз и тканевое дыхание (65%)
Запасание гликогена (5%)
Синтез липидов при

ГЛИКОЛИЗ
Центральный путь метаболизма глюкозы
Самостоятельный способ получения энергии клеткой (в анаэробных условиях)
ГЛИКОЛИЗ
Центральный путь метаболизма глюкозы
Самостоятельный способ получения энергии клеткой (в анаэробных условиях)

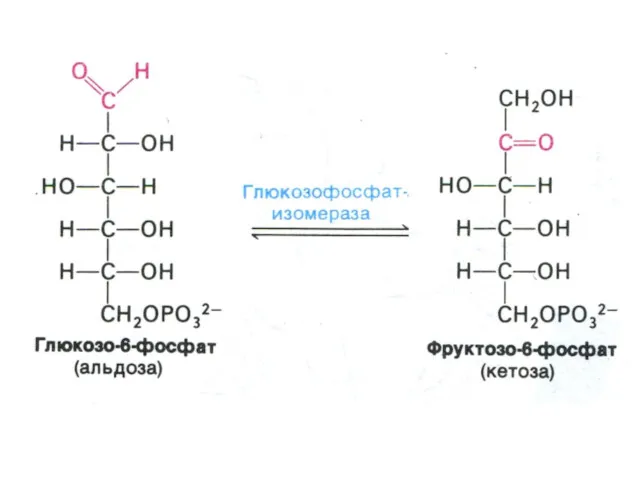
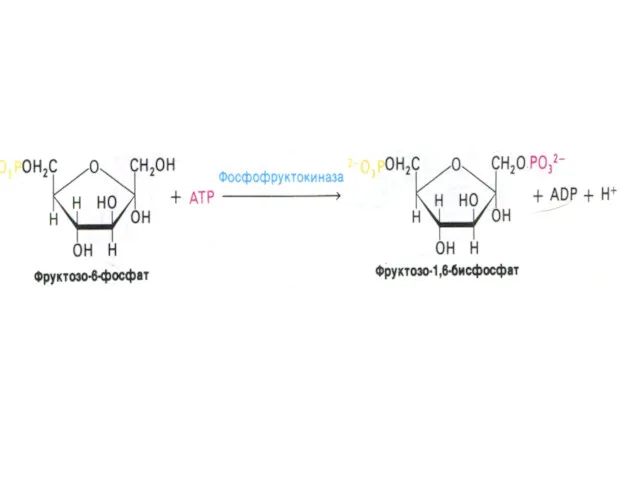
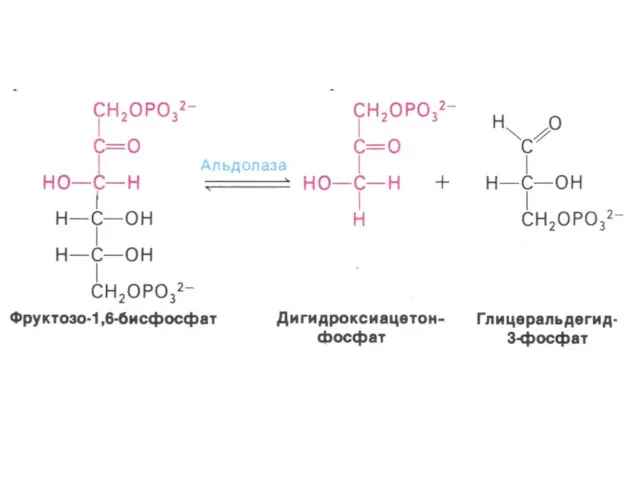
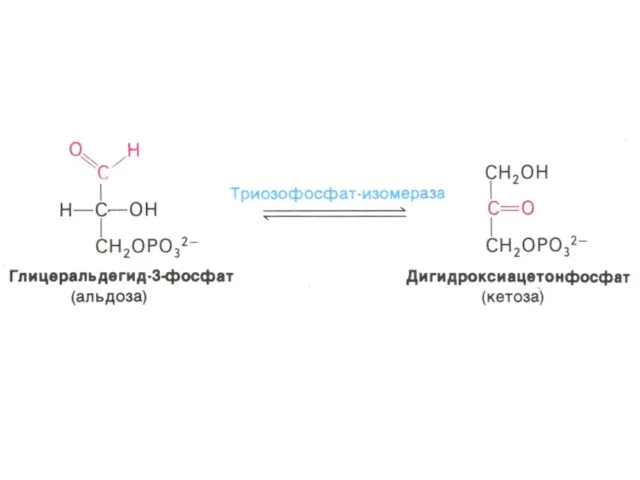
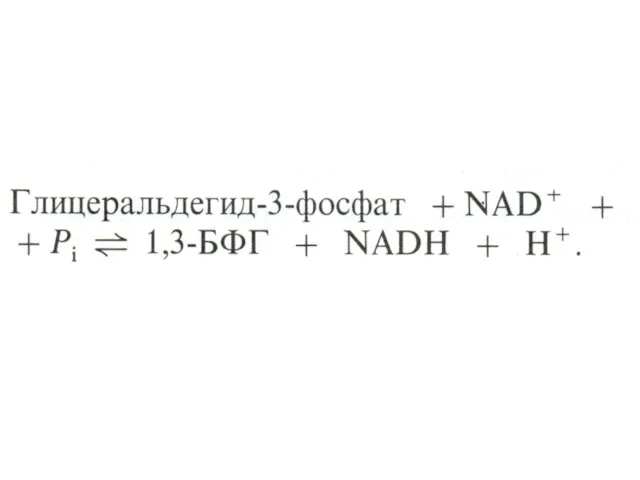

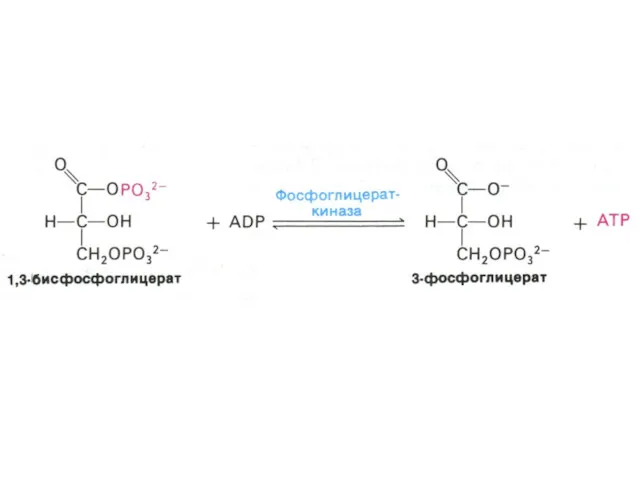
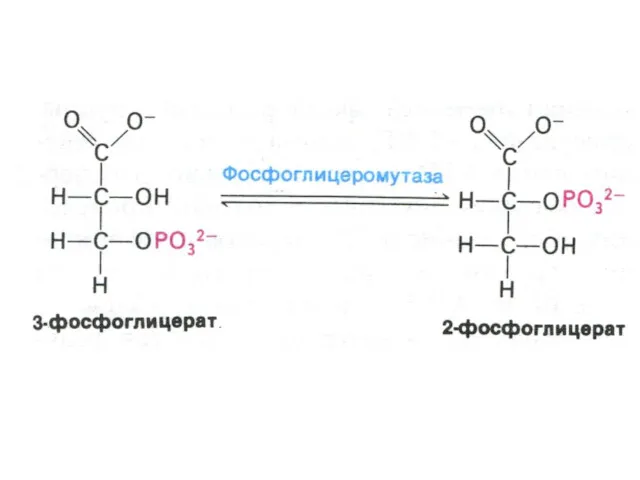
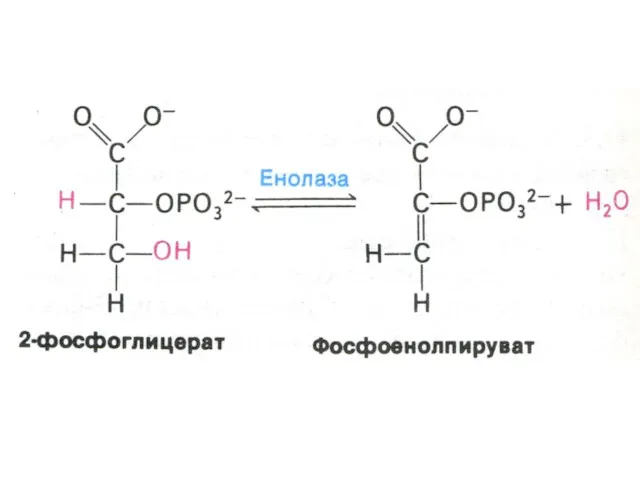
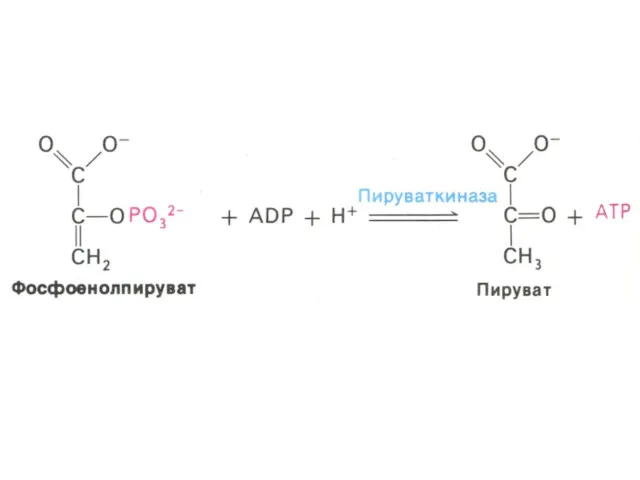


Гликолиз, конечные продукты
Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение).
Гликолиз, конечные продукты
Гликолиз – окисление глюкозы, не требующее присутствия кислорода (брожение).


Особенности гликолиза в разных тканях
Наиболее распространенный шунт окисления глюкозы – пентозофосфатные
Особенности гликолиза в разных тканях
Наиболее распространенный шунт окисления глюкозы – пентозофосфатные

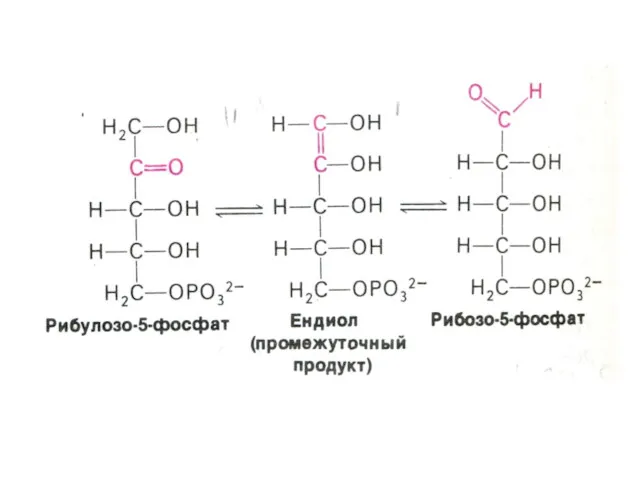
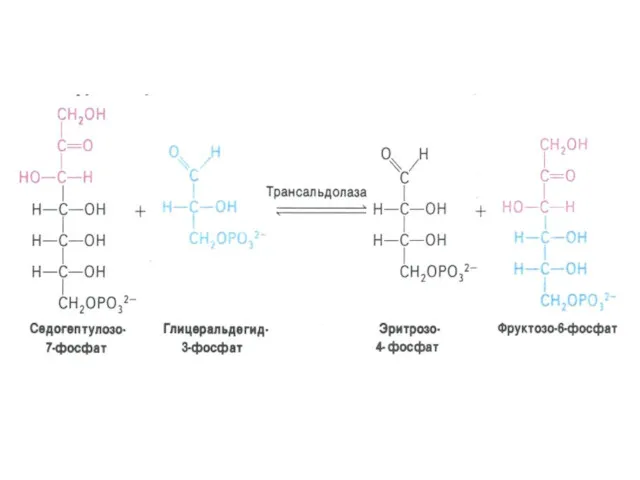
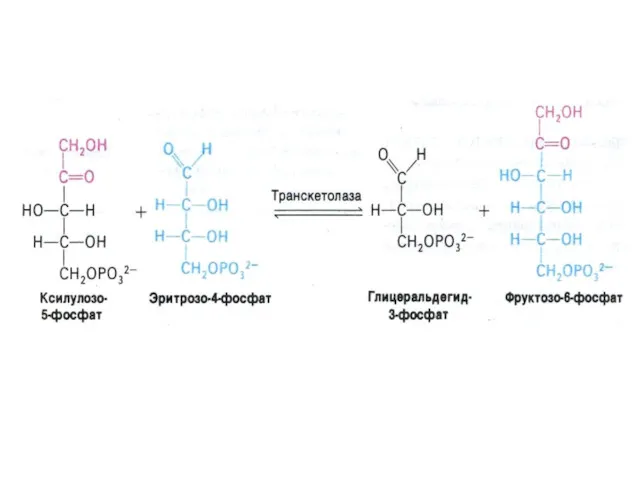
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы
Три ферментные системы, локализованные в цитоплазме:
Окислительно-декарбоксилирующая (образование пентозы
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ окисления глюкозы
Три ферментные системы, локализованные в цитоплазме:
Окислительно-декарбоксилирующая (образование пентозы
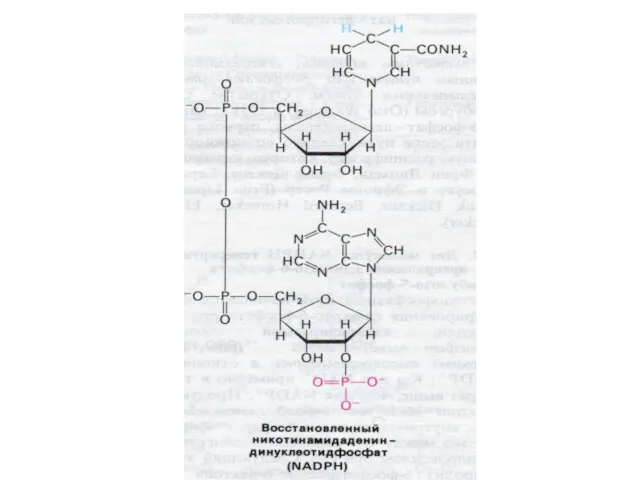


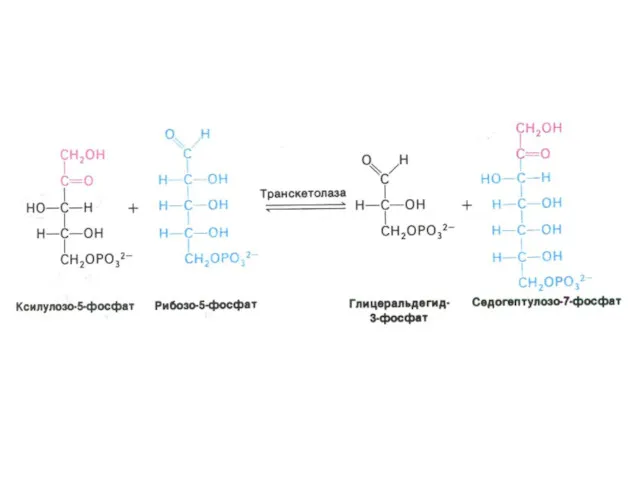

Шунт Раппопорта в эритроцитах
Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося
Шунт Раппопорта в эритроцитах
Особенностью гликолиза в эритроцитах является образование 2,3-глицерофосфата.(особенно усиливающегося


Аэробный метаболизм глюкозы
Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает
Аэробный метаболизм глюкозы
Окисление глюкозы продолжается в матриксе митохондрий, куда пируват поступает


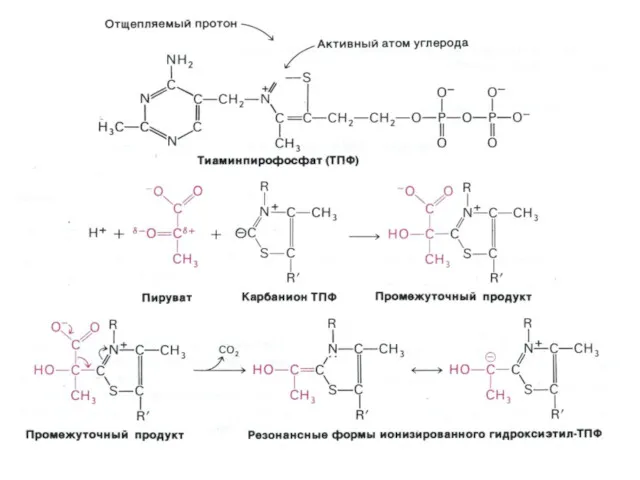
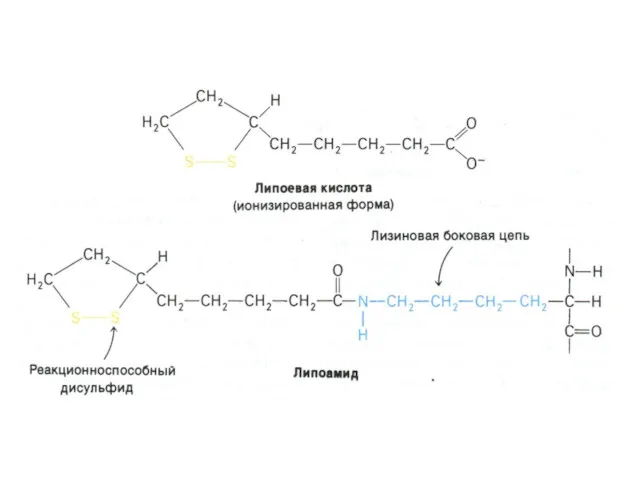
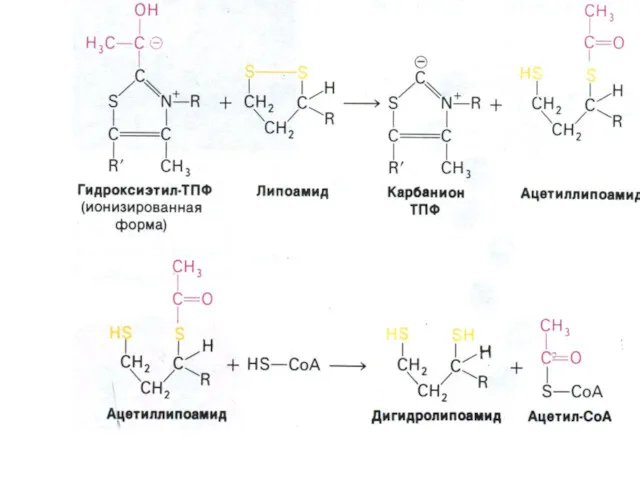

Окисление пирувата
Необратимая реакция.
Продукт окисления и декарбоксилирования – ацетил-КоА.
Протон фиксируется в НАДН.
2-х
Окисление пирувата
Необратимая реакция.
Продукт окисления и декарбоксилирования – ацетил-КоА.
Протон фиксируется в НАДН.
2-х

Структура и функции КоА
Низкомолекулярный термостабильный (не связан с белком) фактор –
Структура и функции КоА
Низкомолекулярный термостабильный (не связан с белком) фактор –

Ацетил-КоА
Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 –
Ацетил-КоА
Ацетил - КоА – универсальное клеточное топливо, окисляется до СО2 –

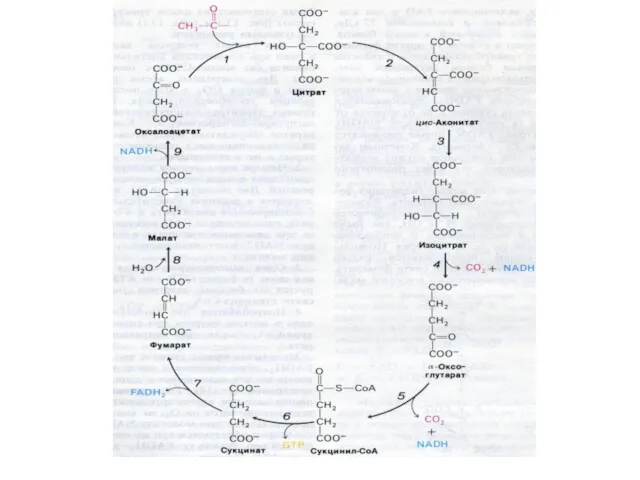
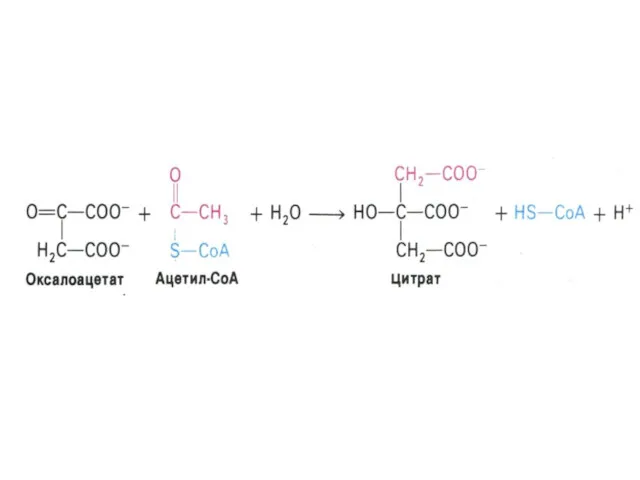
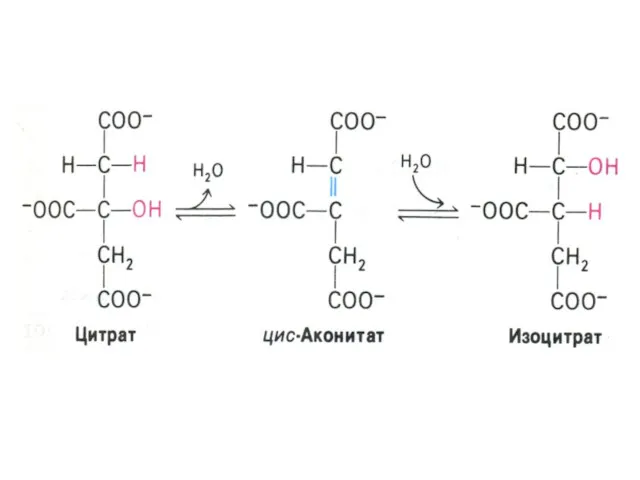
Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса.
8 реакций, замкнутых в цикл
Цикл трикарбоновых кислот (лимонной кислоты), цикл Кребса.
8 реакций, замкнутых в цикл
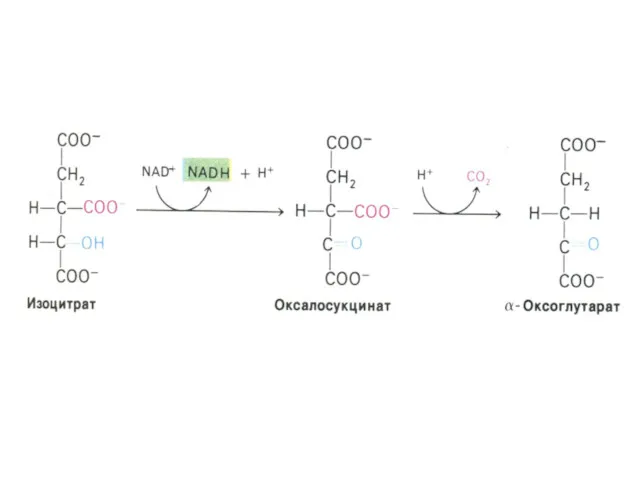

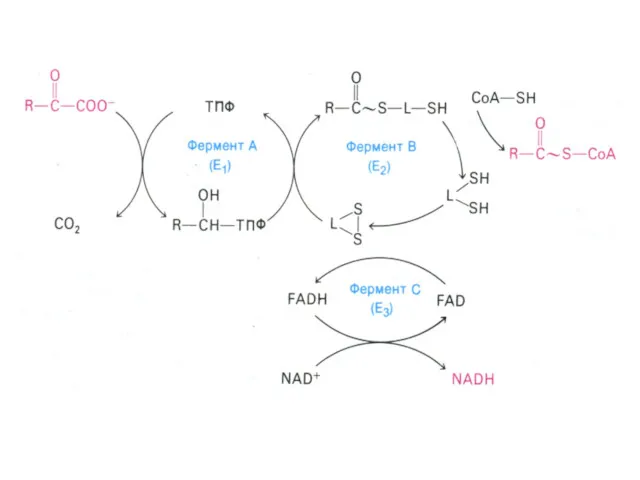
α –кетоглутаратдегидрогеназный комплекс
Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному
α –кетоглутаратдегидрогеназный комплекс
Мультиферментный комплекс, осуществляющий декарбоксилирование и окисление кетоглутарата, аналогично пируватдегидрогеназному

Субстратное фосфорилирование
Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования
Субстратное фосфорилирование
Сукцинил-КоА содержит энергию тиоэфирной связи, достаточную для обеспечения реакции фосфорилирования
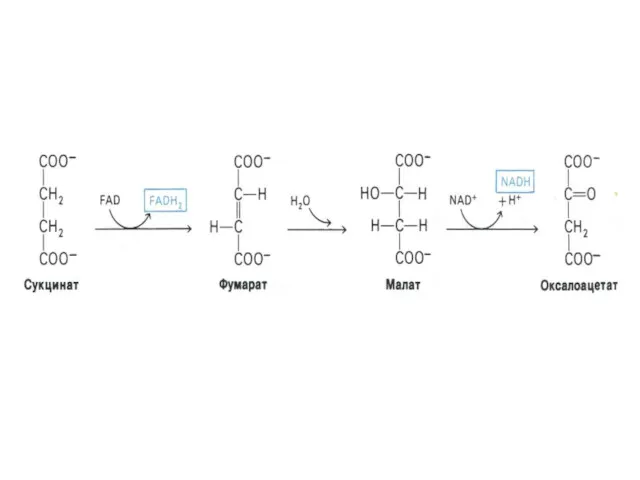
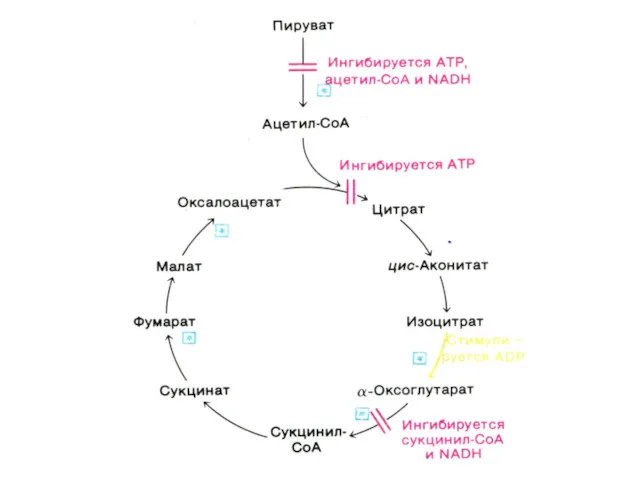

Функции цикла Кребса
Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса
Функции цикла Кребса
Водороддонорная (генерация 4-х пар протонов, поступающих в цепь переноса

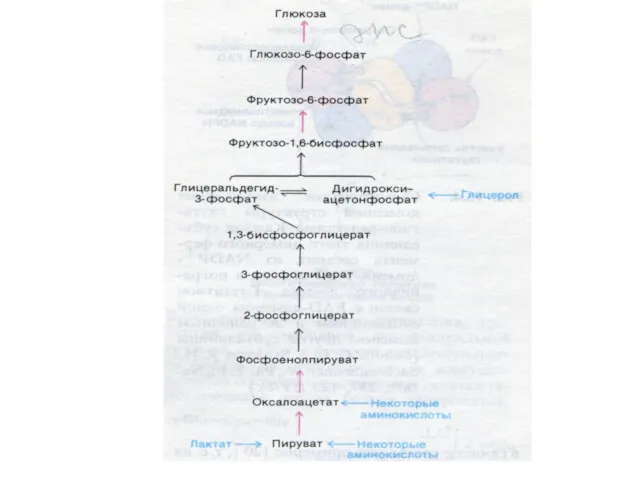
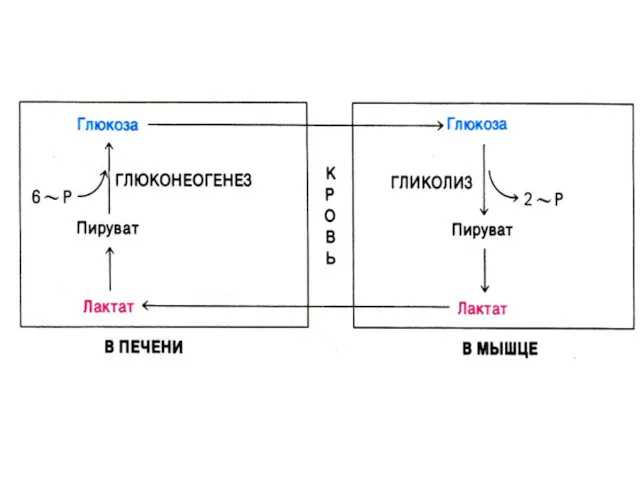
Глюконеогенез
Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК).
4 адаптивных
Глюконеогенез
Образование глюкозы из неуглеводных предшественников (лактата, глицерина, аминокислот, метаболитов ЦТК).
4 адаптивных
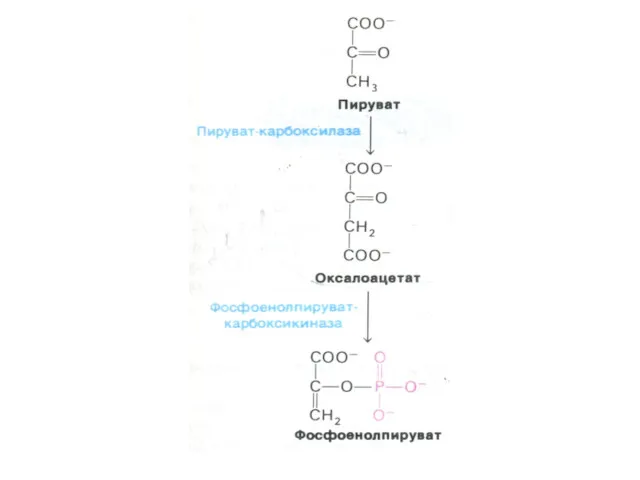
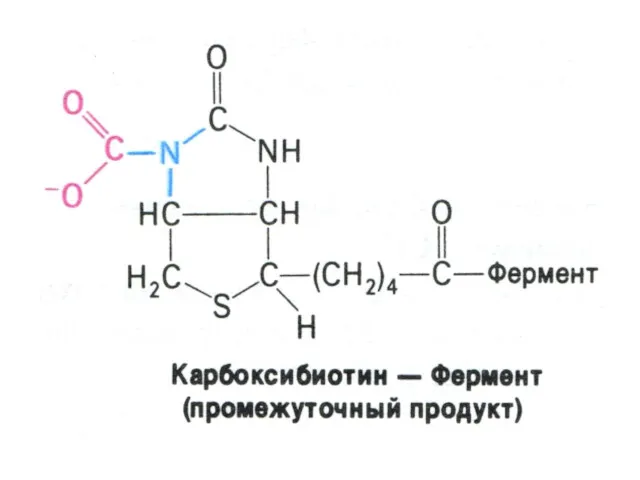
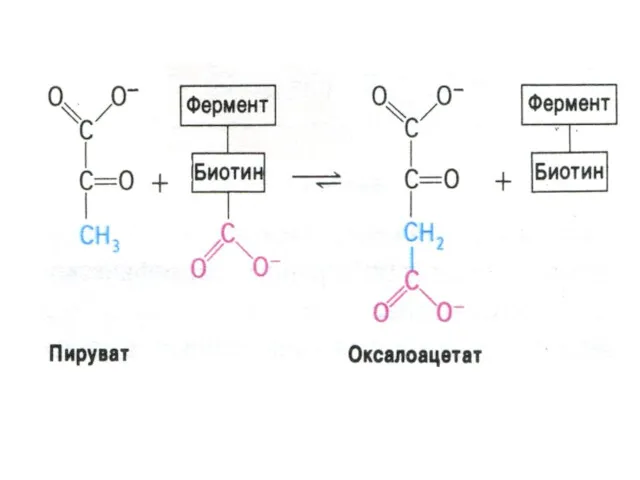

Перемещение оксалоацетата из митохондрий в цитозоль
Пируват, проникнув в митохондрии, карбоксилируется до
Перемещение оксалоацетата из митохондрий в цитозоль
Пируват, проникнув в митохондрии, карбоксилируется до

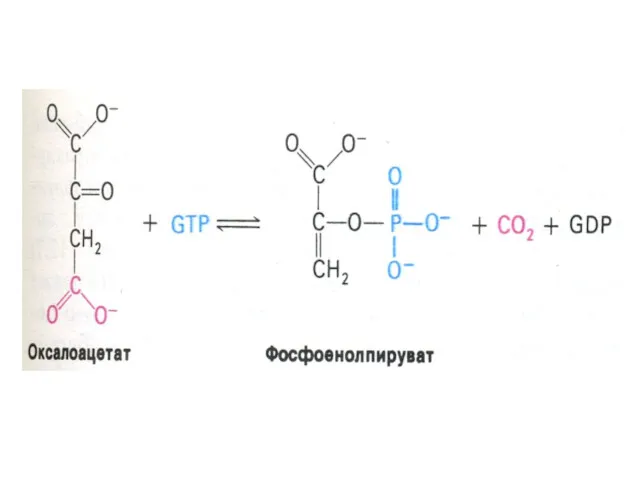
Ферменты глюконеогенеза
Активны в печени, мозге, почках. Нет в скелетных мышцах.
ФЭП- карбоксикиназа;
Ферменты глюконеогенеза
Активны в печени, мозге, почках. Нет в скелетных мышцах.
ФЭП- карбоксикиназа;

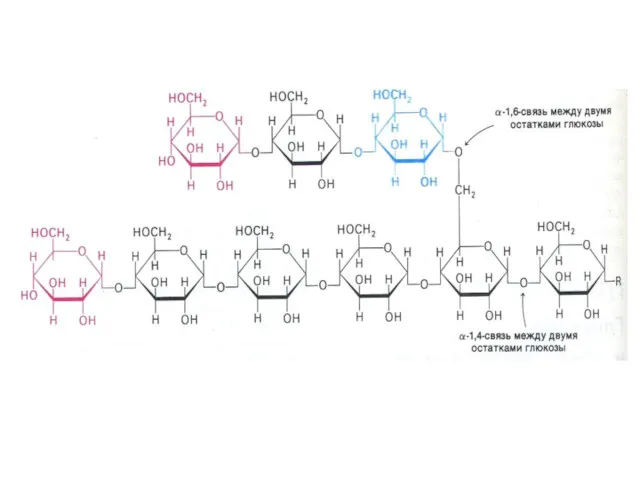
Метаболизм гликогена
Запасы гликогена в клетках разных тканей. Значение. Структура гликогена.
Синтез гликогена.
Метаболизм гликогена
Запасы гликогена в клетках разных тканей. Значение. Структура гликогена.
Синтез гликогена.

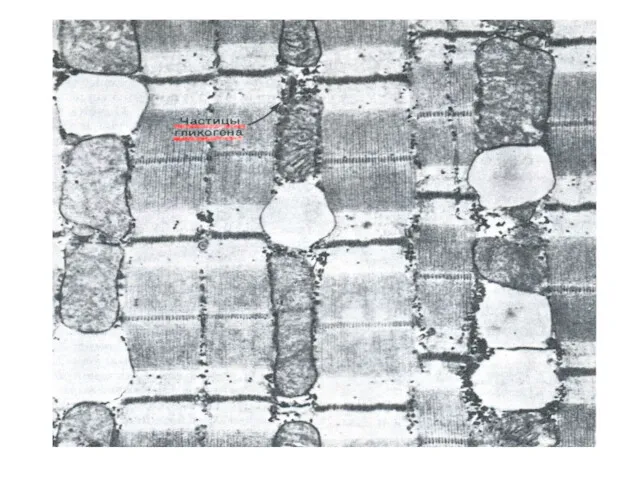

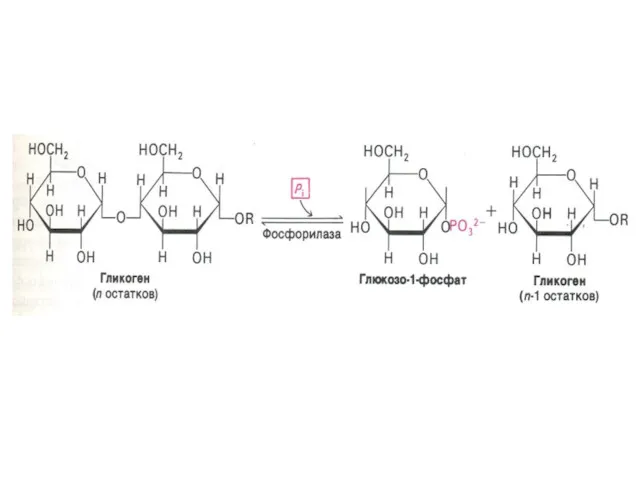
Фосфоролиз гликогена
В отличие от внешнего обмена (ЖКТ) , в клетке гликоген
Фосфоролиз гликогена
В отличие от внешнего обмена (ЖКТ) , в клетке гликоген
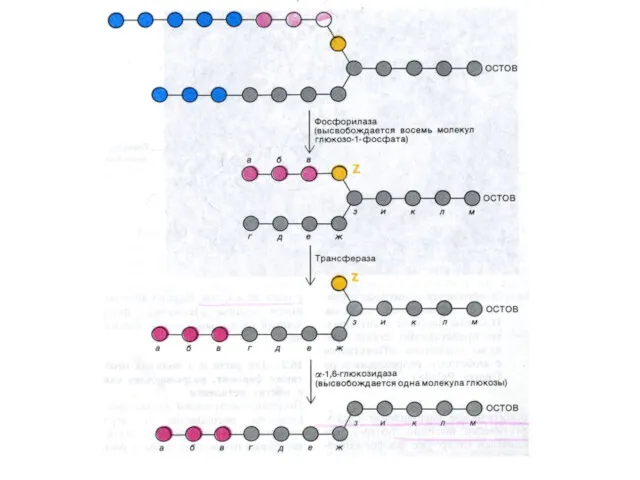
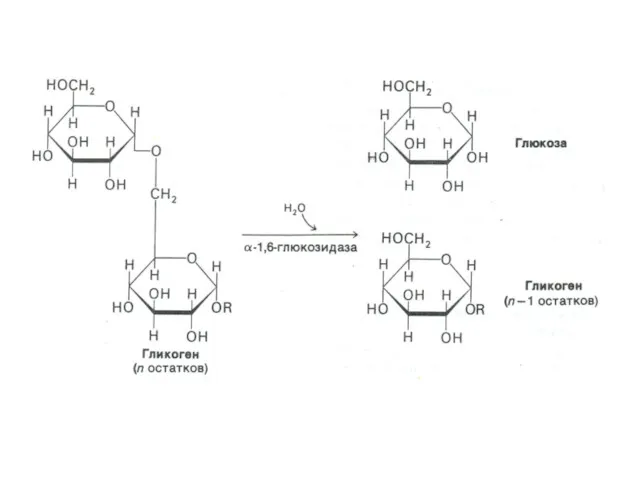

Синтез гликогена
Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1
Синтез гликогена
Субстратом для синтеза гликогена является активированная форма глюкозы УДФ -1

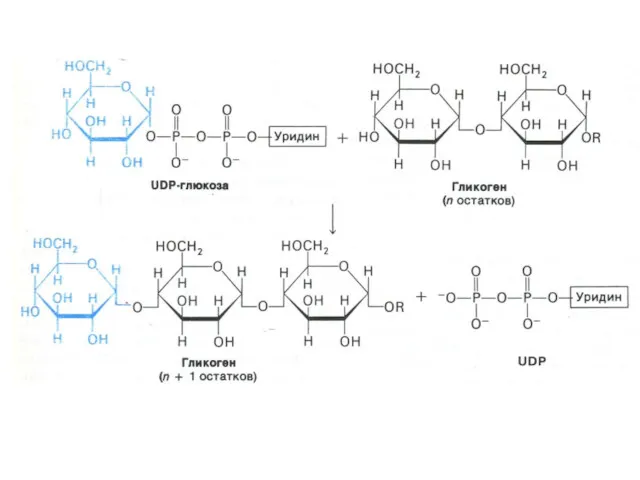


Синтез гликогена
Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6
Синтез гликогена
Синтез гликогена осуществляется ферментами: ГЛИКОГЕНСИНТАЗОЙ и амило -1,4 - 1,6

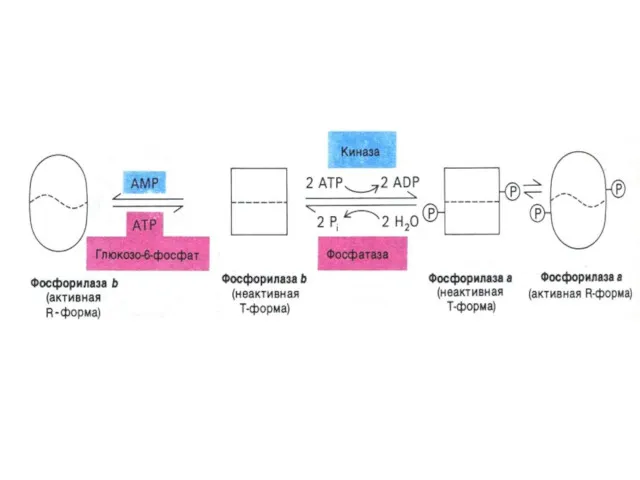
Регуляция метаболизма гликогена
Синтез и распад гликогена регулируются по одним и тем
Регуляция метаболизма гликогена
Синтез и распад гликогена регулируются по одним и тем

Регуляция метаболизма гликогена
Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется
Регуляция метаболизма гликогена
Гликогенсинтаза ингибируется фосфорилированием при участии фосфопротеинкиназы и активируется

Регуляция метаболизма гликогена
В мышцах:
В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ;
При слабых
Регуляция метаболизма гликогена
В мышцах:
В состоянии покоя гликогенфосфорилаза аллостерически активируется АМФ;
При слабых

 Шляпочные грибы
Шляпочные грибы Азбука цветов Кемеровской области
Азбука цветов Кемеровской области Органы чувств
Органы чувств Позвоночный столб вид спереди
Позвоночный столб вид спереди Жабынды эпителий. Бездер. Қан және лимфа
Жабынды эпителий. Бездер. Қан және лимфа Типи поживних речовин
Типи поживних речовин Технология выращивания цветочно-декоративных культур
Технология выращивания цветочно-декоративных культур Макроэволюция. Доказательство макроэволюции
Макроэволюция. Доказательство макроэволюции Реактивы для молекулярной, клеточной биологии и биохимии
Реактивы для молекулярной, клеточной биологии и биохимии Выращивание лука в домашних условиях
Выращивание лука в домашних условиях охорона первоцвітів. Збереження ранніх квітучих рослин
охорона первоцвітів. Збереження ранніх квітучих рослин Сердечнососудистая система
Сердечнососудистая система История изучения клетки 10 класс
История изучения клетки 10 класс Гены, хромосомы, вирусы
Гены, хромосомы, вирусы Проекция ядер ЧМН
Проекция ядер ЧМН Контрольно-обобщающий урок по теме Класс рыб
Контрольно-обобщающий урок по теме Класс рыб Презентация к уроку биологии 7 класс
Презентация к уроку биологии 7 класс Приспособления организмов к совместной жизни
Приспособления организмов к совместной жизни Влияние биостимуляторов роста на развитие и рост растений
Влияние биостимуляторов роста на развитие и рост растений Брусника, голубика, клюква и черника
Брусника, голубика, клюква и черника Структура центра терморегуляции. Роль центра в регуляции теплообмена
Структура центра терморегуляции. Роль центра в регуляции теплообмена Дикие животные тропиков
Дикие животные тропиков Разведение по линиям
Разведение по линиям Масти лошадей
Масти лошадей Дагестанский заповедник у Кизлярского залива
Дагестанский заповедник у Кизлярского залива Полушария мозга
Полушария мозга Овощеводство. Размножение овощных растений
Овощеводство. Размножение овощных растений Раздел Билатеральносимметричные (Bilateria). Подраздел Бесполостные (Acoelomata)
Раздел Билатеральносимметричные (Bilateria). Подраздел Бесполостные (Acoelomata)