- Visual Processing

Содержание
- 2. Visual Processing The Retina The retina lies at the back of the eye Retinal tissue is
- 4. Visual Processing Photoreceptors Visual processing begins in the retina with the division of the sensory receptors
- 5. Visual Processing Photoreceptors The rods and cones differ in three main ways. First, they contain different
- 6. Visual Processing Photoreceptors There are three different types of cones, each containing a different pigment. The
- 7. Visual Processing Photoreceptors Second, the distribution of rods and cones across the retina also differs. Cones
- 8. Visual Processing Photoreceptors Finally, rods and cones are hooked up to the retina’s output layer of
- 9. Visual Processing Photoreceptors The differences in how rods and cones are wired up to other cells
- 10. Visual Processing Photoreceptors By having less summation across multiple photoreceptors, the cone system preserves more fine-grained
- 11. Visual Processing Ganglion Cells Whereas the rods and cones are the “input” part of the retina,
- 12. Visual Processing Ganglion Cells Retinal ganglion cells come in two main types M (midget) cells P
- 13. Visual Processing Pathways from the Retina to the Brain There are two main destinations for visual
- 14. Visual Processing Pathways from the Retina to the Brain The Tectopulvinar Pathway The tectopulvinar path allows
- 15. Visual Processing Pathways from the Retina to the Brain The Tectopulvinar Pathway It is also a
- 16. Visual Processing Pathways from the Retina to the Brain The Geniculostriate Pathway Approximately 90% of optic
- 17. Visual Processing The Geniculostriate Pathway Information from the right sides of both retinas is sent on
- 18. Visual Processing Primary Visual Cortex (Striate Cortex or V1) The first destination within the cortex is
- 19. Visual Processing Primary Visual Cortex (Striate Cortex or V1) The V1 contains a map that is
- 20. Visual Processing Visual Areas beyond the Striate Cortex Striate cortex provides a representation of numerous features
- 21. Visual Processing Dorsal and ventral streams for visual information The striate cortex projects both “downward,” (ventrally),
- 23. Скачать презентацию
Visual Processing
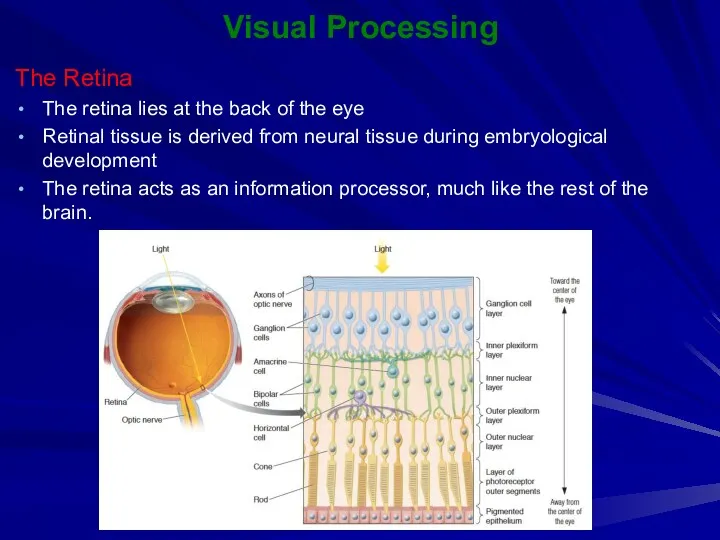
The Retina
The retina lies at the back of the eye
Visual Processing
The Retina
The retina lies at the back of the eye

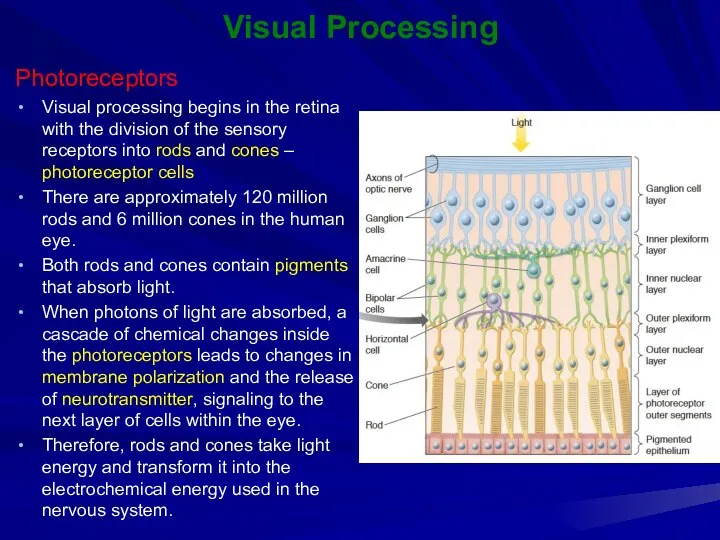
Visual Processing
Photoreceptors
Visual processing begins in the retina with the division of
Visual Processing
Photoreceptors
Visual processing begins in the retina with the division of
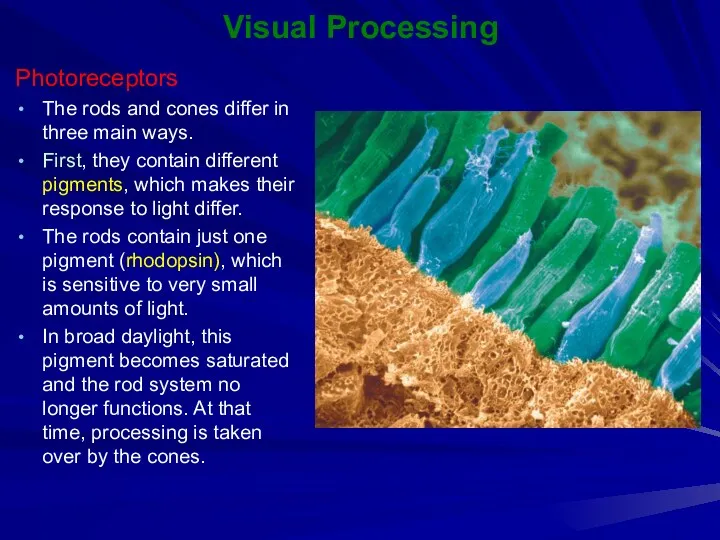
Visual Processing
Photoreceptors
The rods and cones differ in three main ways.
First,
Visual Processing
Photoreceptors
The rods and cones differ in three main ways.
First,
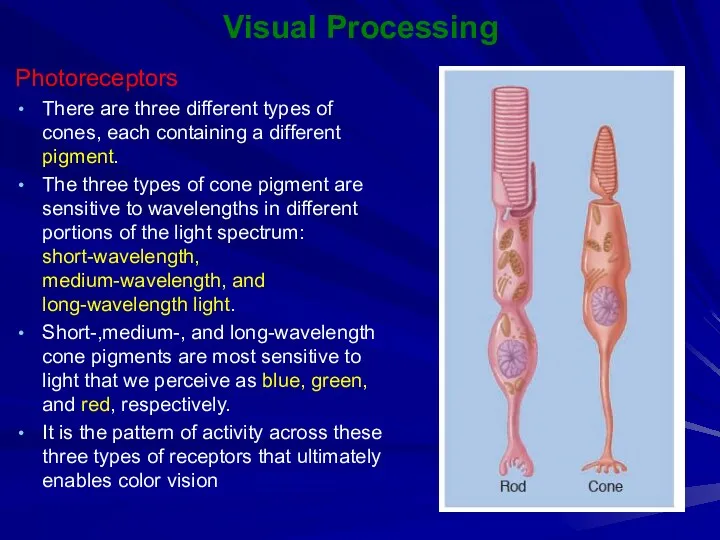
Visual Processing
Photoreceptors
There are three different types of cones, each containing a
Visual Processing
Photoreceptors
There are three different types of cones, each containing a
Visual Processing
Photoreceptors
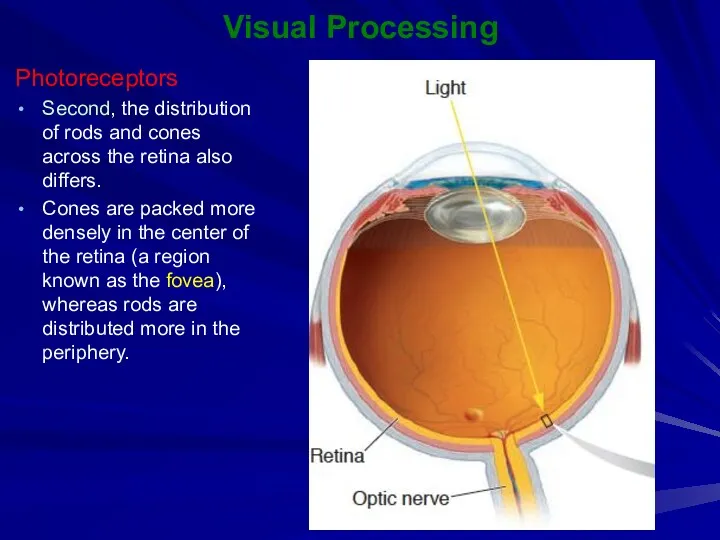
Second, the distribution of rods and cones across the retina
Visual Processing
Photoreceptors
Second, the distribution of rods and cones across the retina
Visual Processing
Photoreceptors
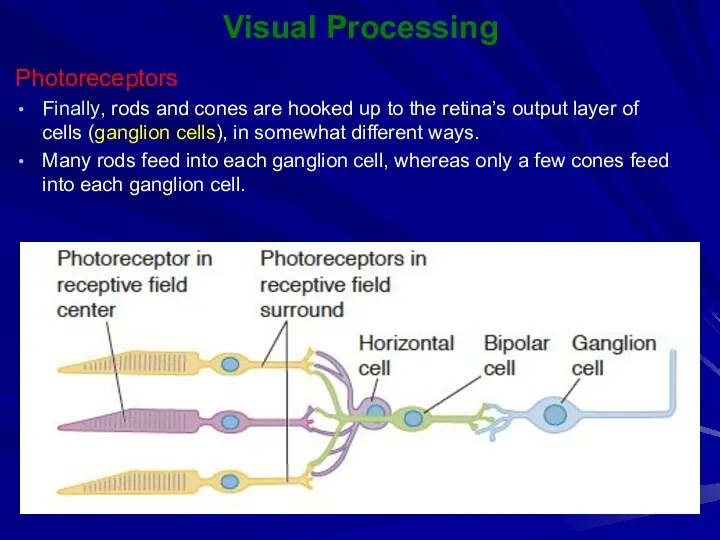
Finally, rods and cones are hooked up to the retina’s
Visual Processing
Photoreceptors
Finally, rods and cones are hooked up to the retina’s
Visual Processing
Photoreceptors
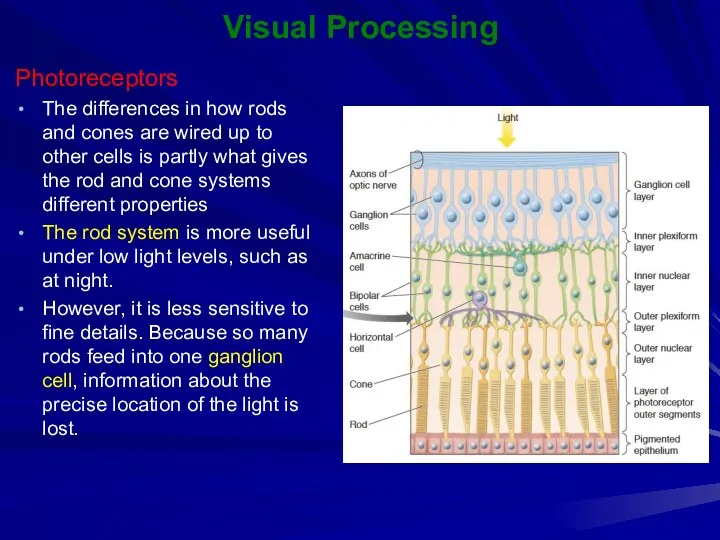
The differences in how rods and cones are wired up
Visual Processing
Photoreceptors
The differences in how rods and cones are wired up
Visual Processing
Photoreceptors
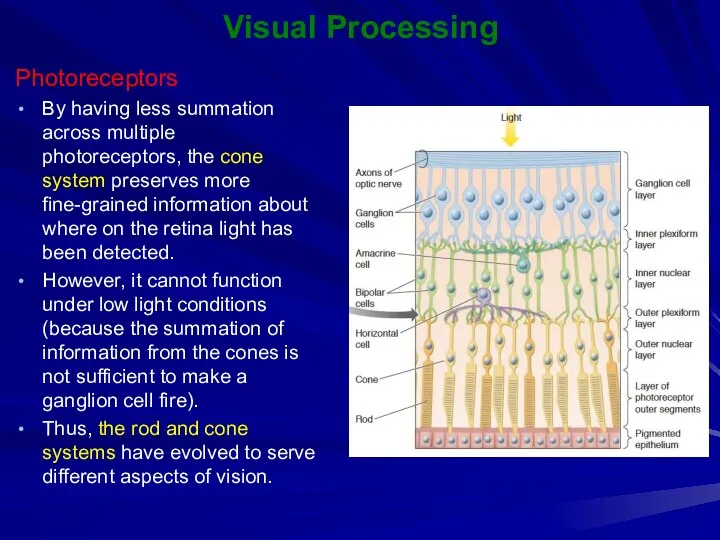
By having less summation across multiple photoreceptors, the cone system
Visual Processing
Photoreceptors
By having less summation across multiple photoreceptors, the cone system
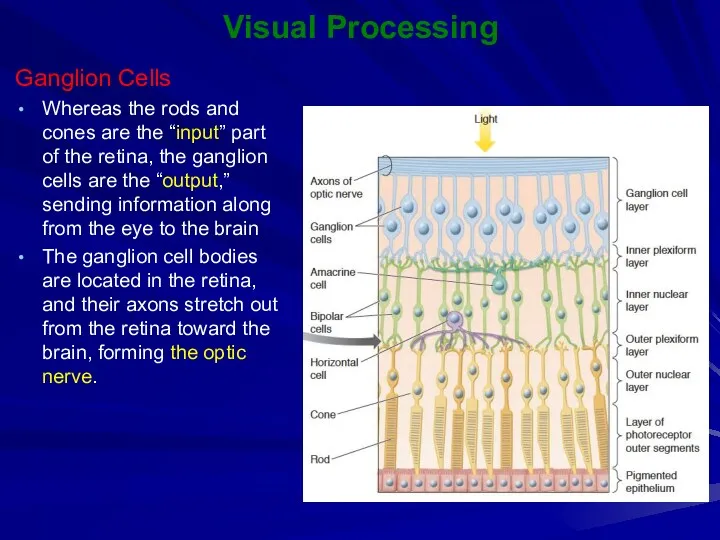
Visual Processing
Ganglion Cells
Whereas the rods and cones are the “input” part
Visual Processing
Ganglion Cells
Whereas the rods and cones are the “input” part
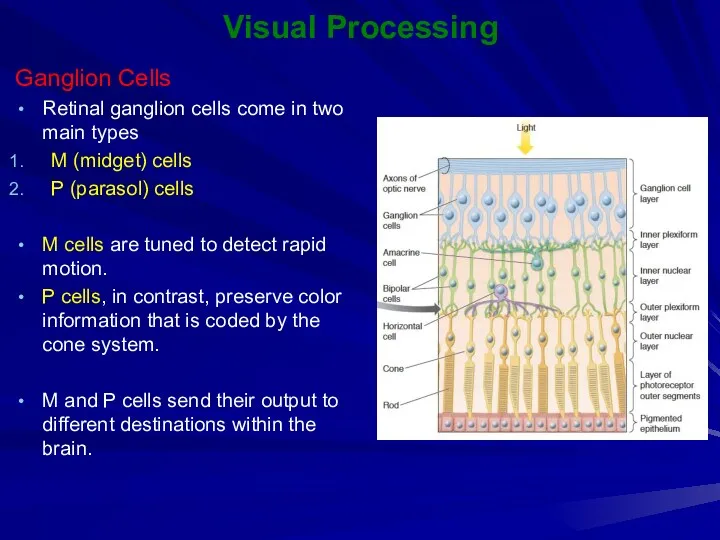
Visual Processing
Ganglion Cells
Retinal ganglion cells come in two main types
M (midget)
Visual Processing
Ganglion Cells
Retinal ganglion cells come in two main types
M (midget)
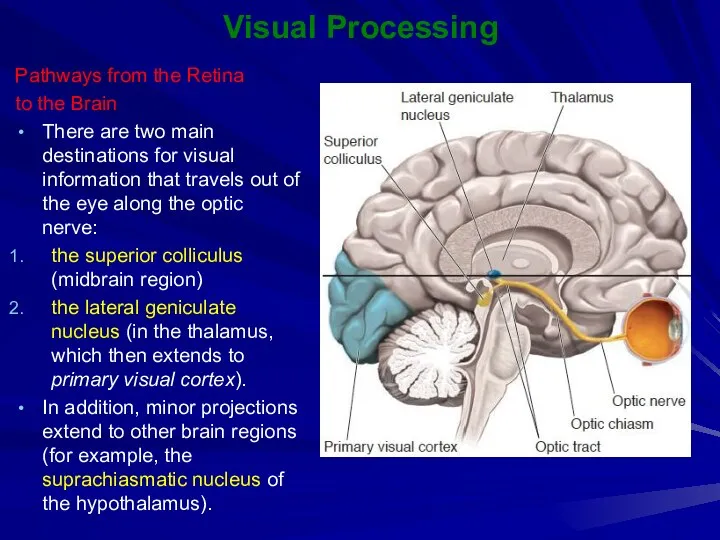
Visual Processing
Pathways from the Retina
to the Brain
There are two main destinations
Visual Processing
Pathways from the Retina
to the Brain
There are two main destinations
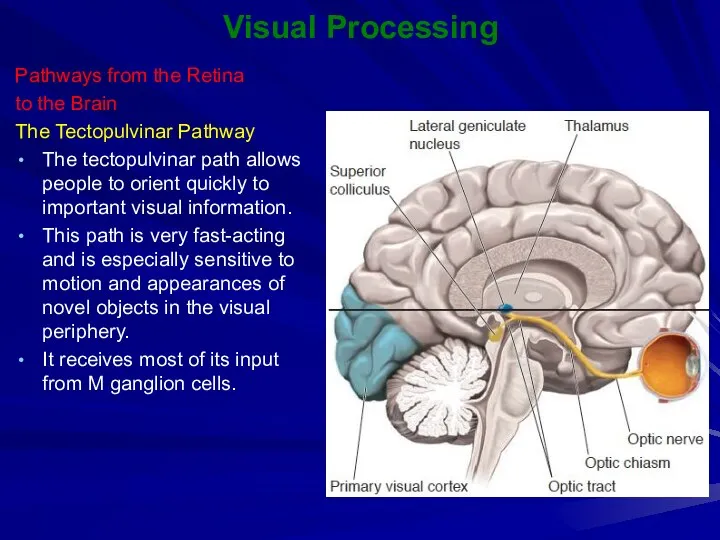
Visual Processing
Pathways from the Retina
to the Brain
The Tectopulvinar Pathway
The tectopulvinar path
Visual Processing
Pathways from the Retina
to the Brain
The Tectopulvinar Pathway
The tectopulvinar path
Visual Processing
Pathways from the Retina
to the Brain
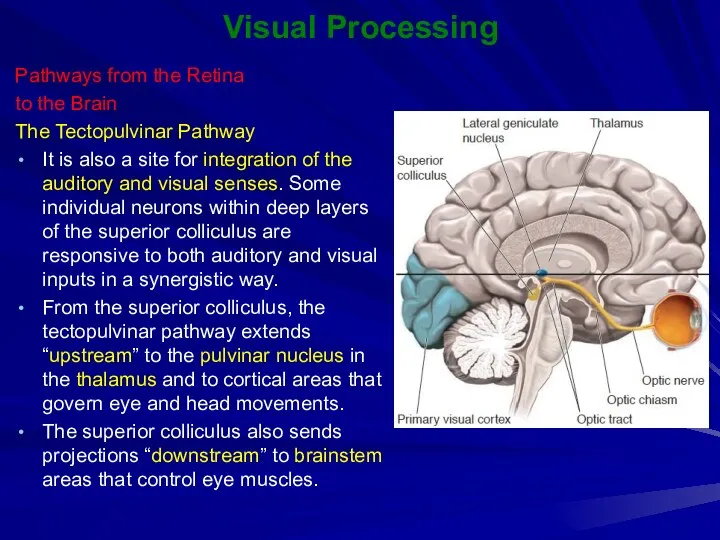
The Tectopulvinar Pathway
It is also
Visual Processing
Pathways from the Retina
to the Brain
The Tectopulvinar Pathway
It is also
Visual Processing
Pathways from the Retina to the Brain
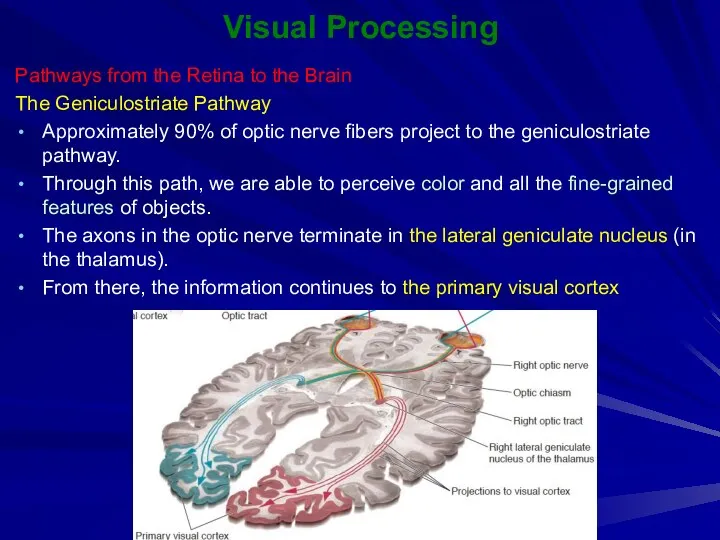
The Geniculostriate Pathway
Approximately 90%
Visual Processing
Pathways from the Retina to the Brain
The Geniculostriate Pathway
Approximately 90%
Visual Processing
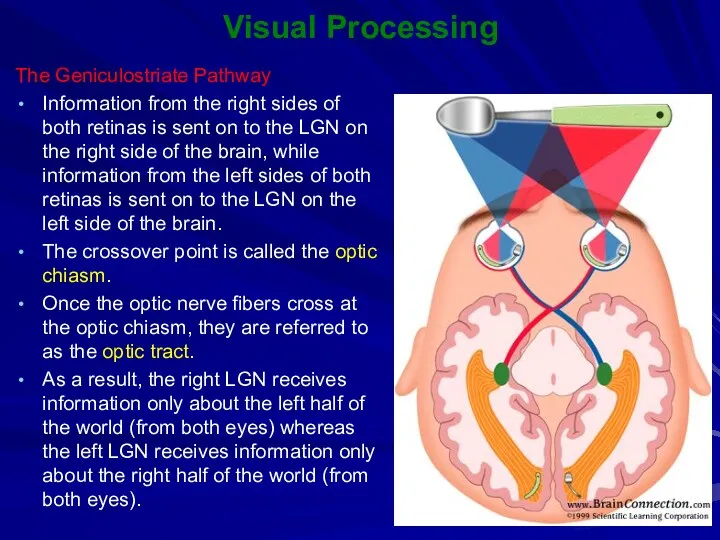
The Geniculostriate Pathway
Information from the right sides of both retinas
Visual Processing
The Geniculostriate Pathway
Information from the right sides of both retinas
Visual Processing
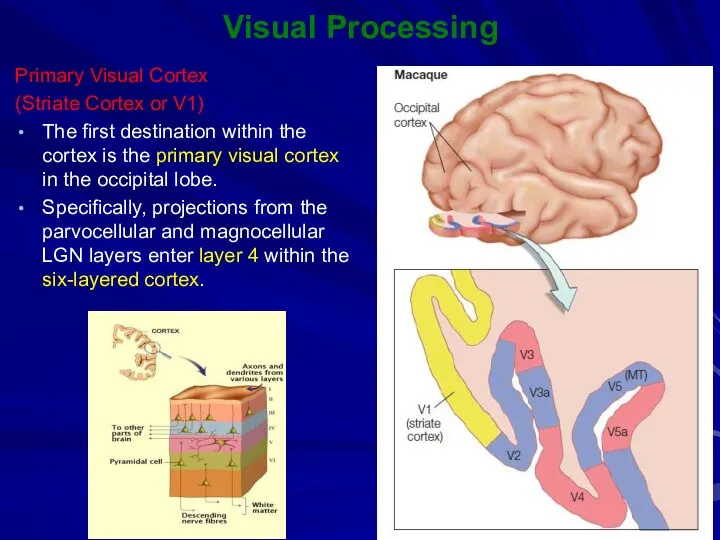
Primary Visual Cortex
(Striate Cortex or V1)
The first destination within the
Visual Processing
Primary Visual Cortex
(Striate Cortex or V1)
The first destination within the

Visual Processing
Primary Visual Cortex (Striate Cortex or V1)
The V1 contains a
Visual Processing
Primary Visual Cortex (Striate Cortex or V1)
The V1 contains a
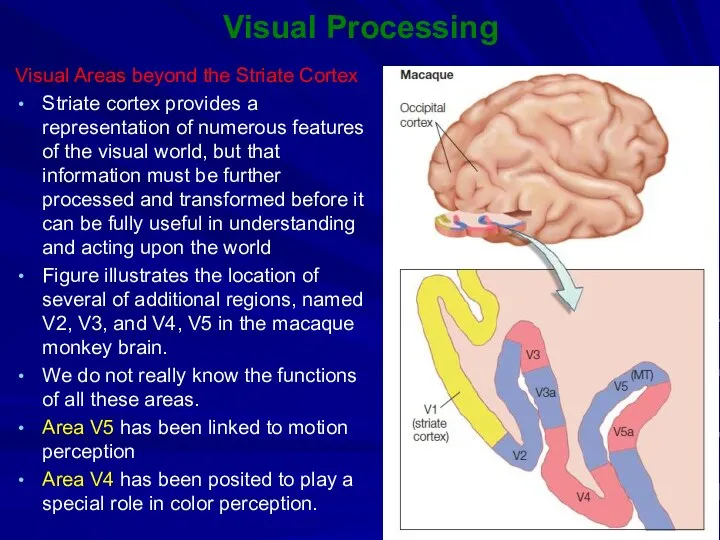
Visual Processing
Visual Areas beyond the Striate Cortex
Striate cortex provides a representation
Visual Processing
Visual Areas beyond the Striate Cortex
Striate cortex provides a representation
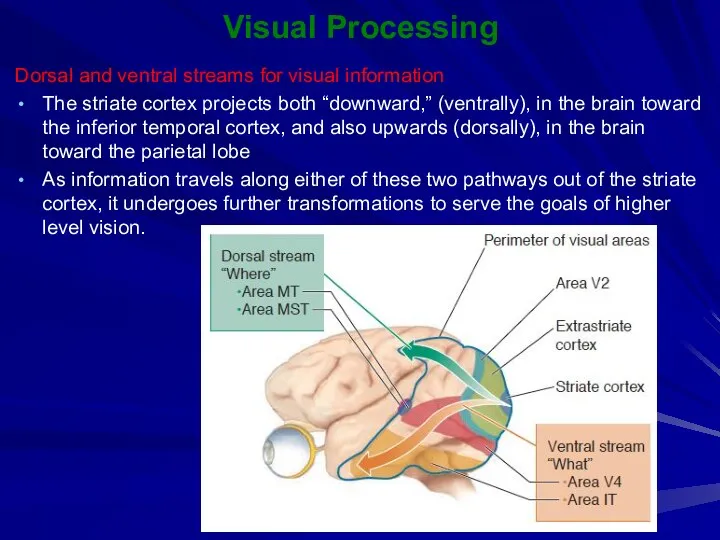
Visual Processing
Dorsal and ventral streams for visual information
The striate cortex projects
Visual Processing
Dorsal and ventral streams for visual information
The striate cortex projects
 Детёныши животных (фотографии)
Детёныши животных (фотографии) Среда обитания растений - фитоценозы
Среда обитания растений - фитоценозы Газоны. Виды газонов
Газоны. Виды газонов Эволюционные теории и взгляды
Эволюционные теории и взгляды Тұзды ортаға төзімді өсімдіктер
Тұзды ортаға төзімді өсімдіктер Нарушения кислотно-щелочного равновесия
Нарушения кислотно-щелочного равновесия Аквариумные рыбки гуппи
Аквариумные рыбки гуппи Движение крови по сосудам. Круги кровообращения
Движение крови по сосудам. Круги кровообращения Рудиментарные органы человека
Рудиментарные органы человека Обмен липидов. (Часть 2)
Обмен липидов. (Часть 2) Чихуахуа
Чихуахуа Водоросли. Общая характеристика водорослей
Водоросли. Общая характеристика водорослей Способы опыления у цветковых растений
Способы опыления у цветковых растений Органы чувств
Органы чувств Ветеринарная диетология
Ветеринарная диетология Спинной мозг
Спинной мозг Урок по ФГОС . История изучения клетки. Методы изучения клетки. Лабораторная работа Строение клеток кожицы чешуи лука.
Урок по ФГОС . История изучения клетки. Методы изучения клетки. Лабораторная работа Строение клеток кожицы чешуи лука. Исследовательский практикум Цветение талой воды
Исследовательский практикум Цветение талой воды Петуния. Выращивание и уход за петунией
Петуния. Выращивание и уход за петунией Орган слуха и равновесия
Орган слуха и равновесия Строение цветка. Соцветие. Значение оплодотворения
Строение цветка. Соцветие. Значение оплодотворения Ленточные черви
Ленточные черви Выращивание и высадка рассады в цветник
Выращивание и высадка рассады в цветник Организация и проведение мероприятий по воспроизводству лесов и лесоразведению
Организация и проведение мероприятий по воспроизводству лесов и лесоразведению Животные Эстонии
Животные Эстонии Антропология тарихына шолу
Антропология тарихына шолу Важнейшие микробиологические процессы. Виды брожения
Важнейшие микробиологические процессы. Виды брожения Метод выделения и разделения лейкоцитов периферической крови в градиенте плотности фиколл-гипак
Метод выделения и разделения лейкоцитов периферической крови в градиенте плотности фиколл-гипак