- Водно-солевой обмен у животных, освоивших сушу

Содержание
- 2. Жизнь на суше Доступность О2 2) Опасность высыхания В широком масштабе переход на сушу совершили Позвоночные
- 3. Проблема потери Н2О Физические факторы, влияющие на испарение. Испарение с водной поверхности > при > Ta.
- 4. Давление водяных паров над свободной водной поверхностью быстро возрастает с повышением температуры (белые кружки, жирная линия).
- 5. Другие факторы, влияющие на скорость испарения воды: Свободная или принудительная конвекция; Барометрическое давление; Размер и форма
- 6. Животные с влажной и сухой кожей «Система, ограниченная паром» (Vapor-limited system) С влажной поверхностью ? интенсивное
- 7. Испарение воды с поверхности тела различных животных при комнатной температуре. Цифры показывают порядок величины; точные значения
- 8. ПАРАДОКС: некоторые амфибии приспособились к жизни в пустыне! Лягушки рода Chiroleptes в Австралии («австралийские жабы»). В
- 9. [испарение с кожи у австралийских лягушек ~= обычным. Лягушки ~~ = жабам с > ороговевшей кожей].
- 10. У лягушки Chiromantis при 25оС и относительной влажности 30% испарение воды идет очень медленно так же,
- 11. б) У южноамериканской квакши р. Phyllomedusa испарение воды с кожи = 1/20 у др. видов. Другой
- 12. Наземные позвоночные Членистоногие Амниоты Водный баланс Поступление воды Потеря воды Питье Поглощение через S (из воды,
- 13. Разная выносливость к потерям воды при жизни в воде и на суше: Млекопитающие: до 10% потерь
- 14. Поступление и потеря воды в засушливых условиях Поступление Питье - может не быть свободной воды; Поглощение
- 15. а) окисление глюкозы C6H12O6 + 6O2 = 6CO2 +6H2O 180 г 192 г 264 г 108
- 16. в) окисление белка. Азот белка образует экскреты, содержащие водород. Этот водород уходит и не окисляется до
- 17. Количество воды, образующееся при окислении различных пищевых веществ (King, 1957; Schmidt-Nielsen, 1964)
- 18. Экскреция аммиака в онтогенезе наземной жабы Bufo bufo и водной шпорцевой лягушки Xenopus laevis, % от
- 19. Рептилии, 4 отряда Крокодилы Змеи Ящерицы Черепахи местообитания (вода) (…+ сухие) (…+ сухие) (…+ сухие) Кожа
- 20. Испарение воды с поверхности тела рептилий при 23-25о С. Все данные выражены в микрограммах воды на
- 21. ?? У рептилий с влажных поверхностей дыхательных путей испаряется > воды, чем с кожи?? Это не
- 22. У рептилий испарение с сухой кожи превышает потерю влаги с поверхности дыхательных путей и составляет от
- 23. Чем суше постоянная среда обитания, тем более приспособлено животное к экономии влаги: у пустынной гремучей змеи
- 24. Рептилии: Многие выводят мочевую кислоту. Потери воды с мочой зависят от конечного продукта белкового обмена. Для
- 25. Соотношение основных форм экскреции азота у разных видов черепах, % от общего азота (по V.Moyle,1949).
- 26. Птицы и млекопитающие
- 27. Многие птицы и млекопитающие используют воду для активной испарительной теплоотдачи (полипноэ, пот). Сложно учесть эти потери
- 28. Водный баланс кенгуровой крысы в пустыне Сев. Америки Сухая пища (семена и растительные материалы). Способны ничего
- 29. Из проекта The Open University “ Animals at the extremes:…”
- 30. Общий водный баланс кенгуровой крысы за время, в течение которого животное потребляет 100 г ячменя (~
- 31. Модель теплообмена в носовых ходах (Schmidt-Nielsen, 1972). Окружающий воздух не насыщен влагой, температура его 28оС, температура
- 32. From Taylor (1977)
- 33. Температура выдыхаемого воздуха у семи видов птиц, измеренная в сухом воздухе (Schmidt-Nielsen et al., 1970). Температура
- 34. Еще о приспособлениях млекопитающих в пустыне…
- 35. Рассчитанная величина испарения, необходимого для поддержания постоянной температуры тела в жаркой пустыне у млекопитающих с различными
- 36. (пустыня в Намибии) From Williams (2001) Из проекта The Open University “ Animals at the extremes:…”
- 37. Из проекта The Open University “ Animals at the extremes:…”
- 38. Большой удодовый жаворонок Alaemon alaudipes обитает в пустынях на Аравийском полуострове, в Сирии, Афганистане, Пакистане и
- 39. Из проекта The Open University “ Animals at the extremes:…”
- 40. У лишенного воды верблюда суточные колебания ректальной температуры могут достигать 5-6оС. Если его поить ежедневно, эти
- 41. У дегидратированного верблюда, который потерял 49 л воды (32%), из межклеточной жидкости убыло значительно больше воды,
- 43. I. Остриженный верблюд (А) испаряет летом на 50% больше воды, чем покрытый шерстью (Б) II. Второй
- 46. и в тропиках…
- 47. Потовые железы у индийского зебувидного скота много крупнее, чем у европейских пород. Общее количество потовых желез
- 48. Частота дыхания при высокой внешней температуре значительно меньше у тепловыносливых тропических пород, чем у пород умеренного
- 49. Морские позвоночные, дышащие воздухом В отличие от рыб избегают многих осмотических проблем, так как дышат через
- 50. Пьют м. воду ? > %C солей в жидкостях тела и они должны выводится в %С
- 51. Выведение соли солевыми железами Рептилии – 3 отряда (ящерицы, змеи, черепахи) имеют представителей в море. Солевые
- 52. Кстати… Слезы человека изоосмотичны по отношению к плазме крови и не играют роль в выведении солей
- 53. Солевые железы морских птиц У всех морских птиц есть парные солевые железы. Расположены во впадине у
- 54. Как и у рептилий, солевые железы птиц работают в ответ на солевую нагрузку. В остальное время
- 55. Концентрация натрия в носовых выделениях различных птиц. Концентрация хлора в каждой пробе почти равна концентрации натрия;
- 56. Поразительная способность выделять соль: В опыте клуша выпивала м.воду до 1/10 от Mb (= 7 л
- 57. Выделения из носа и клоаки у клуши в течение 175 мин после введения в желудок морской
- 58. Поразительная способность выделять соль: В опыте клуша выпивала м.воду до 1/10 от Mb (= 7 л
- 59. Секреция жидкости солевой железой серебристой чайки и почками человека при максимальном водном диурезе
- 60. Поразительная способность выделять соль: В опыте клуша выпивала м.воду до 1/10 от Mb (= 7 л
- 61. Концентрация натрия и калия (в миллимолях на литр) в морской воде и в секрете носовой солевой
- 62. У морских животных солевые железы более специализированы для выделения Na+ и Cl- У наземных - состав
- 63. Концентрация ионов Cl- в экскретах некоторых видов птиц при искусственной солевой нагрузке (по M.Smith, G.Bartholomew,1966)
- 64. Морские млекопитающие Сирены Киты Ластоногие Пища разнообразна по %С. Тюлени и киты едят рыбу, крупных б/п
- 65. Они не нуждаются в солевых железах, так как их почки способны выводить мочу в %С >,
- 66. Влияние 1 л морской воды, введенной через рот, на водный баланс у человека и кита Для
- 71. Важная особенность водного баланса морских млекопитающих - высокая % С их молока повышенное количество жира и
- 72. Состав и энергетическая ценность молока млекопитающих Значения в скобках относятся к общему количеству нежирных твердых веществ,
- 74. Скачать презентацию

Жизнь на суше
Доступность О2 2) Опасность высыхания
В широком масштабе переход
Жизнь на суше
Доступность О2 2) Опасность высыхания
В широком масштабе переход
Проблема потери Н2О
Физические факторы, влияющие на испарение.
Испарение с водной поверхности >
Проблема потери Н2О
Физические факторы, влияющие на испарение.
Испарение с водной поверхности >
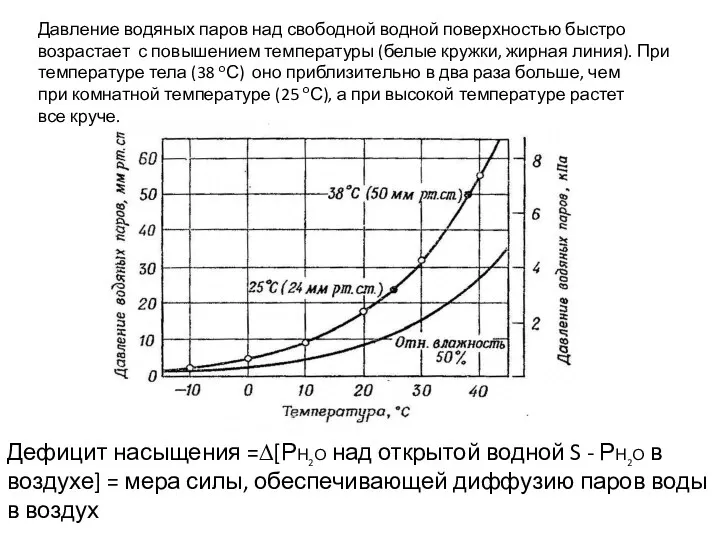
Давление водяных паров над свободной водной поверхностью быстро
возрастает с повышением
Давление водяных паров над свободной водной поверхностью быстро
возрастает с повышением

Другие факторы, влияющие на скорость испарения воды:
Свободная или принудительная конвекция;
Барометрическое давление;
Размер
Другие факторы, влияющие на скорость испарения воды:
Свободная или принудительная конвекция;
Барометрическое давление;
Размер
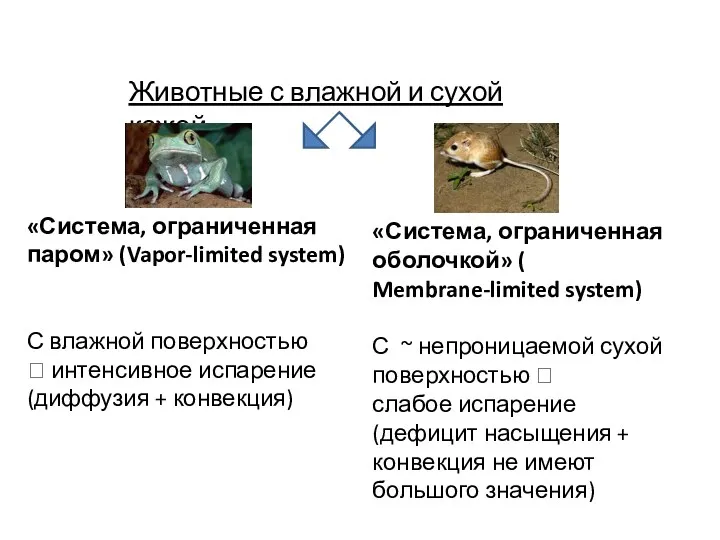
Животные с влажной и сухой кожей
«Система, ограниченная паром» (Vapor-limited system)
С влажной
Животные с влажной и сухой кожей
«Система, ограниченная паром» (Vapor-limited system)
С влажной

Испарение воды с поверхности тела различных животных при комнатной
температуре. Цифры
Испарение воды с поверхности тела различных животных при комнатной
температуре. Цифры

ПАРАДОКС: некоторые амфибии приспособились к жизни
в пустыне!
Лягушки рода
ПАРАДОКС: некоторые амфибии приспособились к жизни
в пустыне!
Лягушки рода
![[испарение с кожи у австралийских лягушек ~= обычным. Лягушки ~~ = жабам с > ороговевшей кожей].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/380722/slide-8.jpg)
< Испарения с кожи не является путем сбережения влаги у австралийских
< Испарения с кожи не является путем сбережения влаги у австралийских
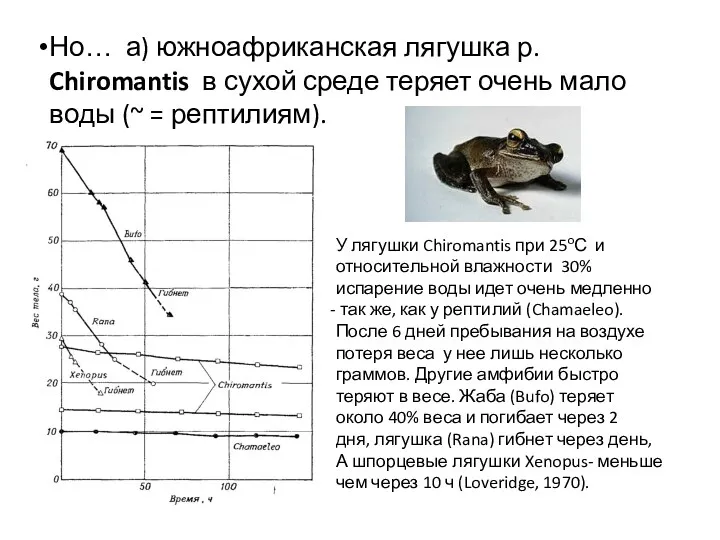
У лягушки Chiromantis при 25оС и
относительной влажности 30%
испарение воды
У лягушки Chiromantis при 25оС и
относительной влажности 30%
испарение воды

б) У южноамериканской квакши р. Phyllomedusa испарение воды с кожи =
б) У южноамериканской квакши р. Phyllomedusa испарение воды с кожи =
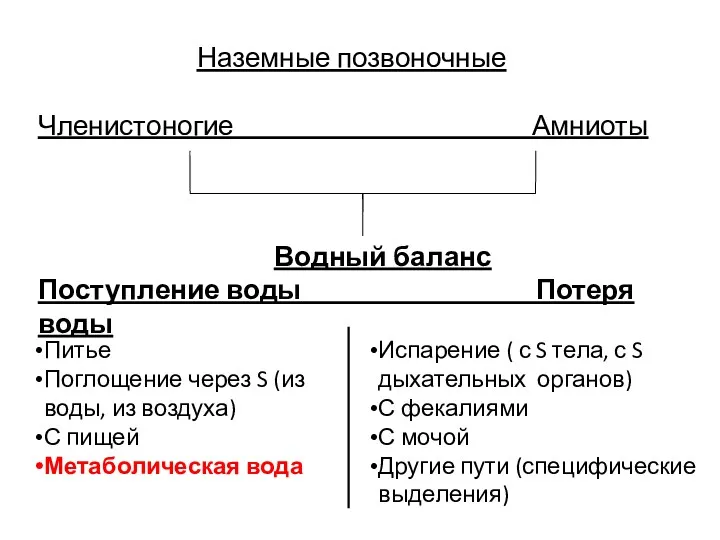
Наземные позвоночные
Членистоногие Амниоты
Водный баланс
Поступление воды Потеря воды
Питье
Поглощение через S
Наземные позвоночные
Членистоногие Амниоты
Водный баланс
Поступление воды Потеря воды
Питье
Поглощение через S

Разная выносливость к потерям воды
при жизни в воде и на
Разная выносливость к потерям воды
при жизни в воде и на

Поступление и потеря воды
в засушливых условиях
Поступление
Питье - может не
Поступление и потеря воды
в засушливых условиях
Поступление
Питье - может не

а) окисление глюкозы
C6H12O6 + 6O2 = 6CO2 +6H2O
180 г 192
а) окисление глюкозы
C6H12O6 + 6O2 = 6CO2 +6H2O
180 г 192

в) окисление белка.
Азот белка образует экскреты, содержащие водород. Этот водород уходит
в) окисление белка.
Азот белка образует экскреты, содержащие водород. Этот водород уходит
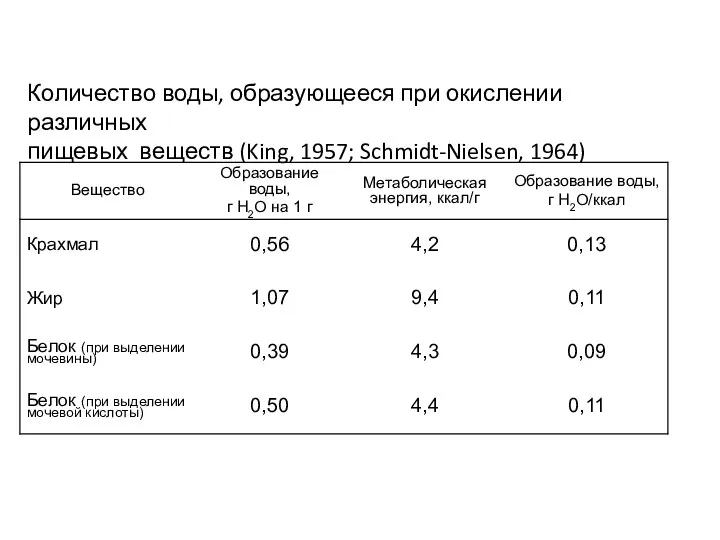
Количество воды, образующееся при окислении различных
пищевых веществ (King, 1957; Schmidt-Nielsen,
Количество воды, образующееся при окислении различных
пищевых веществ (King, 1957; Schmidt-Nielsen,

Экскреция аммиака в онтогенезе наземной жабы Bufo bufo и
водной шпорцевой
Экскреция аммиака в онтогенезе наземной жабы Bufo bufo и
водной шпорцевой
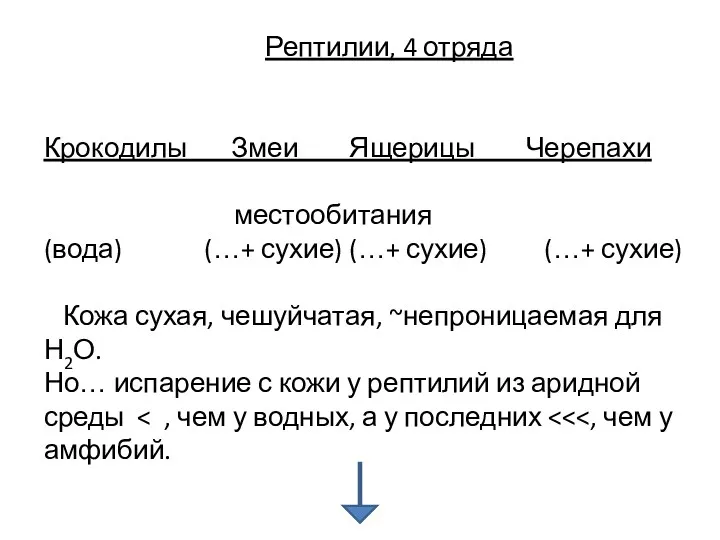
Рептилии, 4 отряда
Крокодилы Змеи Ящерицы Черепахи
местообитания
(вода) (…+ сухие) (…+ сухие)
Рептилии, 4 отряда
Крокодилы Змеи Ящерицы Черепахи
местообитания
(вода) (…+ сухие) (…+ сухие)

Испарение воды с поверхности тела рептилий при 23-25о С. Все данные
Испарение воды с поверхности тела рептилий при 23-25о С. Все данные
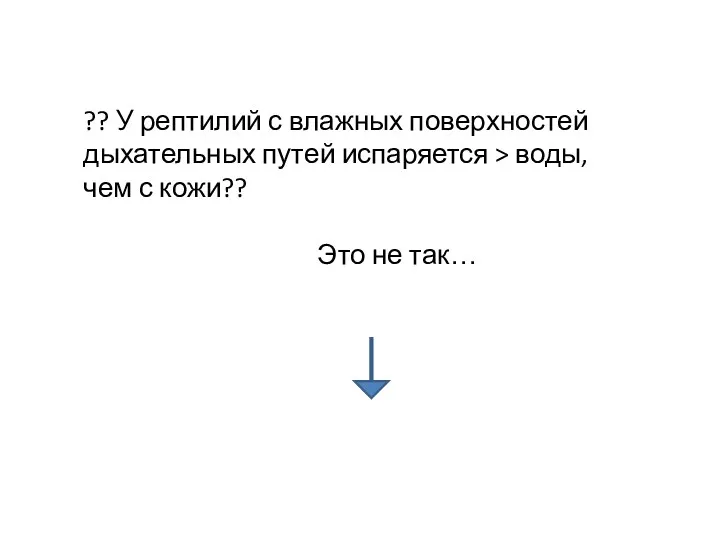
?? У рептилий с влажных поверхностей дыхательных путей испаряется > воды,
?? У рептилий с влажных поверхностей дыхательных путей испаряется > воды,
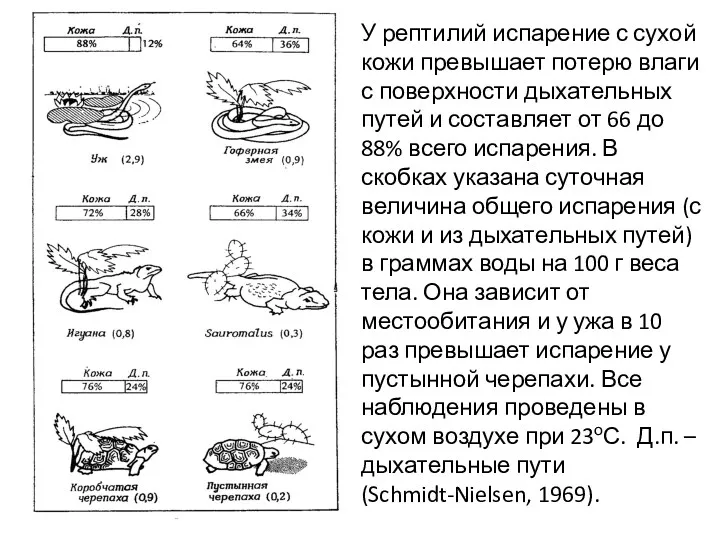
У рептилий испарение с сухой кожи превышает потерю влаги с поверхности
У рептилий испарение с сухой кожи превышает потерю влаги с поверхности

Чем суше постоянная среда обитания, тем более приспособлено животное к экономии
Чем суше постоянная среда обитания, тем более приспособлено животное к экономии

Рептилии:
Многие выводят мочевую кислоту. Потери воды с мочой зависят от
Рептилии:
Многие выводят мочевую кислоту. Потери воды с мочой зависят от
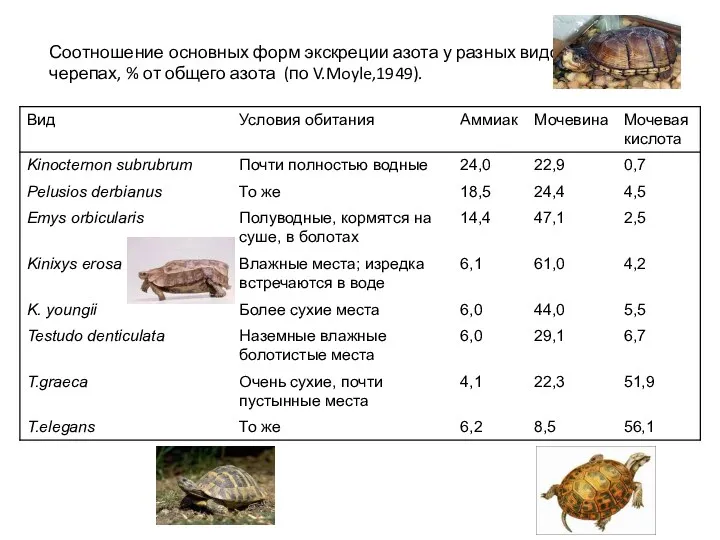
Соотношение основных форм экскреции азота у разных видов
черепах, % от общего
Соотношение основных форм экскреции азота у разных видов
черепах, % от общего

Птицы и млекопитающие
Птицы и млекопитающие

Многие птицы и млекопитающие используют воду
для активной испарительной теплоотдачи
(полипноэ,
Многие птицы и млекопитающие используют воду
для активной испарительной теплоотдачи
(полипноэ,

Водный баланс кенгуровой крысы
в пустыне Сев. Америки
Сухая пища (семена и
Водный баланс кенгуровой крысы
в пустыне Сев. Америки
Сухая пища (семена и
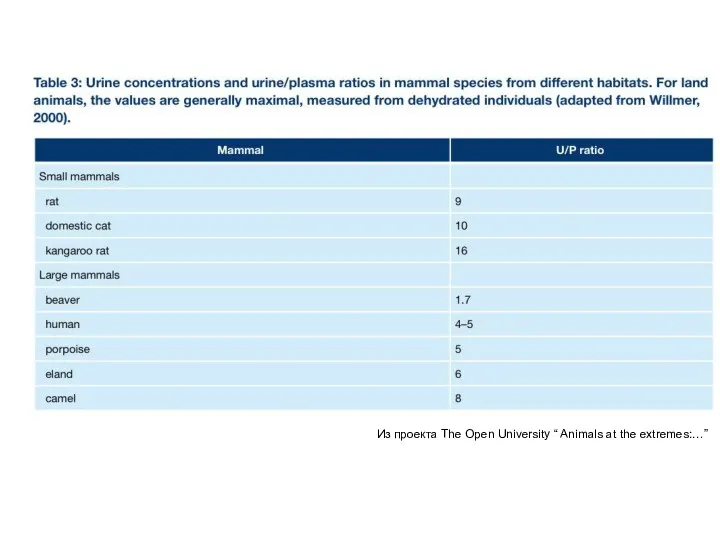
Из проекта The Open University “ Animals at the extremes:…”
Из проекта The Open University “ Animals at the extremes:…”
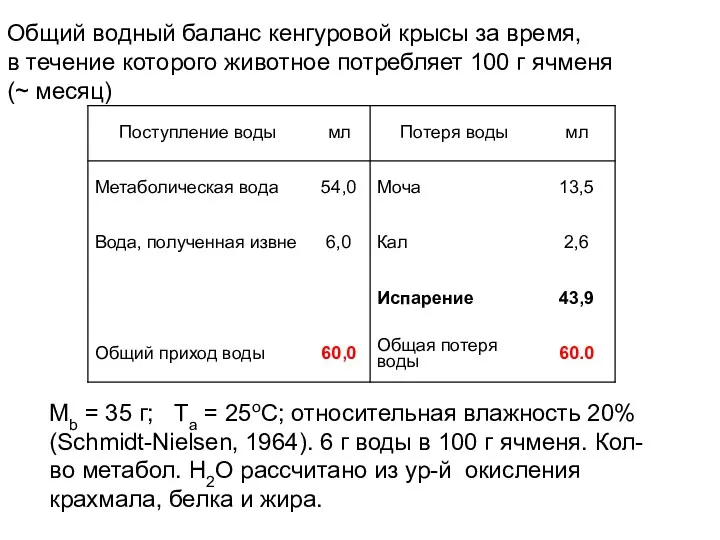
Общий водный баланс кенгуровой крысы за время,
в течение которого животное
Общий водный баланс кенгуровой крысы за время,
в течение которого животное
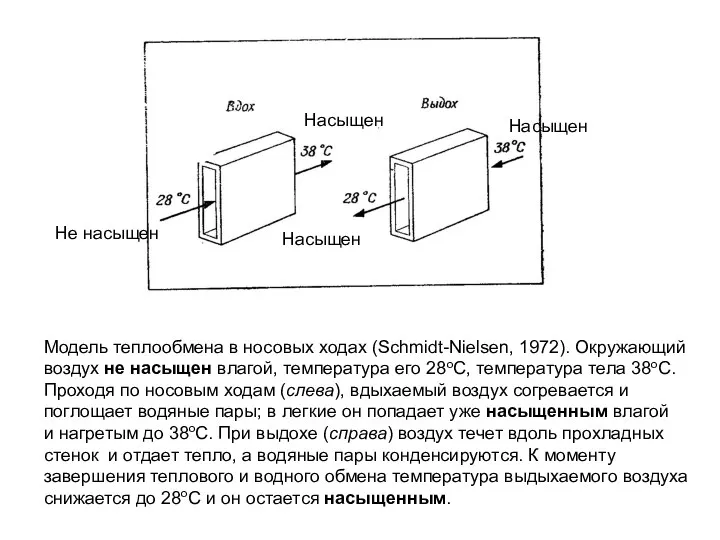
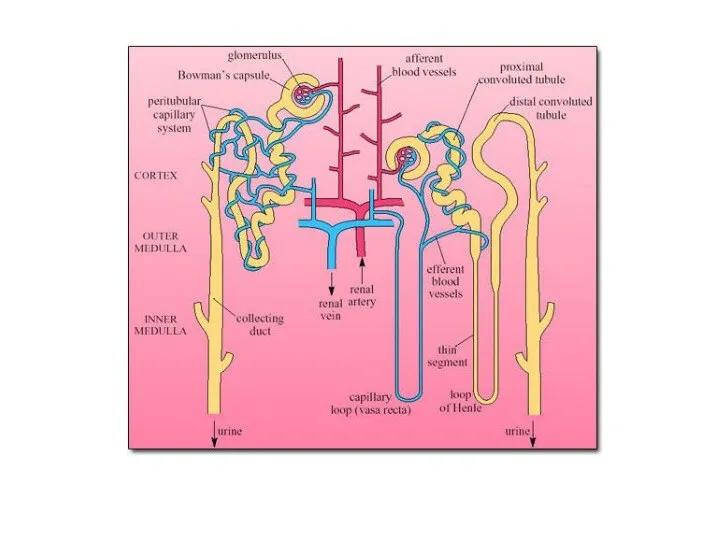
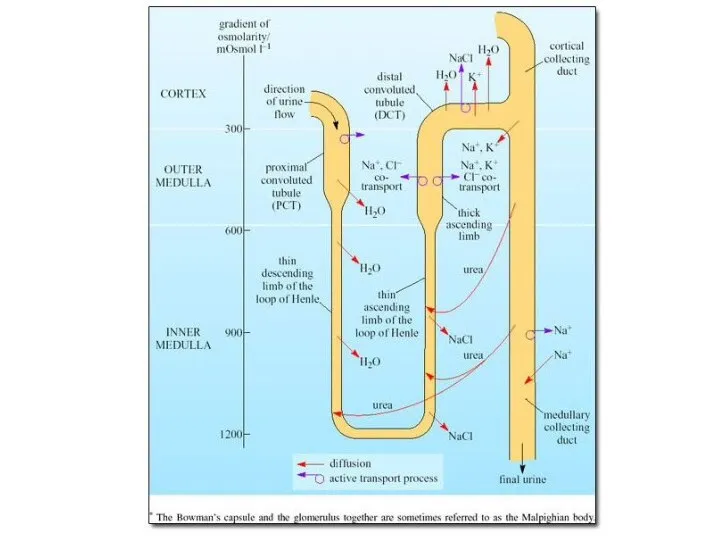
Модель теплообмена в носовых ходах (Schmidt-Nielsen, 1972). Окружающий
воздух не насыщен влагой,
Модель теплообмена в носовых ходах (Schmidt-Nielsen, 1972). Окружающий
воздух не насыщен влагой,
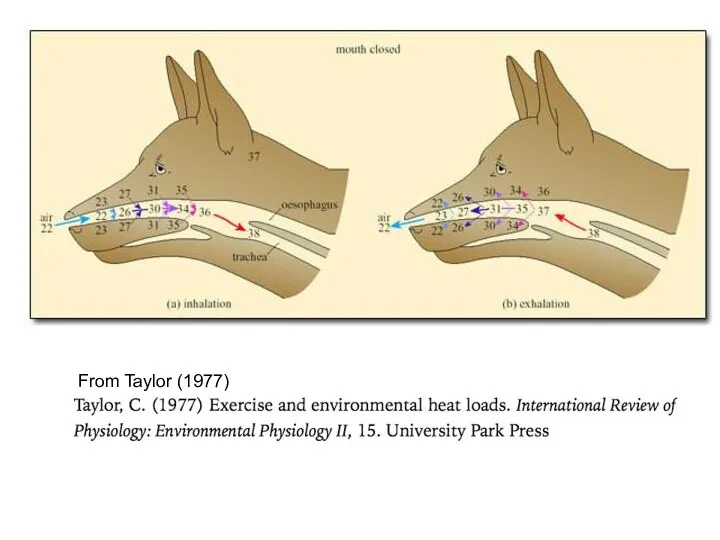
From Taylor (1977)
From Taylor (1977)
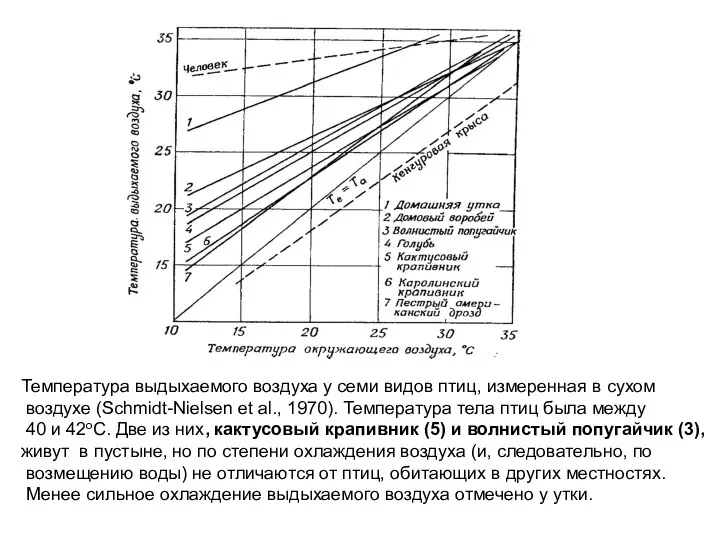
Температура выдыхаемого воздуха у семи видов птиц, измеренная в сухом
воздухе
Температура выдыхаемого воздуха у семи видов птиц, измеренная в сухом
воздухе

Еще о приспособлениях млекопитающих
в пустыне…
Еще о приспособлениях млекопитающих
в пустыне…

Рассчитанная величина испарения, необходимого для поддержания постоянной температуры тела в жаркой
Рассчитанная величина испарения, необходимого для поддержания постоянной температуры тела в жаркой
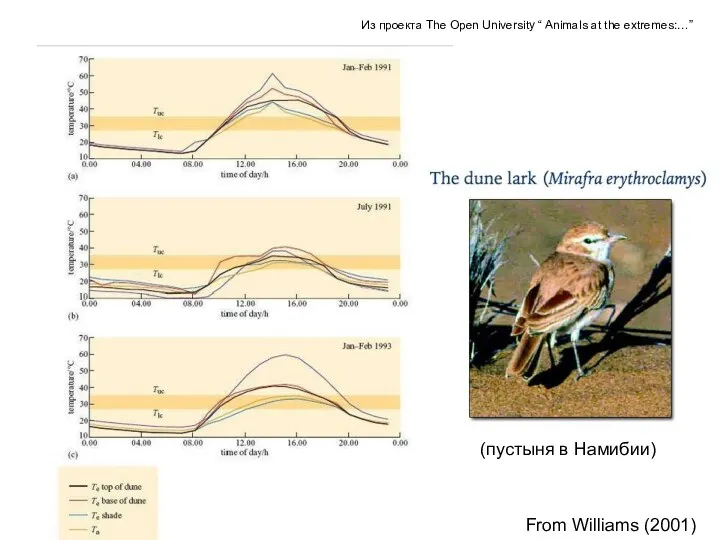
(пустыня в Намибии)
From Williams (2001)
Из проекта The Open University “ Animals
(пустыня в Намибии)
From Williams (2001)
Из проекта The Open University “ Animals
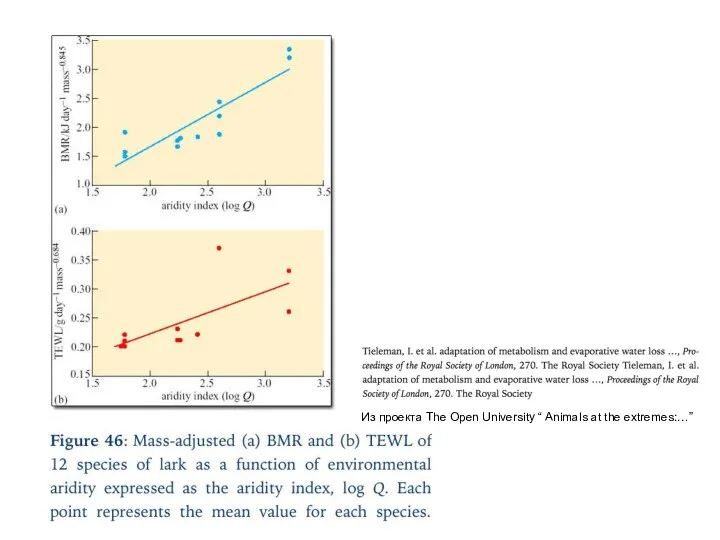
Из проекта The Open University “ Animals at the extremes:…”
Из проекта The Open University “ Animals at the extremes:…”
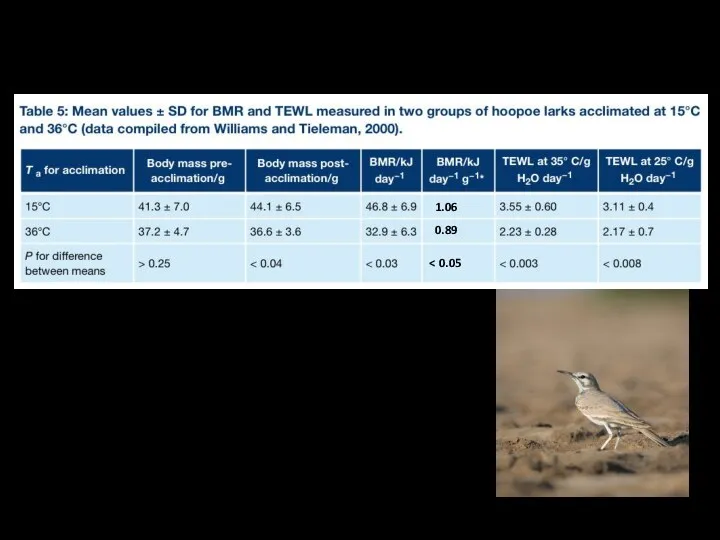
Большой удодовый жаворонок Alaemon alaudipes
обитает в пустынях на Аравийском полуострове, в
Большой удодовый жаворонок Alaemon alaudipes
обитает в пустынях на Аравийском полуострове, в
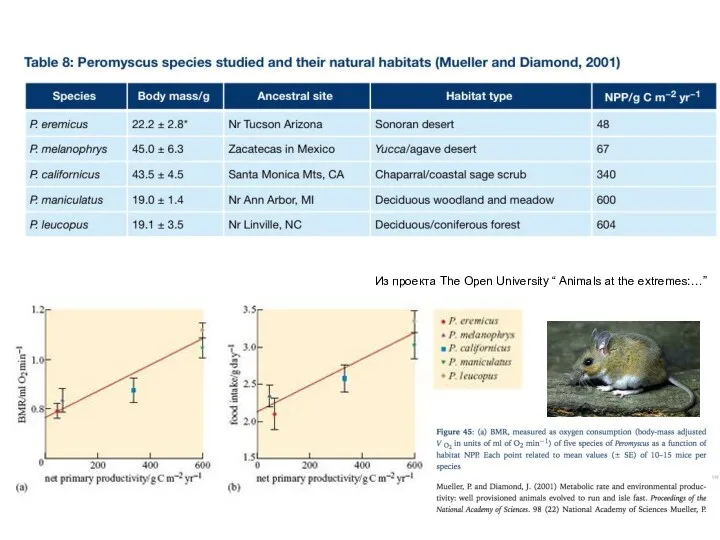
Из проекта The Open University “ Animals at the extremes:…”
Из проекта The Open University “ Animals at the extremes:…”
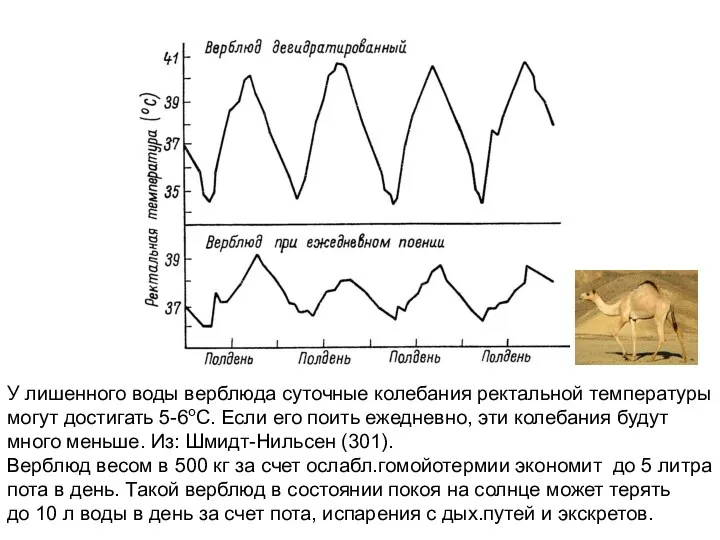
У лишенного воды верблюда суточные колебания ректальной температуры
могут достигать 5-6оС.
У лишенного воды верблюда суточные колебания ректальной температуры
могут достигать 5-6оС.
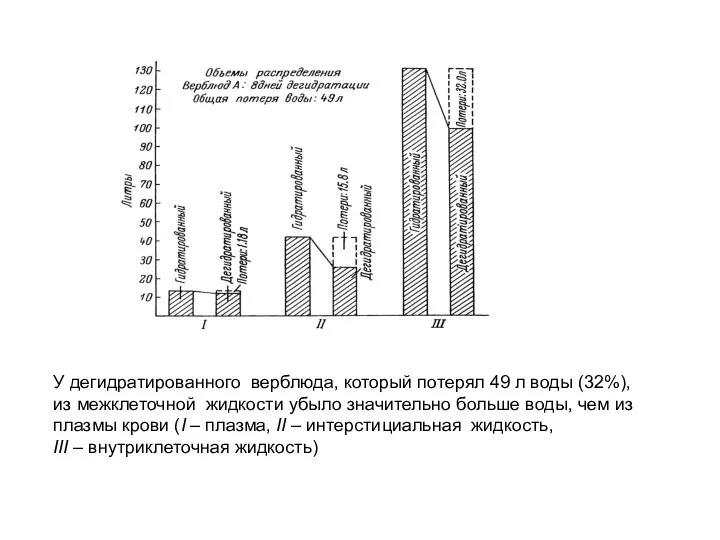
У дегидратированного верблюда, который потерял 49 л воды (32%),
из межклеточной
У дегидратированного верблюда, который потерял 49 л воды (32%),
из межклеточной

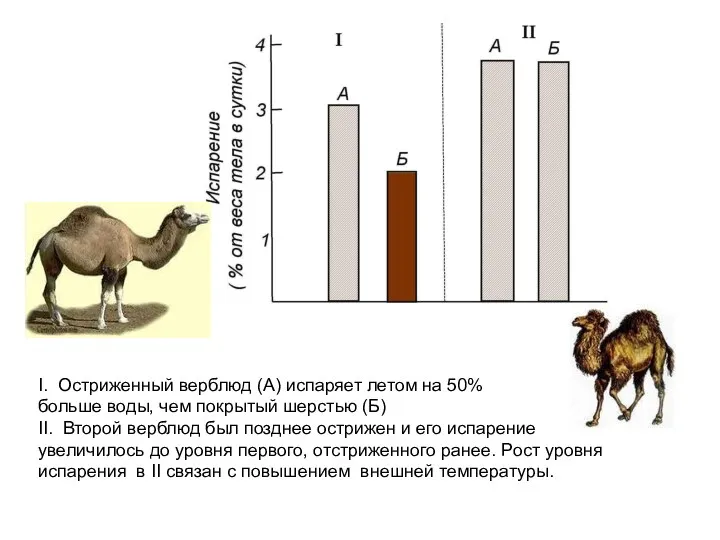
I. Остриженный верблюд (А) испаряет летом на 50%
больше воды, чем
I. Остриженный верблюд (А) испаряет летом на 50%
больше воды, чем



и в тропиках…
и в тропиках…
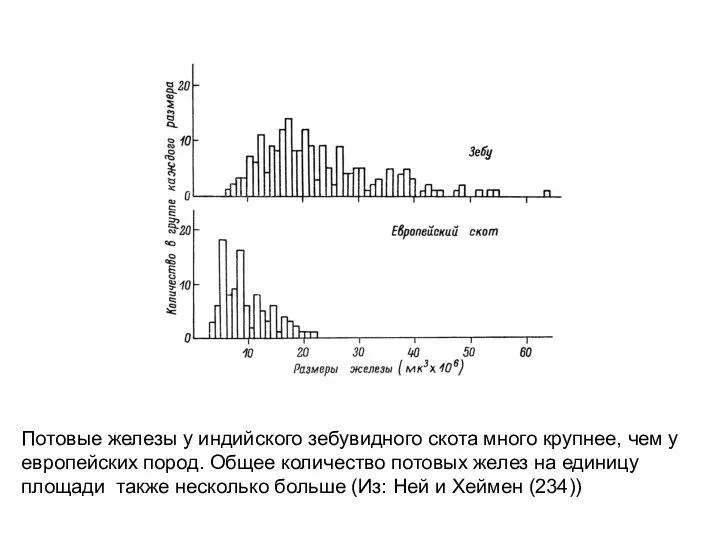
Потовые железы у индийского зебувидного скота много крупнее, чем у
европейских
Потовые железы у индийского зебувидного скота много крупнее, чем у
европейских
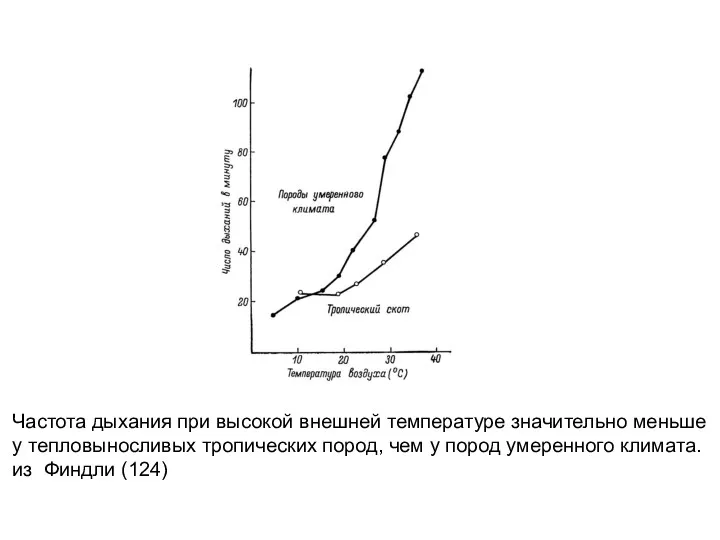
Частота дыхания при высокой внешней температуре значительно меньше
у тепловыносливых тропических пород,
Частота дыхания при высокой внешней температуре значительно меньше
у тепловыносливых тропических пород,

Морские позвоночные, дышащие воздухом
В отличие от рыб избегают многих осмотических проблем,
Морские позвоночные, дышащие воздухом
В отличие от рыб избегают многих осмотических проблем,

Пьют м. воду ?
> %C солей в жидкостях тела
и
Пьют м. воду ?
> %C солей в жидкостях тела
и

Выведение соли солевыми железами
Рептилии – 3 отряда (ящерицы, змеи, черепахи)
имеют
Выведение соли солевыми железами
Рептилии – 3 отряда (ящерицы, змеи, черепахи)
имеют

Кстати…
Слезы человека изоосмотичны по отношению
к плазме крови и не играют
Кстати…
Слезы человека изоосмотичны по отношению
к плазме крови и не играют
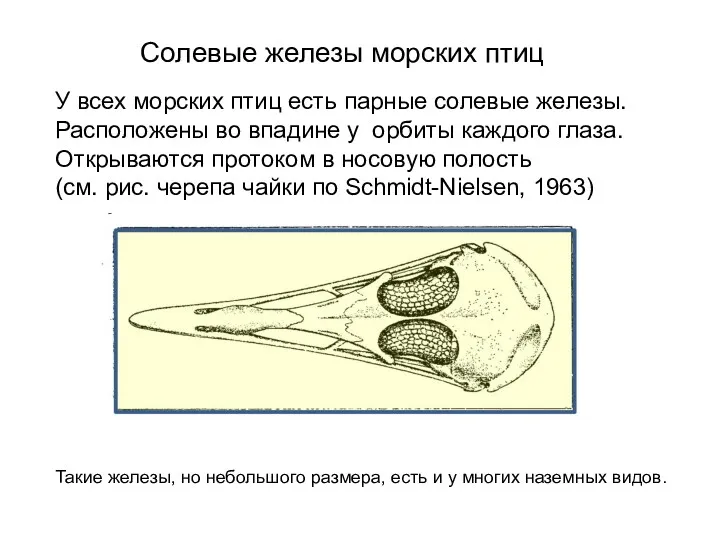
Солевые железы морских птиц
У всех морских птиц есть парные солевые железы.
Солевые железы морских птиц
У всех морских птиц есть парные солевые железы.

Как и у рептилий, солевые железы птиц работают в ответ на
Как и у рептилий, солевые железы птиц работают в ответ на
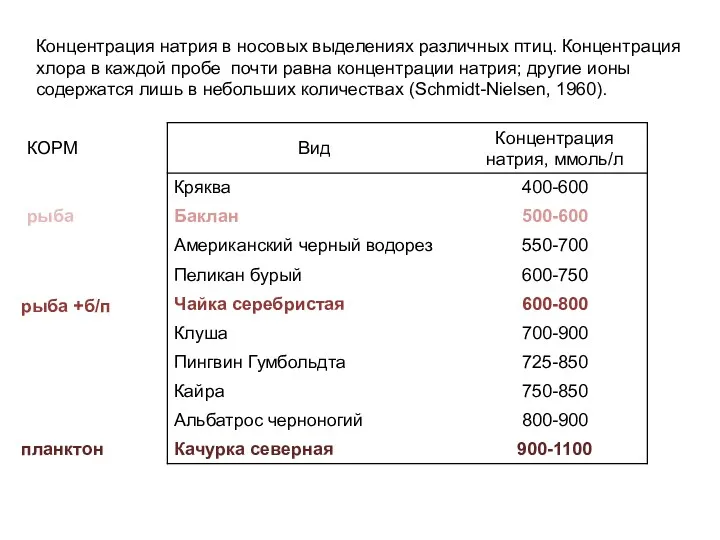
Концентрация натрия в носовых выделениях различных птиц. Концентрация
хлора в каждой пробе
Концентрация натрия в носовых выделениях различных птиц. Концентрация
хлора в каждой пробе

Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
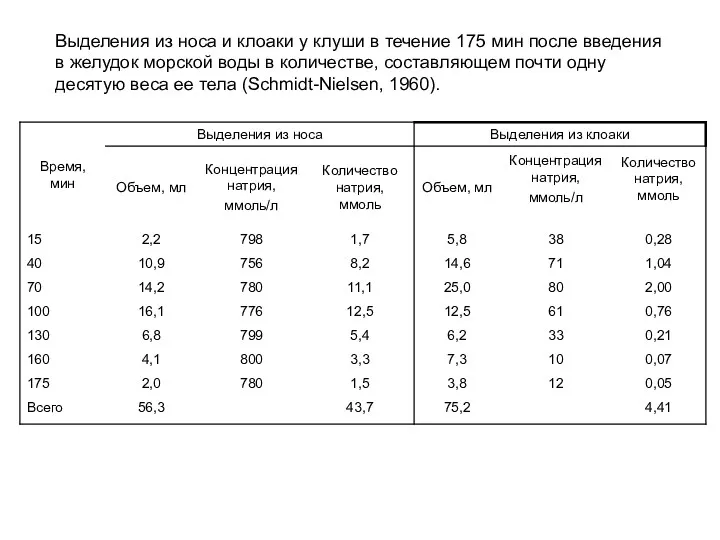
Выделения из носа и клоаки у клуши в течение 175 мин
Выделения из носа и клоаки у клуши в течение 175 мин

Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
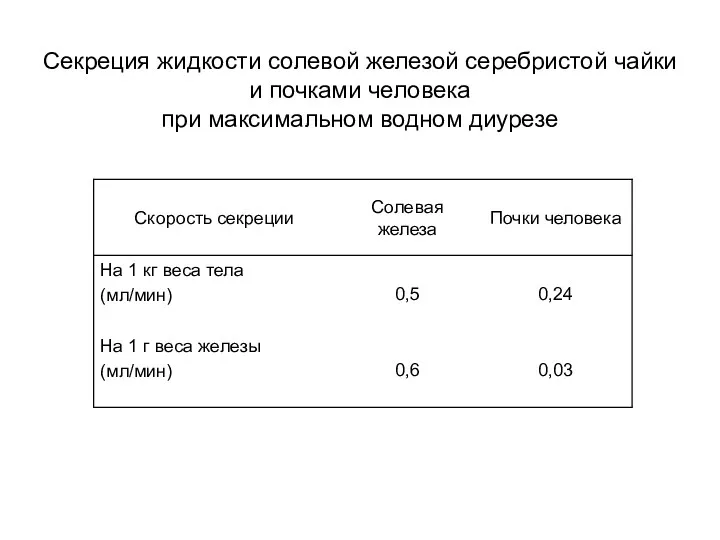
Секреция жидкости солевой железой серебристой чайки
и почками человека
при максимальном водном
Секреция жидкости солевой железой серебристой чайки
и почками человека
при максимальном водном

Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
Поразительная способность выделять соль:
В опыте клуша выпивала м.воду до
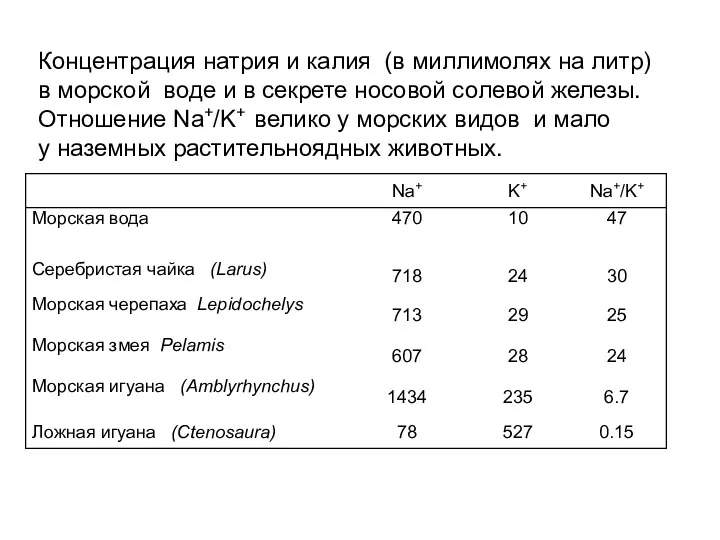
Концентрация натрия и калия (в миллимолях на литр)
в морской воде
Концентрация натрия и калия (в миллимолях на литр)
в морской воде
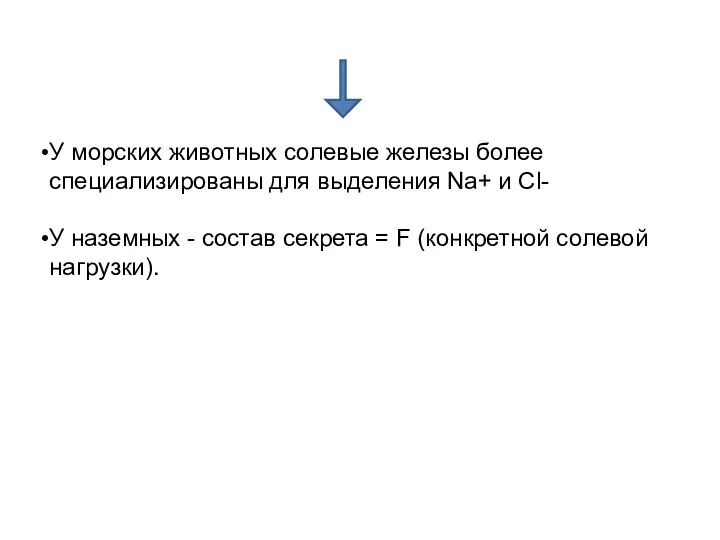
У морских животных солевые железы более специализированы для выделения Na+ и
У морских животных солевые железы более специализированы для выделения Na+ и

Концентрация ионов Cl- в экскретах некоторых видов птиц при
искусственной солевой нагрузке
Концентрация ионов Cl- в экскретах некоторых видов птиц при
искусственной солевой нагрузке

Морские млекопитающие
Сирены
Киты
Ластоногие
Пища разнообразна по %С.
Тюлени и киты едят рыбу, крупных б/п
Морские млекопитающие
Сирены
Киты
Ластоногие
Пища разнообразна по %С.
Тюлени и киты едят рыбу, крупных б/п

Они не нуждаются в солевых железах, так как их
почки способны
Они не нуждаются в солевых железах, так как их
почки способны

Влияние 1 л морской воды, введенной через рот, на водный баланс
Влияние 1 л морской воды, введенной через рот, на водный баланс
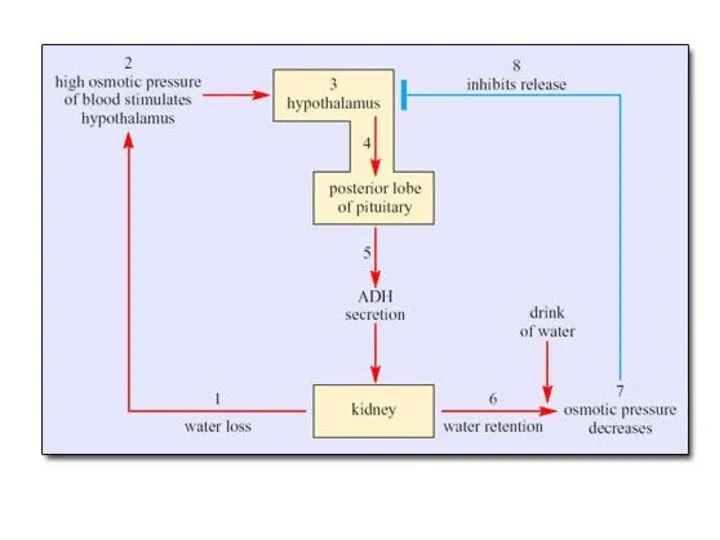
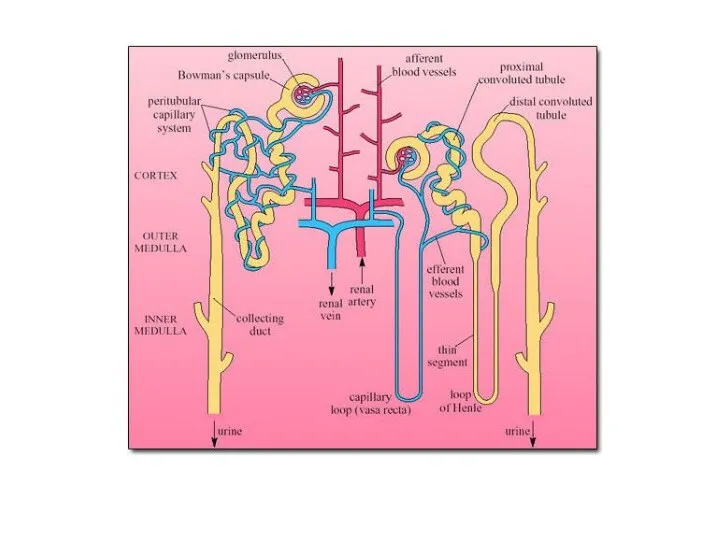

Важная особенность водного баланса морских
млекопитающих - высокая % С их
Важная особенность водного баланса морских
млекопитающих - высокая % С их

Состав и энергетическая ценность молока млекопитающих
Значения в скобках относятся к общему
Состав и энергетическая ценность молока млекопитающих
Значения в скобках относятся к общему
 Методическая разработка к уроку по теме Насекомые
Методическая разработка к уроку по теме Насекомые Современная эмбриология
Современная эмбриология Важность правильного питания
Важность правильного питания Мәдени өсімдіктер мен үй жануарларының шығу орталықтары
Мәдени өсімдіктер мен үй жануарларының шығу орталықтары Методическая разработка заочной экскурсии в мир леса. Хвойные породы. Часть 2
Методическая разработка заочной экскурсии в мир леса. Хвойные породы. Часть 2 Роль кальция в жизни человека
Роль кальция в жизни человека Бактериологический анализ
Бактериологический анализ Rare images from Wildlife Photography
Rare images from Wildlife Photography Породи кішок
Породи кішок Генетические и средовые детерминанты вызванных потенциалов
Генетические и средовые детерминанты вызванных потенциалов Вакуолярная система
Вакуолярная система Грибы. 6 класс
Грибы. 6 класс Однопроходные сумчатые
Однопроходные сумчатые Иммунитет
Иммунитет Законы Грегора Менделя
Законы Грегора Менделя Повторительно-обобщающий урок Биосферный уровень жизни. Тест. 9 класс
Повторительно-обобщающий урок Биосферный уровень жизни. Тест. 9 класс Эволюция. Теория Дарвина
Эволюция. Теория Дарвина Домашние кошки
Домашние кошки Первоцветы Сибири. Примула,
Первоцветы Сибири. Примула, Мембрани. ранспр
Мембрани. ранспр Отряд неполнозубые (edentata)
Отряд неполнозубые (edentata) Использование здоровье-сберегающих технологий в учебно-образовательном процессе
Использование здоровье-сберегающих технологий в учебно-образовательном процессе Скажи, дружок, что за цветок?
Скажи, дружок, что за цветок? Сухоцветы. Составление зимних букетов
Сухоцветы. Составление зимних букетов Особенности молекулярной эволюции
Особенности молекулярной эволюции Wild life in Britain
Wild life in Britain Класс насекомые
Класс насекомые Конспект и презентация урока биологии
Конспект и презентация урока биологии