- Воспроизводительная система рыб

Содержание
- 2. Дифференциация полов 2 Половая дифференциация у рыб - явление уникальное. Рыбы обладают очень пластичной системой репродукции,
- 3. 3.У основных объектов рыборазведения -- карповых, лососевых, представителей осетровых рыб -половых хромосом нет. У этих рыб
- 4. 4.У рыб первичные половые клетки способны развиваться как по женскому, так и по мужскому пути. Многим
- 5. 5.Вторичные половые признаки рыб Вторичные половые признаки рыб, которые особенно ярко проявляются в период нереста, имеют
- 6. 6.Определение пола по форме анального отверстия и форме плавников Вторичные половые признаки рыб: а-спинные плавники Labeo
- 7. Половое поведение рыб перед нерестом 7.Важным половым признаком может служить половое поведение рыб перед нерестом. Это
- 8. 8.Регулирование пола с помощью стероидных гармонов Управление половым детерминизмом при искусственном разведении рыб имеет большое практическое
- 9. 9.Морфология половой системы рыб Развитие мочеполовой системы в эволюции рыб привело к обособлению половых протоков от
- 10. 11.Схема семяизвержения и овуляции у костистых рыб а -самка; 6- самец; 1-почка; 2-гонады; 3 - мочеточник;
- 11. 10.Морфология половой системы рыб А-минога; б- акула; в- осетр; г - лосось; д - карп; 1-
- 12. 11.К моменту полового созревания рыб масса их гонад (семенников и ястыков) возрастает в результате деления половых
- 13. 12.Изменение массы гонад в течении года (г/кг/) массы тела
- 14. 13.Масса зрелых гонад,%
- 15. 14.Плодовитость рыб Плодовитость рыб связана с величиной икринок. Так, у акулы диаметр яйца достигает нескольких сантиметров,
- 16. 15.Объем спермы (мл)у разных видов рыб Объем спермы (мл) у разных видов рыб Спермин, полученные непосредственно
- 17. 16.Высока плодовитость и у фитофильных рыб, откладывающих икру на растительность. Например, достигает 1,5 млн икринок, а
- 18. 17.Оплодотворение У рыб слияние половых клеток происходит, как правило, в воде, т. е. является наружным. Однако
- 19. 18Морфология сперматозоидов Рис. . Проникновение спермия в яйцеклетку при оплодотворении (I)- спермии разных видов рыб (II):
- 20. 19.Дальнейший онтогенез рыб имеет несколько вариантов развития. Так, у речного угря, камбалы, луны-рыбы и некоторых других
- 21. 20.Влияние температуры на продолжительность инкубации Развитие икры требует времени и определенного количества теплоты. При повышении температуры
- 22. 21.Периоды онтогенеза костистой рыбы Рис. Периоды раннего онтогенеза костистой рыбы: 1- икринка; 2- предличинка; 3 -
- 23. 22.Внутриутробное развитие рыб Внутриутробное развитие при внутреннем оплодотворении у рыб имеет различную степень совершенства. Например, у
- 24. 23.Самец Apogon semilineatus с икрой в ротовой полости; вверху показана "плацентарная связь" икринки со слизистой оболочкой
- 25. 24Разновидности плодоношения Разновидности плодоношения: 1, 2-развитие икры в желудке барбуса; 3, 4- развитие икры на брюхе
- 27. Скачать презентацию

Дифференциация полов 2
Половая дифференциация у рыб - явление уникальное. Рыбы обладают
Дифференциация полов 2
Половая дифференциация у рыб - явление уникальное. Рыбы обладают

3.У основных объектов рыборазведения -- карповых, лососевых, представителей осетровых рыб -половых
3.У основных объектов рыборазведения -- карповых, лососевых, представителей осетровых рыб -половых

4.У рыб первичные половые клетки способны развиваться как по женскому, так
4.У рыб первичные половые клетки способны развиваться как по женскому, так

5.Вторичные половые признаки рыб
Вторичные половые признаки рыб, которые особенно ярко проявляются
5.Вторичные половые признаки рыб
Вторичные половые признаки рыб, которые особенно ярко проявляются
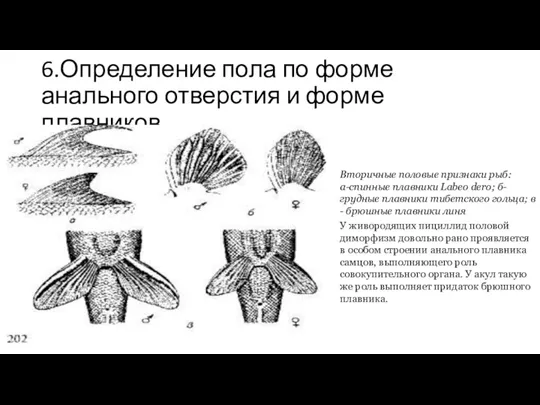
6.Определение пола по форме анального отверстия и форме плавников
Вторичные половые признаки
6.Определение пола по форме анального отверстия и форме плавников
Вторичные половые признаки

Половое поведение рыб перед нерестом
7.Важным половым признаком может служить половое поведение
Половое поведение рыб перед нерестом
7.Важным половым признаком может служить половое поведение

8.Регулирование пола с помощью стероидных гармонов
Управление половым детерминизмом при искусственном разведении
8.Регулирование пола с помощью стероидных гармонов
Управление половым детерминизмом при искусственном разведении

9.Морфология половой системы рыб
Развитие мочеполовой системы в эволюции рыб привело к
9.Морфология половой системы рыб
Развитие мочеполовой системы в эволюции рыб привело к
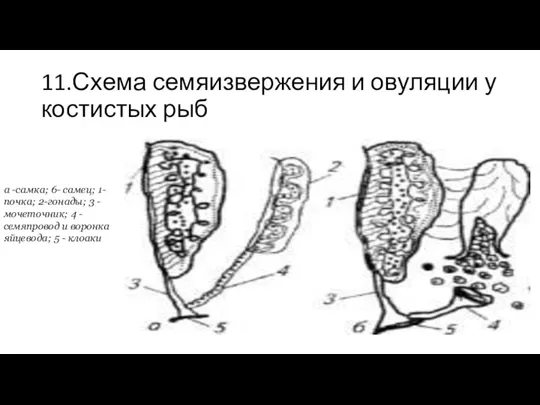
11.Схема семяизвержения и овуляции у костистых рыб
а -самка; 6- самец; 1-почка;
11.Схема семяизвержения и овуляции у костистых рыб
а -самка; 6- самец; 1-почка;
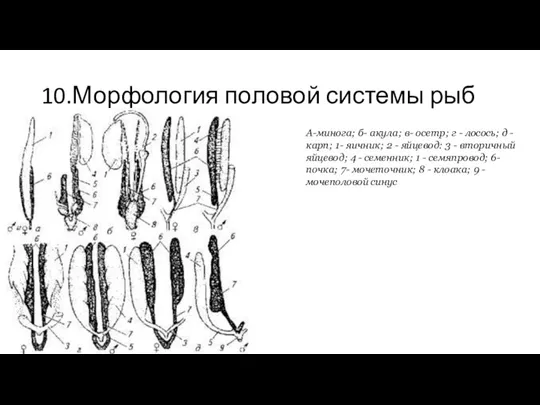
10.Морфология половой системы рыб
А-минога; б- акула; в- осетр; г - лосось;
10.Морфология половой системы рыб
А-минога; б- акула; в- осетр; г - лосось;

11.К моменту полового созревания рыб масса их гонад (семенников и ястыков)
11.К моменту полового созревания рыб масса их гонад (семенников и ястыков)
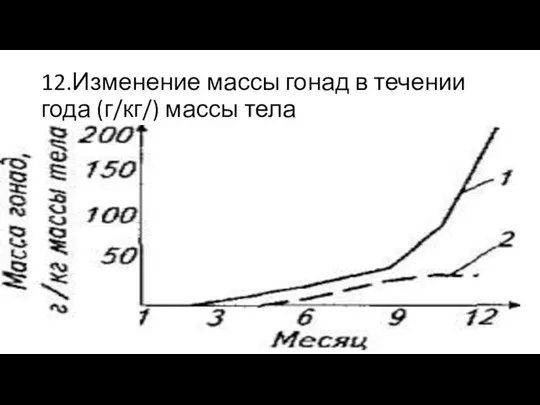
12.Изменение массы гонад в течении года (г/кг/) массы тела
12.Изменение массы гонад в течении года (г/кг/) массы тела
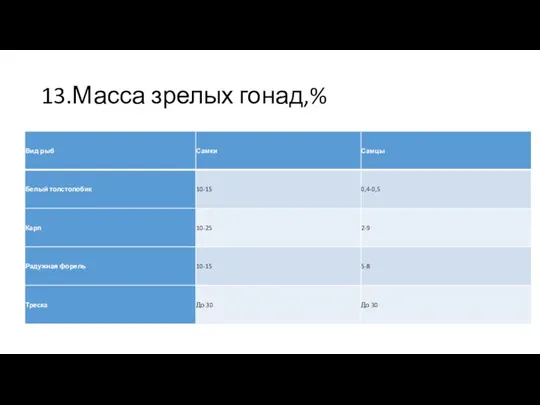
13.Масса зрелых гонад,%
13.Масса зрелых гонад,%
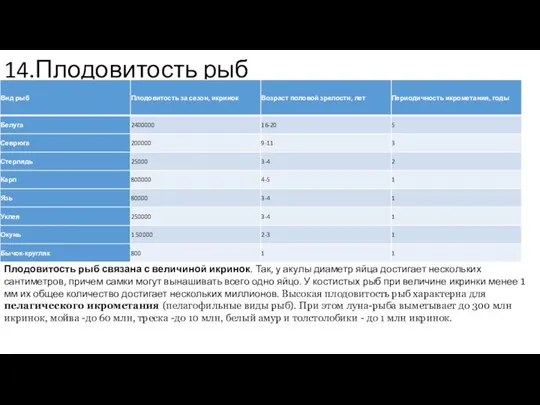
14.Плодовитость рыб
Плодовитость рыб связана с величиной икринок. Так, у акулы диаметр
14.Плодовитость рыб
Плодовитость рыб связана с величиной икринок. Так, у акулы диаметр

15.Объем спермы (мл)у разных видов рыб
Объем спермы (мл) у разных видов
15.Объем спермы (мл)у разных видов рыб
Объем спермы (мл) у разных видов

16.Высока плодовитость и у фитофильных рыб, откладывающих икру на растительность. Например,
16.Высока плодовитость и у фитофильных рыб, откладывающих икру на растительность. Например,

17.Оплодотворение
У рыб слияние половых клеток происходит, как правило, в воде, т.
17.Оплодотворение
У рыб слияние половых клеток происходит, как правило, в воде, т.

18Морфология сперматозоидов
Рис. . Проникновение спермия в яйцеклетку при оплодотворении (I)- спермии
18Морфология сперматозоидов
Рис. . Проникновение спермия в яйцеклетку при оплодотворении (I)- спермии

19.Дальнейший онтогенез рыб имеет несколько вариантов развития. Так, у речного угря,
19.Дальнейший онтогенез рыб имеет несколько вариантов развития. Так, у речного угря,

20.Влияние температуры на продолжительность инкубации
Развитие икры требует времени и определенного количества
20.Влияние температуры на продолжительность инкубации
Развитие икры требует времени и определенного количества

21.Периоды онтогенеза костистой рыбы
Рис. Периоды раннего онтогенеза костистой рыбы:
1- икринка; 2-
21.Периоды онтогенеза костистой рыбы
Рис. Периоды раннего онтогенеза костистой рыбы: 1- икринка; 2-

22.Внутриутробное развитие рыб
Внутриутробное развитие при внутреннем оплодотворении у рыб имеет различную
22.Внутриутробное развитие рыб
Внутриутробное развитие при внутреннем оплодотворении у рыб имеет различную

23.Самец Apogon semilineatus с икрой в ротовой полости; вверху показана "плацентарная связь" икринки со
23.Самец Apogon semilineatus с икрой в ротовой полости; вверху показана "плацентарная связь" икринки со

24Разновидности плодоношения
Разновидности плодоношения:
1, 2-развитие икры в желудке барбуса; 3, 4- развитие
24Разновидности плодоношения
Разновидности плодоношения: 1, 2-развитие икры в желудке барбуса; 3, 4- развитие
 f611e845-2141-4c61-a9b2-34e2049b0658
f611e845-2141-4c61-a9b2-34e2049b0658 Морские свинки
Морские свинки Особенности пищеварения у собак и котов. Питательные и биологически активные вещества
Особенности пищеварения у собак и котов. Питательные и биологически активные вещества Лекция 1. Растениеводство
Лекция 1. Растениеводство Этимология названий лекарственных растений. Мифы и легенды
Этимология названий лекарственных растений. Мифы и легенды Почки растений
Почки растений Турбота про потомство
Турбота про потомство Путешествие в страну Легумия. Систематика покрытосеменных. 6 класс
Путешествие в страну Легумия. Систематика покрытосеменных. 6 класс Нейропрорывы-2017/2018
Нейропрорывы-2017/2018 Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся
Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся Вітаміни. Їх роль у життєдіяльності людини
Вітаміни. Їх роль у життєдіяльності людини Размножение - свойство живых организмов
Размножение - свойство живых организмов Підряд Мавпи
Підряд Мавпи Энергетический обмен. Дыхание
Энергетический обмен. Дыхание Цікаві факти про риб
Цікаві факти про риб Талдағыштар физиологиясы. Талдағыштардың жалпы қасиеттері. Көру талдағыштары
Талдағыштар физиологиясы. Талдағыштардың жалпы қасиеттері. Көру талдағыштары Структура и функции клетки
Структура и функции клетки Размножение и развитие человека
Размножение и развитие человека Покрытосеменные, или Цветковые
Покрытосеменные, или Цветковые Самые страшные животные мира
Самые страшные животные мира Рост и развитие растений
Рост и развитие растений Освоение суши растениями
Освоение суши растениями Хромосомная теория наследственностию. Генетика пола
Хромосомная теория наследственностию. Генетика пола Юрский период (195 -137 млн.)
Юрский период (195 -137 млн.) Презентация Великаны и карлики в животном мире
Презентация Великаны и карлики в животном мире Насекомые. Отряд равнокрылые
Насекомые. Отряд равнокрылые Биотические факторы
Биотические факторы Становление наук о человеке. (8 класс)
Становление наук о человеке. (8 класс)