- Введение в физиологию. Возбуждение. сокращение. Физиология клетки

Содержание
- 2. Тема: Введение в физиологию. Физиология клетки План Основные понятия. Принципы функционирования организма Физиологические свойства тканей Структура
- 3. завершающие дисциплины доклинического образования
- 4. 1. Основные понятия. Принципы функционирования организма Физиология — наука, занимающаяся изучением функционирования живых организмов и составляющих
- 5. Саморегуляция – основа жизнедеятельности живых организмов. Базируется на принципе прямой и обратной связи (Н. Винер) Регулирующее
- 6. 2. Физиологические свойства тканей Раздражимость – способность активно реагировать на раздражители: неспецифические изменения : усиление обмена
- 7. 3. Структура клеточной мембраны. Мембранный транспорт
- 8. Функции клеточной мембраны барьерная транспортная механическая энергетическая рецепторная ферментативная генерация и проведение биопотенциалов маркировка клетки
- 9. Мембранный транспорт
- 12. Активный транспорт – движение против электрохимического градиента затрата энергии (гидролиз АТФ) первично активный вторично активный с
- 13. Транспорт с переносчиками Переносчики (транспортеры) – белки, которые связывают ионы или другие молекулы и затем изменяя
- 14. Везикулярный транспорт Эндоцитоз - перемещение макромолекул путем активной инвагинации мембраны клетки Фагоцитоз - поглощение частиц (бактерий,
- 15. 4. Возбудимость и возбуждение. Мембранный потенциал Возбудимость (свойство) - способность высокоспециализированных тканей реагировать на раздражение комплексом,
- 16. Трансмембранная разность потенциалов (мембранный потенциал) есть у всех клеток: для клетки в покое – это мембранный
- 17. Основы мембранного потенциала покоя Различия концентраций ионов [С] снаружи и внутри клетки [K+in] > [K+out], [Na+in]
- 18. ИОННОЕ РАВНОВЕСИЕ И МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ Ионы перемещаются через мембрану благодаря электрохимическому градиенту по обе стороны
- 19. Ионные насосы (Na/K – АТФ-аза) поддерживают неравновесное распределение Na+ и К+ расщепление 1 АТФ - перенос
- 20. Мембранный потенциал покоя - -70 mV для большинства клеток; -90 mV для нейронов; K+ - основной
- 21. Потенциал действия (ПД) – быстрые колебания трансмембранной разности потенциалов, обусловленные изменением ионной проницаемости мембраны. Фазы ПД
- 22. спайк деполяризация Овершут полная деполяризация реполяризация мембранный потенциал покоя следовая деполяризация следовая гиперполяризация Порог
- 23. Развитие ПД возможно в том случае, если раздражитель достиг пороговой силы (порог раздражения), т.е. в результате
- 24. Потенциал действия является своеобразным триггером, запускающим их специфическую функциональную активность клетки: проведение нервного импульса, сокращение мышцы,
- 25. Фазовые изменения возбудимости во время развития потенциала действия Во время ПД возбудимость клеточной мембраны (способность реагировать
- 26. 5. Механизм мышечного сокращения поперечно-полосатые мышцы (скелетная мускулатура ) гладкие мышцы (стенки полых внутренних органов, сосудов)
- 27. Структура нервно-мышечного соединения концевая пластинка двигательного нерва на мышце концевая мышечная пластинка (постсинаптическая мембрана) Стимуляция нерва
- 28. Последовательность событий в нервно-мышечном синапсе деполяризация двигательной терминали аксона - вход Ca2+ в терминаль аксона выделение
- 29. КЛИНИЧЕСКИЕ АСПЕКТЫ Блокада нервно-мышечной передачи: 1) блокада возбуждения пресинаптической мембране местные анестетики: напр., лидокаин; 2) нарушение
- 30. Сократительные белки мышечных клеток – филаменты тонкие актин: сокращение небулин: длина актина – Са++-кальмодулин; тропонин, тропомиозин:
- 31. Механизм мышечного сокращения: теория скользящих филаментов Хаксли процессы в саркомере скольжение филаментов друг относительно друга толстые
- 32. Электромеханическое сопряжение возбуждение покой Внекл. жидкость СР – депо Са++ Внеклеточная жидкость
- 33. Итак, в результате электромеханического сопряжения ПД, распространяясь вглубь волокна, запускает цикл поперечных мостиков: распространение ПД внутрь
- 34. Цикл поперечных мостиков – АТФ-зависимый процесс 1 цикл – 1 молекула АТФ для диссоциации миозина и
- 35. КЛИНИЧЕСКИЕ КОРРЕЛЯТЫ Фармакотерпия блокаторы Ca2+ каналов L-типа антигипертензивная терапия замедление проводимости в сердце и снижение сократимости
- 37. Скачать презентацию

Тема: Введение в физиологию. Физиология клетки
План
Основные понятия. Принципы функционирования организма
Физиологические свойства
Тема: Введение в физиологию. Физиология клетки
План
Основные понятия. Принципы функционирования организма
Физиологические свойства
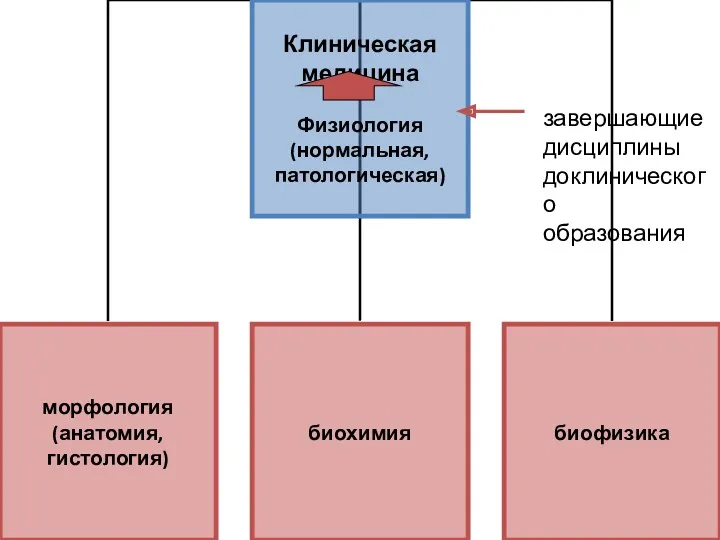
завершающие
дисциплины
доклинического
образования
завершающие
дисциплины
доклинического
образования

1. Основные понятия. Принципы функционирования организма
Физиология — наука, занимающаяся изучением функционирования живых
Физиология — наука, занимающаяся изучением функционирования живых
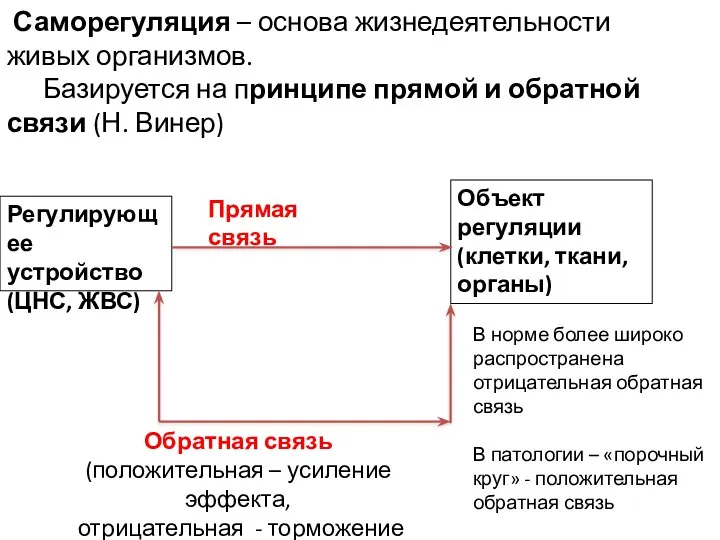
Саморегуляция – основа жизнедеятельности живых организмов.
Базируется на принципе прямой
Саморегуляция – основа жизнедеятельности живых организмов. Базируется на принципе прямой

2. Физиологические свойства тканей
Раздражимость – способность активно реагировать на раздражители:
неспецифические изменения
2. Физиологические свойства тканей
Раздражимость – способность активно реагировать на раздражители:
неспецифические изменения
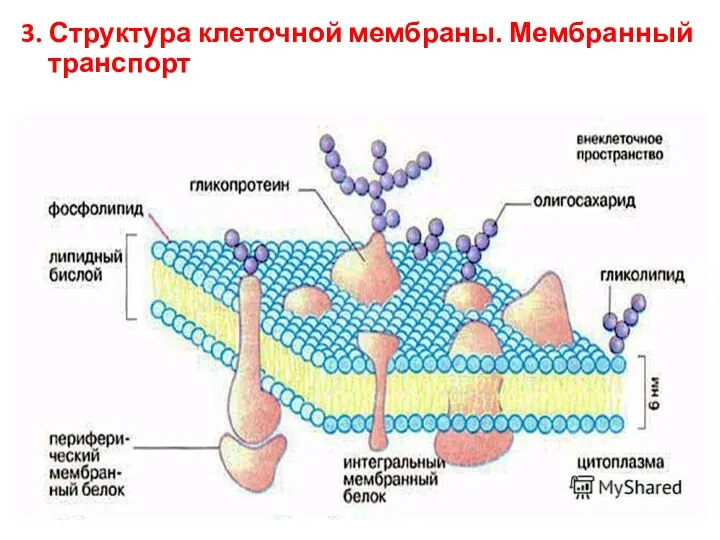
3. Структура клеточной мембраны. Мембранный транспорт
3. Структура клеточной мембраны. Мембранный транспорт

Функции клеточной мембраны
барьерная
транспортная
механическая
энергетическая
рецепторная
ферментативная
генерация и проведение биопотенциалов
маркировка клетки
Функции клеточной мембраны
барьерная
транспортная
механическая
энергетическая
рецепторная
ферментативная
генерация и проведение биопотенциалов
маркировка клетки
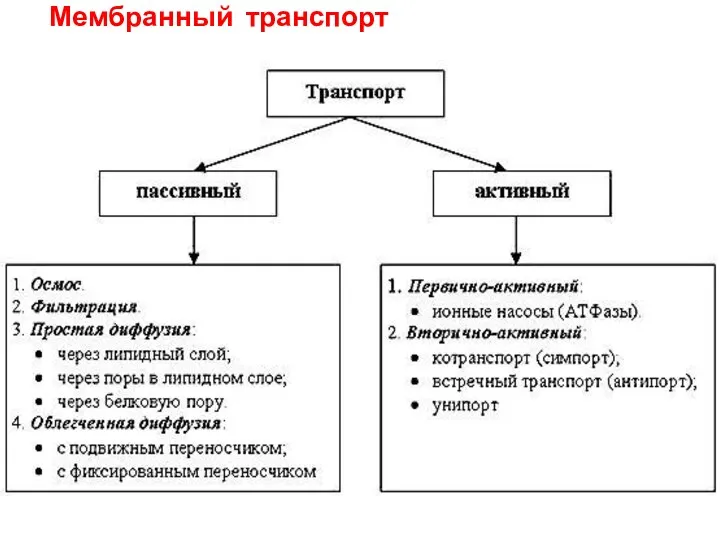
Мембранный транспорт
Мембранный транспорт
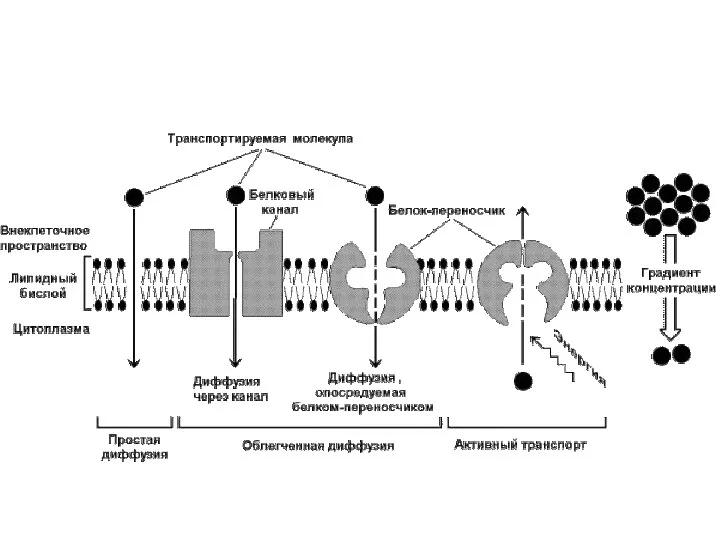
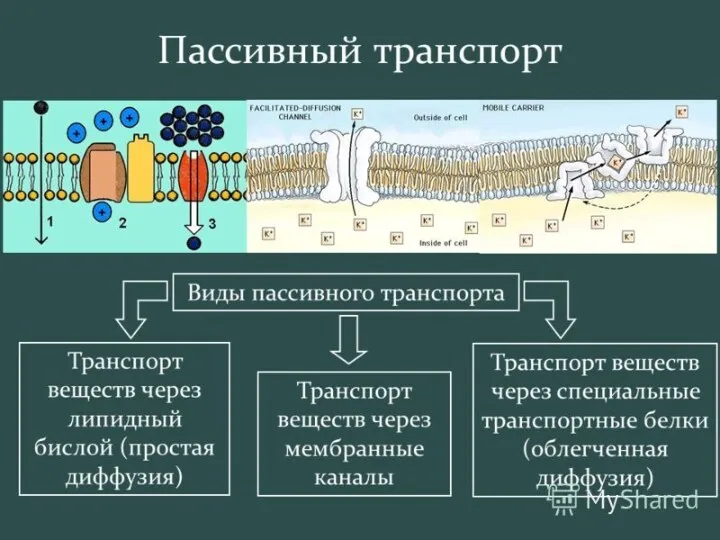

Активный транспорт –
движение против электрохимического градиента
затрата энергии (гидролиз АТФ)
первично активный
вторично
Активный транспорт –
движение против электрохимического градиента
затрата энергии (гидролиз АТФ)
первично активный
вторично
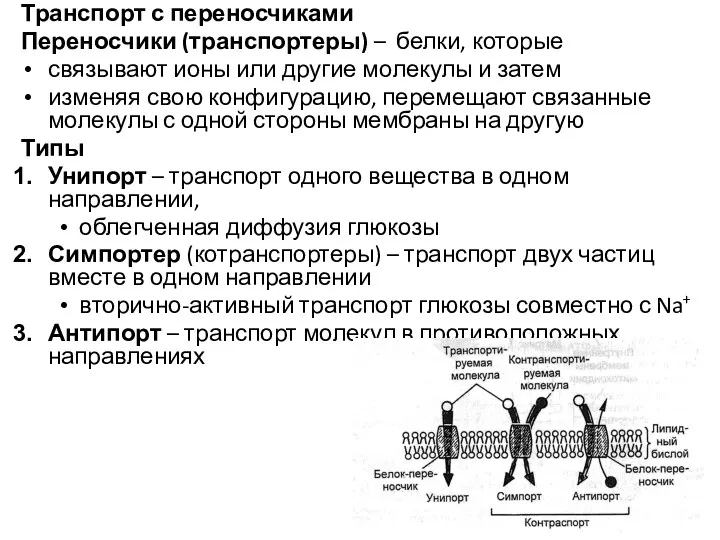
Транспорт с переносчиками
Переносчики (транспортеры) – белки, которые
связывают ионы или другие молекулы
Транспорт с переносчиками
Переносчики (транспортеры) – белки, которые
связывают ионы или другие молекулы
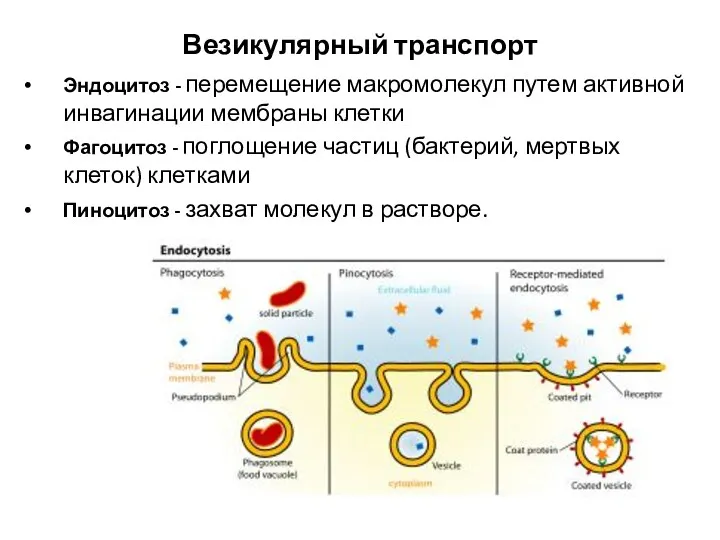
Везикулярный транспорт
Эндоцитоз - перемещение макромолекул путем активной инвагинации мембраны клетки
Фагоцитоз -
Везикулярный транспорт
Эндоцитоз - перемещение макромолекул путем активной инвагинации мембраны клетки
Фагоцитоз -

4. Возбудимость и возбуждение. Мембранный потенциал
Возбудимость (свойство) - способность высокоспециализированных тканей
4. Возбудимость и возбуждение. Мембранный потенциал
Возбудимость (свойство) - способность высокоспециализированных тканей
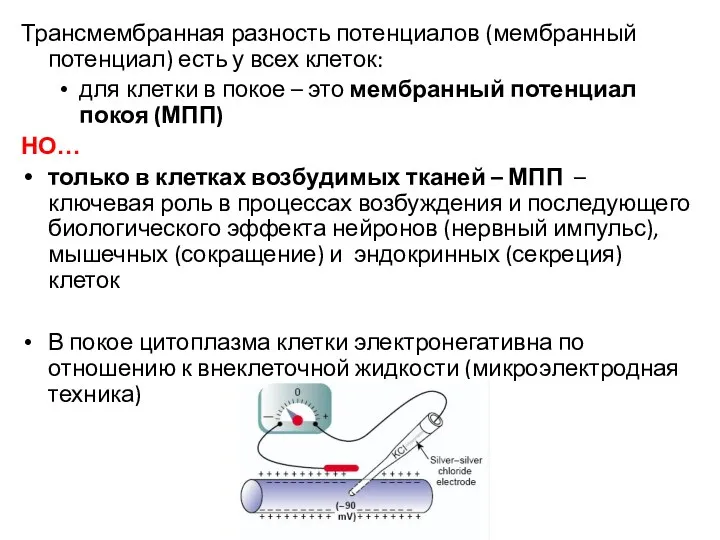
Трансмембранная разность потенциалов (мембранный потенциал) есть у всех клеток:
для клетки в
Трансмембранная разность потенциалов (мембранный потенциал) есть у всех клеток:
для клетки в
![Основы мембранного потенциала покоя Различия концентраций ионов [С] снаружи и](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/373830/slide-16.jpg)
Основы мембранного потенциала покоя
Различия концентраций ионов [С] снаружи и внутри клетки
Основы мембранного потенциала покоя
Различия концентраций ионов [С] снаружи и внутри клетки

ИОННОЕ РАВНОВЕСИЕ И МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
Ионы перемещаются через мембрану благодаря электрохимическому
ИОННОЕ РАВНОВЕСИЕ И МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
Ионы перемещаются через мембрану благодаря электрохимическому
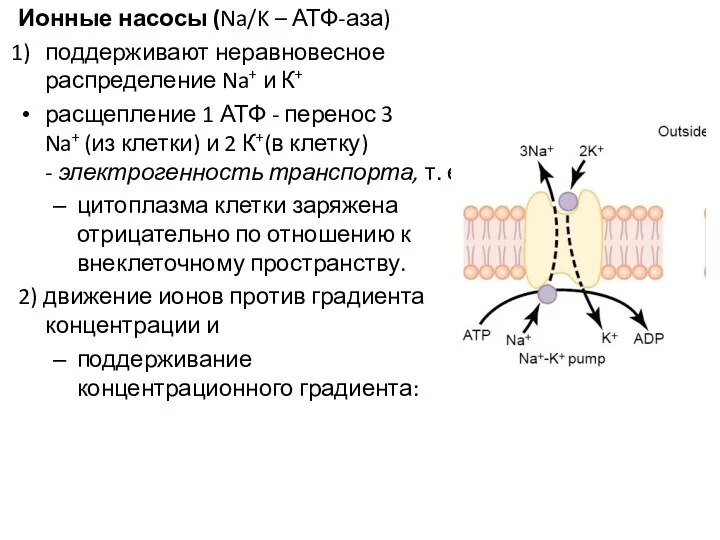
Ионные насосы (Na/K – АТФ-аза)
поддерживают неравновесное распределение Na+ и К+
расщепление 1 АТФ
Ионные насосы (Na/K – АТФ-аза)
поддерживают неравновесное распределение Na+ и К+
расщепление 1 АТФ

Мембранный потенциал покоя -
-70 mV для большинства клеток;
-90
Мембранный потенциал покоя -
-70 mV для большинства клеток;
-90
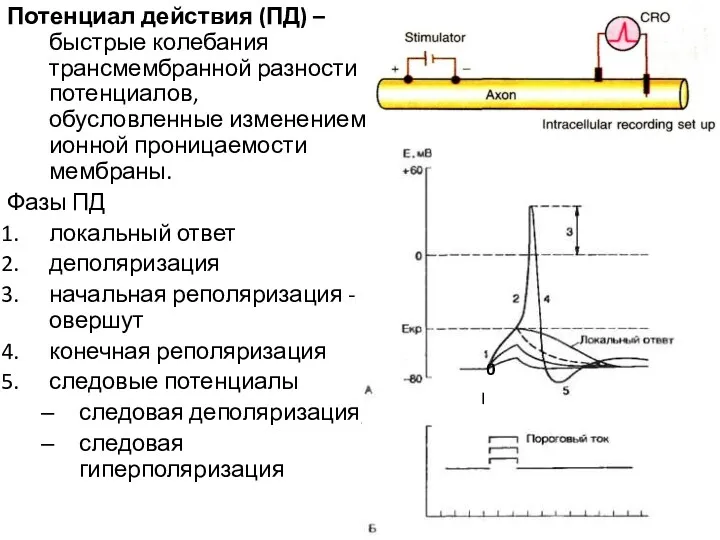
Потенциал действия (ПД) – быстрые колебания трансмембранной разности потенциалов, обусловленные изменением
Потенциал действия (ПД) – быстрые колебания трансмембранной разности потенциалов, обусловленные изменением
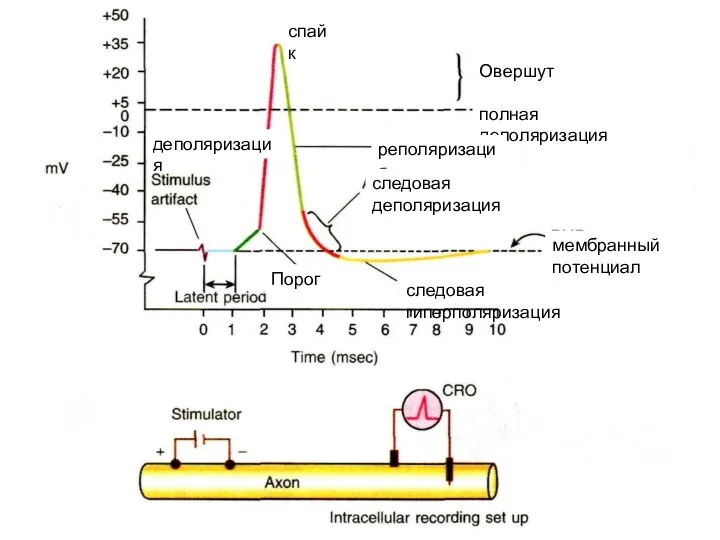
спайк
деполяризация
Овершут
полная деполяризация
реполяризация
мембранный
потенциал покоя
следовая деполяризация
следовая гиперполяризация
Порог
спайк
деполяризация
Овершут
полная деполяризация
реполяризация
мембранный
потенциал покоя
следовая деполяризация
следовая гиперполяризация
Порог
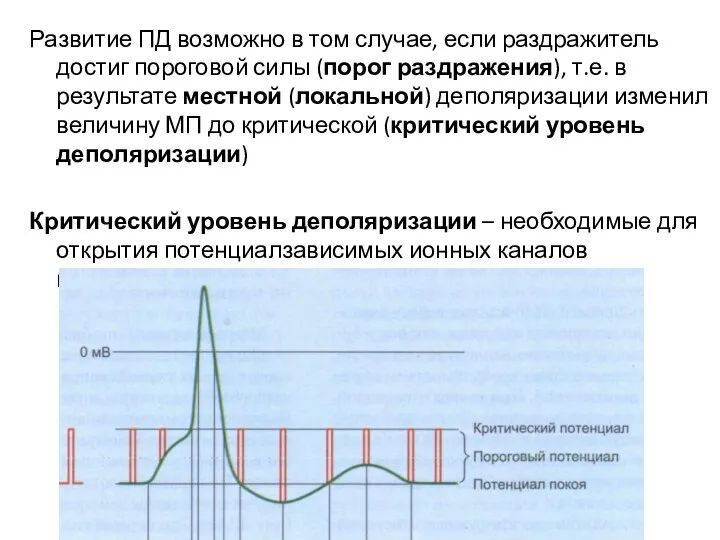
Развитие ПД возможно в том случае, если раздражитель достиг пороговой силы
Развитие ПД возможно в том случае, если раздражитель достиг пороговой силы

Потенциал действия является своеобразным триггером, запускающим их специфическую функциональную активность клетки:
проведение
проведение

Фазовые изменения возбудимости во время развития потенциала действия
Во время ПД возбудимость
Фазовые изменения возбудимости во время развития потенциала действия
Во время ПД возбудимость

5. Механизм мышечного сокращения
поперечно-полосатые мышцы (скелетная мускулатура )
гладкие мышцы (стенки полых
5. Механизм мышечного сокращения
поперечно-полосатые мышцы (скелетная мускулатура )
гладкие мышцы (стенки полых
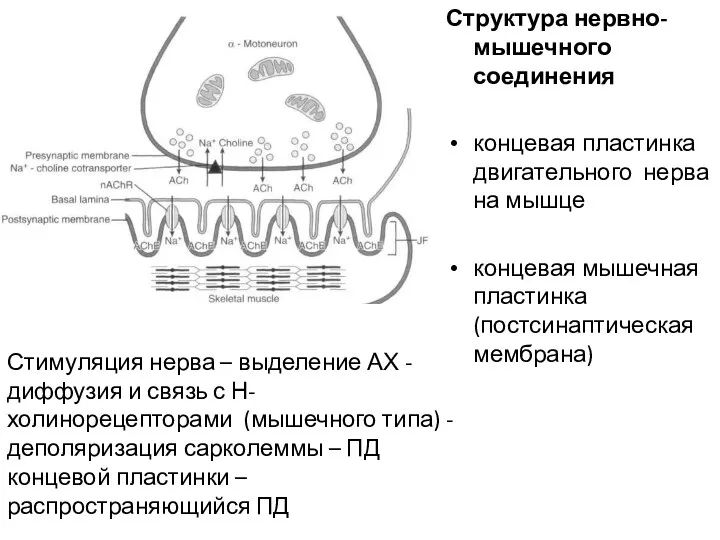
Структура нервно-мышечного соединения
концевая пластинка двигательного нерва на мышце
концевая мышечная пластинка (постсинаптическая
Структура нервно-мышечного соединения
концевая пластинка двигательного нерва на мышце
концевая мышечная пластинка (постсинаптическая

Последовательность событий в нервно-мышечном синапсе
деполяризация двигательной терминали аксона - вход Ca2+
Последовательность событий в нервно-мышечном синапсе
деполяризация двигательной терминали аксона - вход Ca2+

КЛИНИЧЕСКИЕ АСПЕКТЫ
Блокада нервно-мышечной передачи:
1) блокада возбуждения пресинаптической мембране
местные анестетики: напр., лидокаин;
2) нарушение синтеза
КЛИНИЧЕСКИЕ АСПЕКТЫ
Блокада нервно-мышечной передачи:
1) блокада возбуждения пресинаптической мембране
местные анестетики: напр., лидокаин;
2) нарушение синтеза
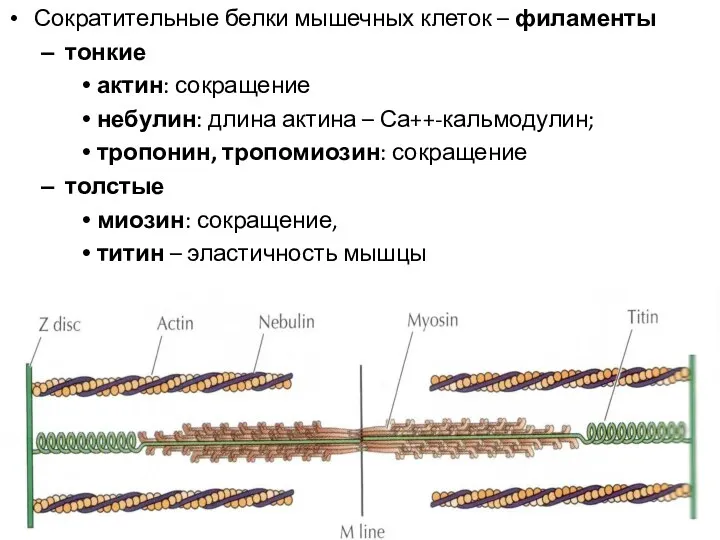
Сократительные белки мышечных клеток – филаменты
тонкие
актин: сокращение
небулин: длина актина –
Сократительные белки мышечных клеток – филаменты
тонкие
актин: сокращение
небулин: длина актина –
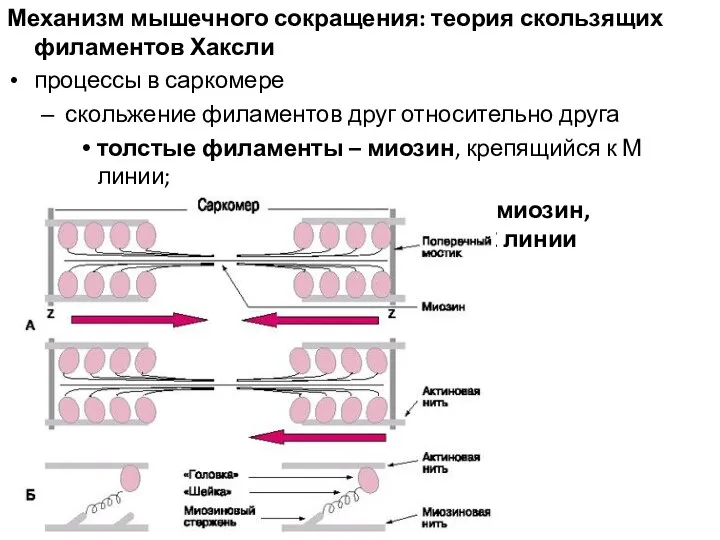
Механизм мышечного сокращения: теория скользящих филаментов Хаксли
процессы в саркомере
скольжение филаментов друг
Механизм мышечного сокращения: теория скользящих филаментов Хаксли
процессы в саркомере
скольжение филаментов друг
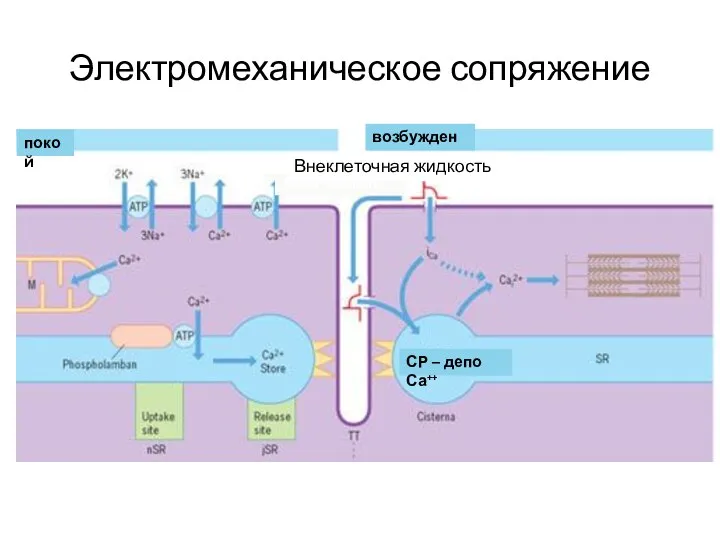
Электромеханическое сопряжение
возбуждение
покой
Внекл. жидкость
СР – депо Са++
Внеклеточная жидкость
Электромеханическое сопряжение
возбуждение
покой
Внекл. жидкость
СР – депо Са++
Внеклеточная жидкость

Итак, в результате электромеханического сопряжения
ПД, распространяясь вглубь волокна, запускает цикл
Итак, в результате электромеханического сопряжения
ПД, распространяясь вглубь волокна, запускает цикл

Цикл поперечных мостиков – АТФ-зависимый процесс
1 цикл – 1 молекула АТФ
Цикл поперечных мостиков – АТФ-зависимый процесс
1 цикл – 1 молекула АТФ

КЛИНИЧЕСКИЕ КОРРЕЛЯТЫ
Фармакотерпия
блокаторы Ca2+ каналов L-типа
антигипертензивная терапия
замедление проводимости в сердце и
КЛИНИЧЕСКИЕ КОРРЕЛЯТЫ
Фармакотерпия
блокаторы Ca2+ каналов L-типа
антигипертензивная терапия
замедление проводимости в сердце и
 Морфология расовых признаков
Морфология расовых признаков Образовательный проект Что мы знаем о лесе
Образовательный проект Что мы знаем о лесе Роль биологических исследований в современной медицине
Роль биологических исследований в современной медицине Программа внеурочной деятельности Юный биолог. (5 класс)
Программа внеурочной деятельности Юный биолог. (5 класс) Эволюция кровеносной системы. Кровь
Эволюция кровеносной системы. Кровь Орган слуха и равновесия
Орган слуха и равновесия Органы размножения. Способы размножения. Оплодотворение
Органы размножения. Способы размножения. Оплодотворение Доказательства эволюции животных
Доказательства эволюции животных Растения. Систематика
Растения. Систематика Звери и птицы осенью
Звери и птицы осенью Вид. Критерии вида. 9 класс
Вид. Критерии вида. 9 класс Насекомые с полным превращением
Насекомые с полным превращением Основные типы гельминтов
Основные типы гельминтов Основные экологические факторы и экологические группы растений
Основные экологические факторы и экологические группы растений Мир микробов
Мир микробов Грибы - гетеротрофы
Грибы - гетеротрофы Техника пикировки растений
Техника пикировки растений Виноград. Качества винограда
Виноград. Качества винограда Закономерности формирования и наследования признаков. Изменчивость
Закономерности формирования и наследования признаков. Изменчивость Обмен белков - 3
Обмен белков - 3 Моллюски, брахиоподы, мшанки, иглокожие
Моллюски, брахиоподы, мшанки, иглокожие презентация по биологии 7 класс
презентация по биологии 7 класс Открытый урок Внешнее строение Насекомых
Открытый урок Внешнее строение Насекомых Основные направления эволюции
Основные направления эволюции Пищеварение в желудке жвачных
Пищеварение в желудке жвачных кошки Иванова
кошки Иванова Собака в городе: жизнь или выживание?
Собака в городе: жизнь или выживание? Класс Млекопитающие. Внешнее строение
Класс Млекопитающие. Внешнее строение