- Выделительная система и осморегуляция рыб

Содержание
- 2. Мальпигиево тельце 1 - артерия; 2 - капсула Шумлянского; 3 - клубочек; 4 - воронки; 5
- 3. Схема строения почек акулы 1 - нефростома (воронка); 2 - мальпигиевы тельца; 3 - кровеносный сосуд,
- 4. Макро и микростроение почек а- головная почка; б, в, д - туловищная почка; г -почка форели;
- 5. Осморегуляция и осморегуляторные приспособления морских и пресноводных рыб В процессах выделения и водно-солевого обмена кроме почек
- 6. Осморегуляция рыб.Схема функциональной системы осморегуляции рыб
- 7. По уровню осмотического давления внутренней среды по отношению к окружающей воде рыбы образуют несколько групп: 1)
- 8. Поддержание солевого состава крови и давления на нужном уровне обусловливается деятельностью: почек, особых клеток стенок почечных
- 9. Основные пути движения воды и солей в процессе осморегуляции у пресноводных и морских костистых рыб Во
- 10. Количество мочи выделяемое рыбами в сутки в зависимости от вида рыб Количество мочи выделяемое рыбами в
- 11. Морские костистые рыбы (с гипотонической кровью), находящиеся в гипертонической среде, постоянно теряют воду – через кожу,
- 12. Морские рыбы выделяют очень мало мочи: в почках у них немного мальпигиевых клубочков, у некоторых их
- 13. Регуляция водно-солевого обмена эвригалинных рыб Эвригалинные организмы, то есть выдерживающие значительное колебание солёности, в частности проходные
- 14. Регуляция водно-солевого обмена у проходных рыб Их почки, кожа и жабры могут функционировать двояко: почки имеют
- 16. Скачать презентацию
 Садовые цветы
Садовые цветы Биология как наука. Методы биологии
Биология как наука. Методы биологии Волокна растительного происхождения
Волокна растительного происхождения Конспект урока. Внутреннее строение рыбы
Конспект урока. Внутреннее строение рыбы Размножение растений семенами. 5 класс
Размножение растений семенами. 5 класс Тест Природа и ваша безопасность
Тест Природа и ваша безопасность Кора больших полушарий
Кора больших полушарий Введение в системную биологию
Введение в системную биологию Вены большого круга кровообращения
Вены большого круга кровообращения Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Клюква
Клюква Майстерність маскування
Майстерність маскування Биополимеры - белки
Биополимеры - белки Анатомия и физиология органов пищеварения
Анатомия и физиология органов пищеварения Многолетние цветущие растения Растения сезонного оформления цветников
Многолетние цветущие растения Растения сезонного оформления цветников Биология – наука о жизни
Биология – наука о жизни Презентация к уроку Обмен веществ и превращение энергии
Презентация к уроку Обмен веществ и превращение энергии Строение семян. 6 класс
Строение семян. 6 класс Антропология. Ранние этапы эволюции человека
Антропология. Ранние этапы эволюции человека Классификация животных. Основные систематические группы. Влияние человека на животных
Классификация животных. Основные систематические группы. Влияние человека на животных РАЗРАБОТКА ПРЕЗЕНТАЦИИ УРОКА ПО БИОЛОГИИ В 11 КЛАССЕ НА ТЕМУ: Движущие силы антропогенеза. Ф. Энгельс о роли труда в процессе превращения обезьяны в человека. Развитие членораздельной речи, сознания и общественных отношений в становлении ч
РАЗРАБОТКА ПРЕЗЕНТАЦИИ УРОКА ПО БИОЛОГИИ В 11 КЛАССЕ НА ТЕМУ: Движущие силы антропогенеза. Ф. Энгельс о роли труда в процессе превращения обезьяны в человека. Развитие членораздельной речи, сознания и общественных отношений в становлении ч Строение и функции отделов нервной системы. Головной мозг
Строение и функции отделов нервной системы. Головной мозг Настоящие грибы или эумицеты
Настоящие грибы или эумицеты Ногти. Строение и функции
Ногти. Строение и функции Жири. Склад жирів. Фізичні властивості. Природні й гідрогенізовані жири. Біологічна роль
Жири. Склад жирів. Фізичні властивості. Природні й гідрогенізовані жири. Біологічна роль Тип Кишечнополостные
Тип Кишечнополостные Мышцы головы
Мышцы головы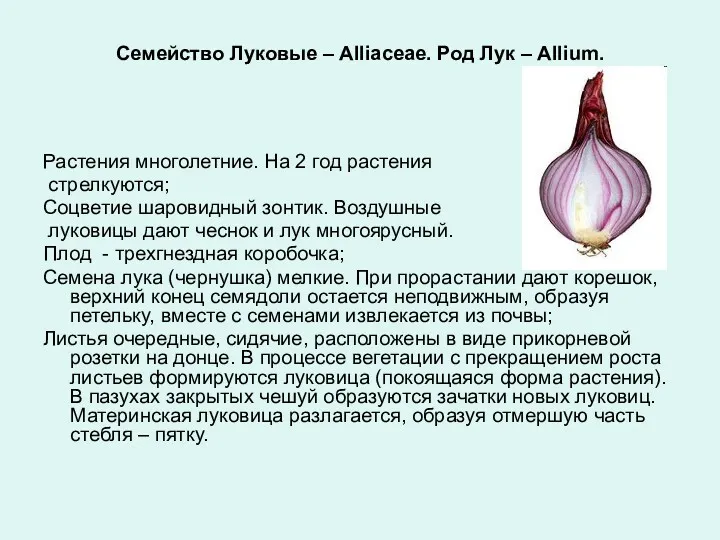 Семейство растений луковые
Семейство растений луковые