- Жасушаның мамандандырылған типін түзуге қатысатын молекулалық-генетикалық механизмдер

Содержание
- 2. Жасушалық жады –жасушаның спецификалық типінің жетілуіне жасуша коммутацияланған болып табылғаннан кейінгі ген экспрессиясының өзгерістерін еске сақтап
- 3. Жоғары сатыдағы эукариоттардың жасушасының жетілуі негізінде ДНҚ тізбегінде қандай да бір өзгерістердің байқалмауы арқылы жүреді. Кейбір
- 4. Осы жетілу механизмі Salmonella бактериясында кездеседі және фазалық вариация түрінде танымал. Геннің ауысуы өлшемі 1000 нуклеотидтік
- 5. Бактерияларда ДНҚ инверсия әдісі арқылы ген экспрессиясының қосылуы
- 6. Нан пісіретін Saccharomyces cerevisiae ашытқысы үш әртүрлі жасуша типіне дифференциациялануға қабілетті. Диплоидты жасушалар екі гаплоидты жасушаның
- 7. Гаплоидтық жасушаның жұптасу типі жалғыз локус арқылы анықталады, яғни жұптасу локусы арқылы (mating-type locus, Mat), ол
- 8. Ашытқылардағы жасуша типін бақылау
- 9. Ашытқы хромосомасында Mat локусының екі жағынан үндемейтін локус бойынша жұптасу типін анықтаушы реттеуші белоктар орналасқан: үндемейтін
- 10. Ашытқылардағы жұптасу типінің ауысуының кассеталық моделі
- 11. Қолайлы жағдайда лямбда бактериофагы E. coli жасушасының ДНҚ-сына орналаса алады және автоматты түрде бактерия бөлінген сайын
- 12. Ген реттелуінің ауысу механизмінің орталығы ретінде вирус арқылы синтезделген екі реттеуші белок табылады: лямбда фагының репрессор
- 13. E. Coli-дің қожайын жасушасындағы лямбда бактериофагының өсу әдісін анықтаушы реттеуші жүйенің оңайлатылған нұсқасы.
- 14. Оң кері байланыстың ілмектері жасушалық жадының қарапайым стратегиясына арналған негіз ретінде қызмет атқарады, яғни ген экспрессиясының
- 15. Оң кері байланыс ілмегінің жасушалық жадыны қалыптастыруын көрсететін схемалық сурет
- 16. Реттеуші тізбекті зерттеу кезінде құрылымның белгілі бір қарапайым типтері әртүрлі түрлердің жасушаларынан табылғандығы анықталды. Мысалы, оң
- 17. Транскрипциялық тізбектердегі желілік мотивтердің кең тараған типтері
- 18. Тура байланыс ілмегінің сигналдың ұзақтығын қалай өлшей алатындығы
- 19. Теңіз кірпісінің дамып келе жатқан эмбрионының бөлігін анықтаушы төтенше күрделі генетикалық тізбегі
- 20. Жердегі тіршілік түн және күннің тәуліктік циклының болу жағдайында дамып отырды және көптеген ағзалар, яғни архейден
- 21. Ми жасушасының мамандандырылған тобының әр участогының ішіндегі әрекет етуші саға ұйқының тәуліктік циклын, сергектікті, дене температурасын,
- 22. Зертхана жасалған қарапайым гендік осциллятор немесе сағат
- 23. Tim (timeless) және Per (period) екі реттеуші белоктардың ыдырауы мен үздікті жиналуы Дрозофиладағы циркадты сағаттардың негізгі
- 24. Дрозофила жасушасындағы циркадты сағат жұмысының механизмінің жеңілдетілген схемасы
- 25. Ген экспрессиясының бақылауы комбинаторлық процес болып табылатындығына қарамастан, бір жалғыз реттеуші белоктың әрекеті белгілі генді өшіру
- 26. Бір реттеуші белок бірнеше әртүрлі гендердің экспрессиясын координациялауы мүмкін
- 27. Сүтқоректілердің қаңқа бұлшықетінің жасушасы миобласт деп аталатын ізашар-жасушалардың құйылысуының нәтижесінде түзілгенжоғары дифференциацияланған гигант жасуша болып табылады
- 28. Бұлшықеттің дамуындағы миогендік реттеуші белоктың рөлі
- 29. Бірнеше реттеуші белоктардан тұратын комбинация ағзаның даму барысындағы жасушаның көптеген типінің қалыптасуын қамтамасыз етеді. Жасушаның комбинаторлық
- 30. Ағзаның дамуына арналған геннің комбинаторлық реттелуінің маңыздылығы
- 31. Егер де бір реттеуші белок белоктардың сәйкес келетін комбинациясын аяқтайтын болса, онда ол геннің тұтас жинағын
- 32. Дрозофиладағы Еу генінің экспрессиясы Ізашар жасушадағы аяқтар аяқтардағы көздің дамуына бастамашылық жасайды
- 34. Дрозофиланың көзінің дамуын анықтаушы реттеуші белоктар. Тоу (Twin of eyeless) және Еу (Eyeless) ұқсас реттеуші белоктарды
- 35. Омыртқалылардың жасушасында цитозиннің метилденуі күшті механизм болып табылады, соның салдарынан ген экспрессиясы профилінің туынды жасушаға берілуі
- 36. 5-метилцитозиннің түзілуі цитозиннің ДНҚ-ның қос спиралінің метилденуі кезінде жүреді.
- 37. Омыртқалылардағы ДНҚ метилдену цитозиннің нуклеотидтерімен CG тізбегінде шектеледі. Осы метилдену типінің ДНҚ-ның туынды тізбегімен тура тұқымқуалауы
- 38. ДНҚ метилдену профилінің нақты тұқымқуалауы қалай жүреді
- 39. ДНҚ метилдену профилі омыртқалылардың даму барысында серпінді өзгереді. Ұрықтанғаннан кейін геном бойынша деметилдену толқыны жүріп өтеді
- 40. Дамудың кеш сатысында метилденудің жаңа типтері сайт спецификалық ДНҚ байланыстырушы белоктары арқылы ДНҚ-ға бағытталған бірнеше de
- 41. Геннің тұрақты репрессиясы арқылы көптеген механизмдер іске қосылған
- 42. Геннің үлкен емес бөлігінің экспрессиясы олардың әкесінен немесе шешесінен берілгендігіне байланысты болады: әзірше геннің аталық көшірмесі
- 43. Тышқандардағы импринтинг
- 44. Тышқандардағы Igf2 гені импринтингінің механизмі. Шешесінен берілген хромосомада CTCF деп аталатын белок Igf2 гені мен энхансер
- 45. CG тізбегі геном бойымен біркелкі емес таралған: ұзындығы 1000-2000 нуклеотидтен тұратын жеке аймақтар бар, оларды CG-аралшықтар
- 46. Сүтқоректілердің «үй шаруашылығы» деп аталатын үш геніндегі промоторларды қоршап алатын CGаралшықтар
- 47. CG тізбегінің айқын ортақ жетіспеушілігін және олардың омыртқалылар геномындағы CG аралшықтарына топтасуын түсіндіретін механизм. Жыныс жасушаларында
- 48. Ағзадағы тұқымқуалаудың эпигенетикалық формасын тудыратын төрт әртүрлі механизм
- 49. Жеке-жеке өскен бір жұмыртқалы егіздер
- 50. Аталық және аналық дарақтар өздерінің жыныс хромосомалары арқылы бір бірінен ерекшеленеді. Аналықта екі Х хромосомасы бар,
- 51. Аналық жасушалардағы Х хромосоманың инактивациясы
- 52. Сүтқоректілерде ген дозасының компенсациясы аналық соматикалық жасушасындағы екі Х-хромосоманың біреуінің транскрипциясының инактивациясы арқылы белгілі мөлшерге жетеді.
- 53. Х хромосома инактивациясы
- 54. Х хромосоманың инактивациясы Х хромосоманың ортасындағы жалғыз участоктан, Х хромосомасының инактиввация орталығынан (X-inactivation center, XIC) басталып,
- 55. Сүтқоректілердегі Х хромосоманың инактивациясы
- 56. Құрамында XISТ-РНК –ның болғанынан бөлек Х хромосоманың гетерохроматині 2А гистонының спецификалық нұсқасының болуымен, Н3 және Н4
- 57. Жасуша әрекетіне кездейсоқ ауытқулардың көп мөлшері тән. Бұл жасуша әрекетінің фундаментальды жағы. Біртіндеп бұл қоршаған ортада
- 59. Скачать презентацию

Жасушалық жады –жасушаның спецификалық типінің жетілуіне жасуша коммутацияланған болып табылғаннан кейінгі
Жасушалық жады –жасушаның спецификалық типінің жетілуіне жасуша коммутацияланған болып табылғаннан кейінгі

Жоғары сатыдағы эукариоттардың жасушасының жетілуі негізінде ДНҚ тізбегінде қандай да бір
Жоғары сатыдағы эукариоттардың жасушасының жетілуі негізінде ДНҚ тізбегінде қандай да бір

Осы жетілу механизмі Salmonella бактериясында кездеседі және фазалық вариация түрінде танымал.
Геннің
Осы жетілу механизмі Salmonella бактериясында кездеседі және фазалық вариация түрінде танымал.
Геннің
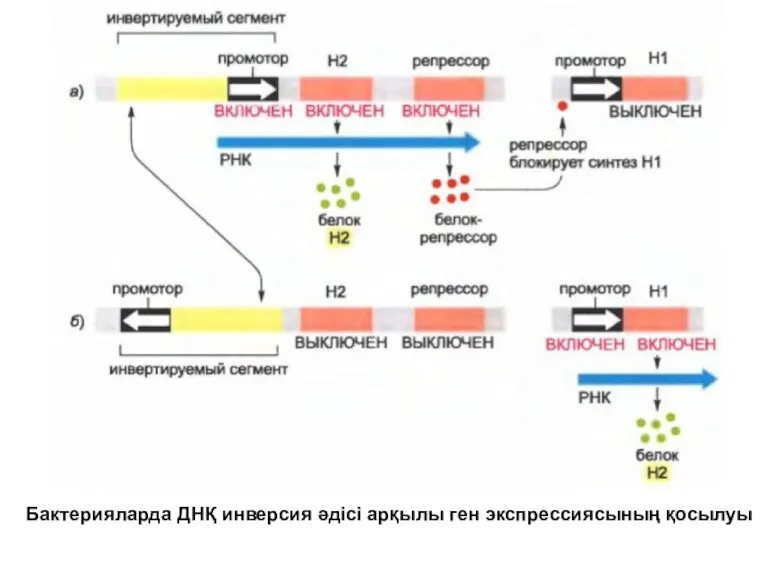
Бактерияларда ДНҚ инверсия әдісі арқылы ген экспрессиясының қосылуы
Бактерияларда ДНҚ инверсия әдісі арқылы ген экспрессиясының қосылуы

Нан пісіретін Saccharomyces cerevisiae ашытқысы үш әртүрлі жасуша типіне дифференциациялануға қабілетті.
Нан пісіретін Saccharomyces cerevisiae ашытқысы үш әртүрлі жасуша типіне дифференциациялануға қабілетті.

Гаплоидтық жасушаның жұптасу типі жалғыз локус арқылы анықталады, яғни жұптасу локусы
Гаплоидтық жасушаның жұптасу типі жалғыз локус арқылы анықталады, яғни жұптасу локусы
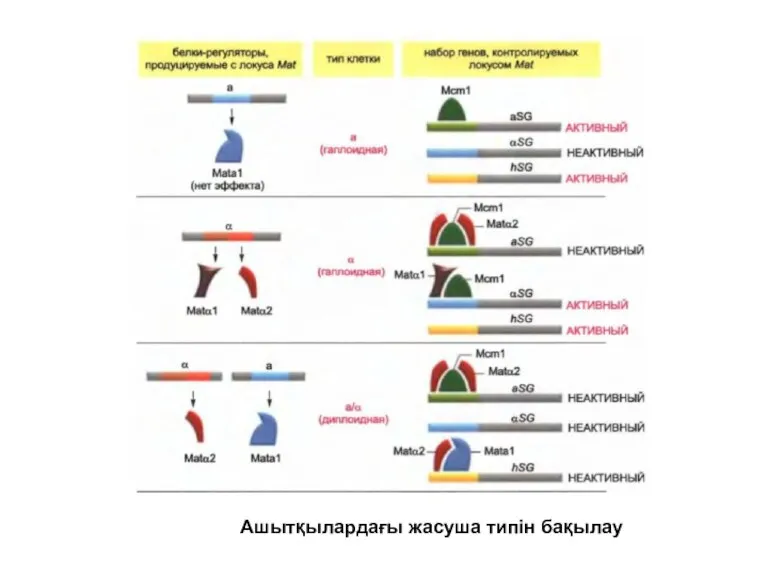
Ашытқылардағы жасуша типін бақылау
Ашытқылардағы жасуша типін бақылау

Ашытқы хромосомасында Mat локусының екі жағынан үндемейтін локус бойынша жұптасу типін
Ашытқы хромосомасында Mat локусының екі жағынан үндемейтін локус бойынша жұптасу типін
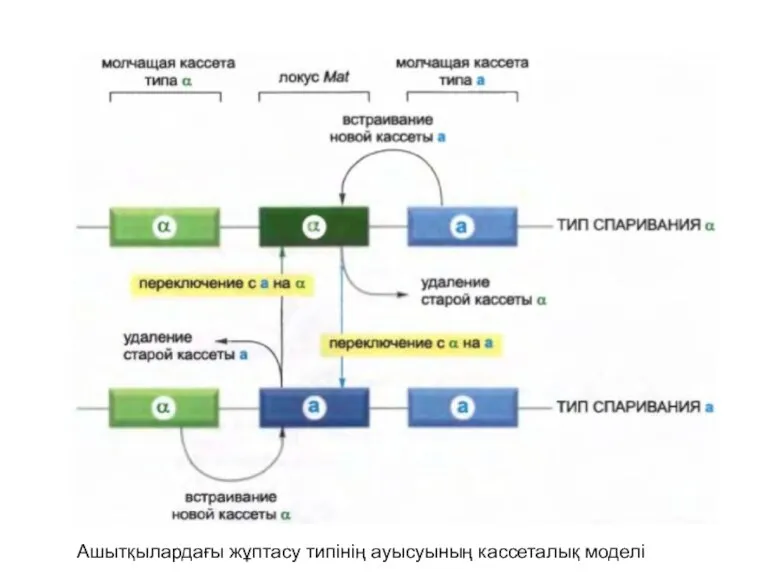
Ашытқылардағы жұптасу типінің ауысуының кассеталық моделі
Ашытқылардағы жұптасу типінің ауысуының кассеталық моделі

Қолайлы жағдайда лямбда бактериофагы E. coli жасушасының ДНҚ-сына орналаса алады және
Қолайлы жағдайда лямбда бактериофагы E. coli жасушасының ДНҚ-сына орналаса алады және

Ген реттелуінің ауысу механизмінің орталығы ретінде вирус арқылы синтезделген екі реттеуші
Ген реттелуінің ауысу механизмінің орталығы ретінде вирус арқылы синтезделген екі реттеуші
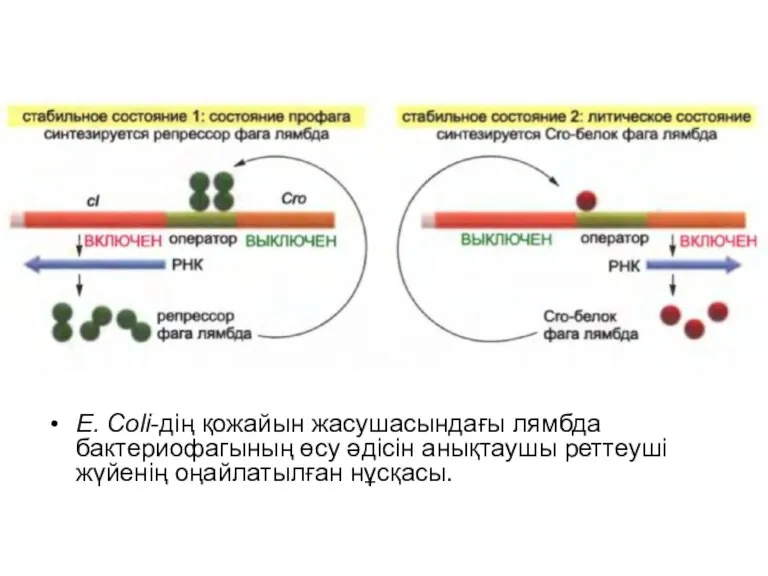
E. Coli-дің қожайын жасушасындағы лямбда бактериофагының өсу әдісін анықтаушы реттеуші жүйенің
E. Coli-дің қожайын жасушасындағы лямбда бактериофагының өсу әдісін анықтаушы реттеуші жүйенің

Оң кері байланыстың ілмектері жасушалық жадының қарапайым стратегиясына арналған негіз ретінде
Оң кері байланыстың ілмектері жасушалық жадының қарапайым стратегиясына арналған негіз ретінде
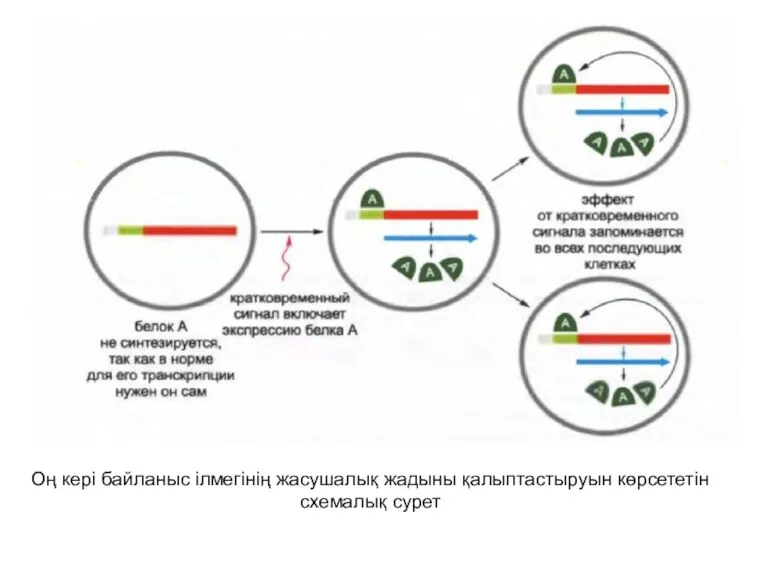
Оң кері байланыс ілмегінің жасушалық жадыны қалыптастыруын көрсететін схемалық сурет
Оң кері байланыс ілмегінің жасушалық жадыны қалыптастыруын көрсететін схемалық сурет

Реттеуші тізбекті зерттеу кезінде құрылымның белгілі бір қарапайым типтері әртүрлі түрлердің
Реттеуші тізбекті зерттеу кезінде құрылымның белгілі бір қарапайым типтері әртүрлі түрлердің
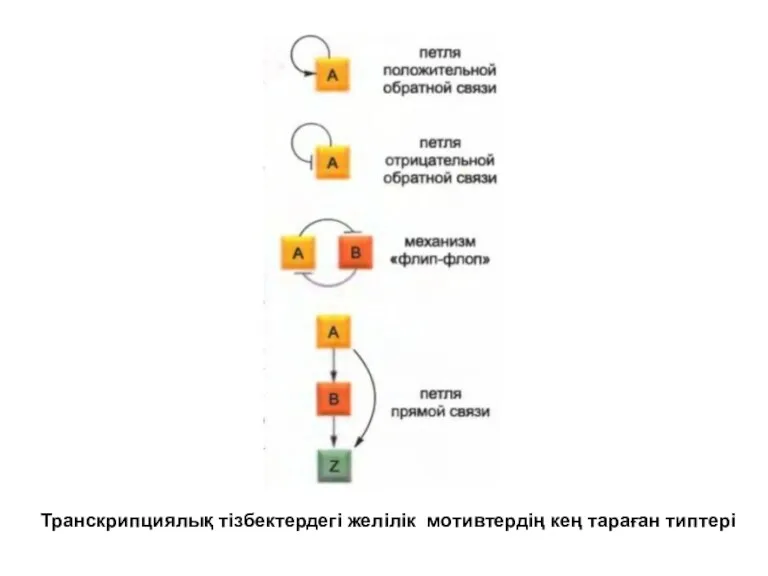
Транскрипциялық тізбектердегі желілік мотивтердің кең тараған типтері
Транскрипциялық тізбектердегі желілік мотивтердің кең тараған типтері
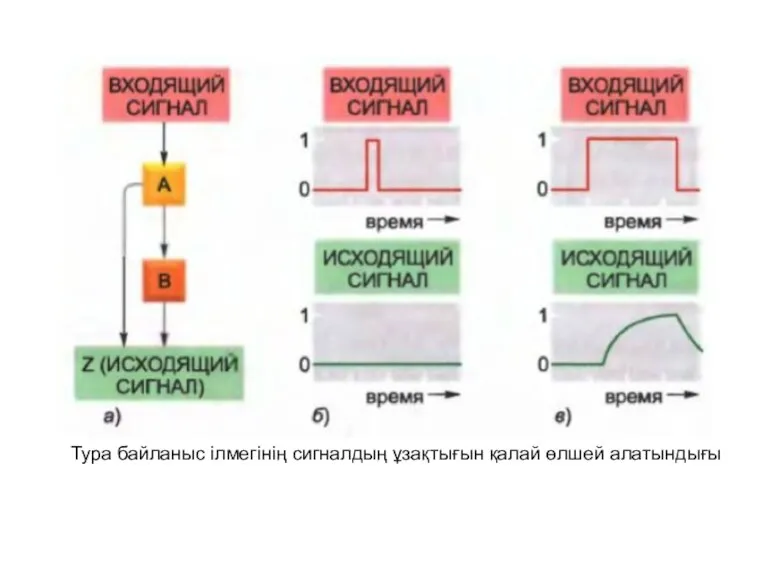
Тура байланыс ілмегінің сигналдың ұзақтығын қалай өлшей алатындығы
Тура байланыс ілмегінің сигналдың ұзақтығын қалай өлшей алатындығы
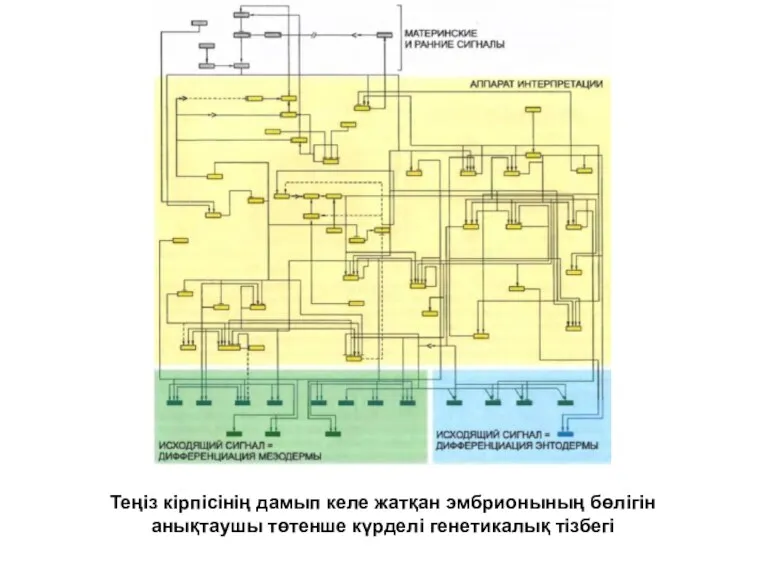
Теңіз кірпісінің дамып келе жатқан эмбрионының бөлігін анықтаушы төтенше күрделі генетикалық
Теңіз кірпісінің дамып келе жатқан эмбрионының бөлігін анықтаушы төтенше күрделі генетикалық

Жердегі тіршілік түн және күннің тәуліктік циклының болу жағдайында дамып отырды
Жердегі тіршілік түн және күннің тәуліктік циклының болу жағдайында дамып отырды

Ми жасушасының мамандандырылған тобының әр участогының ішіндегі әрекет етуші саға ұйқының
Ми жасушасының мамандандырылған тобының әр участогының ішіндегі әрекет етуші саға ұйқының
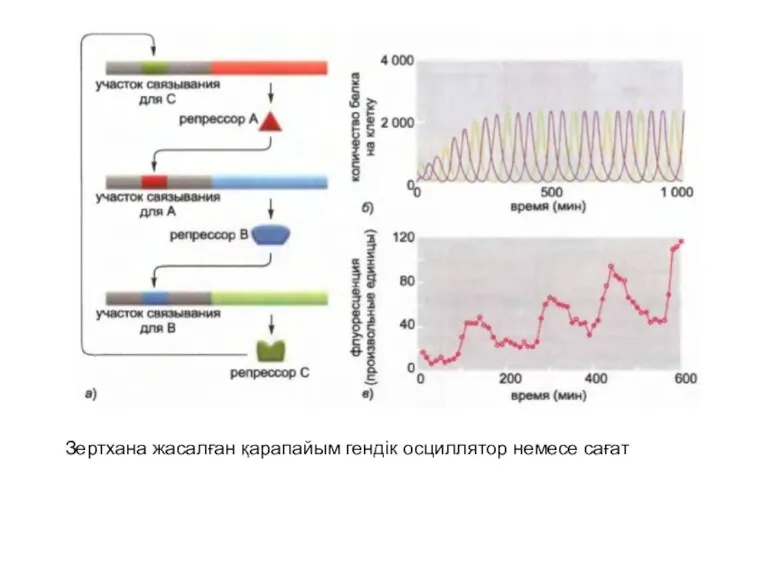
Зертхана жасалған қарапайым гендік осциллятор немесе сағат
Зертхана жасалған қарапайым гендік осциллятор немесе сағат

Tim (timeless) және Per (period) екі реттеуші белоктардың ыдырауы мен үздікті
Tim (timeless) және Per (period) екі реттеуші белоктардың ыдырауы мен үздікті
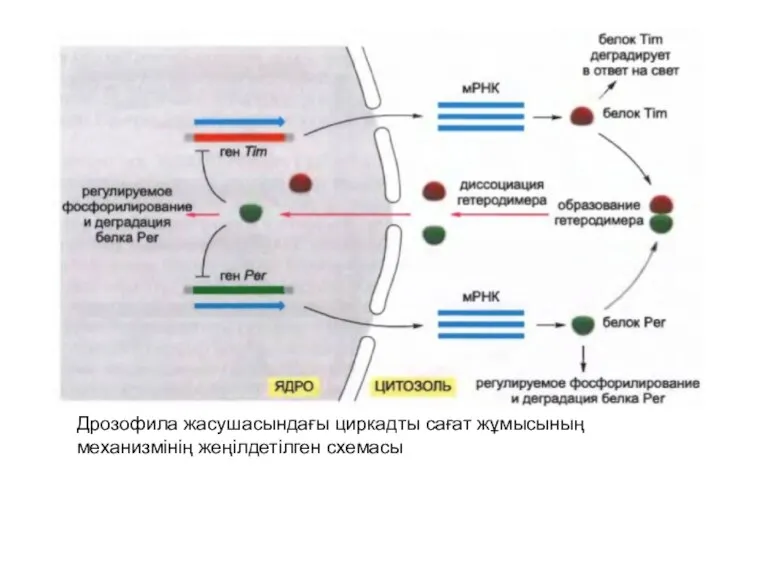
Дрозофила жасушасындағы циркадты сағат жұмысының механизмінің жеңілдетілген схемасы
Дрозофила жасушасындағы циркадты сағат жұмысының механизмінің жеңілдетілген схемасы

Ген экспрессиясының бақылауы комбинаторлық процес болып табылатындығына қарамастан, бір жалғыз реттеуші
Ген экспрессиясының бақылауы комбинаторлық процес болып табылатындығына қарамастан, бір жалғыз реттеуші
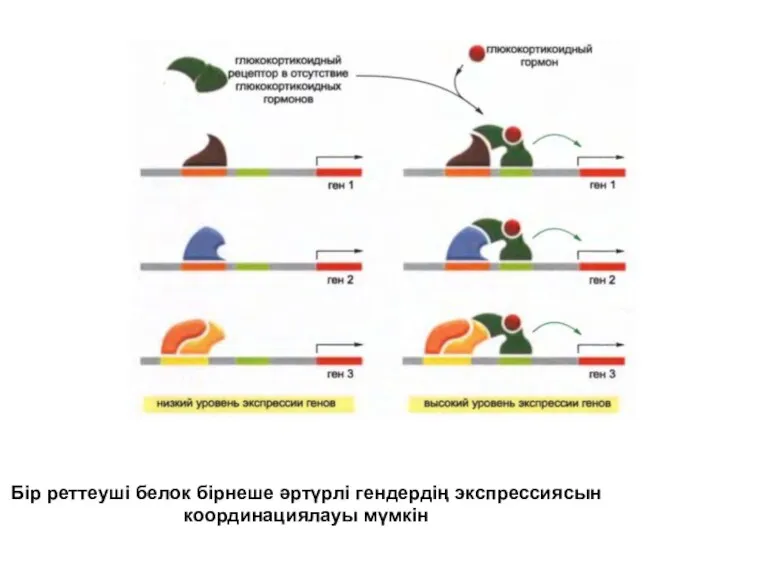
Бір реттеуші белок бірнеше әртүрлі гендердің экспрессиясын координациялауы мүмкін
Бір реттеуші белок бірнеше әртүрлі гендердің экспрессиясын координациялауы мүмкін

Сүтқоректілердің қаңқа бұлшықетінің жасушасы миобласт деп аталатын ізашар-жасушалардың құйылысуының нәтижесінде түзілгенжоғары
Сүтқоректілердің қаңқа бұлшықетінің жасушасы миобласт деп аталатын ізашар-жасушалардың құйылысуының нәтижесінде түзілгенжоғары
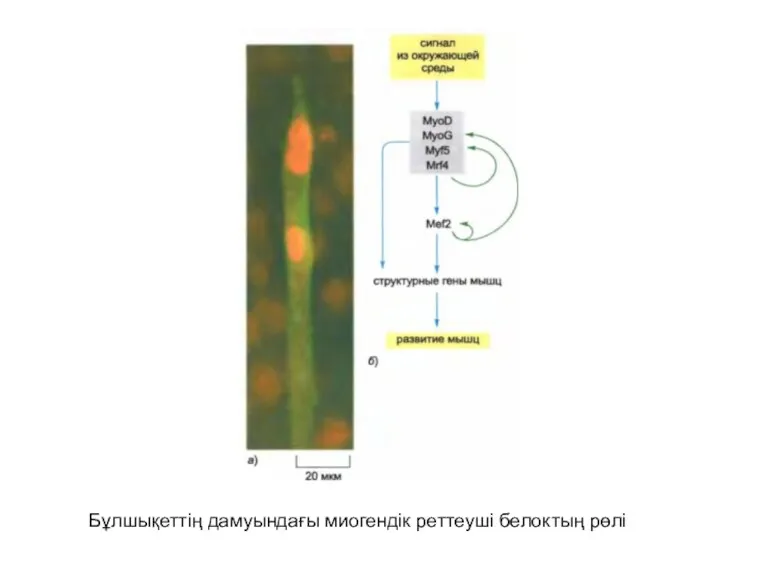
Бұлшықеттің дамуындағы миогендік реттеуші белоктың рөлі
Бұлшықеттің дамуындағы миогендік реттеуші белоктың рөлі

Бірнеше реттеуші белоктардан тұратын комбинация ағзаның даму барысындағы жасушаның көптеген типінің
Бірнеше реттеуші белоктардан тұратын комбинация ағзаның даму барысындағы жасушаның көптеген типінің
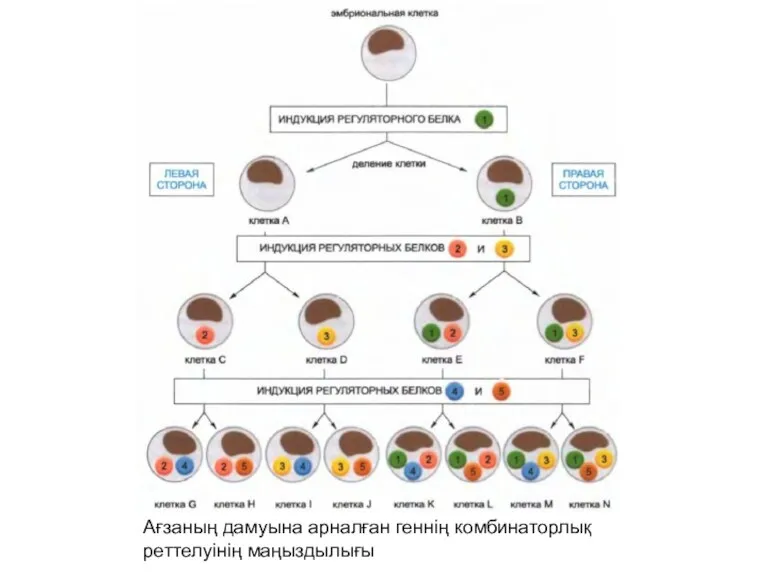
Ағзаның дамуына арналған геннің комбинаторлық реттелуінің маңыздылығы
Ағзаның дамуына арналған геннің комбинаторлық реттелуінің маңыздылығы

Егер де бір реттеуші белок белоктардың сәйкес келетін комбинациясын аяқтайтын болса,
Егер де бір реттеуші белок белоктардың сәйкес келетін комбинациясын аяқтайтын болса,
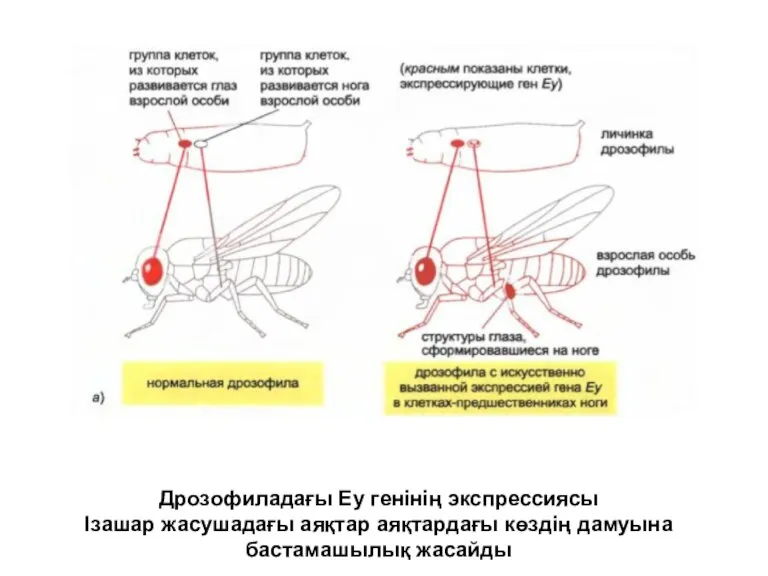
Дрозофиладағы Еу генінің экспрессиясы
Ізашар жасушадағы аяқтар аяқтардағы көздің дамуына бастамашылық
Дрозофиладағы Еу генінің экспрессиясы
Ізашар жасушадағы аяқтар аяқтардағы көздің дамуына бастамашылық

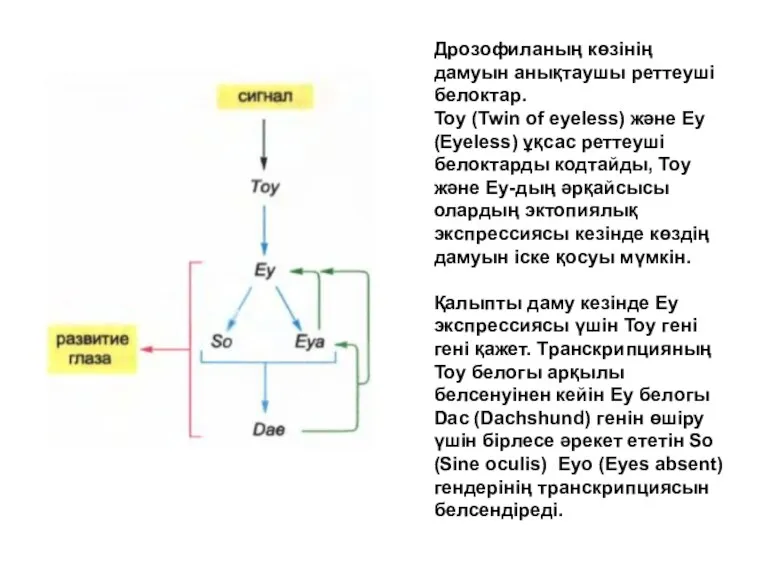
Дрозофиланың көзінің дамуын анықтаушы реттеуші белоктар.
Тоу (Twin of eyeless) және Еу
Дрозофиланың көзінің дамуын анықтаушы реттеуші белоктар.
Тоу (Twin of eyeless) және Еу

Омыртқалылардың жасушасында цитозиннің метилденуі күшті механизм болып табылады, соның салдарынан ген
Омыртқалылардың жасушасында цитозиннің метилденуі күшті механизм болып табылады, соның салдарынан ген
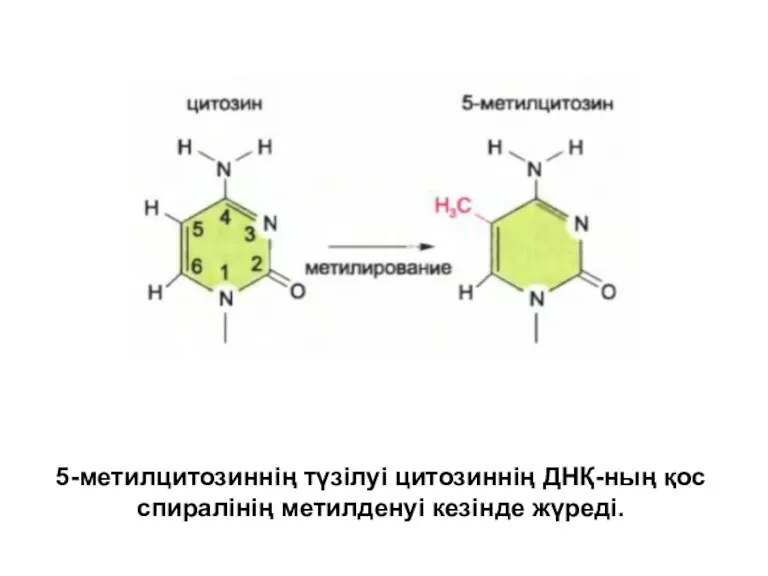
5-метилцитозиннің түзілуі цитозиннің ДНҚ-ның қос спиралінің метилденуі кезінде жүреді.
5-метилцитозиннің түзілуі цитозиннің ДНҚ-ның қос спиралінің метилденуі кезінде жүреді.

Омыртқалылардағы ДНҚ метилдену цитозиннің нуклеотидтерімен CG тізбегінде шектеледі.
Осы метилдену типінің
Омыртқалылардағы ДНҚ метилдену цитозиннің нуклеотидтерімен CG тізбегінде шектеледі.
Осы метилдену типінің
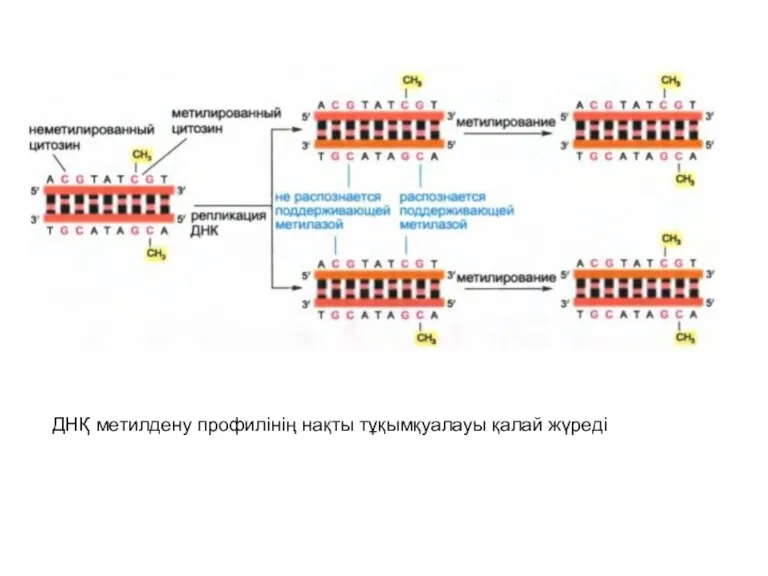
ДНҚ метилдену профилінің нақты тұқымқуалауы қалай жүреді
ДНҚ метилдену профилінің нақты тұқымқуалауы қалай жүреді

ДНҚ метилдену профилі омыртқалылардың даму барысында серпінді өзгереді.
Ұрықтанғаннан кейін геном
ДНҚ метилдену профилі омыртқалылардың даму барысында серпінді өзгереді.
Ұрықтанғаннан кейін геном

Дамудың кеш сатысында метилденудің жаңа типтері сайт спецификалық ДНҚ байланыстырушы белоктары
Дамудың кеш сатысында метилденудің жаңа типтері сайт спецификалық ДНҚ байланыстырушы белоктары
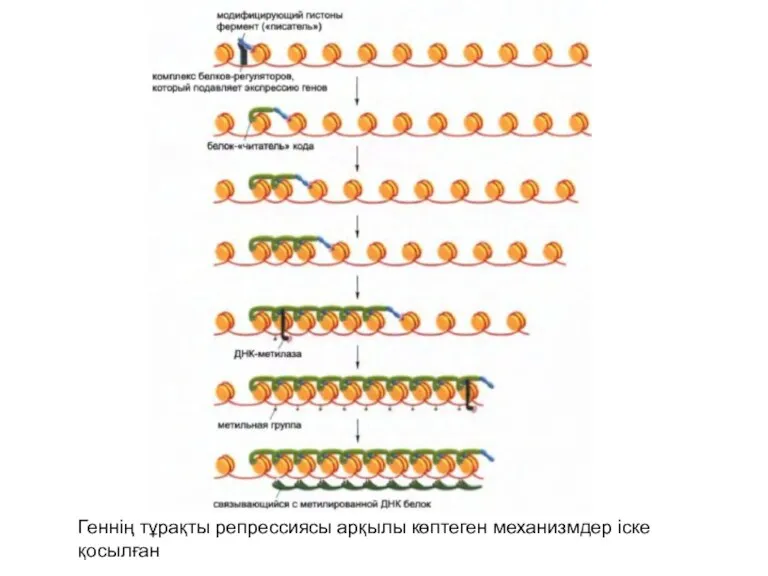
Геннің тұрақты репрессиясы арқылы көптеген механизмдер іске қосылған
Геннің тұрақты репрессиясы арқылы көптеген механизмдер іске қосылған

Геннің үлкен емес бөлігінің экспрессиясы олардың әкесінен немесе шешесінен берілгендігіне байланысты
Геннің үлкен емес бөлігінің экспрессиясы олардың әкесінен немесе шешесінен берілгендігіне байланысты
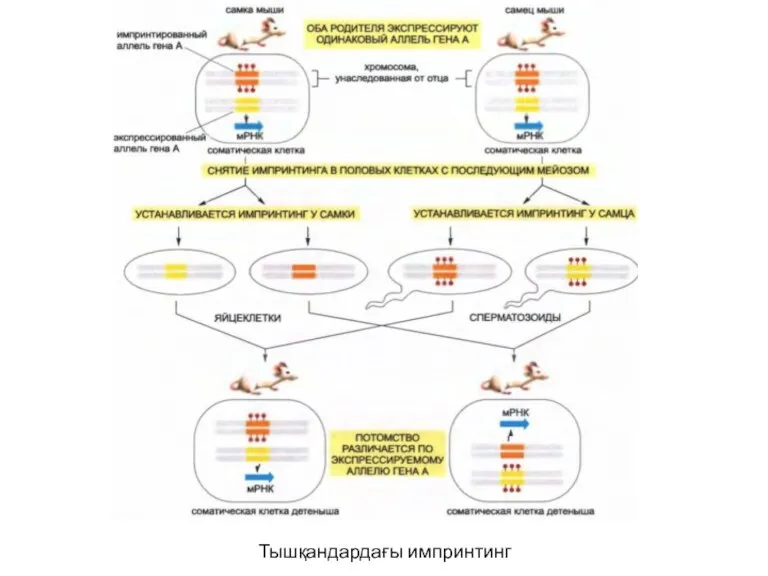
Тышқандардағы импринтинг
Тышқандардағы импринтинг
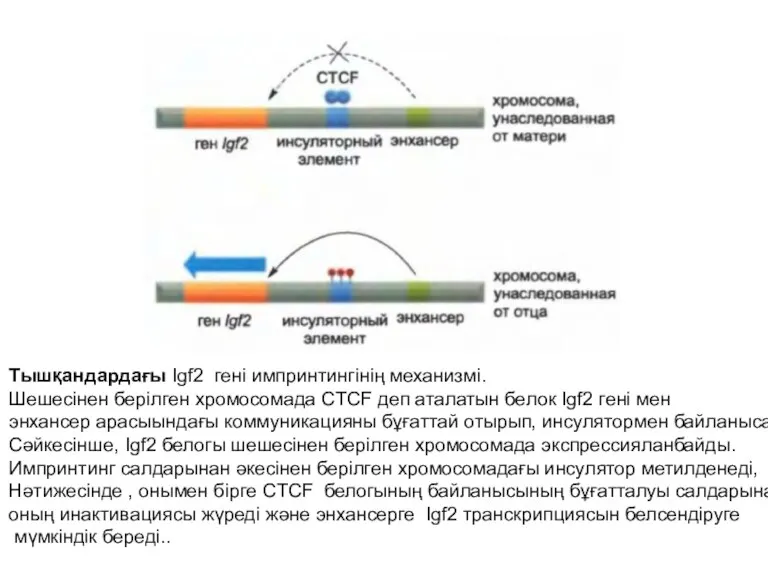
Тышқандардағы Igf2 гені импринтингінің механизмі.
Шешесінен берілген хромосомада CTCF деп аталатын
Тышқандардағы Igf2 гені импринтингінің механизмі.
Шешесінен берілген хромосомада CTCF деп аталатын

CG тізбегі геном бойымен біркелкі емес таралған: ұзындығы 1000-2000 нуклеотидтен тұратын
CG тізбегі геном бойымен біркелкі емес таралған: ұзындығы 1000-2000 нуклеотидтен тұратын
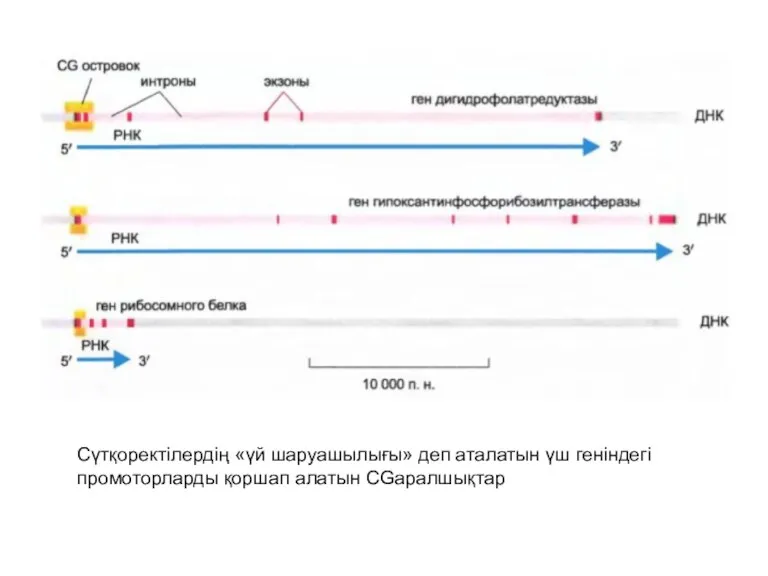
Сүтқоректілердің «үй шаруашылығы» деп аталатын үш геніндегі промоторларды қоршап алатын CGаралшықтар
Сүтқоректілердің «үй шаруашылығы» деп аталатын үш геніндегі промоторларды қоршап алатын CGаралшықтар

CG тізбегінің айқын ортақ жетіспеушілігін және олардың омыртқалылар геномындағы CG аралшықтарына
CG тізбегінің айқын ортақ жетіспеушілігін және олардың омыртқалылар геномындағы CG аралшықтарына
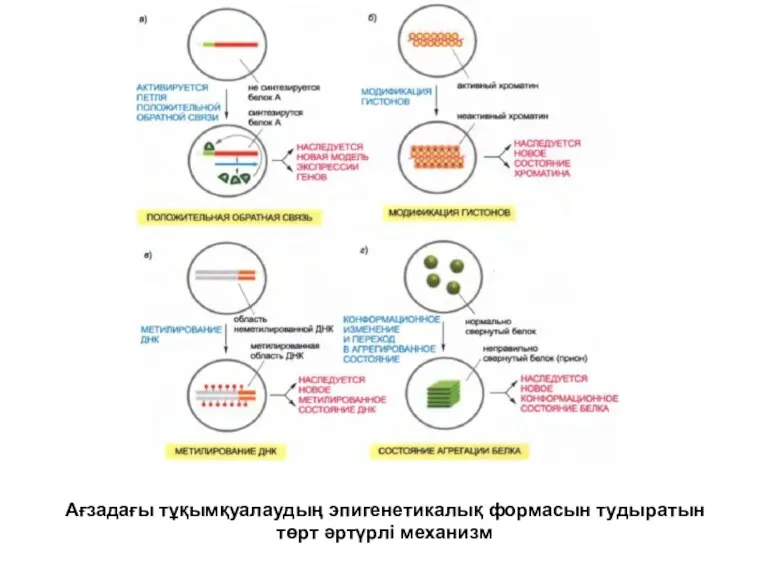
Ағзадағы тұқымқуалаудың эпигенетикалық формасын тудыратын төрт әртүрлі механизм
Ағзадағы тұқымқуалаудың эпигенетикалық формасын тудыратын төрт әртүрлі механизм

Жеке-жеке өскен бір жұмыртқалы егіздер
Жеке-жеке өскен бір жұмыртқалы егіздер

Аталық және аналық дарақтар өздерінің жыныс хромосомалары арқылы бір бірінен ерекшеленеді.
Аталық және аналық дарақтар өздерінің жыныс хромосомалары арқылы бір бірінен ерекшеленеді.
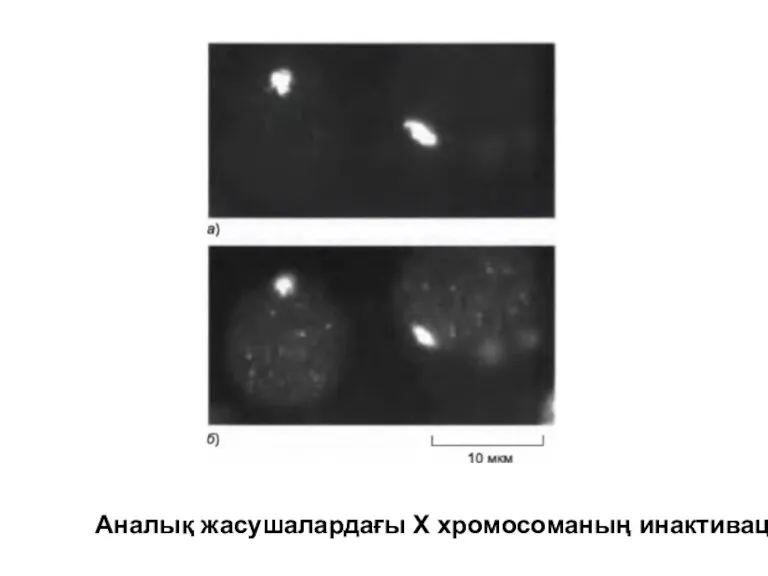
Аналық жасушалардағы Х хромосоманың инактивациясы
Аналық жасушалардағы Х хромосоманың инактивациясы

Сүтқоректілерде ген дозасының компенсациясы аналық соматикалық жасушасындағы екі Х-хромосоманың біреуінің транскрипциясының
Сүтқоректілерде ген дозасының компенсациясы аналық соматикалық жасушасындағы екі Х-хромосоманың біреуінің транскрипциясының
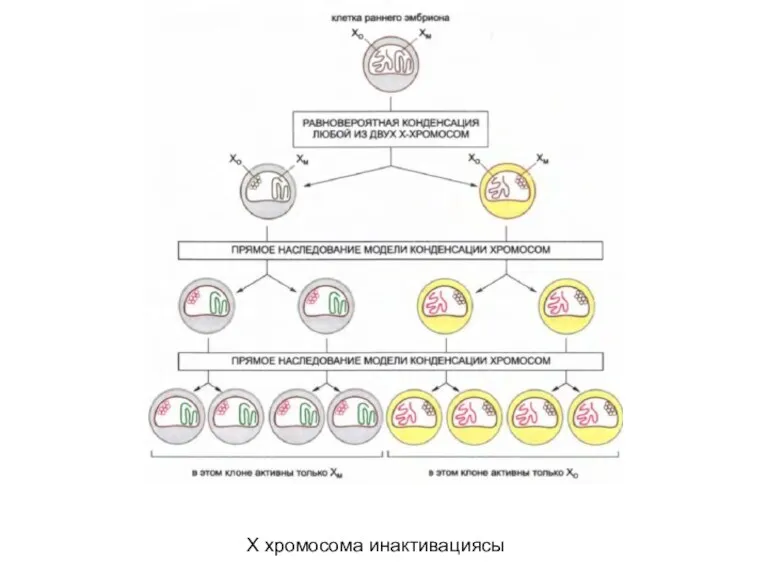
Х хромосома инактивациясы
Х хромосома инактивациясы

Х хромосоманың инактивациясы Х хромосоманың ортасындағы жалғыз участоктан, Х хромосомасының инактиввация
Х хромосоманың инактивациясы Х хромосоманың ортасындағы жалғыз участоктан, Х хромосомасының инактиввация
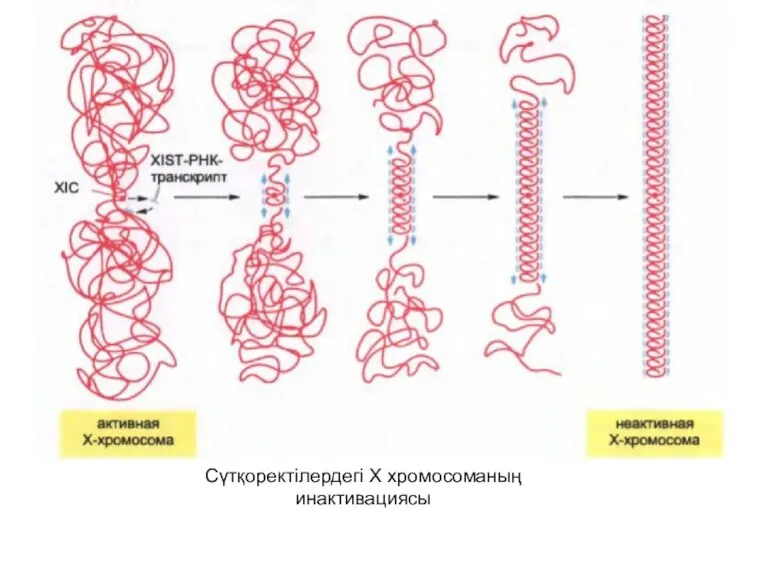
Сүтқоректілердегі Х хромосоманың инактивациясы
Сүтқоректілердегі Х хромосоманың инактивациясы

Құрамында XISТ-РНК –ның болғанынан бөлек Х хромосоманың гетерохроматині 2А гистонының спецификалық
Құрамында XISТ-РНК –ның болғанынан бөлек Х хромосоманың гетерохроматині 2А гистонының спецификалық

Жасуша әрекетіне кездейсоқ ауытқулардың көп мөлшері тән. Бұл жасуша әрекетінің фундаментальды
Жасуша әрекетіне кездейсоқ ауытқулардың көп мөлшері тән. Бұл жасуша әрекетінің фундаментальды
 Витамины. Определение, классификация
Витамины. Определение, классификация Информационная биология. Семантическая и прагматическая информация. (Тема 5)
Информационная биология. Семантическая и прагматическая информация. (Тема 5) Круги кровообращения
Круги кровообращения Строение нервной системы. Спинной мозг
Строение нервной системы. Спинной мозг ЕГЭ по биологии
ЕГЭ по биологии Физиология центральной нервной системы
Физиология центральной нервной системы Осенние приметы. (Урок 15. 1 класс)
Осенние приметы. (Урок 15. 1 класс) Открытый урок биологии по теме Движение 6 класс (УМК Н.И.Сонин) , автор учитель биологии Сизова С.В.
Открытый урок биологии по теме Движение 6 класс (УМК Н.И.Сонин) , автор учитель биологии Сизова С.В. Класс Пресмыкающиеся или Рептилии (Reptilia)
Класс Пресмыкающиеся или Рептилии (Reptilia) Фотосинтез
Фотосинтез Организм человека. Анатомия человека
Организм человека. Анатомия человека Биология кошек
Биология кошек Технология выращивания томатов
Технология выращивания томатов Теории происхождения человека
Теории происхождения человека Породи свиней
Породи свиней Бактериологическая разведка и индикация бактериологического (биологического) оружия
Бактериологическая разведка и индикация бактериологического (биологического) оружия Методы оценки биоразнообразия
Методы оценки биоразнообразия Размножение и развитие земноводных. Сезонные явления в жизни земноводных
Размножение и развитие земноводных. Сезонные явления в жизни земноводных Мал азықтық жемдік дақылдар
Мал азықтық жемдік дақылдар Газообмен в легких и тканях. Дыхательные движения. Регуляция дыхания
Газообмен в легких и тканях. Дыхательные движения. Регуляция дыхания Бабочки
Бабочки Нуклеиновые кислоты
Нуклеиновые кислоты Молекулярные механизмы образования хромосомных перестроек с учетом структурной организации хромосомных районов
Молекулярные механизмы образования хромосомных перестроек с учетом структурной организации хромосомных районов Царство грибы. Общая характеристика
Царство грибы. Общая характеристика презентация к уроку биологии в 8 классе
презентация к уроку биологии в 8 классе Изготовление компоста из листьев
Изготовление компоста из листьев Гепард — хищное млекопитающее семейства кошачьих
Гепард — хищное млекопитающее семейства кошачьих Внутреннее строение корня
Внутреннее строение корня