- Развитие экосистемы

Содержание
- 2. Экологическая сукцессия Наблюдение над полем, заброшенным на несколько лет, показывает, что его последовательно завоевывают сначала многолетние
- 3. Пример сукцессии, приводящей к смене одного сообщества другим, — зарастание небольшого озера с последующим появлением на
- 4. Экологическая сукцессия Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении экологической сукцессии следует учитывать
- 5. Экологическая сукцессия Типы сукцессионных смен. Выделяют два главных типа сукцессионных смен: 1 — с участием автотрофного
- 6. Первичной сукцессией называется процесс развития и смены экосистем на незаселенных ранее участках, начинающихся с их колонизации.
- 7. Первичная сукцессия
- 8. Вторичная сукцессия — это восстановление экосистемы, когда-то уже существовавшей на данной территории. В.с. начинается в том
- 9. Вторичная сукцессия
- 10. Вторичная сукцессия сибирского темно-хвойного леса (пихтово-кедровой тайги) после опустошительного лесного пожара (по Н. Ф. Реймерсу, 1990)
- 11. Климаксовоя экосистема Сукцессия завершается стадией, когда все виды экосистемы, размножаясь, сохраняют относительно постоянную численность и дальнейшей
- 12. Специфические закономерности сукцессии Американский эколог Ф. Клементс, наиболее полно разработавший учение о сукцессиях, считает, что в
- 13. Специфические закономерности сукцессии Наряду с теорией моноклимакса существует точка зрения, в соответствии с которой в одном
- 14. Антропогенные сукцессии Наряду с природными факторами, причинами динамики экосистем выступает человек. К настоящему времени им разрушено
- 15. Общие закономерности сукцессионного процесса 1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. По
- 16. Общие закономерности сукцессионного процесса 5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов,
- 17. Кривая изменения общей биомассы (B) обычно имеет выпуклую форму, тогда как кривая изменения валовой первичной продукции
- 18. Положение сообществ различных типов в классификации, основанной на метаболизме (по: Одум, 1975). Направления автотрофной и гетеротрофной
- 19. Изменение признаков экосистемы в ходе сукцессии
- 20. Законы развития экосистем Закон последовательности прохождения фаз развития: фазы развития природной системы могут следовать лишь в
- 22. Скачать презентацию

Экологическая сукцессия
Наблюдение над полем, заброшенным на несколько лет, показывает, что его
Экологическая сукцессия
Наблюдение над полем, заброшенным на несколько лет, показывает, что его
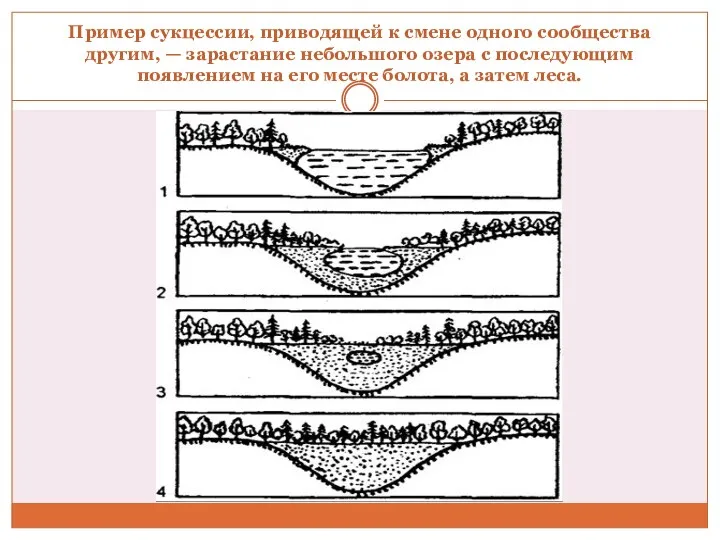
Пример сукцессии, приводящей к смене одного сообщества другим, — зарастание небольшого
Пример сукцессии, приводящей к смене одного сообщества другим, — зарастание небольшого

Экологическая сукцессия
Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении
Экологическая сукцессия
Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении

Экологическая сукцессия
Типы сукцессионных смен. Выделяют два главных типа сукцессионных смен: 1
Экологическая сукцессия
Типы сукцессионных смен. Выделяют два главных типа сукцессионных смен: 1

Первичной сукцессией называется процесс развития и смены экосистем на незаселенных ранее
Первичной сукцессией называется процесс развития и смены экосистем на незаселенных ранее

Первичная сукцессия
Первичная сукцессия

Вторичная сукцессия — это восстановление экосистемы, когда-то уже существовавшей на данной
Вторичная сукцессия — это восстановление экосистемы, когда-то уже существовавшей на данной

Вторичная сукцессия
Вторичная сукцессия

Вторичная сукцессия сибирского темно-хвойного леса
(пихтово-кедровой тайги) после опустошительного лесного пожара
Вторичная сукцессия сибирского темно-хвойного леса (пихтово-кедровой тайги) после опустошительного лесного пожара

Климаксовоя экосистема
Сукцессия завершается стадией, когда все виды экосистемы, размножаясь, сохраняют относительно
Климаксовоя экосистема
Сукцессия завершается стадией, когда все виды экосистемы, размножаясь, сохраняют относительно

Специфические закономерности сукцессии
Американский эколог Ф. Клементс, наиболее полно разработавший учение о
Специфические закономерности сукцессии
Американский эколог Ф. Клементс, наиболее полно разработавший учение о

Специфические закономерности сукцессии
Наряду с теорией моноклимакса существует точка зрения, в соответствии
Специфические закономерности сукцессии
Наряду с теорией моноклимакса существует точка зрения, в соответствии

Антропогенные сукцессии
Наряду с природными факторами, причинами динамики экосистем выступает человек. К
Антропогенные сукцессии
Наряду с природными факторами, причинами динамики экосистем выступает человек. К

Общие закономерности сукцессионного процесса
1. На начальных стадиях видовое разнообразие незначительно, продуктивность
Общие закономерности сукцессионного процесса
1. На начальных стадиях видовое разнообразие незначительно, продуктивность

Общие закономерности сукцессионного процесса
5. Скорость сукцессионного процесса в большей мере зависит
Общие закономерности сукцессионного процесса
5. Скорость сукцессионного процесса в большей мере зависит
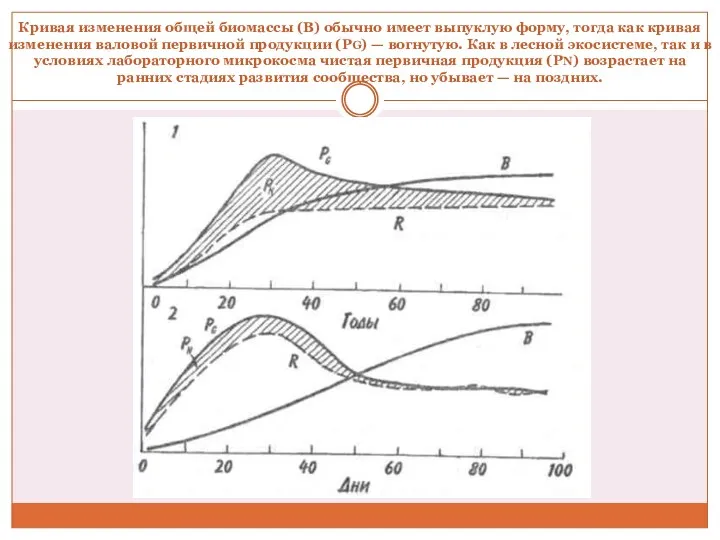
Кривая изменения общей биомассы (B) обычно имеет выпуклую форму, тогда как
Кривая изменения общей биомассы (B) обычно имеет выпуклую форму, тогда как

Положение сообществ различных типов в классификации, основанной на метаболизме (по: Одум,
Положение сообществ различных типов в классификации, основанной на метаболизме (по: Одум,

Изменение признаков экосистемы в ходе сукцессии
Изменение признаков экосистемы в ходе сукцессии

Законы развития экосистем
Закон последовательности прохождения фаз развития: фазы развития природной системы
Законы развития экосистем
Закон последовательности прохождения фаз развития: фазы развития природной системы
 Современные экологические проблемы и способы их решения
Современные экологические проблемы и способы их решения Экология, как наука. Задания на лето
Экология, как наука. Задания на лето Water Pollution
Water Pollution Захоронение твердых бытовых и промышленных отходов на полигонах. (Лекция 5)
Захоронение твердых бытовых и промышленных отходов на полигонах. (Лекция 5) Global warming
Global warming Экологические последствия урагана Катрина
Экологические последствия урагана Катрина Глобальные экологические прблемы
Глобальные экологические прблемы Отчет о проведении районной экологической акции Птицы Кубани, операция День птиц
Отчет о проведении районной экологической акции Птицы Кубани, операция День птиц Поток энергии и цепи питания
Поток энергии и цепи питания Загрязнение литосферы. Причины
Загрязнение литосферы. Причины Промышленные предприятия и их воздействие на природу
Промышленные предприятия и их воздействие на природу Водоёмы Саратовской области и их охрана
Водоёмы Саратовской области и их охрана Национальные парки и заповедники России
Национальные парки и заповедники России Круговорот веществ в природе
Круговорот веществ в природе Общие законы действия факторов среды на организмы
Общие законы действия факторов среды на организмы Green BIM технологии в архитектурном проектировании
Green BIM технологии в архитектурном проектировании Загрязнение и изменения литосферы
Загрязнение и изменения литосферы What is ecology? What is environment?
What is ecology? What is environment? Қоршаған орта үшін мүмкін болатын қолайсыз эффектілердің қауіптілігін бағалау
Қоршаған орта үшін мүмкін болатын қолайсыз эффектілердің қауіптілігін бағалау Презентация Редкие растения и их охрана
Презентация Редкие растения и их охрана Экология как наука
Экология как наука Социальный проект Экологическая тропа. Путешествие по просторам родного края
Социальный проект Экологическая тропа. Путешествие по просторам родного края Енергозбереження. Проблеми екології
Енергозбереження. Проблеми екології Топ-10 радиациялық заттардың құрбаны болған жерлер
Топ-10 радиациялық заттардың құрбаны болған жерлер Отношение христианина к природе
Отношение христианина к природе Живая планета
Живая планета Экологическое нормирование
Экологическое нормирование Круговорот веществ в биосфере
Круговорот веществ в биосфере