- Особливості біологічних об’єктів як термодинамічних структур

Содержание
- 2. Однією з умов нормального функціонування окремої клітини і цілого організму є підтримання постійності їх параметрів (концентрації
- 3. Термодинаміку розділяють на два розділи: класичну (рівноважну) і термодинаміку необоротних процесів (нерівноважну). Рівноважна термодинаміка вивчає в
- 4. В термодинаміці ізольованих процесів основним поняттям є стаціонарний стан системи. В стаціонарному стані параметри не змінюються
- 5. Така ситуація, а також градієнт концентрацій сприяють просочуванню іонів Na+ всередину клітини, незважаючи на низьку проникливість
- 6. Перше начало термодинаміки в хімії і біології Перше начало термодинаміки δQ=dU+δA (1) де Q – кількість
- 7. при V=const δQ=dU (4) при p=const δQ=dU+pdV=d(U+H)=dH, (5) де Н=U+pV- ентальпія, функція стану, що визначає кількість
- 8. В біологічних системах процеси здійснюються в основному при постійному тиску, тому тепловий ефект біохімічних реакцій дорівнює
- 9. Друге начало термодинаміки для відкритих систем Згідно другого начала термодинаміки, зміна ентропії (dS) є більшою або
- 10. Це критерій Клаузіуса: ізольована система прагне досягнути кінцевого рівноважного стану з максимальною ентропією. У процесі функціонування
- 11. Запишемо загалбну зміну ентропії в такій системі (3) Для оборотних процесів diS=0, для необоротних diS>0. Якщо
- 12. В загальному випадку може спостерігатись один із трьох процесів: Перший випадок >0 відповідає патологічному стану організму,
- 13. Другий випадок Третій випадок =0 відповідає встановленню стаціонарного стану в системі. В цілому в системі Земля-Сонце
- 15. Скачать презентацию

Однією з умов нормального функціонування окремої клітини і цілого організму є
Однією з умов нормального функціонування окремої клітини і цілого організму є

Термодинаміку розділяють на два розділи: класичну (рівноважну) і термодинаміку необоротних процесів
Термодинаміку розділяють на два розділи: класичну (рівноважну) і термодинаміку необоротних процесів

В термодинаміці ізольованих процесів основним поняттям є стаціонарний стан системи. В
В термодинаміці ізольованих процесів основним поняттям є стаціонарний стан системи. В

Така ситуація, а також градієнт концентрацій сприяють просочуванню іонів Na+ всередину
Така ситуація, а також градієнт концентрацій сприяють просочуванню іонів Na+ всередину

Перше начало термодинаміки в хімії і біології
Перше начало термодинаміки
δQ=dU+δA (1)
де
Перше начало термодинаміки в хімії і біології
Перше начало термодинаміки
δQ=dU+δA (1)
де

при V=const
δQ=dU (4)
при p=const
δQ=dU+pdV=d(U+H)=dH, (5)
де Н=U+pV- ентальпія, функція стану,
при V=const
δQ=dU (4)
при p=const
δQ=dU+pdV=d(U+H)=dH, (5)
де Н=U+pV- ентальпія, функція стану,

В біологічних системах процеси здійснюються в основному при постійному тиску, тому
В біологічних системах процеси здійснюються в основному при постійному тиску, тому

Друге начало термодинаміки
для відкритих систем
Згідно другого начала термодинаміки, зміна ентропії
Друге начало термодинаміки
для відкритих систем
Згідно другого начала термодинаміки, зміна ентропії

Це критерій Клаузіуса:
ізольована система прагне досягнути кінцевого рівноважного стану з максимальною
Це критерій Клаузіуса:
ізольована система прагне досягнути кінцевого рівноважного стану з максимальною

Запишемо загалбну зміну ентропії в такій системі
(3)
Для оборотних процесів diS=0, для
Запишемо загалбну зміну ентропії в такій системі
(3)
Для оборотних процесів diS=0, для

В загальному випадку може спостерігатись один із трьох процесів:
Перший випадок >0
В загальному випадку може спостерігатись один із трьох процесів:
Перший випадок >0

Другий випадок <0 відповідає підвищенню рівня організації організму, наприклад при
Другий випадок <0 відповідає підвищенню рівня організації організму, наприклад при
 Цепные передачи
Цепные передачи Хвильова оптика
Хвильова оптика Атом водорода в квантовой физике. (Лекция 8)
Атом водорода в квантовой физике. (Лекция 8) Инфракрасная Фурье спектроскопия
Инфракрасная Фурье спектроскопия Влажность воздуха
Влажность воздуха Бойове застосування КЗА 86Ж6. Система електроживлення, вентиляції, кондиціювання. Розподільний пристрій 64Т6. (Тема 9.3)
Бойове застосування КЗА 86Ж6. Система електроживлення, вентиляції, кондиціювання. Розподільний пристрій 64Т6. (Тема 9.3) Решение задач на применение уравнения состояния идеального газа и газовых законов
Решение задач на применение уравнения состояния идеального газа и газовых законов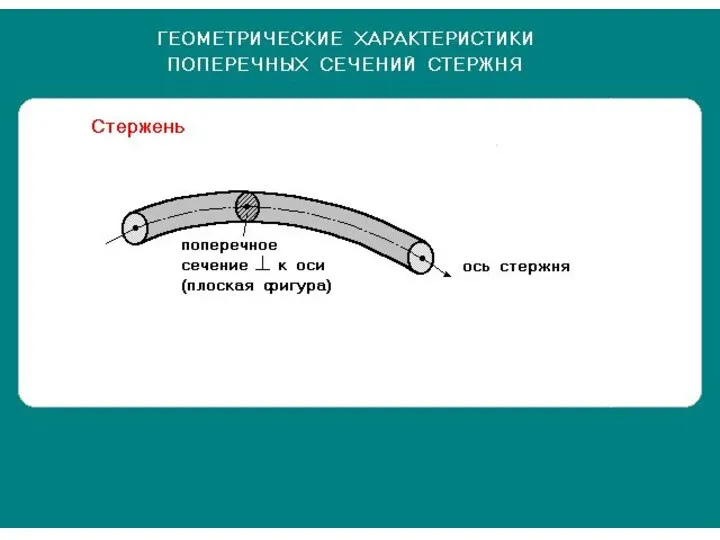 Геометрические характеристики поперечных сечений стержня
Геометрические характеристики поперечных сечений стержня Зеркала
Зеркала VALVEDIM – программа для расчета пропускной способности и подбора регулирующих клапанов
VALVEDIM – программа для расчета пропускной способности и подбора регулирующих клапанов Сила трения. Трение в природе и технике
Сила трения. Трение в природе и технике Вакуумные винтовые сухие безмасляные насосы LGB
Вакуумные винтовые сухие безмасляные насосы LGB Решение задач по темам Архимедова сила, Условия плавания тел. 7 класс
Решение задач по темам Архимедова сила, Условия плавания тел. 7 класс Повышение качества образования по предмету физика через систему использования интегрированных уроков в условиях модернизации образовательной системы РФ
Повышение качества образования по предмету физика через систему использования интегрированных уроков в условиях модернизации образовательной системы РФ Презентация по теме Спектральный анализ и его применение 9 класс
Презентация по теме Спектральный анализ и его применение 9 класс Равновесие при наличии трения
Равновесие при наличии трения Физика Солнца
Физика Солнца Тест как средство контроля знаний и умений учащихся по физике
Тест как средство контроля знаний и умений учащихся по физике Домашняя лабораторная работа. Плотность куска мыла
Домашняя лабораторная работа. Плотность куска мыла Структура пограничного слоя при продольном обтекании сотовой поверхности
Структура пограничного слоя при продольном обтекании сотовой поверхности Электронные свойства низкоразмерных электронных систем. Принцип размерного квантования
Электронные свойства низкоразмерных электронных систем. Принцип размерного квантования Жидкостная хроматография Waters Breeze
Жидкостная хроматография Waters Breeze Механические колебания
Механические колебания Взаимное притяжение и отталкивание молекул
Взаимное притяжение и отталкивание молекул Действие магнитного поля на проводник с током и движущиеся заряды
Действие магнитного поля на проводник с током и движущиеся заряды Методы исследования. Вещества
Методы исследования. Вещества Итоговый урок, физика 8 класс
Итоговый урок, физика 8 класс Обозначение физической величины
Обозначение физической величины