- Система комплемента как часть иммунной системы

Содержание
- 2. foot and mouth virus Influenza virus E. coli Anthrax Candida albicans
- 3. Комплемент Иммунная система Врождённый иммунитет Приобретённый иммунитет
- 4. Immune system Effector arm Regulatory arm Humoral response Cellular response Antibodies Lectin C Pathway Alternative C
- 5. Система комплемента Впервые была обнаружена в 1889 г. как составляющая нормальной сыворотки, чувствительная к высоким температурам
- 6. Система комплемента Функции: Лизис: клеток, бактерий (грам-негативных), паразитов, вирусов Опсонизация комплексов антиген-антитело с помощью продуктов расщепления
- 7. Что делает комплемент?
- 8. Компоненты комплемента Протеины и гликопротеины, составляющие систему комплемента, синтезируются в основном гепатоцитами печени, а также кровяными
- 9. Растворимые белки комплемента
- 10. Три способа защиты от инфекции:
- 11. Ранние события всех трёх путей активации это ряд реакций расщепления, завершает которые образование С3 конвертазы, которая
- 12. Classical pathway C1 C4, C2 Activation C3 convertase C4b,2a C4b,2a,3b C5 convertase C5 C5b,6,7,8,9n (MAC) C6,C7,C8,nC9
- 13. Mannan-binding lectin
- 14. Alternative pathway C3(H2O) B,D C3(H2O)Bb C3 C3b B, D C3b,Bb C3b,Bb,3b C5 C5b,6,7,8,9n (MAC) C6,C7,C8,nC9 I.
- 15. II.
- 16. DAF/CD55 MCP/CD46, CR1
- 17. Factor H
- 18. Тиоэфирная связь в белках: C3b и C4b C3, C4 C3a, C4a C3b, C4b
- 19. Classical pathway Lectin pathway C1 MBL/ficolin MASP C4, C2 C4, C2 Activation C3 convertase C4b,2a C4b,2a
- 20. I. Фагоцитоз: рецепторы комплемента
- 21. CR1-C3b C5aR-C5a Фагоцитоз I. Фагоцитоз: рецепторы комплемента
- 22. II.Воспаление Маленькие фрагменты комплемента – C3a, C4a, C5a - активируют специфические рецепторы и вызывают локальные воспаления.
- 23. III. МАК - Конечные белки комплемента
- 24. Образование комплекса C5b-9 C5b C6 C7 C6 C5b C5b C5b C6 C6 C7 C7 C7 C8
- 25. MAC
- 26. C9 – «палач» системы комплемента Гидрофильный белок Гидрофобный белок
- 27. PolyC9 21 nm 15 nm Transmembrane channel Membrane lipids rearrangement Loss of membrane integrity Комплексы атакующие
- 28. Necrosis MACs Sodium influx Penetration of water Cell death
- 29. Mitochondria are damaged within 5 min of MAC attack Papadimitriou et al. J. Immunol. 147: 212,
- 30. Complement in pathogenesis Alzheimer’s disease (Rogers et al., 1992) Allotransplantation (Pruitt and Bollinger, 1991) Asthma (Regal
- 31. Complement in pathogenesis
- 32. Регуляция системы комплемента
- 33. Soluble complement regulatory proteins: Factor H, Factor I, C4bp, Properdin, S protein (Vitronectin), Clusterin (SP40,40), C1INH
- 34. Complement resistance of tumor cells: basal and induced mechanisms Fishelson et al., 1999, Mol Imm
- 35. Эффект комплемента на опухолевые клетки неоднократно демонстрировался, тем не менее однозначных доказательств анти-раковых функций показано не
- 36. Известно, что клетки, обладающие ядрами, отличаются друг от друга в чувствительности к комплементу, эти различия принято
- 37. mCRPs (membrane Complement Regulatory Proteins) Устойчивость опухолевых клеток к системе комплемента Complement resistance of tumor cells:
- 38. Устойчивость опухолевых клеток к системе комплемента Complement resistance of tumor cells: basal and induced mechanisms Fishelson
- 39. Устойчивость опухолевых клеток к системе комплемента Complement resistance of tumor cells: basal and induced mechanisms Fishelson
- 40. CD59 CD59 binds to C8 and C9 and prevents further build up of the Membrane Attack
- 41. mCRPs находящиеся на поверхности нормальных и опухолевых клеток и уровень их экспрессии даже в одних и
- 42. Expression of membrane-bound complement regulators on K562 cells, PMN and erythrocytes. Cells were treated first with
- 43. Важность mCRPs как механизма устойчивости к комплементу была также показана с помощью экспериментов с блокированием этих
- 44. Effect of neutralization of membrane complement regulatory proteins on complement-mediated cell lysis. Cells were treated with
- 45. Основные механизмы устойчивости к комплементу: I. экспрессия растворимых белков-регуляторов
- 46. Индуцируемые механизмы устойчивости к комплементу Tumor cell DAF MCP CD59 I II III B Ca2+ MAC
- 47. Интересно, что одним из наиболее потенциальных подобных агентов является сам МАК. Для того чтобы лизировать клетки
- 48. Также было показано, что клетки К562 могут быть защищены от лизиса комплементом с помощью обработки с
- 49. Обработка клеток с помощью суб-литического комплемента приводит в действие клеточный механизм, который убирает МАК с поверхности
- 50. Mortalin/GRP75 promotes release of membrane vesicles from immune attacked cells and protection from complement-mediated lysis D.
- 51. Белки, участвующие в везикуляции Method: * NHS-normal Human Sera HI-NHS-Heat Inactivated Normal Human Sera
- 52. NHS HI-NHS Mortalin/GRP75 promotes release of membranevesicles from immune attacked cells and protection from complement-mediated lysis.
- 53. Mortalin Mitochondria Unfolding of proteins and importing them into the mitochondria Cytosol Intracellular trafficking P53 inactivation
- 54. Mortalin is secreted from cells after sub-lytic complement attack K562 cells treated with sub-lytic complement (anti-K562
- 55. Mortalin and C9 are associated with membrane vesicles K562 treated with sub-lytic complement Sedimentation at 5000Xg
- 56. HI-NHS NHS HI-NHS NHS Anti-Mortalin Ab Anti-C9 Ab Anti-Mortalin Ab Anti-C9 Ab Results: HI-NHS NHS HI-NHS
- 57. NHS + anti-mortalin Mortalin is translocated to the cell cortex after sub-lytic complement attack NHS +
- 58. Addition of anti-Mortalin Ab to the experiments Anti-Mortalin Ab lowers cells’ protection against complement K562 cells
- 59. Results:
- 60. Increased sensitivity of early apoptotic cells to complement-mediated lysis Gitit Attali, Dana Gancz and Zvi Fishelson,
- 61. Интеракция белков комплемента с апоптотическими клетками была продемонстрирована, однако эта тема мало изучена. Апоптотические клетки могут
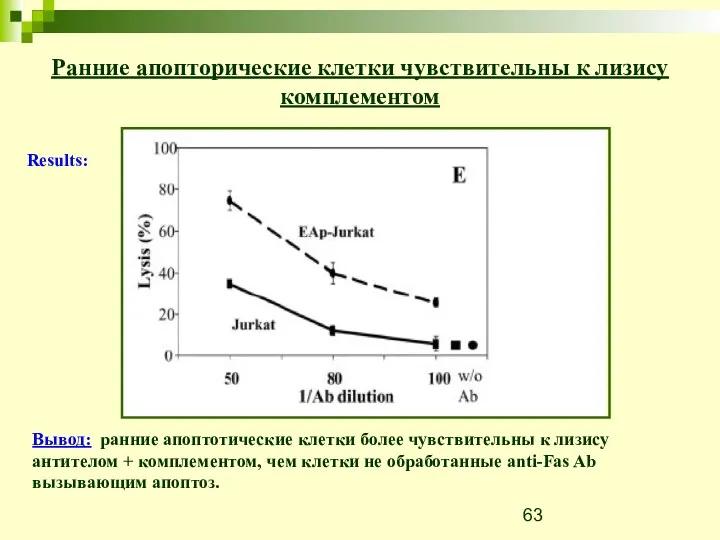
- 62. Ранние апопторические клетки чувствительны к лизису комплементом Цель: проверить интеракцию между ранними апоптотическими клетками и системой
- 63. Вывод: ранние апоптотические клетки более чувствительны к лизису антителом + комплементом, чем клетки не обработанные anti-Fas
- 64. Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases) Цель: проверить участвуют ли каспазы к
- 65. Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases) Вывод: 10 uM частично уменьшили гибель
- 66. Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом Анти-апоптотические эффекты Bcl-2 хорошо известны. Этот белок
- 67. Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом Results: Вывод: Наблюдается прямая связь между over-expression
- 69. Скачать презентацию
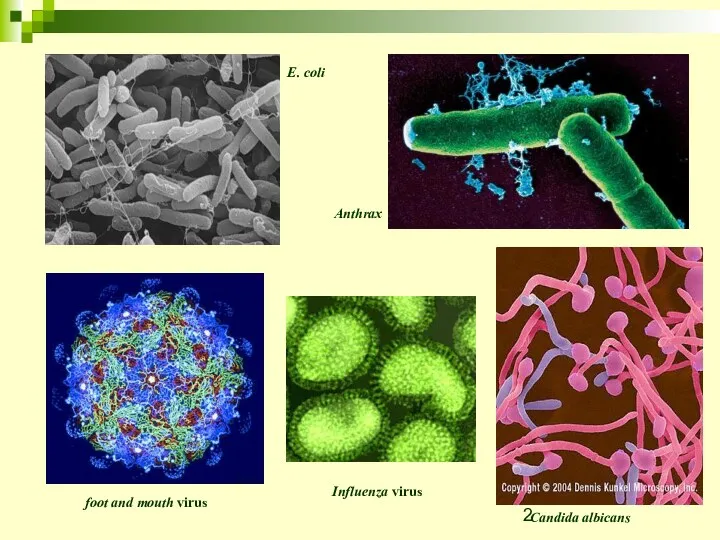
foot and mouth virus
Influenza virus
E. coli
Anthrax
Candida albicans
foot and mouth virus
Influenza virus
E. coli
Anthrax
Candida albicans
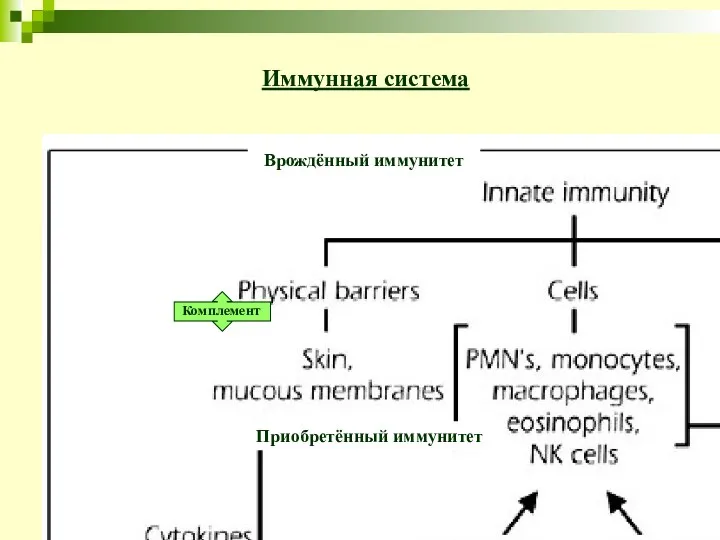
Комплемент
Иммунная система
Врождённый иммунитет
Приобретённый иммунитет
Комплемент
Иммунная система
Врождённый иммунитет
Приобретённый иммунитет
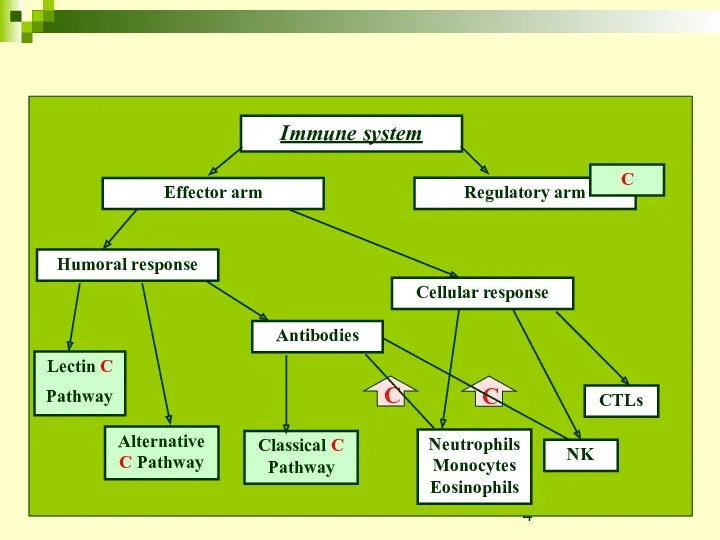
Immune system
Effector arm
Regulatory arm
Humoral response
Cellular response
Antibodies
Lectin C
Pathway
Alternative C Pathway
Classical C Pathway
CTLs
NK
Neutrophils
Monocytes
Eosinophils
C
C
C
Immune system
Effector arm
Regulatory arm
Humoral response
Cellular response
Antibodies
Lectin C
Pathway
Alternative C Pathway
Classical C Pathway
CTLs
NK
Neutrophils
Monocytes
Eosinophils
C
C
C

Система комплемента
Впервые была обнаружена в 1889 г. как составляющая нормальной сыворотки,
Система комплемента
Впервые была обнаружена в 1889 г. как составляющая нормальной сыворотки,

Система комплемента
Функции:
Лизис: клеток, бактерий (грам-негативных), паразитов, вирусов
Опсонизация комплексов антиген-антитело с
Система комплемента
Функции:
Лизис: клеток, бактерий (грам-негативных), паразитов, вирусов
Опсонизация комплексов антиген-антитело с

Что делает комплемент?
Что делает комплемент?

Компоненты комплемента
Протеины и гликопротеины, составляющие систему комплемента, синтезируются в основном гепатоцитами
Компоненты комплемента
Протеины и гликопротеины, составляющие систему комплемента, синтезируются в основном гепатоцитами
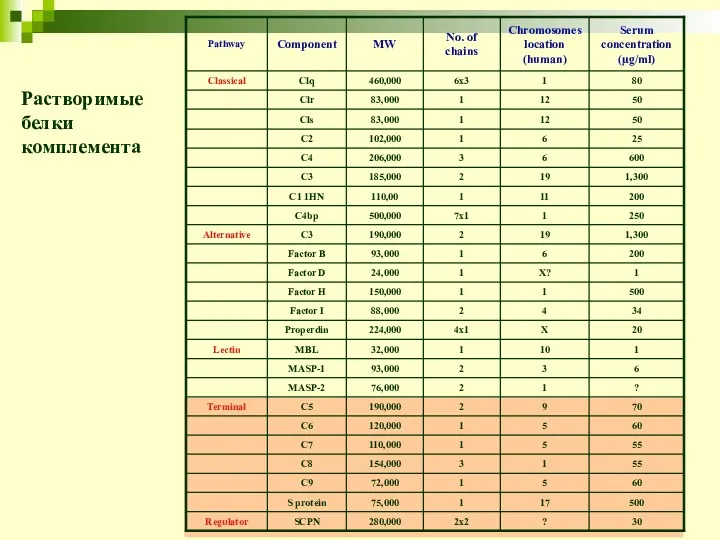
Растворимые белки комплемента
Растворимые белки комплемента
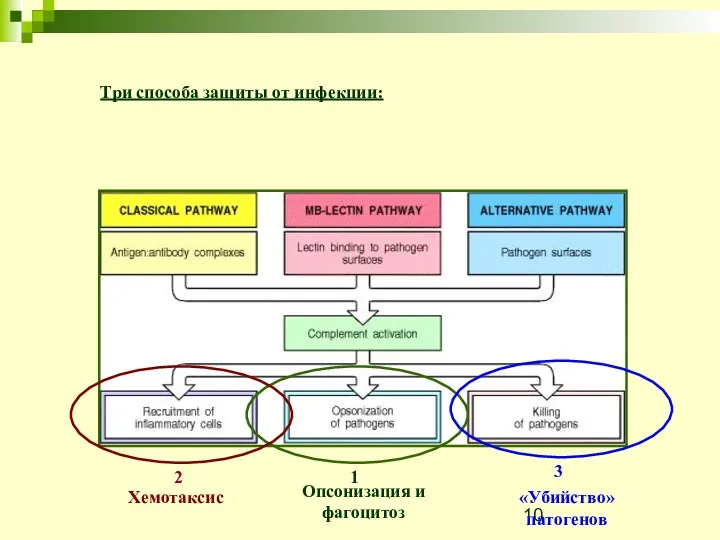
Три способа защиты от инфекции:
Три способа защиты от инфекции:
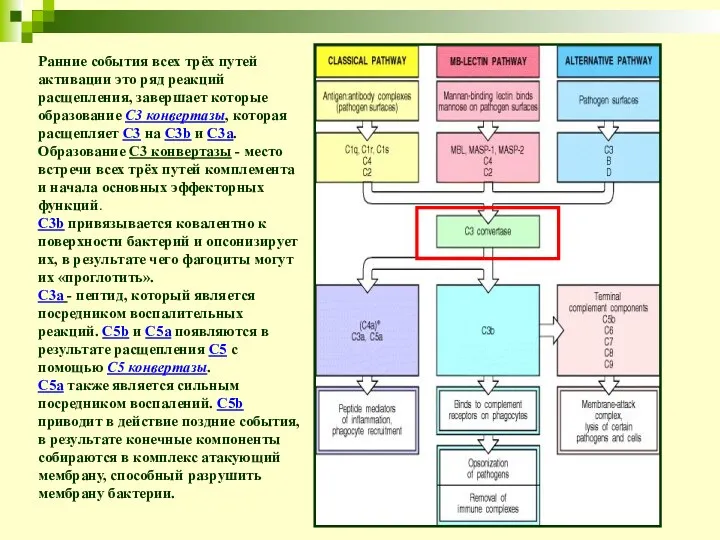
Ранние события всех трёх путей активации это ряд реакций расщепления, завершает
Ранние события всех трёх путей активации это ряд реакций расщепления, завершает
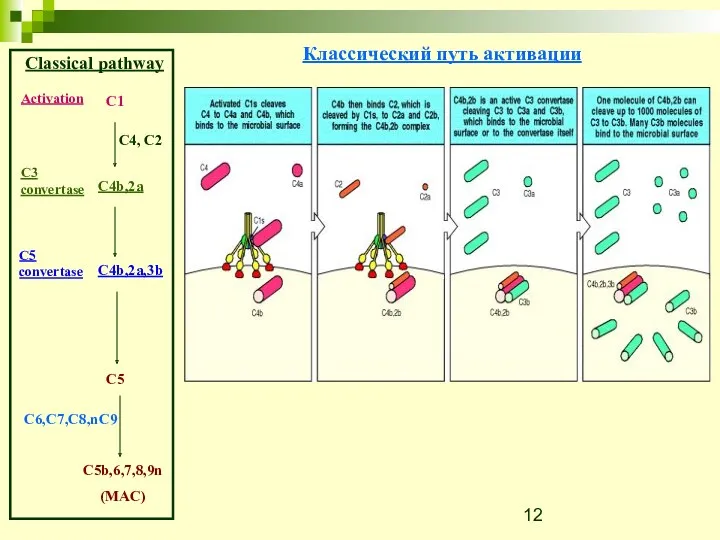
Classical pathway
C1
C4, C2
Activation
C3 convertase
C4b,2a
C4b,2a,3b
C5 convertase
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
C1 комплекс
Classical pathway
C1
C4, C2
Activation
C3 convertase
C4b,2a
C4b,2a,3b
C5 convertase
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
C1 комплекс
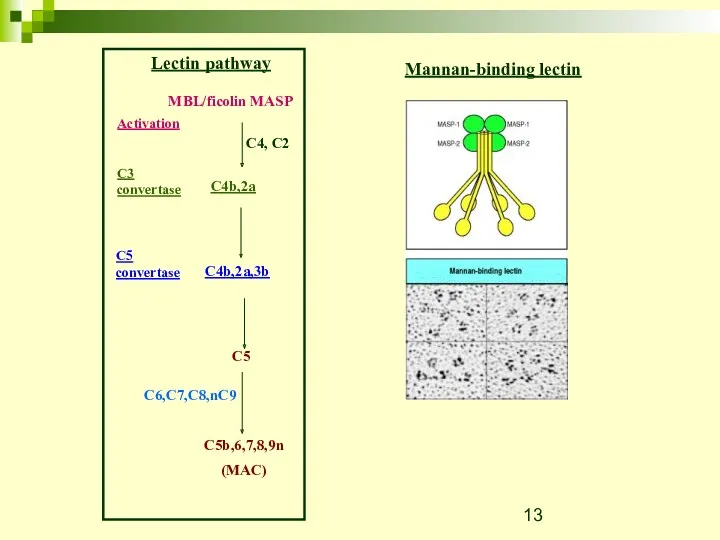
Mannan-binding lectin
Mannan-binding lectin
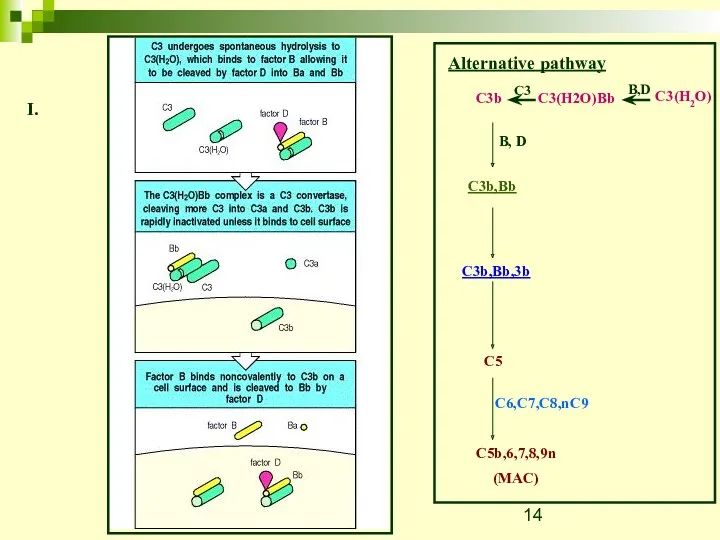
Alternative pathway
C3(H2O)
B,D
C3(H2O)Bb
C3
C3b
B, D
C3b,Bb
C3b,Bb,3b
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
I.
Alternative pathway
C3(H2O)
B,D
C3(H2O)Bb
C3
C3b
B, D
C3b,Bb
C3b,Bb,3b
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
I.
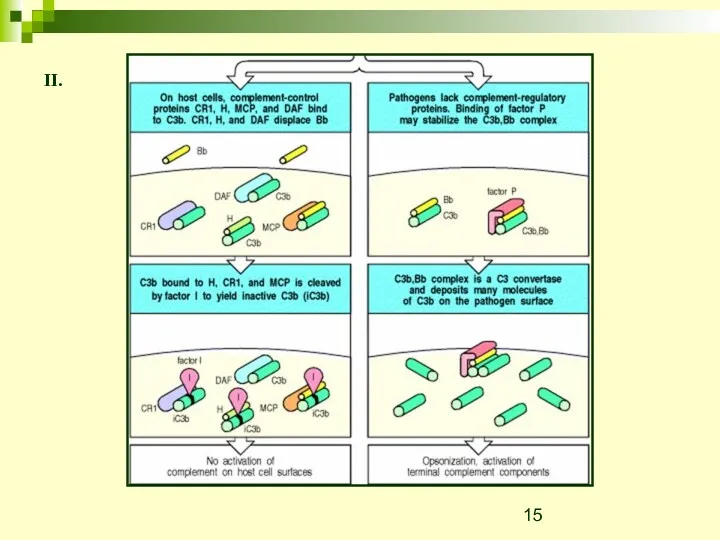
II.
II.
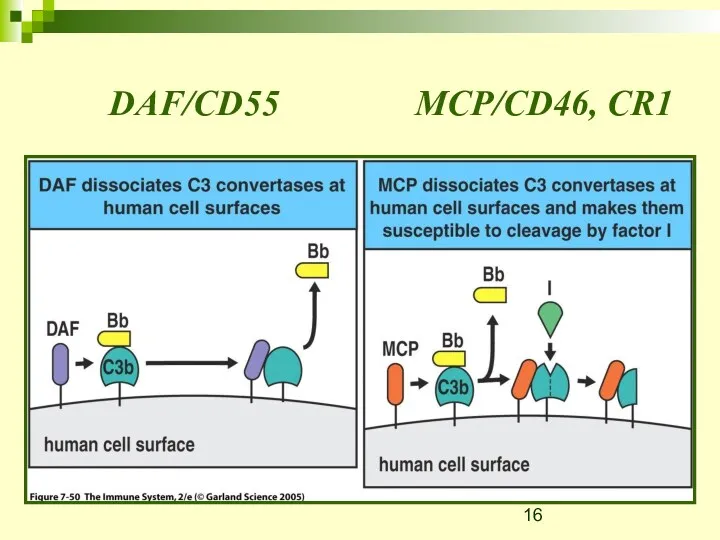
DAF/CD55
MCP/CD46, CR1
DAF/CD55
MCP/CD46, CR1
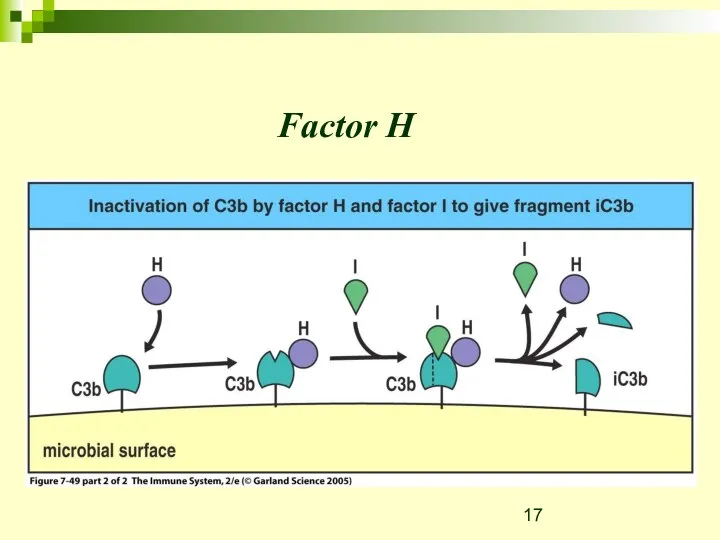
Factor H
Factor H
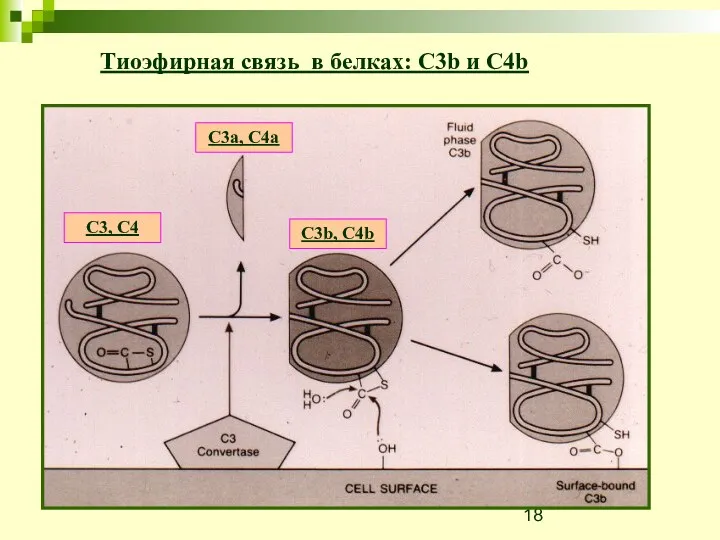
Тиоэфирная связь в белках: C3b и C4b
C3, C4
C3a, C4a
C3b, C4b
Тиоэфирная связь в белках: C3b и C4b
C3, C4
C3a, C4a
C3b, C4b
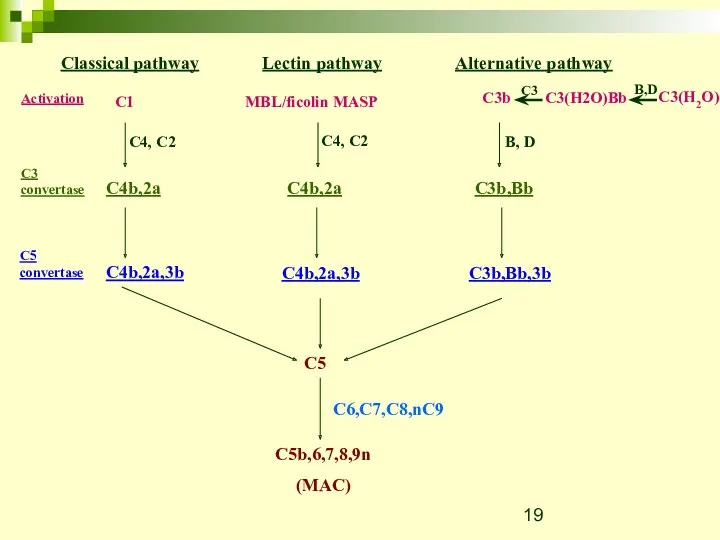
Classical pathway
Lectin pathway
C1
MBL/ficolin MASP
C4, C2
C4, C2
Activation
C3 convertase
C4b,2a
C4b,2a
C4b,2a,3b
C4b,2a,3b
C5 convertase
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
Classical pathway
Lectin pathway
C1
MBL/ficolin MASP
C4, C2
C4, C2
Activation
C3 convertase
C4b,2a
C4b,2a
C4b,2a,3b
C4b,2a,3b
C5 convertase
C5
C5b,6,7,8,9n
(MAC)
C6,C7,C8,nC9
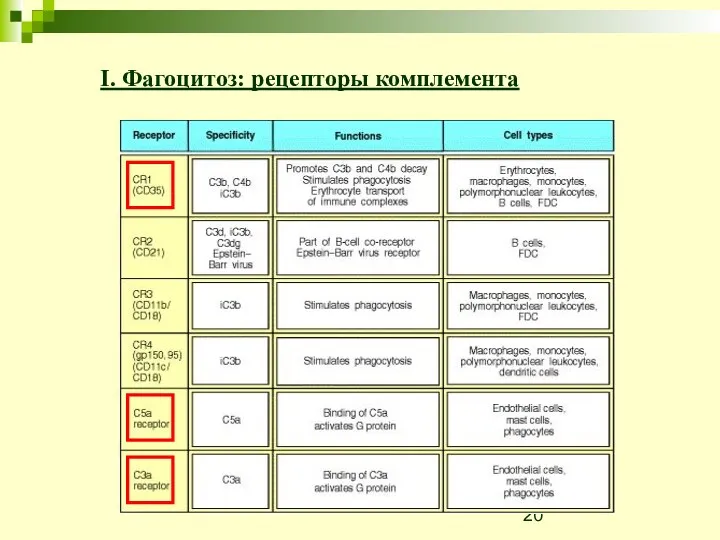
I. Фагоцитоз: рецепторы комплемента
I. Фагоцитоз: рецепторы комплемента
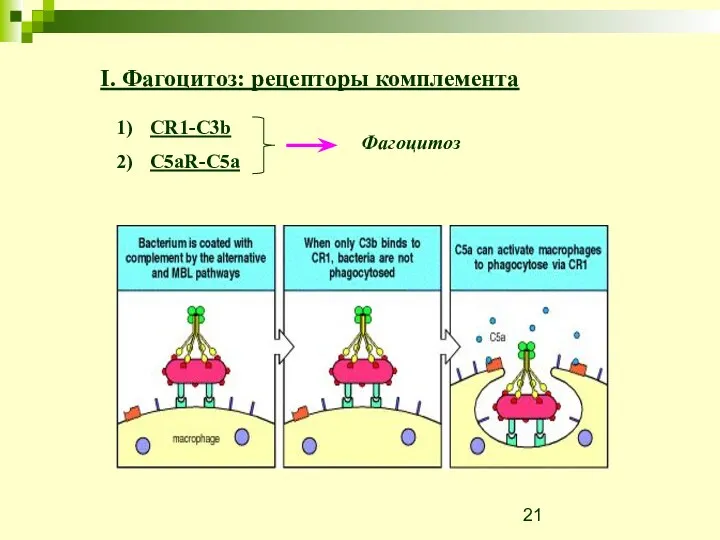
CR1-C3b
C5aR-C5a
Фагоцитоз
I. Фагоцитоз: рецепторы комплемента
CR1-C3b
C5aR-C5a
Фагоцитоз
I. Фагоцитоз: рецепторы комплемента
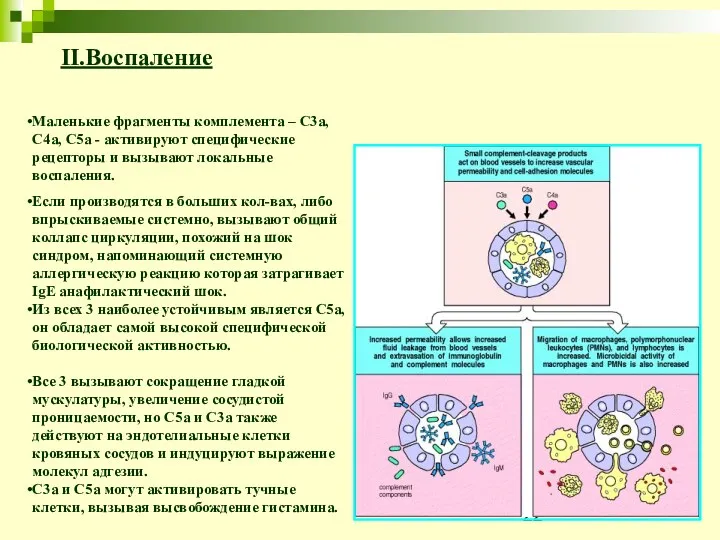
II.Воспаление
Маленькие фрагменты комплемента – C3a, C4a, C5a - активируют специфические рецепторы
II.Воспаление
Маленькие фрагменты комплемента – C3a, C4a, C5a - активируют специфические рецепторы

III. МАК - Конечные белки комплемента
III. МАК - Конечные белки комплемента
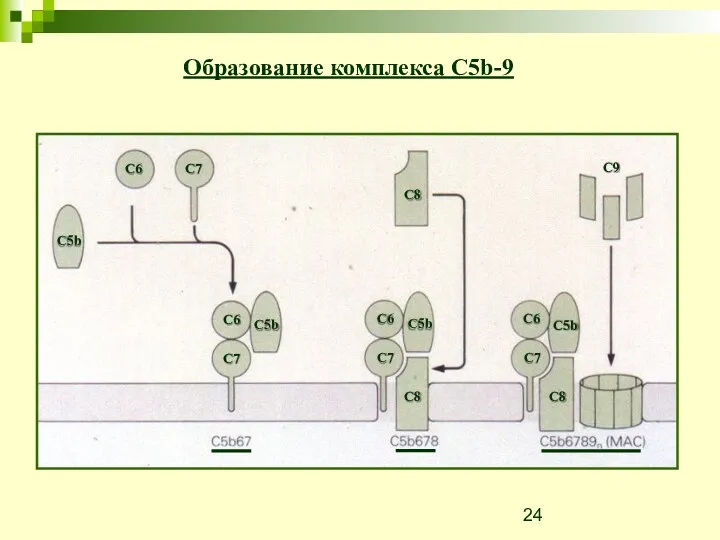
Образование комплекса C5b-9
C5b
C6
C7
C6
C5b
C5b
C5b
C6
C6
C7
C7
C7
C8
C8
C8
C9
Образование комплекса C5b-9
C5b
C6
C7
C6
C5b
C5b
C5b
C6
C6
C7
C7
C7
C8
C8
C8
C9
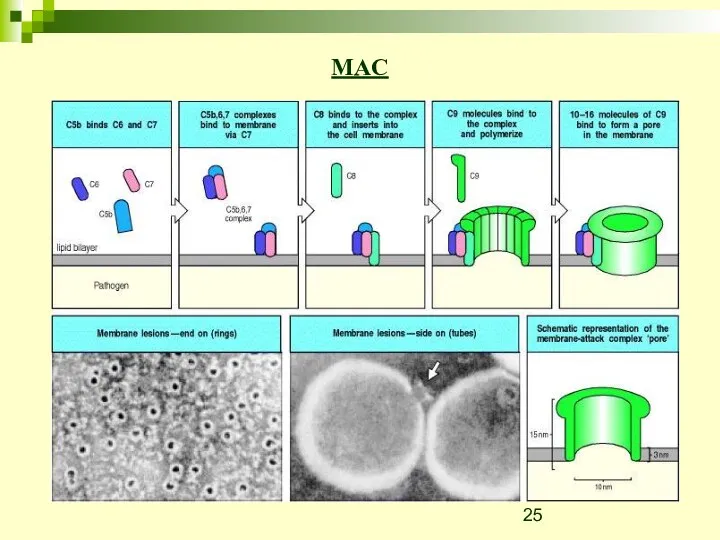
MAC
MAC
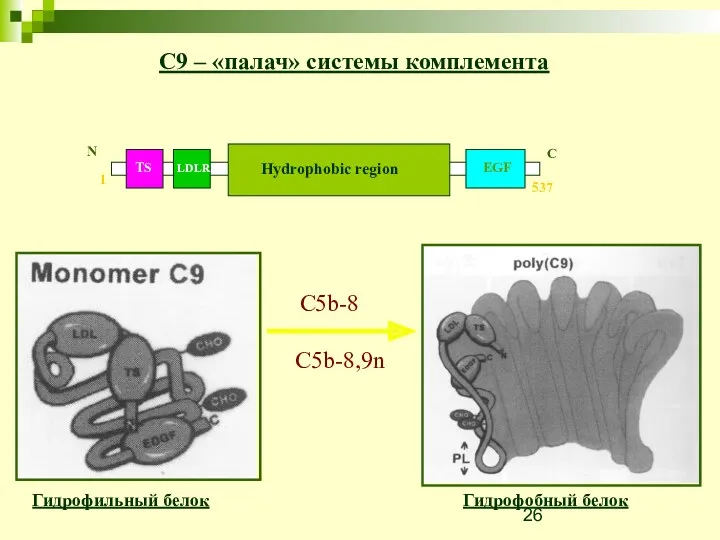
C9 – «палач» системы комплемента
Гидрофильный белок
Гидрофобный белок
C9 – «палач» системы комплемента
Гидрофильный белок
Гидрофобный белок
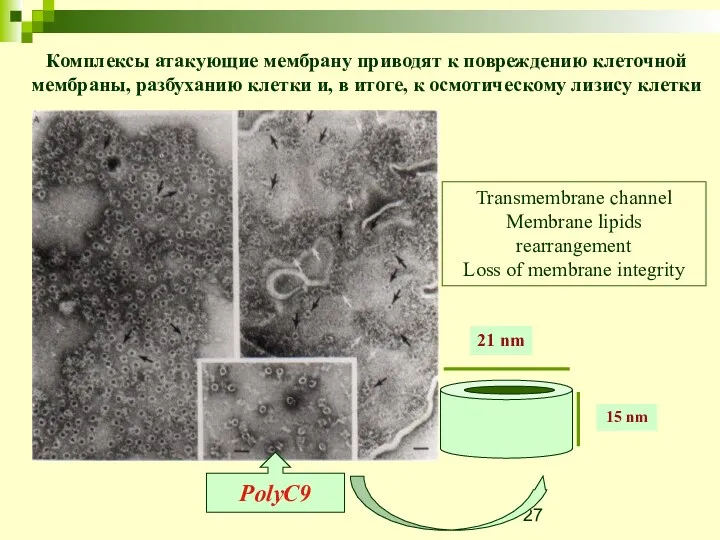
PolyC9
21 nm
15 nm
Transmembrane channel
Membrane lipids rearrangement
Loss of membrane integrity
Комплексы атакующие мембрану
PolyC9
21 nm
15 nm
Transmembrane channel
Membrane lipids rearrangement
Loss of membrane integrity
Комплексы атакующие мембрану

Necrosis
MACs
Sodium influx
Penetration of water
Cell death
Necrosis
MACs
Sodium influx
Penetration of water
Cell death
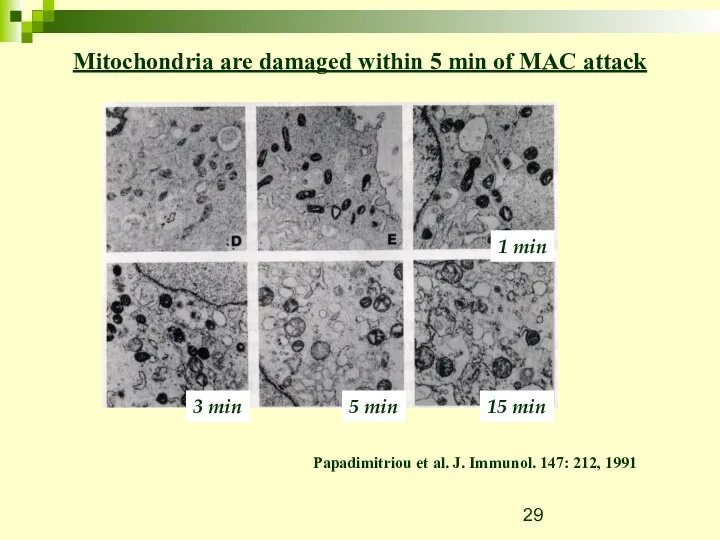
Mitochondria are damaged within 5 min of MAC attack
Papadimitriou et al.
Mitochondria are damaged within 5 min of MAC attack
Papadimitriou et al.

Complement in pathogenesis
Alzheimer’s disease (Rogers et al., 1992)
Allotransplantation (Pruitt and Bollinger,
Complement in pathogenesis
Alzheimer’s disease (Rogers et al., 1992)
Allotransplantation (Pruitt and Bollinger,

Complement in pathogenesis
Complement in pathogenesis
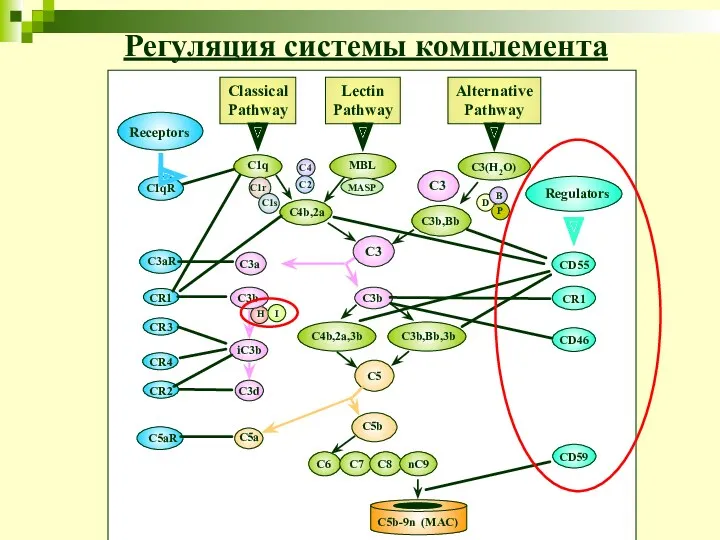
Регуляция системы комплемента
Регуляция системы комплемента

Soluble complement regulatory proteins:
Factor H, Factor I, C4bp, Properdin,
S protein (Vitronectin),
Soluble complement regulatory proteins:
Factor H, Factor I, C4bp, Properdin,
S protein (Vitronectin),

Complement resistance of tumor cells:
basal and induced mechanisms
Fishelson et al.,
Complement resistance of tumor cells:
basal and induced mechanisms
Fishelson et al.,

Эффект комплемента на опухолевые клетки неоднократно демонстрировался, тем не менее однозначных
Эффект комплемента на опухолевые клетки неоднократно демонстрировался, тем не менее однозначных

Известно, что клетки, обладающие ядрами, отличаются друг от друга в чувствительности
Известно, что клетки, обладающие ядрами, отличаются друг от друга в чувствительности
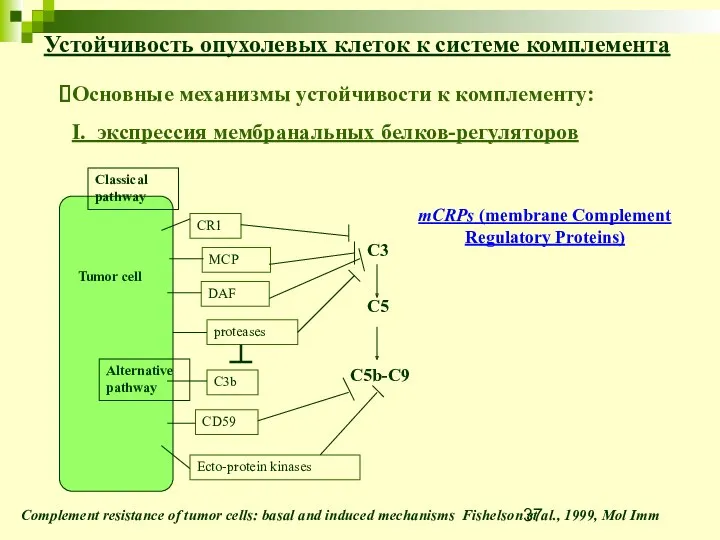
mCRPs (membrane Complement Regulatory Proteins)
Устойчивость опухолевых клеток к системе комплемента
Complement resistance
Устойчивость опухолевых клеток к системе комплемента
Complement resistance
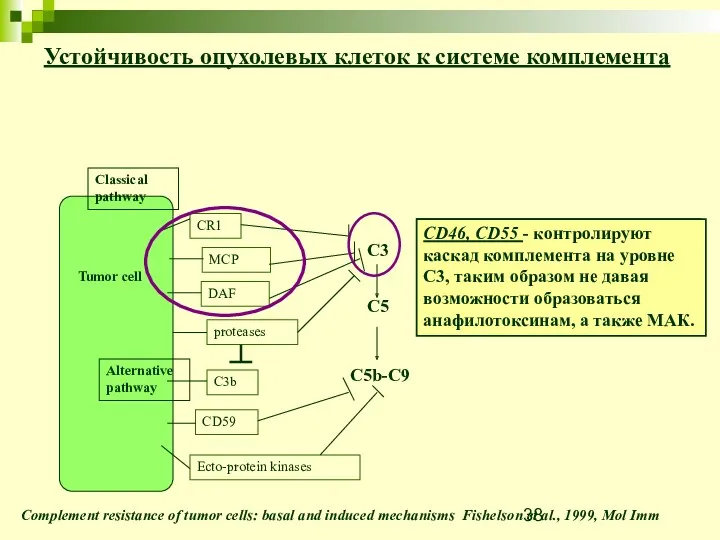
Устойчивость опухолевых клеток к системе комплемента
Complement resistance of tumor cells: basal
Устойчивость опухолевых клеток к системе комплемента
Complement resistance of tumor cells: basal
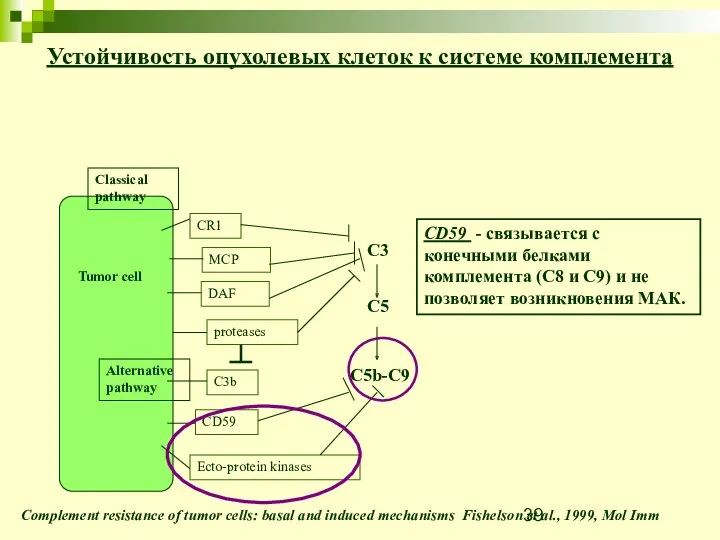
Устойчивость опухолевых клеток к системе комплемента
Complement resistance of tumor cells: basal
Устойчивость опухолевых клеток к системе комплемента
Complement resistance of tumor cells: basal

CD59
CD59 binds to C8 and C9 and prevents further
build up
CD59
CD59 binds to C8 and C9 and prevents further
build up

mCRPs находящиеся на поверхности нормальных и опухолевых клеток и уровень
mCRPs находящиеся на поверхности нормальных и опухолевых клеток и уровень
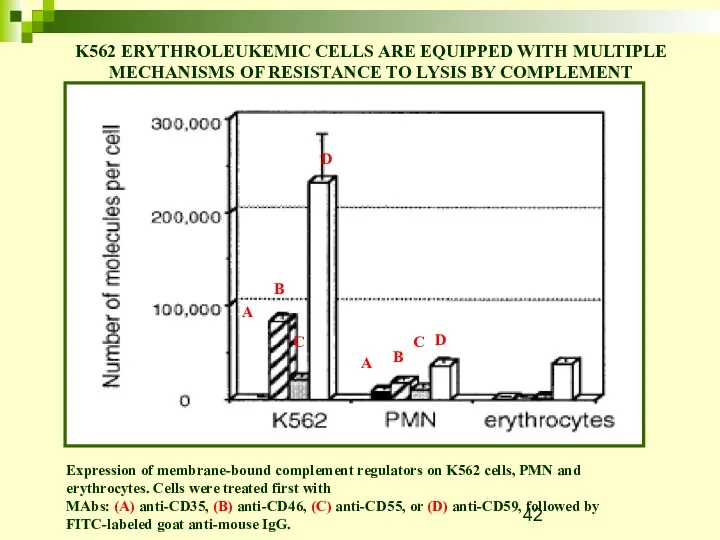
Expression of membrane-bound complement regulators on K562 cells, PMN and erythrocytes.
Expression of membrane-bound complement regulators on K562 cells, PMN and erythrocytes.

Важность mCRPs как механизма устойчивости к комплементу была также показана с
Важность mCRPs как механизма устойчивости к комплементу была также показана с
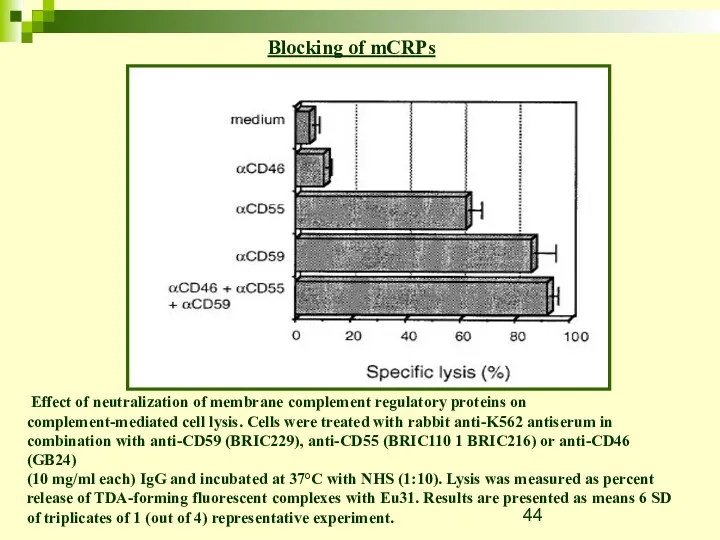
Effect of neutralization of membrane complement regulatory proteins on complement-mediated
Effect of neutralization of membrane complement regulatory proteins on complement-mediated

Основные механизмы устойчивости к комплементу:
I. экспрессия растворимых белков-регуляторов
Основные механизмы устойчивости к комплементу:
I. экспрессия растворимых белков-регуляторов
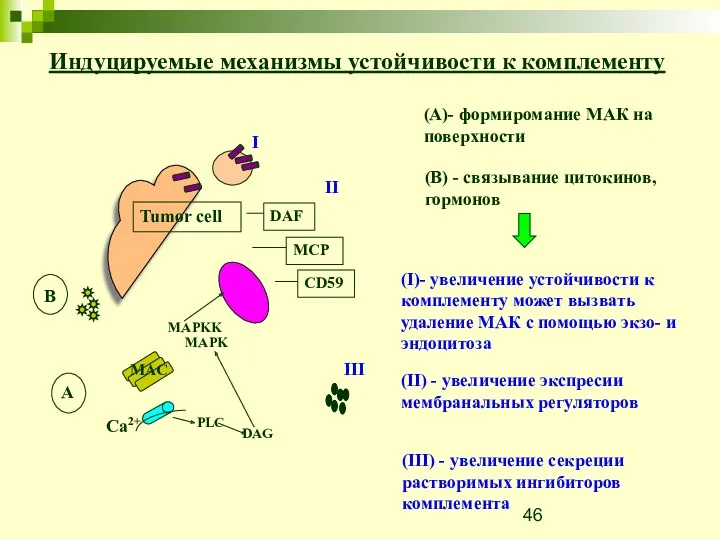
Индуцируемые механизмы устойчивости к комплементу
Tumor cell
DAF
MCP
CD59
I
II
III
B
Ca2+
MAC
A
PLC
DAG
MAPK
MAPKK
(I)- увеличение устойчивости к комплементу
Индуцируемые механизмы устойчивости к комплементу
Tumor cell
DAF
MCP
CD59
I
II
III
B
Ca2+
MAC
A
PLC
DAG
MAPK
MAPKK
(I)- увеличение устойчивости к комплементу

Интересно, что одним из наиболее потенциальных подобных агентов является сам МАК.
Для
Интересно, что одним из наиболее потенциальных подобных агентов является сам МАК.
Для

Также было показано, что клетки К562 могут быть защищены от лизиса
Также было показано, что клетки К562 могут быть защищены от лизиса

Обработка клеток с помощью суб-литического комплемента приводит в действие клеточный механизм,
Обработка клеток с помощью суб-литического комплемента приводит в действие клеточный механизм,

Mortalin/GRP75 promotes release of membrane
vesicles from immune attacked cells and protection
Mortalin/GRP75 promotes release of membrane
vesicles from immune attacked cells and protection
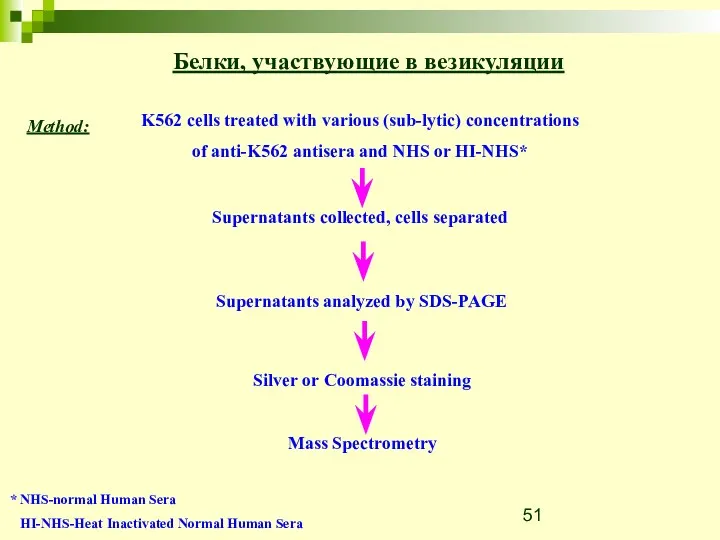
Белки, участвующие в везикуляции
Method:
* NHS-normal Human Sera
HI-NHS-Heat Inactivated Normal Human
Белки, участвующие в везикуляции
Method:
* NHS-normal Human Sera
HI-NHS-Heat Inactivated Normal Human
NHS
HI-NHS
Mortalin/GRP75 promotes release of membranevesicles from immune attacked cells and protection
from
NHS
HI-NHS
Mortalin/GRP75 promotes release of membranevesicles from immune attacked cells and protection
from
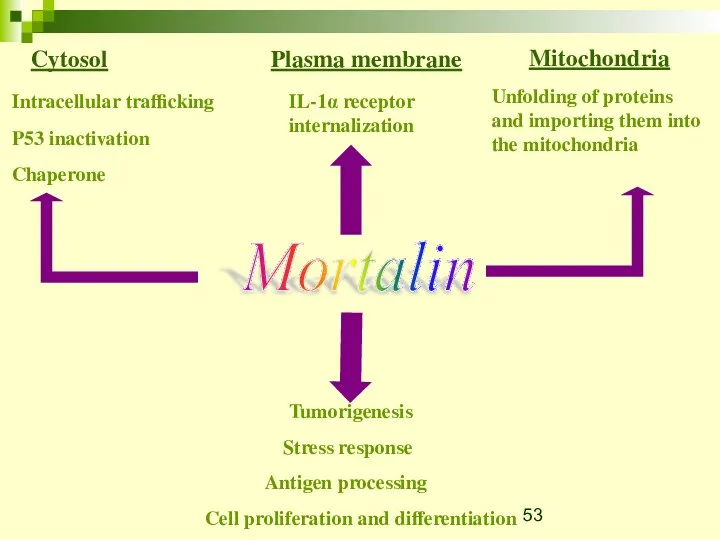
Mortalin
Mitochondria
Unfolding of proteins and importing them into the mitochondria
Cytosol
Intracellular trafficking
Mortalin
Mitochondria
Unfolding of proteins and importing them into the mitochondria
Cytosol
Intracellular trafficking
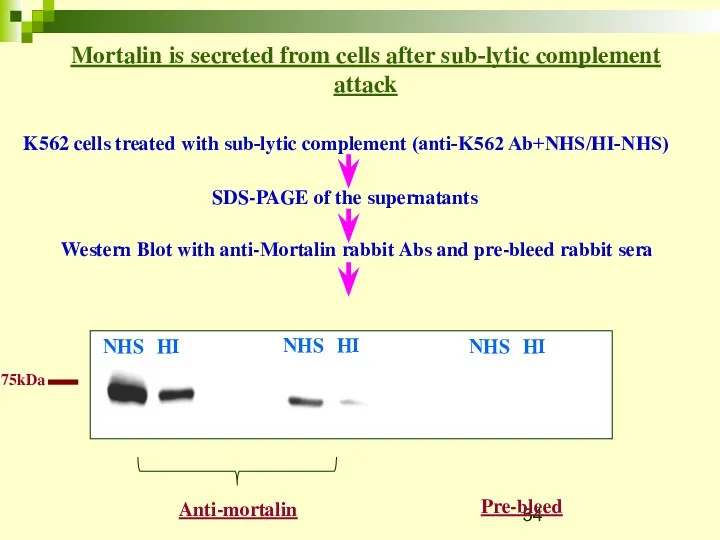
Mortalin is secreted from cells after sub-lytic complement attack
K562 cells treated
Mortalin is secreted from cells after sub-lytic complement attack
K562 cells treated

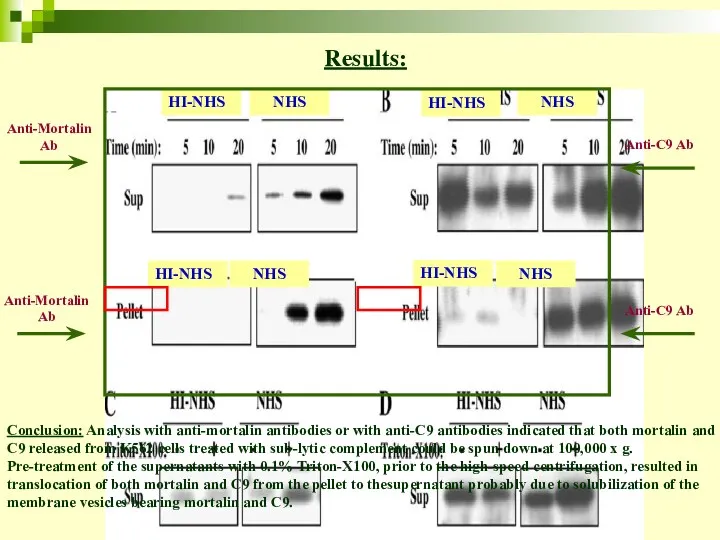
Mortalin and C9 are associated with membrane vesicles
K562 treated with sub-lytic
Mortalin and C9 are associated with membrane vesicles
K562 treated with sub-lytic
HI-NHS
NHS
HI-NHS
NHS
Anti-Mortalin Ab
Anti-C9 Ab
Anti-Mortalin Ab
Anti-C9 Ab
Results:
HI-NHS
NHS
HI-NHS
NHS
Conclusion: Analysis with anti-mortalin antibodies or with
HI-NHS
NHS
HI-NHS
NHS
Anti-Mortalin Ab
Anti-C9 Ab
Anti-Mortalin Ab
Anti-C9 Ab
Results:
HI-NHS
NHS
HI-NHS
NHS
Conclusion: Analysis with anti-mortalin antibodies or with
NHS +
anti-mortalin
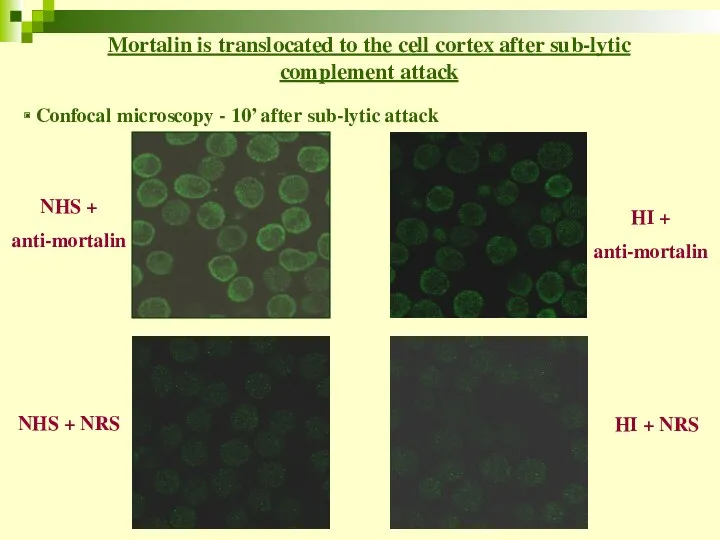
Mortalin is translocated to the cell cortex after sub-lytic complement
NHS +
anti-mortalin
Mortalin is translocated to the cell cortex after sub-lytic complement

Addition of anti-Mortalin Ab to the experiments
Anti-Mortalin Ab lowers cells’
Addition of anti-Mortalin Ab to the experiments
Anti-Mortalin Ab lowers cells’
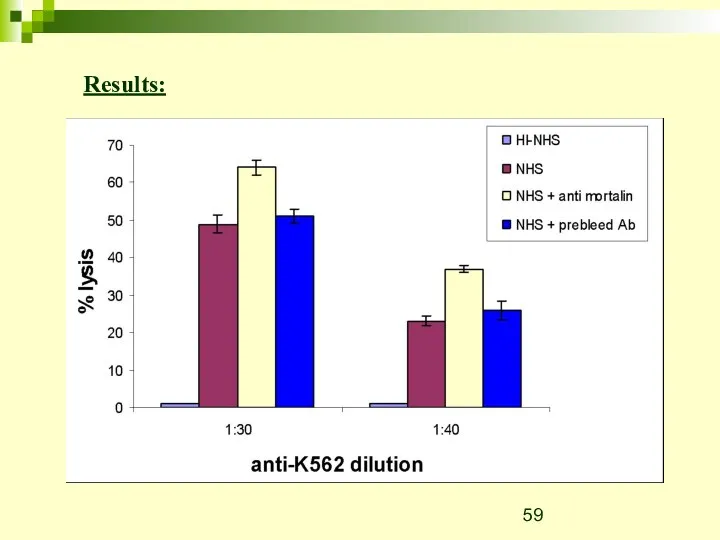
Results:
Results:

Increased sensitivity of early apoptotic cells to
complement-mediated lysis
Gitit Attali, Dana Gancz
Increased sensitivity of early apoptotic cells to
complement-mediated lysis
Gitit Attali, Dana Gancz

Интеракция белков комплемента с апоптотическими клетками была продемонстрирована, однако эта тема
Интеракция белков комплемента с апоптотическими клетками была продемонстрирована, однако эта тема

Ранние апопторические клетки чувствительны к лизису комплементом
Цель: проверить интеракцию между ранними
Ранние апопторические клетки чувствительны к лизису комплементом
Цель: проверить интеракцию между ранними
Вывод: ранние апоптотические клетки более чувствительны к лизису антителом + комплементом,
Вывод: ранние апоптотические клетки более чувствительны к лизису антителом + комплементом,

Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases)
Цель: проверить
Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases)
Цель: проверить
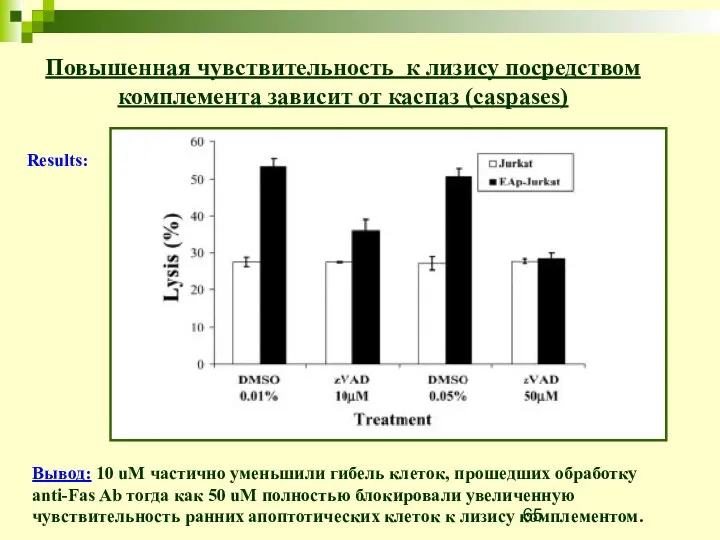
Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases)
Вывод: 10
Повышенная чувствительность к лизису посредством комплемента зависит от каспаз (caspases)
Вывод: 10

Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом
Анти-апоптотические эффекты Bcl-2
Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом
Анти-апоптотические эффекты Bcl-2
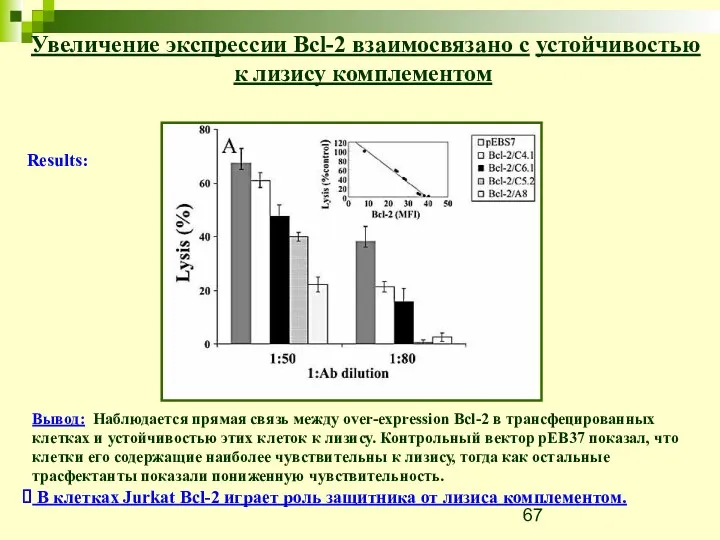
Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом
Results:
Вывод: Наблюдается прямая
Увеличение экспрессии Bcl-2 взаимосвязано с устойчивостью к лизису комплементом
Results:
Вывод: Наблюдается прямая
 Тыныс алудың маңызы. Тыныс алу мүшелері көмей бронхылар кеңірдек
Тыныс алудың маңызы. Тыныс алу мүшелері көмей бронхылар кеңірдек Металды қалыпталған сауытты дайындауда қолданылатын болат маркасы
Металды қалыпталған сауытты дайындауда қолданылатын болат маркасы Средства, влияющие на Н-холинорецепторы. Н-холиномиметики. Ганглиоблокаторы. Миорелаксанты периферического действия (Лекция 5)
Средства, влияющие на Н-холинорецепторы. Н-холиномиметики. Ганглиоблокаторы. Миорелаксанты периферического действия (Лекция 5) Особенности фармакокинетики и фармакотерапии в неонатальном периоде
Особенности фармакокинетики и фармакотерапии в неонатальном периоде Острые промиелоцитарные лейкозы
Острые промиелоцитарные лейкозы Журектiн аускультациясы патологияда
Журектiн аускультациясы патологияда Промывание желудка
Промывание желудка Осложнения желудочно-кишечного тракта внутриутробного развития
Осложнения желудочно-кишечного тракта внутриутробного развития Продуктивное воспаление
Продуктивное воспаление 05. медичне страхування
05. медичне страхування Рестриктивные кардиомиопатии. Формы РКМП
Рестриктивные кардиомиопатии. Формы РКМП Опухоли слюнных желез
Опухоли слюнных желез Изосерологическая несовместимость крови матери и плода
Изосерологическая несовместимость крови матери и плода Болезнь Лайма
Болезнь Лайма Рак яичников
Рак яичников Введение в доказательную медицину
Введение в доказательную медицину Вплив алкоголю на дітей та підлітків
Вплив алкоголю на дітей та підлітків Дерматомиозит
Дерматомиозит Кавказские Минеральные Воды
Кавказские Минеральные Воды Профессиональная гигиена полости рта
Профессиональная гигиена полости рта Т-20С.31 янв
Т-20С.31 янв Дифференциально-диагностический поиск у пациентов с гипогликемическим синдромом
Дифференциально-диагностический поиск у пациентов с гипогликемическим синдромом Сердце, аорта, кровь, вены, капилляры
Сердце, аорта, кровь, вены, капилляры Ларингит
Ларингит Проблемы нормативного обеспечения в сфере информатизации здравоохранения
Проблемы нормативного обеспечения в сфере информатизации здравоохранения Қан айналымды зерттеу әдістері. Интегралдық және регионарлық реография. Реография
Қан айналымды зерттеу әдістері. Интегралдық және регионарлық реография. Реография Анемия у детей
Анемия у детей Клиническая характеристика больных и оказание неотложной помощи при почечной колике
Клиническая характеристика больных и оказание неотложной помощи при почечной колике