- В-лимфоциты – основные эффекторы гуморального иммунного ответа. Лекция 5

Содержание
- 2. Адаптивный B-клеточный иммунный ответ работает по принципу клональной селекции. - Стволовая клетка дает начало огромному разнообразию
- 3. BCR – активирующий рецептор. Igα и Igβ -инвариантные сигнальные субъединицы. Между собой связаны дисульфидной связью и
- 4. Развитие и дифференцировка В-лимфоцитов Elodie Foulquier and Marie-Paule Lefranc. 2001 АГ специфичность В-клетки определяется на ранних
- 5. Первые стадии развития В-клетки проходят в костном мозге, где осуществляется негативная селекция BCRs и В-клеток (участвуют
- 6. При дифференцировке в костном мозге В-клеткам (от стадии предшественника до стадии незрелой/immature В-клетки) необходим тесный контакт
- 7. Развивающаяся В-клетка находится в контакте с клетками стромы и передвигается в костном мозге
- 8. Стадии дифференцировки В-лимфоцитов отличаются по экспрессии цепей иммуноглобулинов и поверхностных маркеров. Развитие В-клетки в костном мозге
- 9. В про-В-клетке правильно перестроенный ИГ VH-ген немедленно экспрессируется (транскрипция идет всегда) и тестируется в виде пре-BCR.
- 10. У легких цепей ИГ есть несколько попыток V-J перестройки! Непродуктивная перестройка V-J генных сегментов легких цепей
- 11. IgHa/a IgHb/b IgHa/b Для генов иммуноглобулинов – аллельное исключение в В-клетках. В каждой В-клетке экспрессируется только
- 12. Дифференцировка на первых стадиях развития контролируется перестройками генных сегментов BCR. При VDJ или VJ перестройках –
- 13. Дилемма для В-клеток Огромное Разнообразие BCRs - потенциально против всех возможных патогенов Отсутствие реактивности BCRs c
- 14. Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге. Центральная толерантность. КАК? По взаимодействию BCR
- 15. Редактирование BCR (Receptor editing) с помощью замены легкой цепи и, соответственно, их антигенной специфичности «спасает» некоторые
- 16. Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге. Центральная толерантность. КАК? По взаимодействию BCR
- 17. Центральная толерантность – не идеальна, позволяет редким аутореактивным В-клеткам созревать. Зачем? - Для поддержания баланса между
- 18. Наивная зрелая В-клетка-созревание в фолликулах вторичных лимфоидных органов(фолликулы – первичные/вторичные) Кровь Встреча с АГ (+цитокины, +ко-стимулирующие
- 19. BCR – В-клеточный рецептор Для чего нужен BCR? - Для взаимодействия В-клетки с окружающей внеклеточной средой.
- 20. BCR – активирующий рецептор: sIgM + Igα и Igβ -инвариантные сигнальные субъединицы. Между собой связаны дисульфидной
- 21. Связывание рецептора с лигандом (кросс-сшивка BCR с АГ приводит к кластеризации рецептора на мембране лимфоцита Кросс-сшивка
- 22. Как работают ITAM-содержащие рецепторы? Лиганд кросс-сшивает рецептор. Тирозины в ITAM-мотиве фосфорилируются тирозин-киназой, ассоциированной с рецептором. После
- 23. Связывание АГ с BCRs приводит к их кластеризации, что вызывает взаимо-активацию ассоциированных с рецептором тирозин-киназ семейства
- 24. BCR signaling pathway
- 25. Сигналинг через BCR усиливается ко-рецепторами, которые связывают тот же самый лиганд: ко-рецепторный комплекс CD19/ CD21 (CR2)/
- 26. Все антигены, на которые отвечает В-клетка делятся на: Т-независимые (LPS) и Т-зависимые (белки, в основном) Ответ
- 27. Для эффективной активации наивной В-клетке требуется, как правило, два сигнала: 1 – от BCR, 2- от
- 28. Для получения «помощи» от Т-лимфоцита В-клетка должна сначала интернализовать АГ, и затем «представить» его на своей
- 29. Активация В-клеток происходит во вторичных лимфоидных органах, где В- и Т-клетки занимают строго определенные гистологические компартменты.
- 30. Лимфоузлы имеют похожие на селезенку клеточные компартменты. Но АГ и АГ-несущие дендритные клетки и АГ-несущие макрофаги
- 31. Вторичный лимфоидный фолликул селезенки мыши с зародышевым центром FDC – фолликулярные дендритные клетки, имеют длинные отростки
- 32. Зародышевые центры – компартменты, где происходит размножение и дифференцировка активированных В-клеток (в случае хронической инфекции). Два
- 33. Как отбираются клетки с высоко аффинным BCR? Считается, что В-клетки с более высоко аффинным BCR сильнее
- 34. Аффинное созревание иммунного ответа в зародышевом центре. Точечные мутации в СDRs V-областей ИГ, постепенно накапливаются по
- 35. ФОЛЛИКУЛЯРНЫЕ ДЕНДРИТНЫЕ КЛЕТКИ (FDC) – уникальная популяция клеток, необходимая для развития зародышевого центра и продукции высоко
- 36. Локализация FDC – первичные В-клеточные фолликулы и светлая зона GC вторичных В-клеточных фолликулов. Экспрессируют хемокин CXCL13,
- 37. Balthasar A. Heesters et al., Nature Reviews Immunology 2014 FDC отвечают за доступность антигена для В-клетки
- 38. Balthasar A. Heesters et al., Nature Reviews Immunology 2014 Уникальная особенность FDC поглощают и длительно сохраняют
- 39. FDC - резервуары для HIV HIV спонтанно связывает C3d и через CR2 попадает в недеградирующие условия
- 40. FDC, образующие зародышевые центры в нетипичных местах при патологических состояниях: а) в суставе при ревматоидном артрите,
- 41. Ранние стадии ответа – секреция IgM, далее класс-переключение – IgG и IgA. Как регулируется класс-переключение? По-видимому,
- 42. Все антигены, на которые отвечает В-клетка делятся на: Т-независимые (LPS) и Т-зависимые B1 клетки, MZ B
- 43. Поздние стадии В-клеточной дифференцировки регулируются множеством транскрипционных факторов Nutt et al., Nature Review Immunology, 2015 Активированная
- 44. Плазматическая клетка – терминальная стадия дифференцировки В-клетки. Сильные морфологические и функциональные изменения по сравнению со зрелой
- 46. Скачать презентацию

Адаптивный B-клеточный иммунный ответ работает по принципу клональной селекции.
- Стволовая клетка
Адаптивный B-клеточный иммунный ответ работает по принципу клональной селекции.
- Стволовая клетка
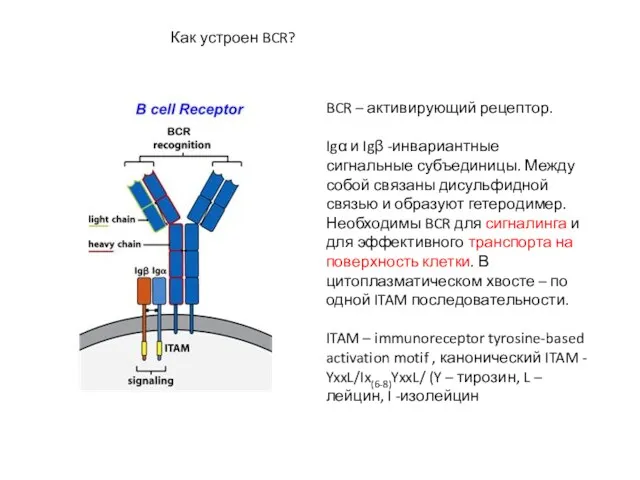
BCR – активирующий рецептор.
Igα и Igβ -инвариантные сигнальные субъединицы. Между собой
BCR – активирующий рецептор.
Igα и Igβ -инвариантные сигнальные субъединицы. Между собой
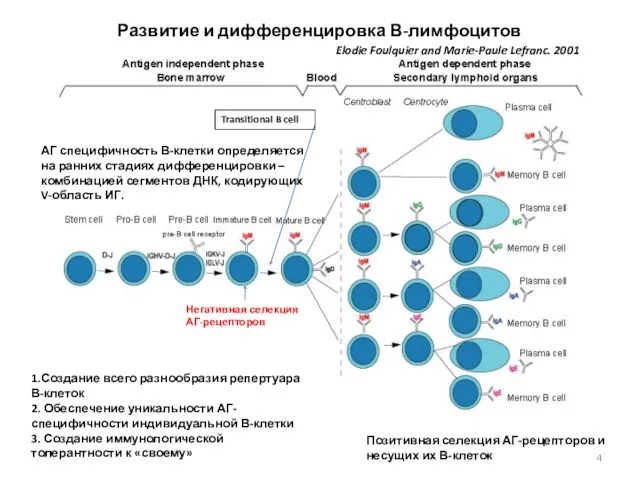
Развитие и дифференцировка В-лимфоцитов
Elodie Foulquier and Marie-Paule Lefranc. 2001
АГ специфичность
Развитие и дифференцировка В-лимфоцитов
Elodie Foulquier and Marie-Paule Lefranc. 2001
АГ специфичность
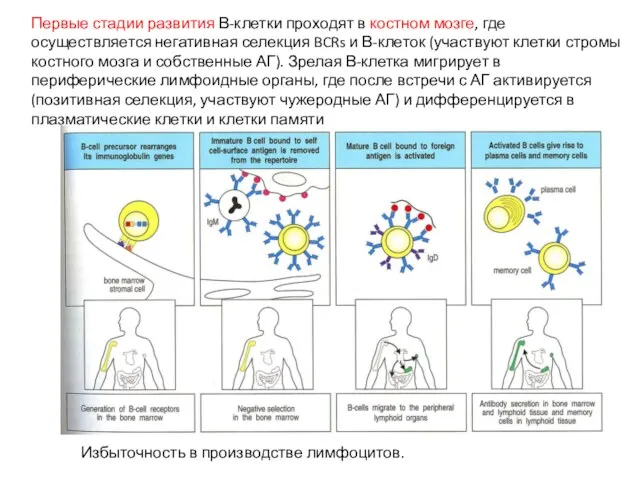
Первые стадии развития В-клетки проходят в костном мозге, где осуществляется негативная
Первые стадии развития В-клетки проходят в костном мозге, где осуществляется негативная
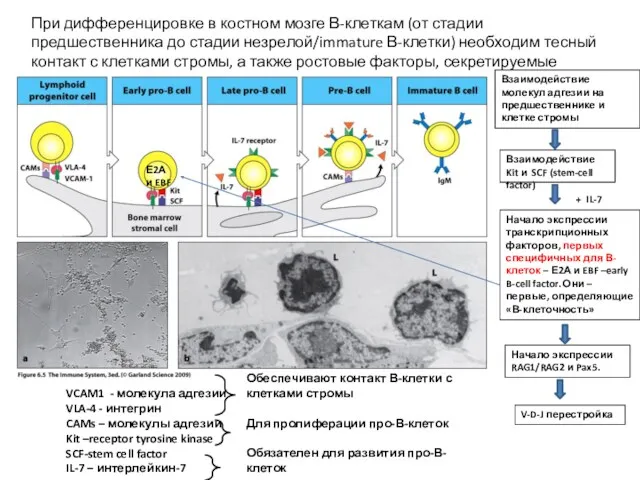
При дифференцировке в костном мозге В-клеткам (от стадии предшественника до стадии
При дифференцировке в костном мозге В-клеткам (от стадии предшественника до стадии
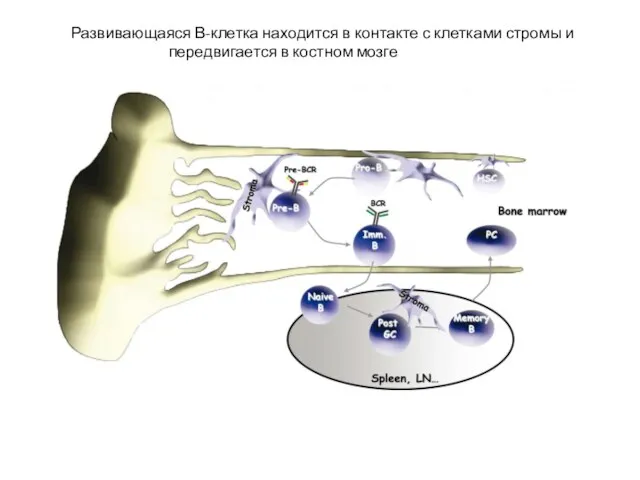
Развивающаяся В-клетка находится в контакте с клетками стромы и
передвигается в
Развивающаяся В-клетка находится в контакте с клетками стромы и
передвигается в
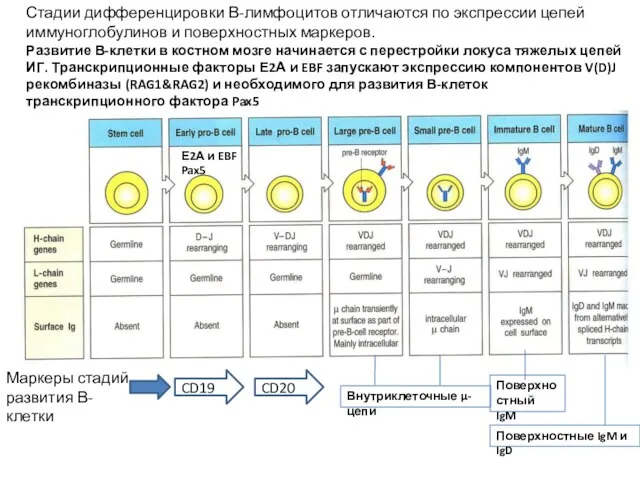
Стадии дифференцировки В-лимфоцитов отличаются по экспрессии цепей иммуноглобулинов и поверхностных маркеров.
Развитие
Стадии дифференцировки В-лимфоцитов отличаются по экспрессии цепей иммуноглобулинов и поверхностных маркеров.
Развитие
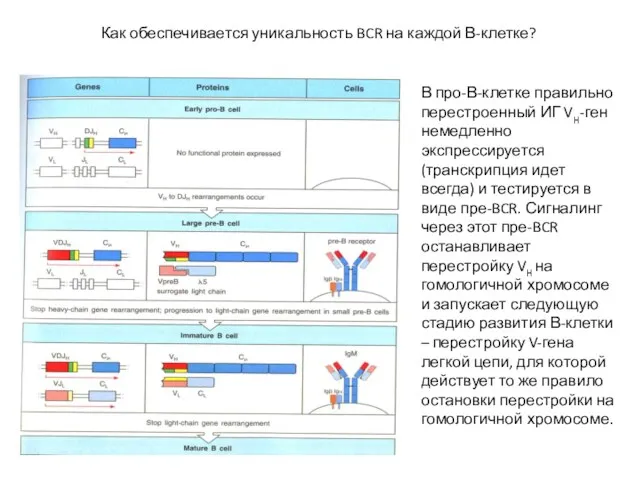
В про-В-клетке правильно перестроенный ИГ VH-ген немедленно экспрессируется (транскрипция идет всегда)
В про-В-клетке правильно перестроенный ИГ VH-ген немедленно экспрессируется (транскрипция идет всегда)
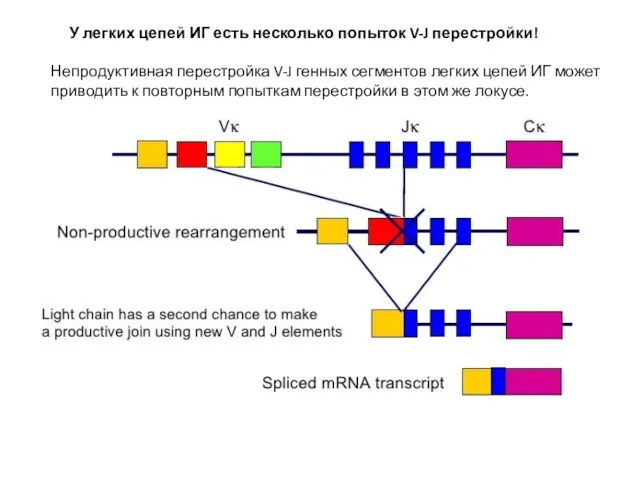
У легких цепей ИГ есть несколько попыток V-J перестройки!
Непродуктивная
У легких цепей ИГ есть несколько попыток V-J перестройки!
Непродуктивная
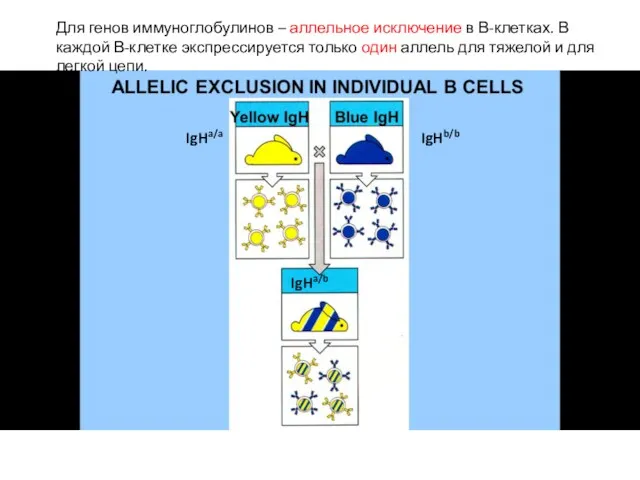
IgHa/a IgHb/b
IgHa/b
Для генов иммуноглобулинов – аллельное исключение в В-клетках. В
IgHa/a IgHb/b
IgHa/b
Для генов иммуноглобулинов – аллельное исключение в В-клетках. В
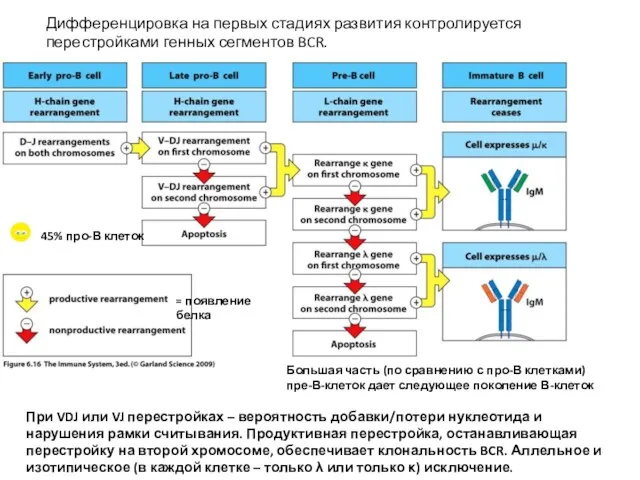
Дифференцировка на первых стадиях развития контролируется перестройками генных сегментов BCR.
При VDJ
Дифференцировка на первых стадиях развития контролируется перестройками генных сегментов BCR.
При VDJ
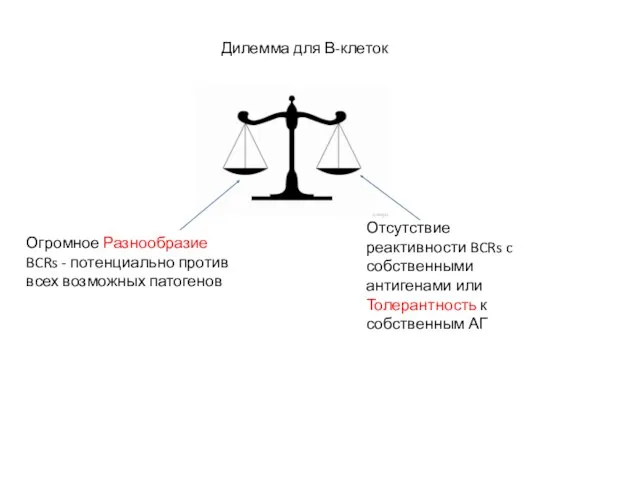
Дилемма для В-клеток
Огромное Разнообразие BCRs - потенциально против всех возможных патогенов
Отсутствие
Дилемма для В-клеток
Огромное Разнообразие BCRs - потенциально против всех возможных патогенов
Отсутствие
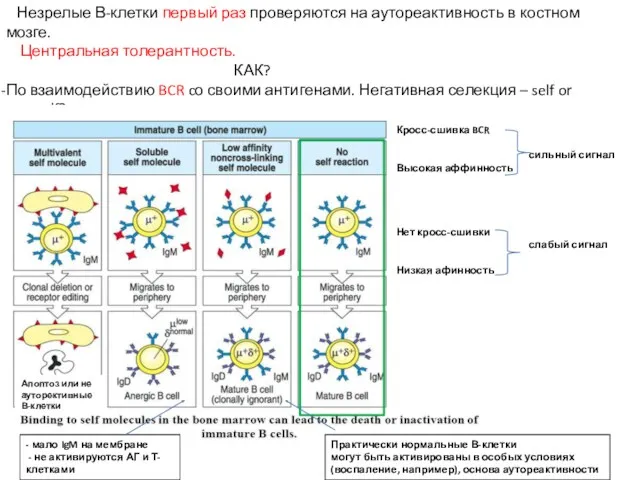
Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге.
Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге.
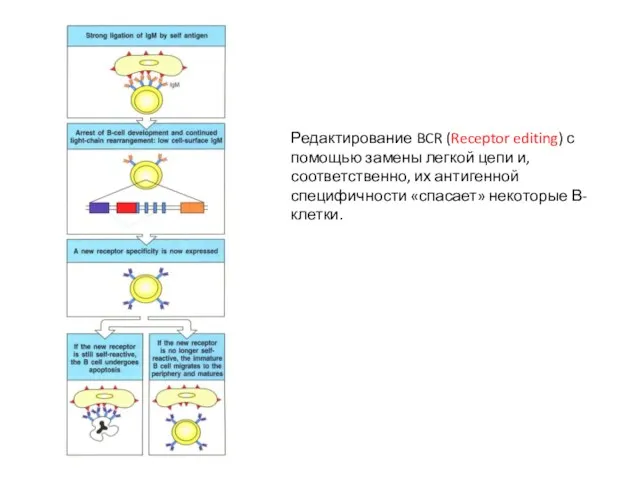
Редактирование BCR (Receptor editing) с помощью замены легкой цепи и, соответственно,
Редактирование BCR (Receptor editing) с помощью замены легкой цепи и, соответственно,
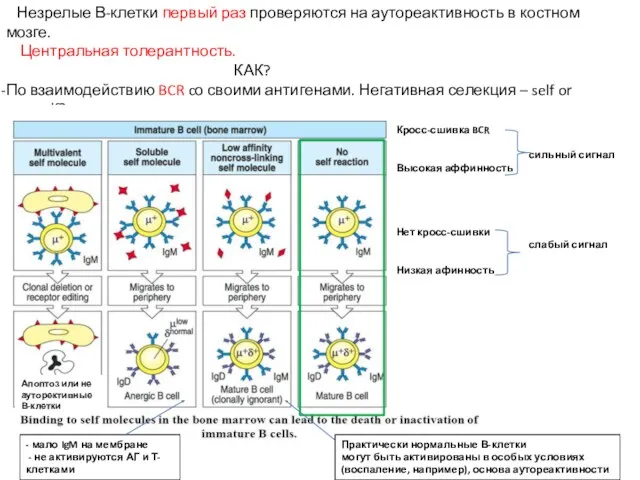
Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге.
Незрелые В-клетки первый раз проверяются на аутореактивность в костном мозге.
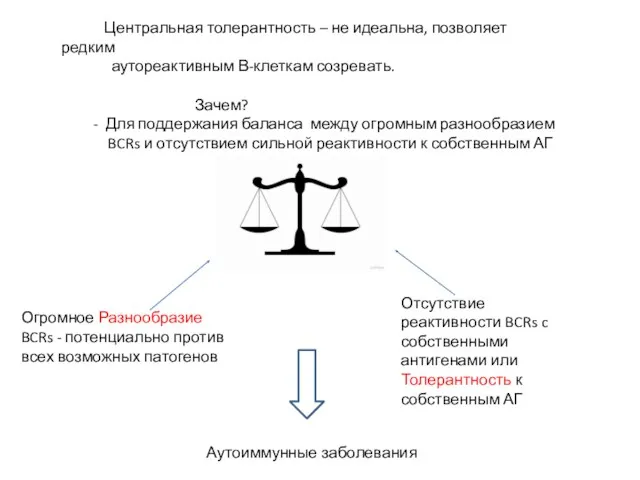
Центральная толерантность – не идеальна, позволяет редким
аутореактивным В-клеткам
Центральная толерантность – не идеальна, позволяет редким
аутореактивным В-клеткам
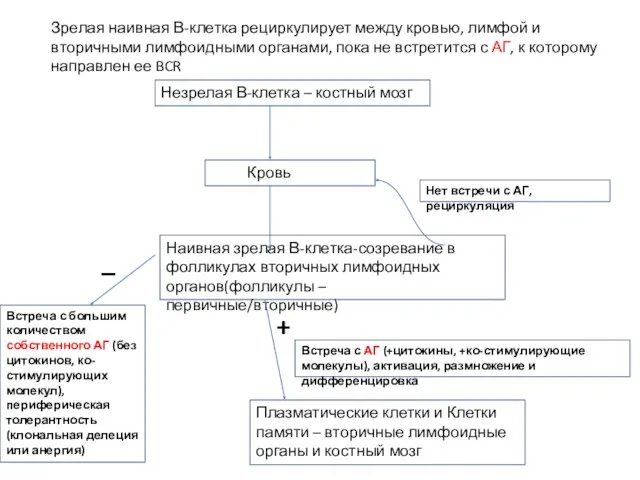
Наивная зрелая В-клетка-созревание в фолликулах вторичных лимфоидных органов(фолликулы – первичные/вторичные)
Кровь
Наивная зрелая В-клетка-созревание в фолликулах вторичных лимфоидных органов(фолликулы – первичные/вторичные)
Кровь

BCR – В-клеточный рецептор
Для чего нужен BCR?
- Для взаимодействия В-клетки
BCR – В-клеточный рецептор
Для чего нужен BCR?
- Для взаимодействия В-клетки
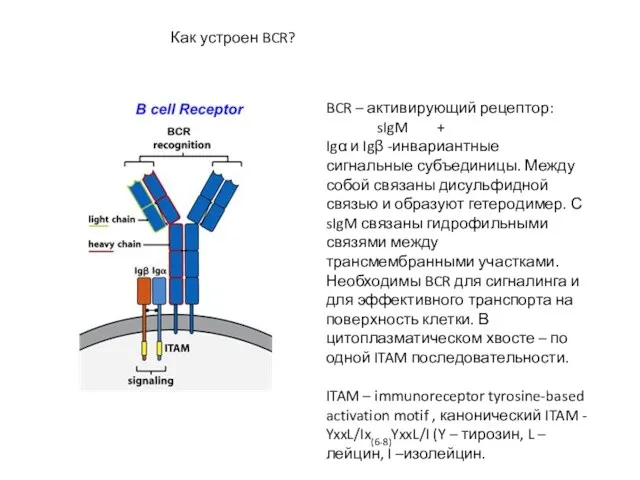
BCR – активирующий рецептор:
sIgM +
Igα и Igβ -инвариантные сигнальные
BCR – активирующий рецептор:
sIgM +
Igα и Igβ -инвариантные сигнальные
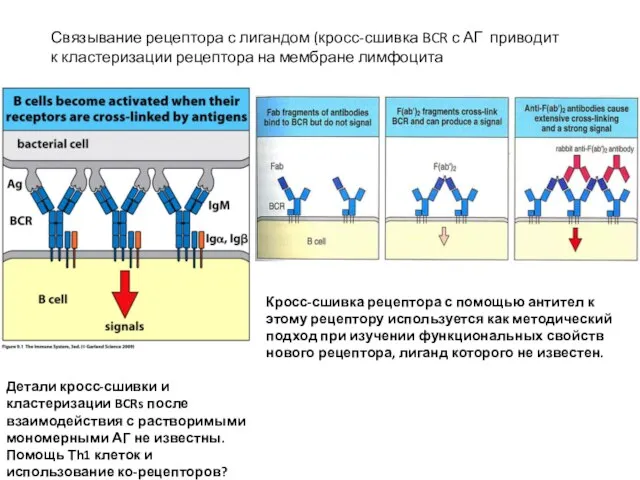
Связывание рецептора с лигандом (кросс-сшивка BCR с АГ приводит к кластеризации
Связывание рецептора с лигандом (кросс-сшивка BCR с АГ приводит к кластеризации
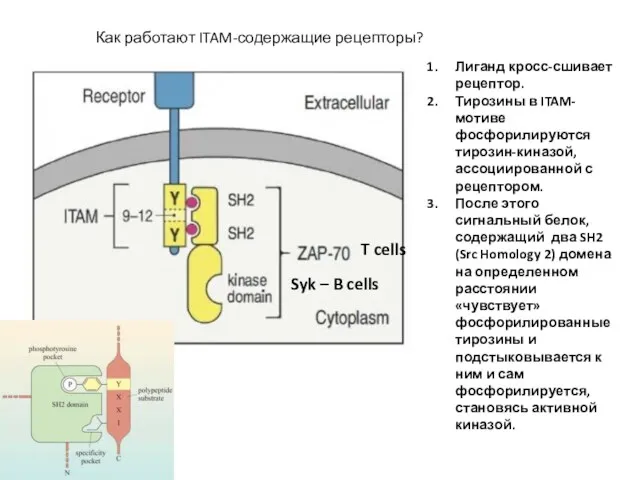
Как работают ITAM-содержащие рецепторы?
Лиганд кросс-сшивает рецептор.
Тирозины в ITAM-мотиве фосфорилируются тирозин-киназой, ассоциированной
Как работают ITAM-содержащие рецепторы?
Лиганд кросс-сшивает рецептор.
Тирозины в ITAM-мотиве фосфорилируются тирозин-киназой, ассоциированной
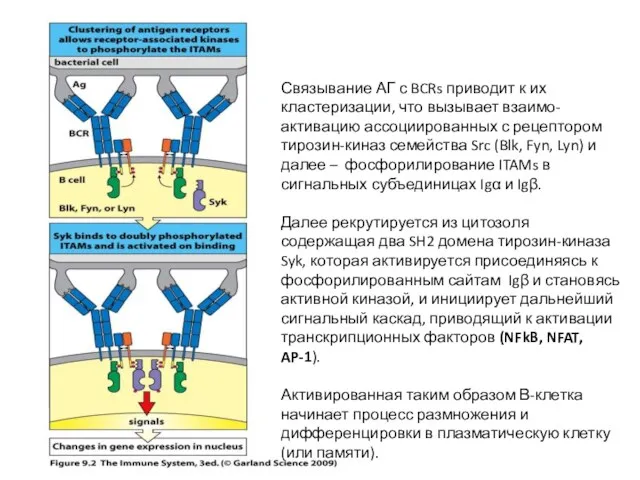
Связывание АГ с BCRs приводит к их кластеризации, что вызывает взаимо-активацию
Связывание АГ с BCRs приводит к их кластеризации, что вызывает взаимо-активацию
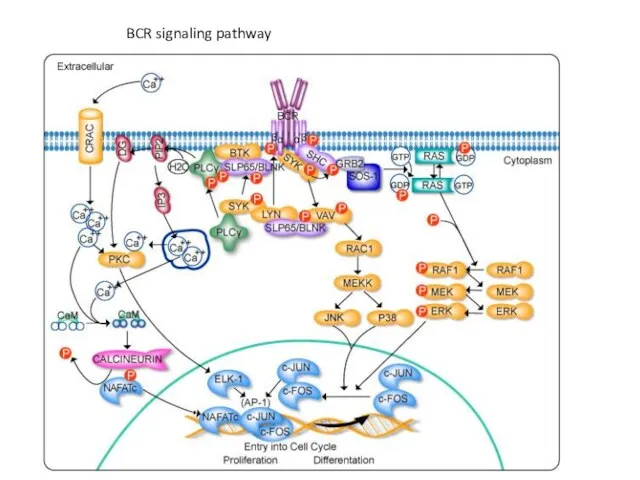
BCR signaling pathway
BCR signaling pathway
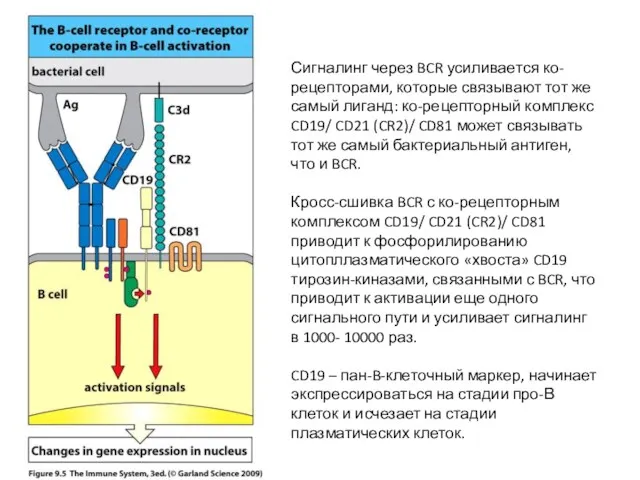
Сигналинг через BCR усиливается ко-рецепторами, которые связывают тот же самый лиганд:
Сигналинг через BCR усиливается ко-рецепторами, которые связывают тот же самый лиганд:
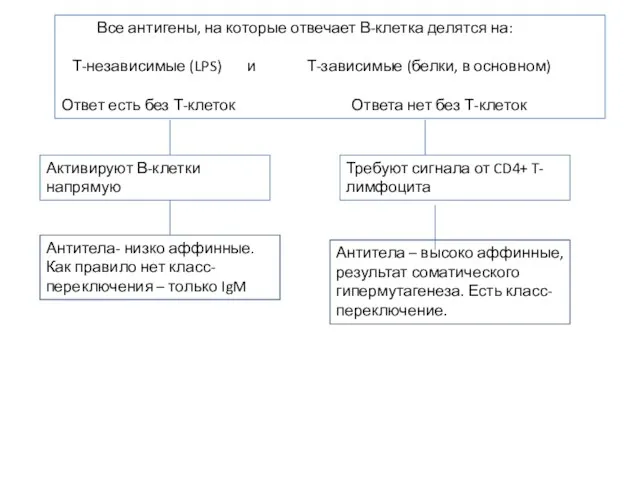
Все антигены, на которые отвечает В-клетка делятся на:
Т-независимые
Все антигены, на которые отвечает В-клетка делятся на:
Т-независимые
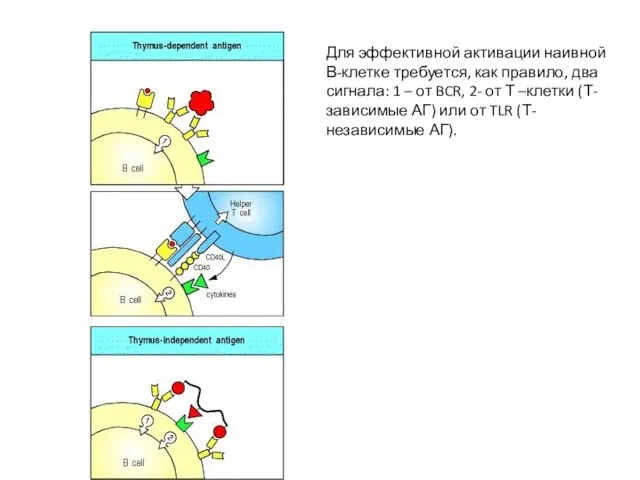
Для эффективной активации наивной В-клетке требуется, как правило, два сигнала: 1
Для эффективной активации наивной В-клетке требуется, как правило, два сигнала: 1
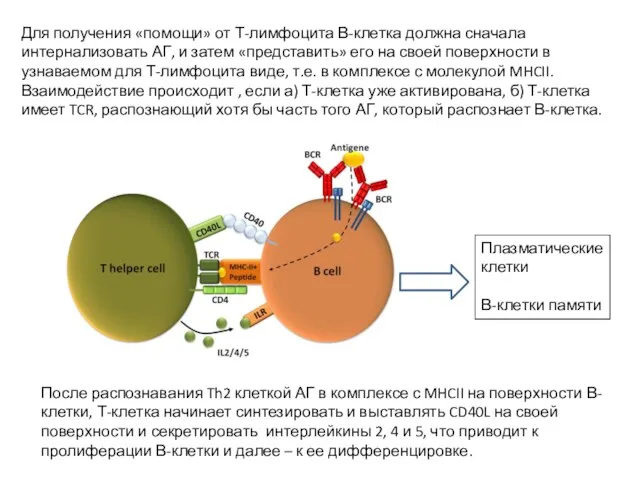
Для получения «помощи» от Т-лимфоцита В-клетка должна сначала интернализовать АГ, и
Для получения «помощи» от Т-лимфоцита В-клетка должна сначала интернализовать АГ, и
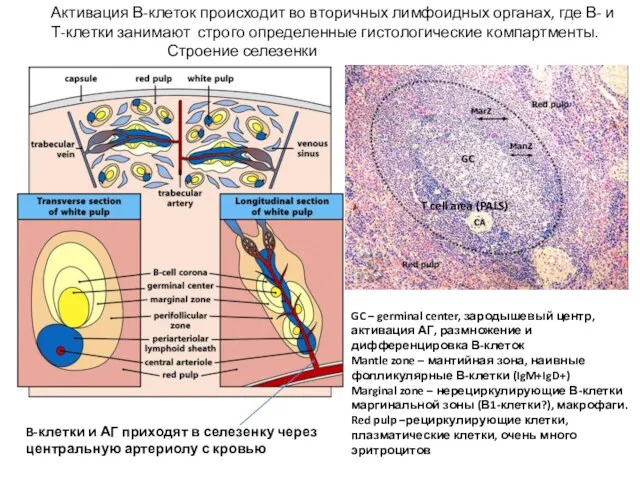
Активация В-клеток происходит во вторичных лимфоидных органах, где В- и Т-клетки
Активация В-клеток происходит во вторичных лимфоидных органах, где В- и Т-клетки
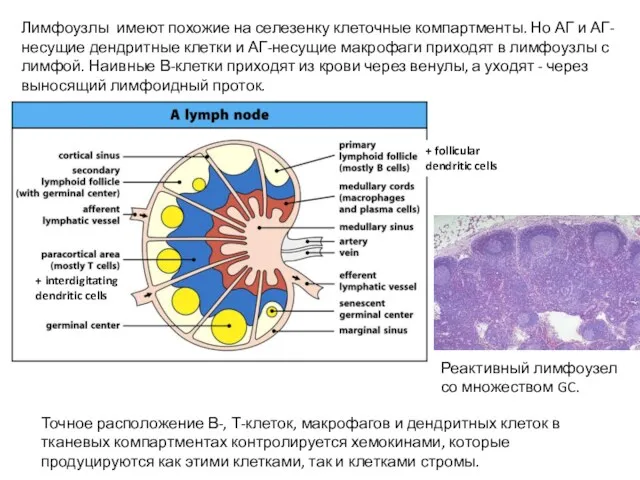
Лимфоузлы имеют похожие на селезенку клеточные компартменты. Но АГ и АГ-несущие
Лимфоузлы имеют похожие на селезенку клеточные компартменты. Но АГ и АГ-несущие
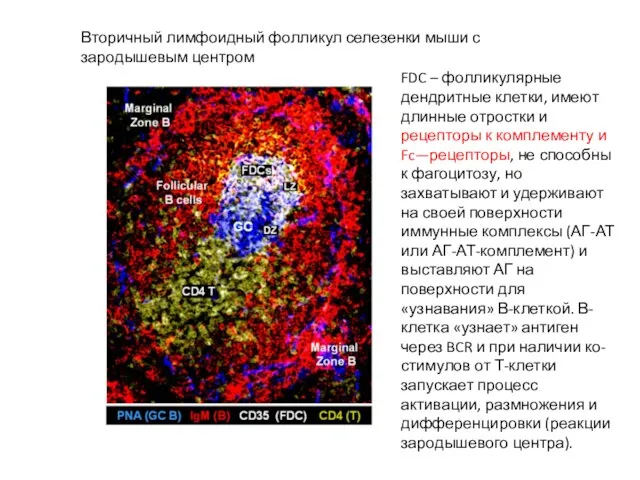
Вторичный лимфоидный фолликул селезенки мыши с зародышевым центром
FDC – фолликулярные дендритные
Вторичный лимфоидный фолликул селезенки мыши с зародышевым центром
FDC – фолликулярные дендритные
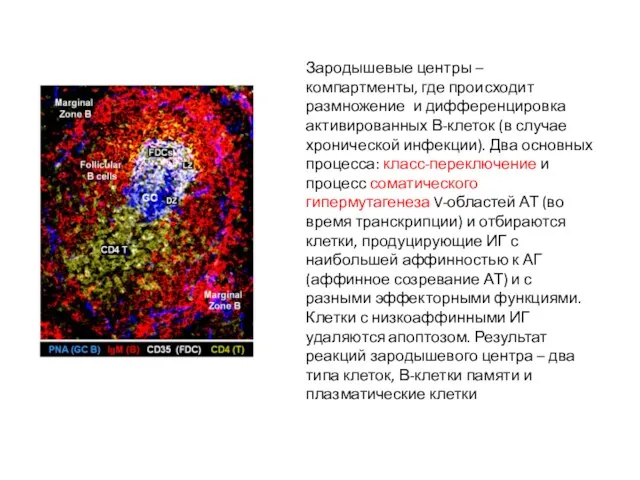
Зародышевые центры – компартменты, где происходит размножение и дифференцировка активированных В-клеток
Зародышевые центры – компартменты, где происходит размножение и дифференцировка активированных В-клеток
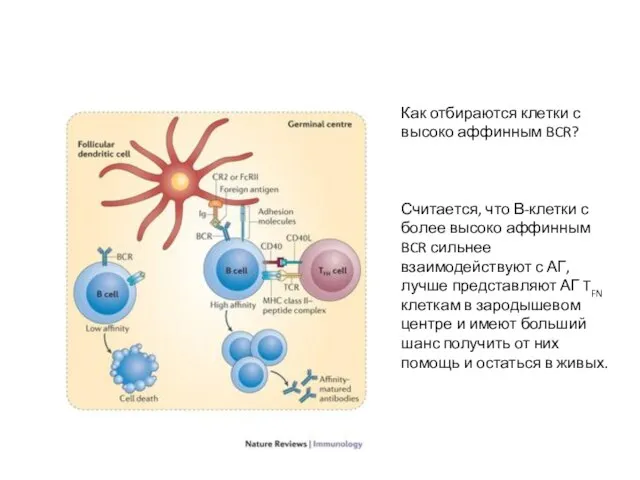
Как отбираются клетки с высоко аффинным BCR?
Считается, что В-клетки с более
Как отбираются клетки с высоко аффинным BCR?
Считается, что В-клетки с более
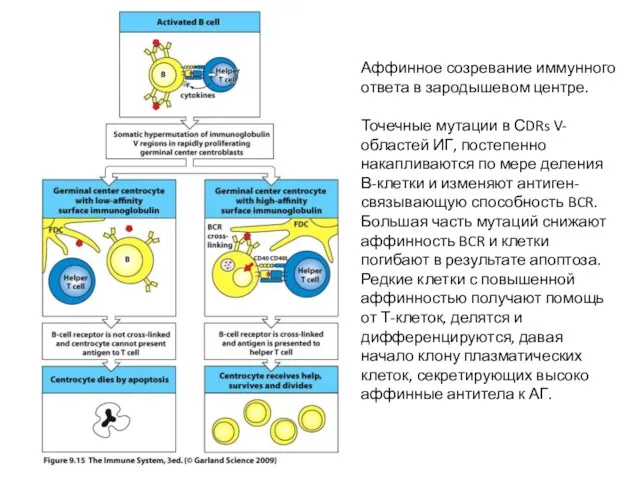
Аффинное созревание иммунного ответа в зародышевом центре.
Точечные мутации в СDRs V-областей
Аффинное созревание иммунного ответа в зародышевом центре.
Точечные мутации в СDRs V-областей

ФОЛЛИКУЛЯРНЫЕ ДЕНДРИТНЫЕ КЛЕТКИ (FDC)
– уникальная популяция клеток, необходимая для развития
ФОЛЛИКУЛЯРНЫЕ ДЕНДРИТНЫЕ КЛЕТКИ (FDC)
– уникальная популяция клеток, необходимая для развития
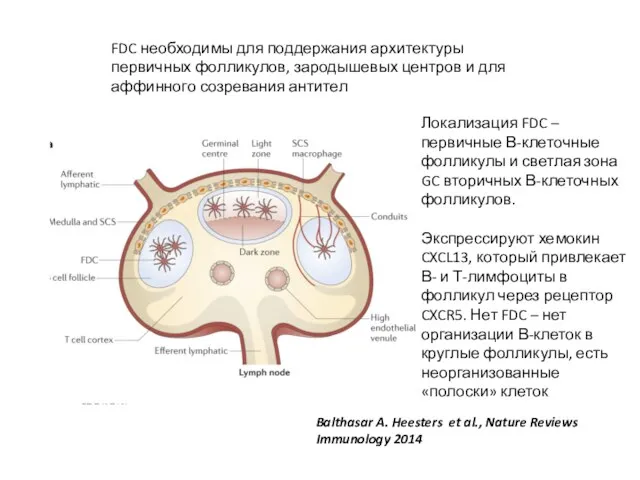
Локализация FDC – первичные В-клеточные фолликулы и светлая зона GC вторичных
Локализация FDC – первичные В-клеточные фолликулы и светлая зона GC вторичных
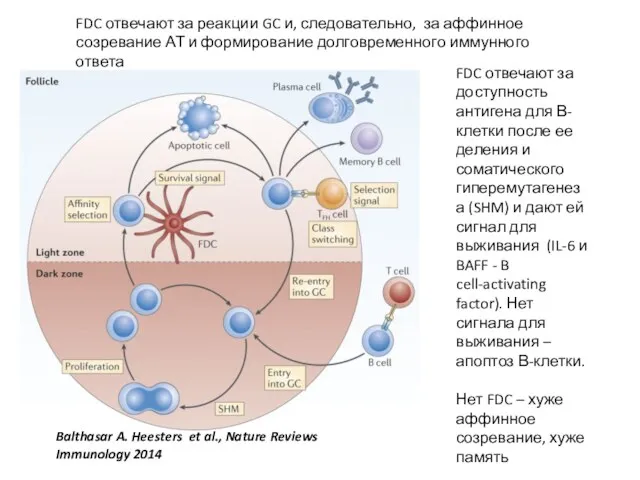
Balthasar A. Heesters et al., Nature Reviews Immunology 2014
FDC отвечают
Balthasar A. Heesters et al., Nature Reviews Immunology 2014
FDC отвечают
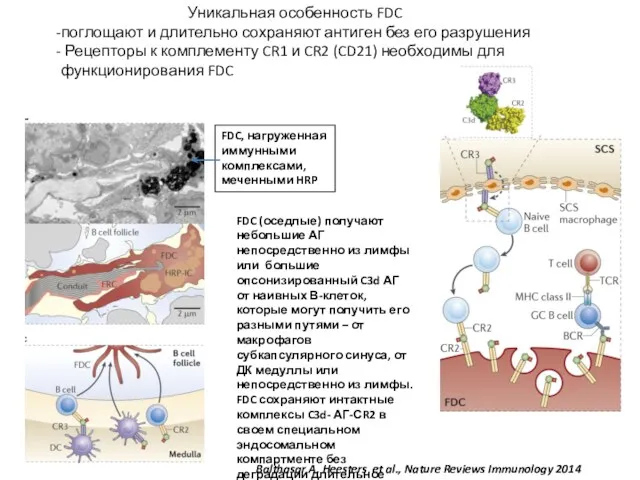
Balthasar A. Heesters et al., Nature Reviews Immunology 2014
Уникальная
Balthasar A. Heesters et al., Nature Reviews Immunology 2014
Уникальная
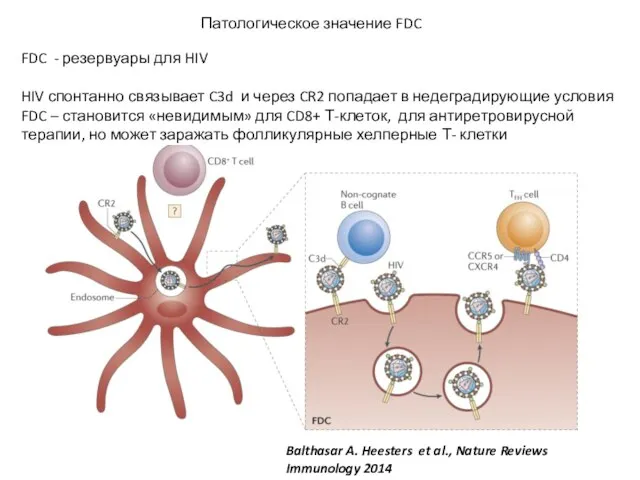
FDC - резервуары для HIV
HIV спонтанно связывает C3d и через CR2
FDC - резервуары для HIV
HIV спонтанно связывает C3d и через CR2
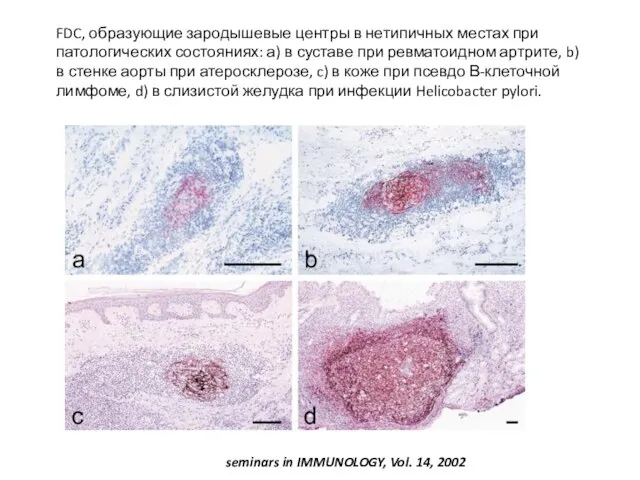
FDC, образующие зародышевые центры в нетипичных местах при патологических состояниях: а)
FDC, образующие зародышевые центры в нетипичных местах при патологических состояниях: а)
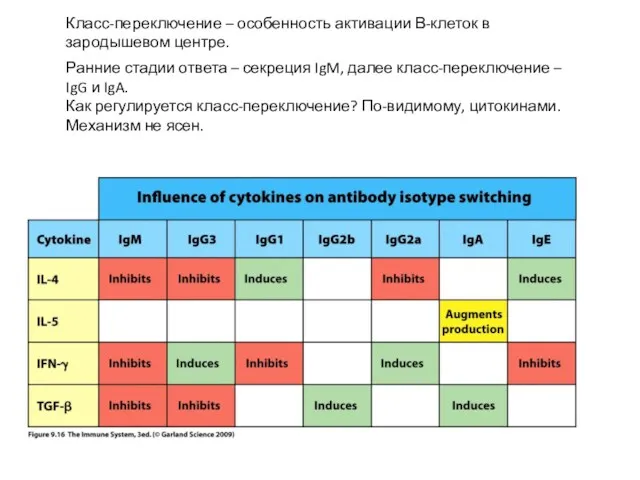
Ранние стадии ответа – секреция IgM, далее класс-переключение – IgG и
Ранние стадии ответа – секреция IgM, далее класс-переключение – IgG и
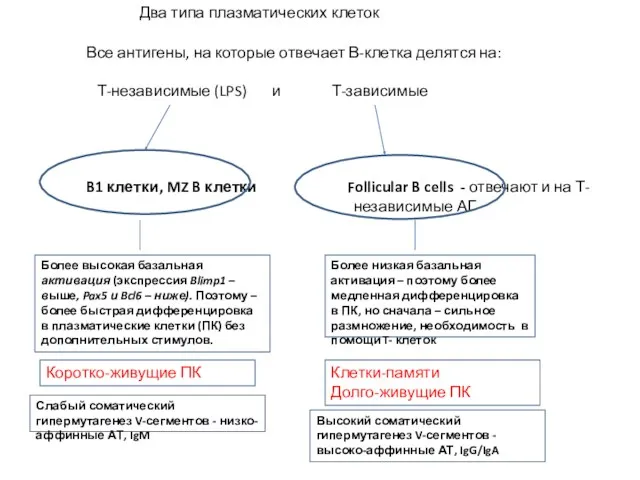
Все антигены, на которые отвечает В-клетка делятся на:
Т-независимые (LPS)
Все антигены, на которые отвечает В-клетка делятся на:
Т-независимые (LPS)
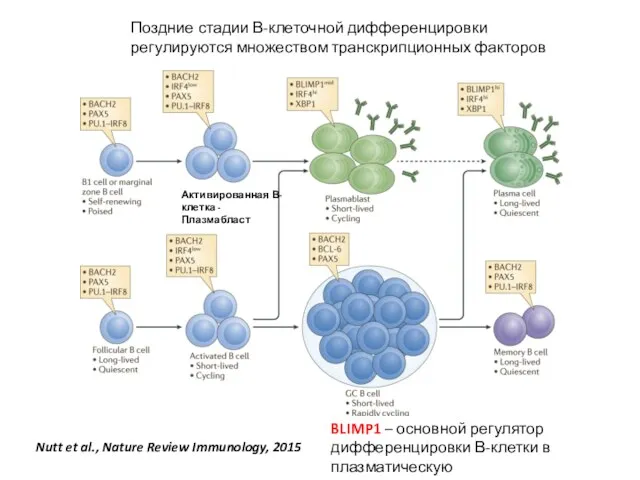
Поздние стадии В-клеточной дифференцировки регулируются множеством транскрипционных факторов
Nutt et al., Nature
Поздние стадии В-клеточной дифференцировки регулируются множеством транскрипционных факторов
Nutt et al., Nature
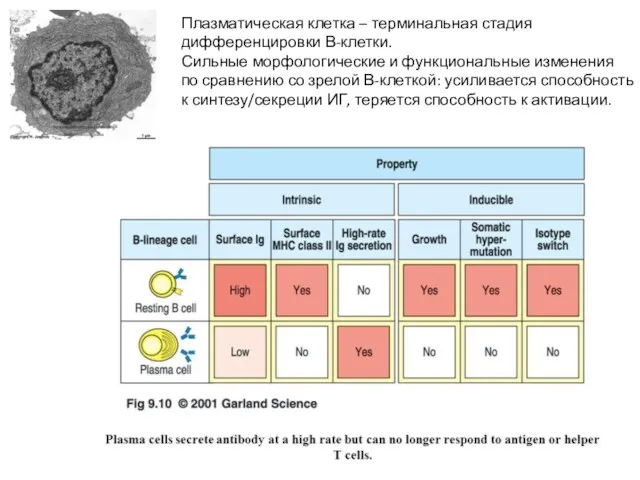
Плазматическая клетка – терминальная стадия дифференцировки В-клетки.
Сильные морфологические и функциональные изменения
Плазматическая клетка – терминальная стадия дифференцировки В-клетки.
Сильные морфологические и функциональные изменения
 Гигиенаның қалыптасуы және даму тарихы
Гигиенаның қалыптасуы және даму тарихы Ишемическая болезнь сердца
Ишемическая болезнь сердца Осложнения острого хоолецистита. Механическая желтуха
Осложнения острого хоолецистита. Механическая желтуха Молекулярно-генетические механизмы онтогенеза. Нарушения онтогенеза и их место в патологии человека
Молекулярно-генетические механизмы онтогенеза. Нарушения онтогенеза и их место в патологии человека Питание дома. Питание в школьной столовой. Поведение в школьной столовой
Питание дома. Питание в школьной столовой. Поведение в школьной столовой Балалардағы В 12 тапшылықты анемия
Балалардағы В 12 тапшылықты анемия Патофизиология липидного обмена. Атеросклероз
Патофизиология липидного обмена. Атеросклероз Наборы инстрементов на операции
Наборы инстрементов на операции Загальна характеристика вірусних гепатитів. Вірусні гепатити з фекально-оральним механізмом зараження
Загальна характеристика вірусних гепатитів. Вірусні гепатити з фекально-оральним механізмом зараження Эпидемиология как наука. Предмет, задачи и методы
Эпидемиология как наука. Предмет, задачи и методы Асептика и антисептика. История вопроса. Виды антисептики
Асептика и антисептика. История вопроса. Виды антисептики Строение тазобедренного сустава
Строение тазобедренного сустава Недоношенные новорожденные
Недоношенные новорожденные Сюрпризы погоды и их влияние на здоровье человека
Сюрпризы погоды и их влияние на здоровье человека Энцефалиты
Энцефалиты Ранняя реабилитация пациентов, перенесших инсульт. Роль медицинской сестры
Ранняя реабилитация пациентов, перенесших инсульт. Роль медицинской сестры Геморрагический васкулит
Геморрагический васкулит ДТЗ (Диффузно-токсический зоб)
ДТЗ (Диффузно-токсический зоб) Гиподинамия, гипердинамия
Гиподинамия, гипердинамия Энтеральные вирусные гепатиты
Энтеральные вирусные гепатиты Современные методы обследования больных с опухолями головы и шеи
Современные методы обследования больных с опухолями головы и шеи Пародонт аурулары. Түсінік, жүйесі, этиологиясы, патогенезі. Терминдер
Пародонт аурулары. Түсінік, жүйесі, этиологиясы, патогенезі. Терминдер Coagularea intravasculară diseminată
Coagularea intravasculară diseminată Диагностика варикозной болезни и облитерирующего атеросклероза вен нижних конечностей
Диагностика варикозной болезни и облитерирующего атеросклероза вен нижних конечностей Волосатоклеточный лейкоз
Волосатоклеточный лейкоз ОРВИ у детей
ОРВИ у детей Искусственный интеллект (ИИ) в медицине (примеры использования)
Искусственный интеллект (ИИ) в медицине (примеры использования) Заболевания, передающиеся половым путём
Заболевания, передающиеся половым путём