- Биологические мембраны и перенос вещества. Лекция 3

Содержание
- 2. Структура и физико-химические свойства биологических и искусственных мембран. Ленгмюровские монослои.. Явления переноса, активный и пассивный транспорт
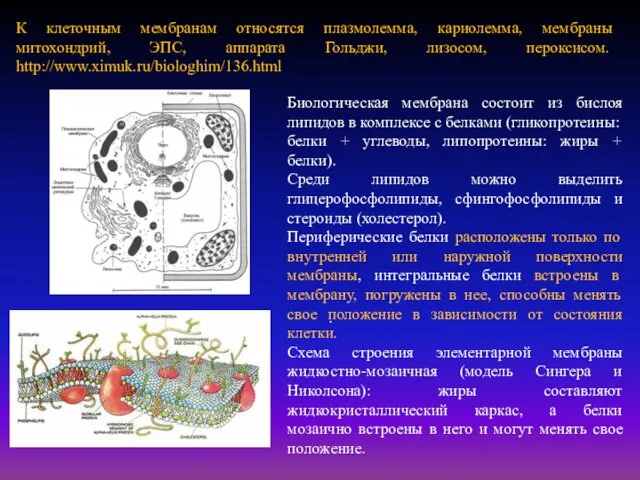
- 3. . К клеточным мембранам относятся плазмолемма, кариолемма, мембраны митохондрий, ЭПС, аппарата Гольджи, лизосом, пероксисом. http://www.ximuk.ru/biologhim/136.html Биологическая
- 4. Функции: 1) барьерная ; 2) структурная (придание определенной формы клеткам); 3) защитная (за счет избирательной проницаемости,
- 5. Структура липидов модульная —они состоят из головок, хвостов и промежуточных сегментов в разных сочетаниях, что обеспечивает
- 6. Химические формулы распространенных липидов биологических мембран: 1 -фосфатидилэтаноламин; 2 - фосфатидилсерин; 3 - фосфатидилинозит; 4 -
- 7. Распределение липидов между наружной (а) и внутренней (б) сторонами бислоя в мембранах эритроцитов (I), вируса гриппа
- 8. Липосомы — самопроизвольно образующиеся в смесях фосфолипида с водой замкнутые пузырьки. Диаметр липосом варьирует от 20
- 9. Ленгмюровские монослои Ванна и весы Ленгмюра для измерения поверхностного давления монослоя. Под действием внешней силы плавучий
- 10. Монослои амфифильных молекул можно перенести с поверхности воды на твердую подложку методом Ленгмюра – Блоджетт (вверху)
- 11. Барьерная роль плазмалеммы Гликокаликс представляет собой внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки
- 12. Барьерная роль плазмалеммы заключается также в ограничении свободной диффузии веществ. Модельные опыты на искусственных липидных мембранах
- 13. Простая диффузия неэлектролитов. Законы Фика. Вещества, перемещающиеся через мембрану путём свободной диффузии, не образуют каких-либо химических
- 14. Так как концентрационный градиент клеточной мембраны определить трудно, то для описания диффузии веществ через клеточные мембраны
- 15. Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационных и электрических градиентов. Липидный бислой мембраны
- 16. Явления переноса, активный и пассивный транспорт ионов Пассивный транспорт осуществляется в результате диффузии веществ в сторону
- 17. Клеточные мембраны проницаемы не только для веществ, диффундирующих через липидный слой, но и для многих ионов,
- 18. Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа. Способность ионного канала пропускать ионы
- 19. унипорт, когда переносится одно вещество в одном направлении (например, глюкоза в клетках печени или потенциал-зависимый натриевый
- 20. Вторично- активным транспортом называют системы транспорта через мембраны, которые переносят вещества из области их низкой концентрации
- 21. Один из самых интересных способов активного транспорта состоит в том, чтобы каким-либо образом удержать внутри клетки
- 22. (K+/Na+)-нacoc: 1 — участок связывания Na+; 2 — участок связывания К+; 3 — мембрана Существует несколько
- 23. Участие АТФаз в активном транспорте ионов через биологические мембраны. Опытным путем было установлено, что: а) Транспорт
- 24. Основные этапы работы Na+ K+ АТФаз таковы: 1. Присоединение снаружи двух ионов K+ и одной молекулы
- 25. Кальций-транспортная АТФаза - сравнительно небольшой белок, состоящий из одной полипептидной цепи. Он выполняет важнейшую функцию -
- 26. Поддержание низкой концентрации ионов кальция в цитоплазме покоящихся клеток создает возможность регуляции клеточных функций путем увеличения
- 27. Водородно-калиевая аденозинтрифосфатаза (Н+/К+-АТФа́за) В гастроэнтерологии и фармацевтике, ориентированной на органы пищеварения, вместо водородно-калиевая аденозинтрифосфатаза обычно используют
- 28. Схема укладки a- и b-субъединиц Н+, К+-АТФазы в липидном бислое (А) и схема, иллюстрирующая каталитический цикл
- 29. Выход Cl- из клеток обеспечивается градиентом электрохимического потенциала через каналы апикальной мембраны, которые активируются цАМФ; Н+
- 30. Эндоцитоз и экзоцитоз Макромолекулы - белки и нуклеиновые кислоты - не могут проникнуть через плазматическую мембрану
- 31. Схема процессов, включающих экзо- и эндоцитоз. А. Белок, синтезированный в гранулярном эндоплазматическом ретикулуме, транспортируется посредством аппарата
- 33. Скачать презентацию

Структура и физико-химические свойства биологических и искусственных мембран.
Ленгмюровские монослои..
Ленгмюровские монослои..
.
К клеточным мембранам относятся плазмолемма, кариолемма, мембраны митохондрий, ЭПС, аппарата Гольджи,
.
К клеточным мембранам относятся плазмолемма, кариолемма, мембраны митохондрий, ЭПС, аппарата Гольджи,

Функции:
1) барьерная ;
2) структурная (придание определенной формы клеткам);
3) защитная (за счет избирательной проницаемости, рецепции
Функции:
1) барьерная ;
2) структурная (придание определенной формы клеткам);
3) защитная (за счет избирательной проницаемости, рецепции
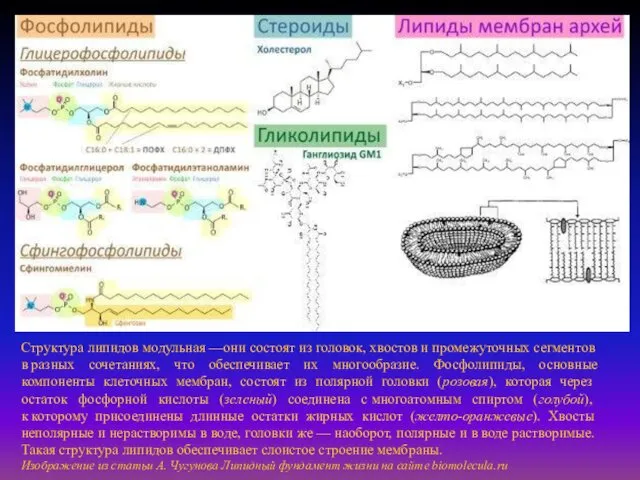
Структура липидов модульная —они состоят из головок, хвостов и промежуточных сегментов в разных
Структура липидов модульная —они состоят из головок, хвостов и промежуточных сегментов в разных
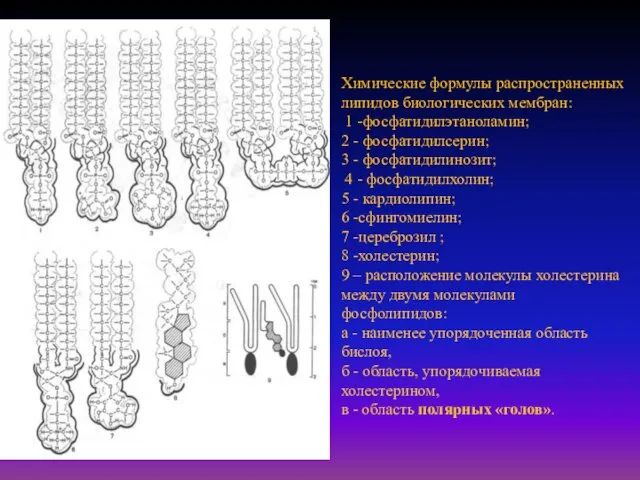
Химические формулы распространенных липидов биологических мембран:
1 -фосфатидилэтаноламин;
2 - фосфатидилсерин;
Химические формулы распространенных липидов биологических мембран:
1 -фосфатидилэтаноламин;
2 - фосфатидилсерин;
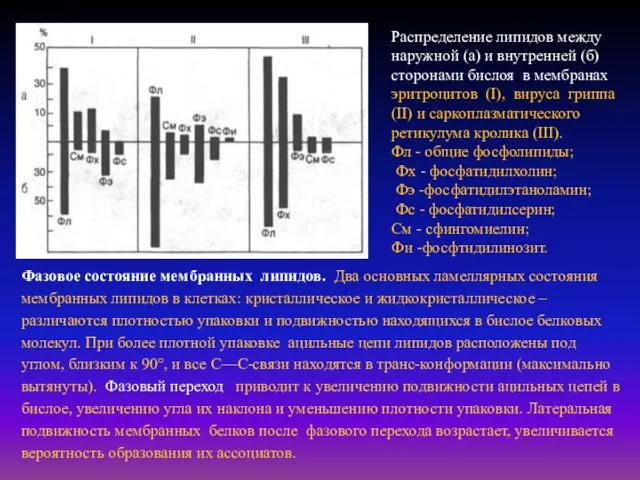
Распределение липидов между наружной (а) и внутренней (б) сторонами бислоя в
Распределение липидов между наружной (а) и внутренней (б) сторонами бислоя в

Липосомы — самопроизвольно образующиеся в смесях фосфолипида с водой замкнутые пузырьки. Диаметр
Липосомы — самопроизвольно образующиеся в смесях фосфолипида с водой замкнутые пузырьки. Диаметр
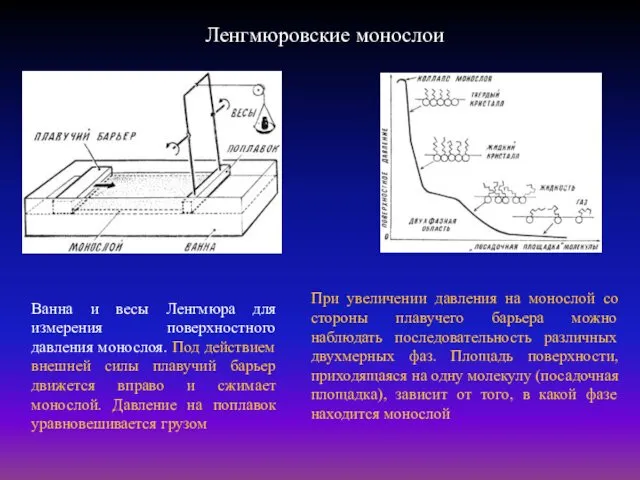
Ленгмюровские монослои
Ванна и весы Ленгмюра для измерения поверхностного давления монослоя. Под
Ленгмюровские монослои
Ванна и весы Ленгмюра для измерения поверхностного давления монослоя. Под

Монослои амфифильных молекул можно перенести с поверхности воды на твердую подложку
Монослои амфифильных молекул можно перенести с поверхности воды на твердую подложку
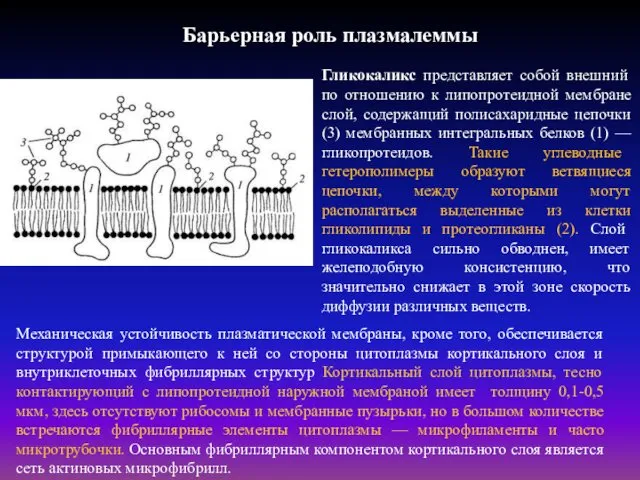
Барьерная роль плазмалеммы
Гликокаликс представляет собой внешний по отношению к липопротеидной мембране
Барьерная роль плазмалеммы
Гликокаликс представляет собой внешний по отношению к липопротеидной мембране

Барьерная роль плазмалеммы заключается также в ограничении свободной диффузии веществ. Модельные
Барьерная роль плазмалеммы заключается также в ограничении свободной диффузии веществ. Модельные

Простая диффузия неэлектролитов. Законы Фика.
Вещества, перемещающиеся через мембрану путём свободной диффузии,
Простая диффузия неэлектролитов. Законы Фика.
Вещества, перемещающиеся через мембрану путём свободной диффузии,

Так как концентрационный градиент клеточной мембраны определить трудно, то для описания
Так как концентрационный градиент клеточной мембраны определить трудно, то для описания

Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационных и
Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационных и
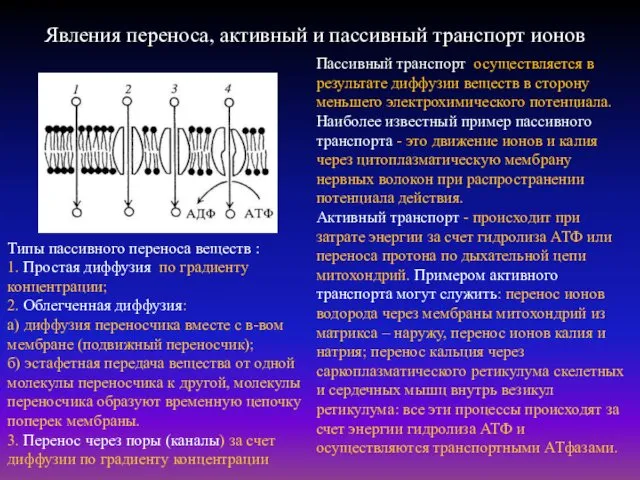
Явления переноса, активный и пассивный транспорт ионов
Пассивный транспорт осуществляется в результате
Явления переноса, активный и пассивный транспорт ионов
Пассивный транспорт осуществляется в результате
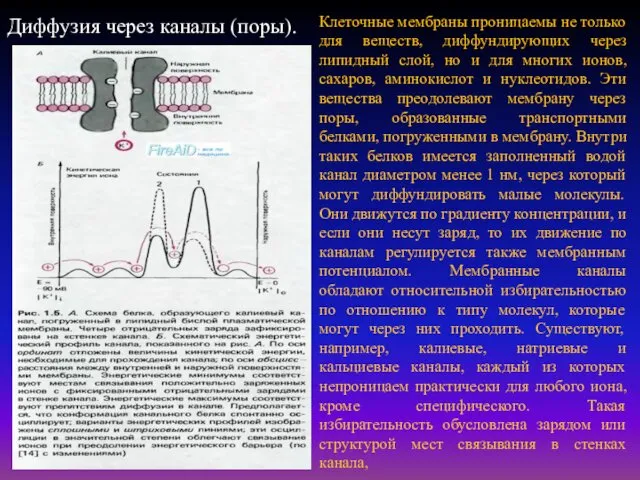
Клеточные мембраны проницаемы не только для веществ, диффундирующих через липидный слой,
Клеточные мембраны проницаемы не только для веществ, диффундирующих через липидный слой,

Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.
Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.

унипорт, когда переносится одно вещество в одном направлении (например, глюкоза в
унипорт, когда переносится одно вещество в одном направлении (например, глюкоза в

Вторично- активным транспортом называют системы транспорта через мембраны, которые переносят вещества
Вторично- активным транспортом называют системы транспорта через мембраны, которые переносят вещества

Один из самых интересных способов активного транспорта состоит в том, чтобы
Один из самых интересных способов активного транспорта состоит в том, чтобы

(K+/Na+)-нacoc:
1 — участок связывания Na+;
2 — участок связывания К+;
(K+/Na+)-нacoc:
1 — участок связывания Na+;
2 — участок связывания К+;

Участие АТФаз в активном транспорте ионов через биологические мембраны.
Опытным путем
Участие АТФаз в активном транспорте ионов через биологические мембраны.
Опытным путем

Основные этапы работы Na+ K+ АТФаз таковы:
1. Присоединение снаружи двух ионов
Основные этапы работы Na+ K+ АТФаз таковы:
1. Присоединение снаружи двух ионов

Кальций-транспортная АТФаза - сравнительно небольшой белок, состоящий из одной полипептидной цепи.
Кальций-транспортная АТФаза - сравнительно небольшой белок, состоящий из одной полипептидной цепи.

Поддержание низкой концентрации ионов кальция в цитоплазме покоящихся клеток создает возможность
Поддержание низкой концентрации ионов кальция в цитоплазме покоящихся клеток создает возможность

Водородно-калиевая аденозинтрифосфатаза (Н+/К+-АТФа́за)
В гастроэнтерологии и фармацевтике, ориентированной на органы пищеварения, вместо
Водородно-калиевая аденозинтрифосфатаза (Н+/К+-АТФа́за)
В гастроэнтерологии и фармацевтике, ориентированной на органы пищеварения, вместо
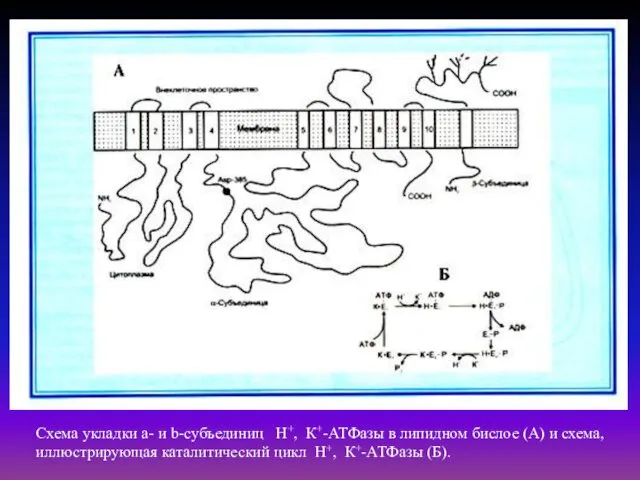
Схема укладки a- и b-субъединиц Н+, К+-АТФазы в липидном бислое (А)
Схема укладки a- и b-субъединиц Н+, К+-АТФазы в липидном бислое (А)

Выход Cl- из клеток обеспечивается градиентом электрохимического потенциала через каналы апикальной
Выход Cl- из клеток обеспечивается градиентом электрохимического потенциала через каналы апикальной

Эндоцитоз и экзоцитоз
Макромолекулы - белки и нуклеиновые кислоты - не могут
Эндоцитоз и экзоцитоз
Макромолекулы - белки и нуклеиновые кислоты - не могут
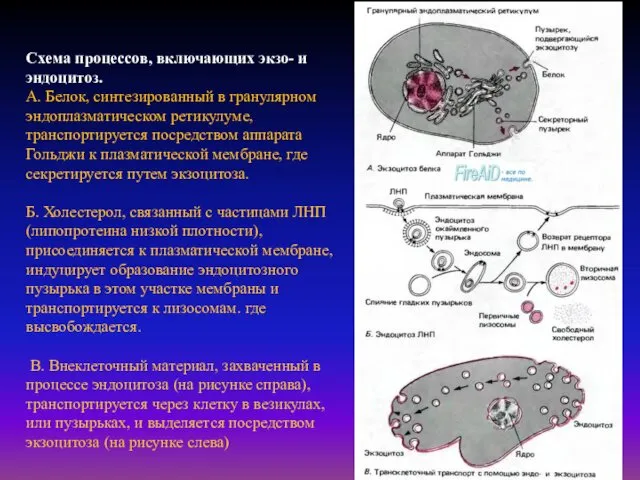
Схема процессов, включающих экзо- и эндоцитоз.
А. Белок, синтезированный в гранулярном
Схема процессов, включающих экзо- и эндоцитоз.
А. Белок, синтезированный в гранулярном
 fvkhfykkhvpyschdyvamyvzham
fvkhfykkhvpyschdyvamyvzham Новое качество образования
Новое качество образования Обработка информации с помощью текстовых процессоров
Обработка информации с помощью текстовых процессоров Отчет о проведении Публичной презентации результатов педагогической деятельности и инновационной работы - 2014.
Отчет о проведении Публичной презентации результатов педагогической деятельности и инновационной работы - 2014. Многонациональный Крым
Многонациональный Крым Презентация Предметно-развивающая среда группы раннего возраста
Презентация Предметно-развивающая среда группы раннего возраста Рентгенодиагностика и дифференциальная диагностика частных форм туберкулеза. Туберкулёма
Рентгенодиагностика и дифференциальная диагностика частных форм туберкулеза. Туберкулёма Производство сильфонных компенсаторов
Производство сильфонных компенсаторов Лингвистический анализ художественного текста
Лингвистический анализ художественного текста Резьбы
Резьбы Г. Ибсен Кукольный дом
Г. Ибсен Кукольный дом Транспорт. Какие величины характеризуют движение?
Транспорт. Какие величины характеризуют движение? Смесительный полупроводниковый диод
Смесительный полупроводниковый диод Образование Верещагинского района: вчера, сегодня, завтра. Виртуальный музей
Образование Верещагинского района: вчера, сегодня, завтра. Виртуальный музей Технология конструкционных материалов. Оптимальная точность обработки
Технология конструкционных материалов. Оптимальная точность обработки Проектная деятельность в старших классах 25.09.2018
Проектная деятельность в старших классах 25.09.2018 Музейный урок Быт и культура Кубани
Музейный урок Быт и культура Кубани Ильменский заповедник
Ильменский заповедник Города воинской славы
Города воинской славы Деньги и их функции
Деньги и их функции Особенности проведения деловых переговоров в Японии
Особенности проведения деловых переговоров в Японии Сепсис. Этиология
Сепсис. Этиология Участие в областном конкурсе Учитель года 2016. Методический семинар.
Участие в областном конкурсе Учитель года 2016. Методический семинар. Я-гражданин России.
Я-гражданин России. Обучение детей с расстройствами аутистического спектра: создание ресурсного класса
Обучение детей с расстройствами аутистического спектра: создание ресурсного класса Лапароскопия в детской хирургии
Лапароскопия в детской хирургии Интеллектуальная игра для 1 класса Хочу всё знать.
Интеллектуальная игра для 1 класса Хочу всё знать. На основе дипломной работы Повышение здоровьесозидающего потенциала уроков химии в школе продолжается работа по разработке уроков, с использованием детских исследовательских проектов.
На основе дипломной работы Повышение здоровьесозидающего потенциала уроков химии в школе продолжается работа по разработке уроков, с использованием детских исследовательских проектов.