- Биотехнология в селекции растений. Часть 5. Селекция на устойчивость к болезням

Содержание
- 2. Большое количество защитных генов растений Большинство кодируют устойчивость лишь к ограниченному кругу патогенов или даже расам
- 3. ДЕФЕНЗИНЫ РАСТЕНИЙ небольшие (около 5 kD) пептиды обычно основные богаты цистеиновыми остатками связаны дисульфидными мостиками экспрессируются
- 4. БЕЛКОВЫЕ ИНГИБИТОРЫ ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ - локализованы в межклеточном пространстве и клеточной стенке - являются регуляторами эндогенных
- 5. Протеолиз является одним из важнейших процессов жизнедеятельности как животных, так и растений. Протеолитические ферменты участвуют в
- 6. Фитопатогенные микроорганизмы используют протеиназы в качестве атакующего агента. Растения выработали различные защитные механизмы Неотъемлемыми компонентами являются
- 7. ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ - участвуют в регуляции внутриклеточных процессов - играют активную роль в передаче внутриклеточных сигналов
- 8. Участие протеиназ в регуляции внутриклеточных процессов Могут играть активную роль в передаче внутриклеточных сигналов, процессинге и
- 10. Вакуолярная протеиназа – один из ферментов, осуществляющих процессинг запасных белков у ряда растений специфически расщепляет в
- 11. В плазматической мембране листьев томатов присутствует белок, который избирательно связывает пептидный гормон системин. Связывание сопровождается гидролизом
- 12. Участие протеиназ во взаимоотношениях растение-патоген Была обнаружена системная индукция синтеза протеолитических ферментов в ответ на повреждение
- 13. Фитопатогенные микроорганизмы наряду с другими ферментами продуцируют активные экстрацеллюлярные протеазы Корреляция между активностью экстрацеллюлярных протеаз фитопатогена
- 14. Реакция сверхчувствительности при взаимодействии растений с несовместимыми расами фитопатогенных микроорганизмов Напоминает апоптоз у животных Гибель клеток
- 15. Особую группу составляют сериновые протеиназы, вовлеченные в защитные механизмы при поражении растений фитопатогенными микроорганизмами, а также
- 16. ИНГИБИТОРЫ ПРОТЕИНАЗ РАСТЕНИЙ Белковые ингибиторы протеолитических ферментов - являются регуляторами эндогенных протеаз - широко представлены в
- 17. Локализация и индукция действия ингибиторов протеиназ растений Характерной особенностью всех белков, связанных с патогенезом, в том
- 18. Секреция ингибиторов протеиназ в экстрацеллюларное пространство происходит в процессе прорастания семян. Так, ингибиторы трипсина и химотрипсина
- 19. Действие грибных и бактериальных протеиназ на растения индуцирует ответ растительной клетки, что, прежде всего, связано с
- 20. Способность к образованию PIIF в ответ на повреждение была обнаружена у представителей семейств Fabaceae, Cucurbitaceae и
- 21. Другим полипептидом, индуцирующим синтез ингибиторов протеиназ, является системин, выделенный из плодов и листьев томатов Установлена полная
- 23. Предполагается, что существуют по крайней мере два различных пути распространения сигнала. Первый путь - при поранении
- 24. Второй путь реализуется при действии на растение фитoпaтoгeнныx микроорганизмов Индукция олигоуронидами, в том числе и PIIF,
- 25. Участие ингибиторов протеиназ в регуляции активности внутриклеточных ферментов растений Ингибиторы протеолитических ферментов принимают участие в регуляции
- 26. Во многих случаях ингибиторы, действующие на эндогенные протеиназы растений, отличаются от ингибиторов трипсина, химотрипсина и протеиназ
- 27. Ингибиторы протеиназ являются полифункциональными Классический ингибитор сериновых протеиназ сои STI способен подавлять активность цистеиновых протеиназ Из
- 28. Ингибиторы протеиназ в системе защиты растений от биотических стрессов Механизмы действия ингибиторов протеиназ при защите от
- 29. Ингибиторы протеиназ в системе защиты растений от насекомых В середине 60-х годов из семян сои и
- 30. Многие белки растительного происхождения, первоначально описанные как ингибиторы трипсина и химотрипсина, являются эффективными ингибиторами протеиназ, содержащихся
- 31. Модифицированный ген ингибитора ингибиторе трипсина из коровьего гороха (Vigna unguiculata) был перенесен в растение табака и
- 32. Ингибиторы протеиназ в системе защиты растений от патогенных микроорганизмов Многие белки-ингибиторы протеаз растительного происхождения способны подавлять
- 33. Подавление протеолитической активности может приводить к нарушению способности утилизировать растительные белки и соответственно к угнетению роста
- 34. Белковые ингибиторы протеиназ из семян и листьев разных видов люпина подавляли активность протеаз рода Fusarium, задерживали
- 35. В тканях картофеля и томатов присутствуют ингибитор I и ингибитор II, биосинтез которых кодируется различными негомологичными
- 36. У растений обнаружены белки, обладающие специфической ингибиторной активностью против протеиназ микроорганизмов: у бобовых (фасоль обыкновенная, коровий
- 37. Белок-ингибитор, действующий на микробные протеиназы и не активный по отношению к трипсину и собственным протеиназам, специфически
- 38. Ингибиторы цистеиновых протеиназ из растений также способны подавлять рост и развитие фитопатогенных грибов, например, Bot. cynerea
- 39. Индукция синтеза ингибиторов протеиназ наблюдалась и при бактериальной инфекции. Инокуляция листьев томатов патогенной бактерией Ps. syringae
- 40. Из семян одной из разновидностей проса (Pennisetum glaucum) был выделен ингибитор цистеиновых протеиназ, который подавлял рост
- 41. В последние годы появились данные, указывающие на то, что ингибиторы протеиназ могут играть существенную роль также
- 42. Характеристика белковых ингибиторов сериновых протеиназ гречихи Большинство известных и исследованных в настоящее время ингибиторов протеиназ из
- 43. В экспериментах установлено, что аминокислотный составы трех очищенных анионных ингибиторов протеиназ гречихи (BWI-1a, BWI-2a и BWI-4a)
- 44. Анализ аминокислотной последовательности ингибитора BWI-1a показал, что она на 65 % идентична последовательности ингибитора трипсина и
- 45. Влияние ингибиторов BWI-1а, BWI-2а и BWI-4a на активности различных протеолитических ферментов
- 46. Показано, что BWI-1а и BWI-2а кроме активности трипсина ингибируют также активность химотрипсина, хотя и с меньшей
- 47. Влияние BWI-1а из семян гречихи на рост мицелия фитопатогенных грибов Изучение влияния исследованных ингибиторов на рост
- 48. Сравнение констант ингибирования сериновых протеиназ ингибиторами протеаз из семян гречихи и других растительных источников (по Цыбиной,
- 49. ДЕФЕНЗИНЫ РАСТЕНИЙ Растительные дефензины принадлежат к большому семейству антимикробных пептидов с цистеин-стабилизированным αβ-мотивом Достаточно широко представлены
- 50. Структура дефензинов растений Дефензины растений – это семейство небольших (около 5 kD), обычно основных, пептидов, богатых
- 51. Сравнение последовательностей растительных дефензинов и растительных дефензин-подобных протеинов. Цистеины обозначены рамкой. Цистеин-стабилизирующие отпечатки α-спирали обозначены звездочками
- 52. При этом были выявлены консервативные положения восьми цистеинов, двух глицинов в позициях 13 и 34, серина
- 53. Локализация и предполагаемый механизм действия дефензинов растений Было показано, что дефензины растений экспрессируются почти в каждом
- 54. Методами иммунологической или in situ-гибридизации была показана локализация и аккумуляция конститутивно экспрессируемых дефензинов растений преимущественно в
- 55. По крайней мере, на четырех видах растений, а именно в горохе, табаке, редисе и арабидопсисе, было
- 56. Было установлено, что экспрессию дефензина арабидопсиса PDF1.2 индуцирует экзогенное применение или метилжасмоната (MeJA), или этилена. Измерение
- 57. Дефензины редьки Rs-AFP2 и георгина Dm-AMP1 не образовывали ион-проникающих пор в искусственных фосфолипидных мембранах Растительный дефензин
- 58. Один из предполагаемых механизмов действия состоит в том, что мембранный рецептор позволяет растительным дефензинам встраиваться в
- 59. Классификация дефензинов растений Морфогенные дефензины растений приводят к редукции роста гифов и одновременному увеличению числа ветвей,
- 60. Напротив, дефензины, выделенные из Dahlia merckii, Clitoria ternatea, Aesculus hippocastanum, относящиеся к неморфогенному типу, ингибируют элонгацию
- 61. Фунгицидная активность дефензинов растений Анализ антимикробной активности различных дефензинов показал, что область от cys-27 до cys-47
- 62. При использовании точечного мутагенеза было показано, что замена нейтральных остатков gly-9 и val-39 заряженным аргинином (G9R
- 63. Фунгицидная активность как морфогенных, так и неморфогенных дефензинов растений уменьшается с увеличением ионной силы в среде
- 64. Фунгицидная активность различных пептидов
- 65. Бактерицидная активность дефензинов растений Антибактериальные активности пептидов были измерены на четырех грам-положительных (Bacillus subtilis, Micrococcus luteus,
- 66. Вклад дефензинов в защиту растения-хозяина Дефензины играют важную роль в защите семян или всходов от проникновения
- 67. Растения табака были протрансформированы химерной генной конструкцией, включающей кодирующую область гена Rs-AFP2 дефензина редьки под контролем
- 68. Болезни картофеля Картофель относится к числу культур, в сильной степени поражаемых болезнями и вредителями, которые нередко
- 69. Фитофтороз Возбудитель - гриб Phytophthora infestans D.В. Первые признаки - на нижних листьях по краям листа
- 70. Ризоктониоз (белая ножка, черная парша клубней) Возбудитель - гриб Rhizoctonia solani Kuhn. На растениях образуются глубокие
- 71. Фузариозное увядание, сухая гниль Возбудитель - гриб Fusarium spp. На растениях нижняя часть стебля буреет, при
- 72. Парша обыкновенная Возбудитель - актиномицеты Streptomyces scabies Waks et Henr. и другие виды. На поверхности клубней,
- 73. Рак картофеля Возбудитель - гриб Sinchytrium endobioticum (Shilb). Проявляется в виде различного рода разрастании тканей (наростов).
- 74. Черная ножка и бактериальная мягкая (мокрая) гниль Возбудитель - бактерии Erwinia caratovora. var. atroseptica и Erwinia
- 75. Мягкая бактериальная гниль поражает клубни. В месте прикрепления клубня к столону отмечается размягчение бесцветного или светло-желтого
- 76. Кольцевая гниль Возбудитель - бактерии Clavibacter michiganensis ssp. sepedonicus. Поражаются растения и клубни. Развитие кольцевой гнили
- 77. Крапчатая (обыкновенная, мягкая) мозаика картофеля Возбудитель - Х-вирус картофеля. Проявляется в виде расплывчатых светло-зеленных пятен на
- 78. Морщинистая и полосчатая мозаики. Возбудитель - Y-вирус картофеля. Различают три основных штамма вируса Yo, Yn, Yc.
- 79. Скручивание листьев картофеля Возбудитель - L-вирус картофеля (Вирус скручивания листьев картофеля - ВСЛК). В год заражения
- 80. Ведьмина метла картофеля Возбудитель - фитоплазма. Куст становится низкорослым, с большим количеством измельченных тонких, округлого сечения
- 81. Бронзовость листьев Возникает при недостаточном поступлении калия в растения. Листья покрываются мелкими точками отмершей ткани, становятся
- 82. Коричневая пятнистость стеблей и клубней Причина возникновения - избыточное поступление в растение железа и алюминия. На
- 83. Картофельная нематода (бледная и золотистая) Globodera rostochiensis и Globodera pallida. На пораженных почвах вырастают малорослые, слабые
- 84. Стеблевая нематода Ditylenchus destructor. Видимые признаки в период вегетации отсутствуют. Может вызывать деформацию кустов, изогнутость стеблей.
- 85. Галловая нематода Meloidogyne marioni. Пораженные растения отстают в росте и увядают. На корневой системе образуются небольшие
- 86. Генетическая инженерия картофеля В сочетании с современными технологиями возделывания различные виды пестицидов способны значительно сократить потери
- 87. Картофель, наряду с другим известным представителем семейства Solanaceae Nicotiana tabacum был одним из первых растений, подвергшихся
- 88. Основные факторы, влияющие на эффективность генетической модификации растений
- 89. Основным фактором, определяющим эффективность получения трансгенных растений, является восприимчивость эксплантов конкретного сорта к трансформации и способность
- 90. При селекции картофеля в полевых условиях необходимо провести оценку 50-100 тысяч сеянцев, то in vitro можно
- 92. Использование патогенов в клеточной селекции Непосредственное культивирование в присутствии патогена, если мало известно о патогенном начале
- 93. Некоторые грибы растут очень быстро, что не позволяет идентифицировать устойчивые клетки С середины 60-х гг. Изучение
- 94. Использование патотоксинов в клеточной селекции Большинство штаммов грибов производит микотоксины и поражает широкий круг растений Применение
- 95. Патотоксины: 1 категория – токсины, не являющиеся определяющими в заболевании, но обладающие неспецифическим токсическим действием по
- 96. Выбор схемы селекции зависит от механизма действия: Одни патогены поражают клетки, затем выделяют токсин Другие выделяют
- 97. Получены устойчивые формы гвоздики, томатов, люцерны, клевера к фузариозу путем культивирования изолированных клеток, тканей и органов
- 98. Для ряда культур отбор с использованием селективного фактора фузариевой кислоты (5-н-бутил-2-пиридин-карбоновая кислота), продуцируемого различными фузариумами. Японские
- 99. ВНИИ льна – лен-долгунец – изолированные пыльники и каллусная ткань – высокая комплексная устойчивость к фузариозу
- 100. Использование культурального фильтрата патогена в клеточной селекции Следует учитывать: - способ получения КФ - агрессивность штаммов
- 101. Фузариоз на томатах ступенчатый отбор – 2n=48, 2n=64, прямой отбор – 2n=32 Клевер, люцерна (10 %
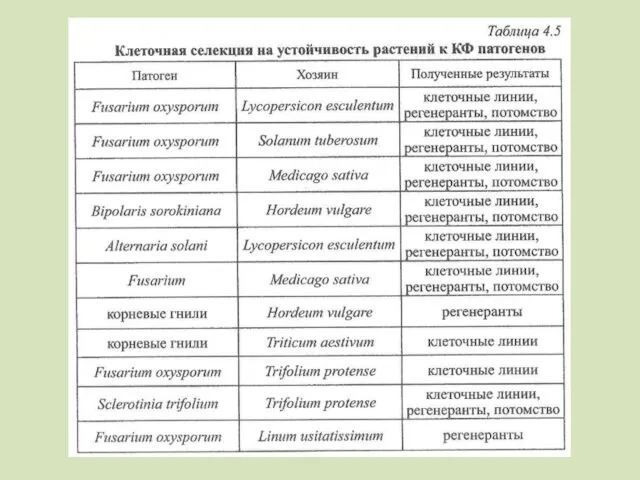
- 102. Сельдерей – септориоз – каллусные и суспензионные культуры из молодых черешков Важно изучение корреляции устойчивости к
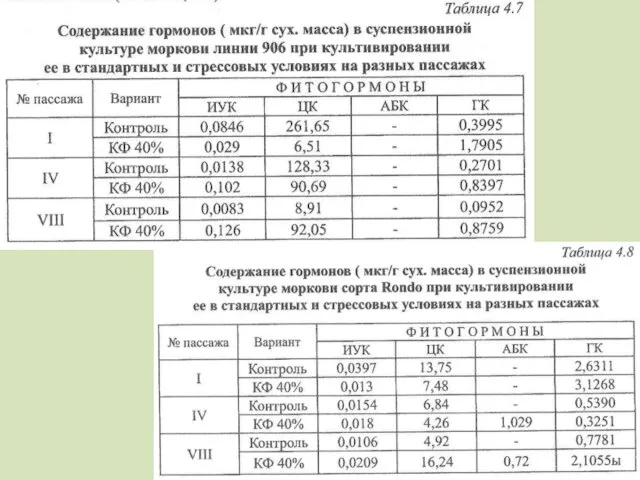
- 104. Участие фитогормонов в клеточной селекции Под действием факторов биотической природы происходит быстрый синтез и интенсивное накопление
- 105. Синтез эндогенной ГК увеличивается с увеличение концентрации КФ, ИУК и цитокининов – снижается (более крупные и
- 107. Полученные в результате селекции растения имели больше ауксинов, гиббереллинов и цитокининов В адаптации играют роль АБК
- 108. Устойчивость против проникновения фитопатогенов Накопление и отложение лигнина в местах внедрения патогена. Лигнифицированные ткани устойчивы к
- 109. Соотношение ИУК/АБК – величина постоянная у более устойчивых сортов (Дезире, Жуковский ранний)
- 110. В стандартных условиях уменьшение цитокининов, при стрессе – увеличение, особенно у более устойчивых сортов Утолщение клеточных
- 111. Участие фенольных соединений Механизм защиты и адаптации Поражение патогенами активизирует синтез фенольных соединений, в том числе
- 113. Скачать презентацию

Большое количество защитных генов растений
Большинство кодируют устойчивость лишь к ограниченному кругу
Большое количество защитных генов растений
Большинство кодируют устойчивость лишь к ограниченному кругу

ДЕФЕНЗИНЫ РАСТЕНИЙ
небольшие (около 5 kD) пептиды
обычно основные
богаты цистеиновыми остатками
связаны дисульфидными мостиками
экспрессируются
ДЕФЕНЗИНЫ РАСТЕНИЙ
небольшие (около 5 kD) пептиды
обычно основные
богаты цистеиновыми остатками
связаны дисульфидными мостиками
экспрессируются
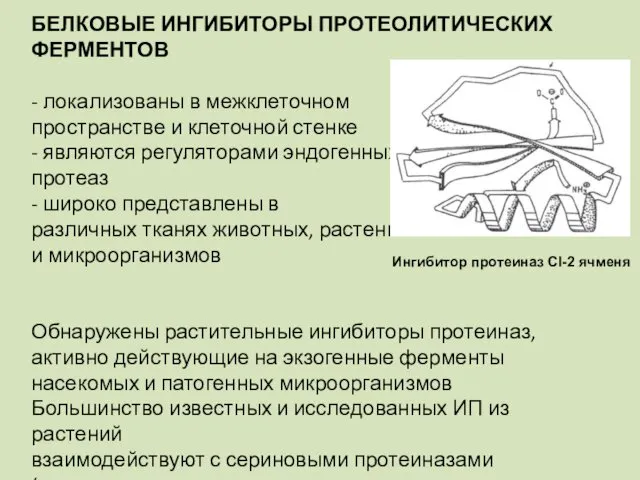
БЕЛКОВЫЕ ИНГИБИТОРЫ ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
- локализованы в межклеточном
пространстве и клеточной стенке
-
БЕЛКОВЫЕ ИНГИБИТОРЫ ПРОТЕОЛИТИЧЕСКИХ ФЕРМЕНТОВ
- локализованы в межклеточном
пространстве и клеточной стенке
-

Протеолиз является одним из важнейших процессов жизнедеятельности как животных, так и
Протеолиз является одним из важнейших процессов жизнедеятельности как животных, так и
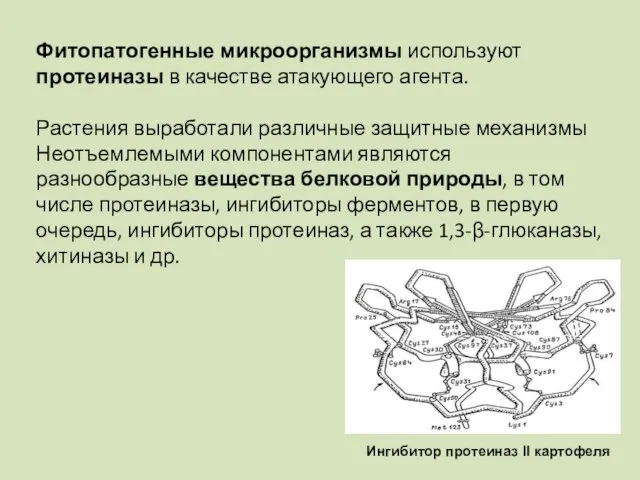
Фитопатогенные микроорганизмы используют протеиназы в качестве атакующего агента.
Растения выработали различные
Фитопатогенные микроорганизмы используют протеиназы в качестве атакующего агента.
Растения выработали различные

ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
- участвуют в регуляции внутриклеточных процессов
- играют активную роль в
ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
- участвуют в регуляции внутриклеточных процессов
- играют активную роль в

Участие протеиназ в регуляции внутриклеточных процессов
Могут играть активную роль в передаче
Участие протеиназ в регуляции внутриклеточных процессов
Могут играть активную роль в передаче

Вакуолярная протеиназа – один из ферментов, осуществляющих процессинг запасных белков у
Вакуолярная протеиназа – один из ферментов, осуществляющих процессинг запасных белков у

В плазматической мембране листьев томатов присутствует белок, который избирательно связывает пептидный
В плазматической мембране листьев томатов присутствует белок, который избирательно связывает пептидный

Участие протеиназ во взаимоотношениях растение-патоген
Была обнаружена системная индукция синтеза протеолитических ферментов
Участие протеиназ во взаимоотношениях растение-патоген
Была обнаружена системная индукция синтеза протеолитических ферментов

Фитопатогенные микроорганизмы наряду с другими ферментами продуцируют активные экстрацеллюлярные протеазы
Корреляция
Фитопатогенные микроорганизмы наряду с другими ферментами продуцируют активные экстрацеллюлярные протеазы
Корреляция

Реакция сверхчувствительности при взаимодействии растений с несовместимыми расами фитопатогенных микроорганизмов
Напоминает апоптоз
Реакция сверхчувствительности при взаимодействии растений с несовместимыми расами фитопатогенных микроорганизмов
Напоминает апоптоз

Особую группу составляют сериновые протеиназы, вовлеченные в защитные механизмы при поражении
Особую группу составляют сериновые протеиназы, вовлеченные в защитные механизмы при поражении

ИНГИБИТОРЫ ПРОТЕИНАЗ РАСТЕНИЙ
Белковые ингибиторы протеолитических ферментов
- являются регуляторами эндогенных протеаз
ИНГИБИТОРЫ ПРОТЕИНАЗ РАСТЕНИЙ
Белковые ингибиторы протеолитических ферментов
- являются регуляторами эндогенных протеаз

Локализация и индукция действия ингибиторов протеиназ растений
Характерной особенностью всех белков, связанных
Локализация и индукция действия ингибиторов протеиназ растений
Характерной особенностью всех белков, связанных

Секреция ингибиторов протеиназ в экстрацеллюларное пространство происходит в процессе прорастания семян.
Секреция ингибиторов протеиназ в экстрацеллюларное пространство происходит в процессе прорастания семян.

Действие грибных и бактериальных протеиназ на растения индуцирует ответ растительной клетки,
Действие грибных и бактериальных протеиназ на растения индуцирует ответ растительной клетки,

Способность к образованию PIIF в ответ на повреждение была обнаружена у
Способность к образованию PIIF в ответ на повреждение была обнаружена у

Другим полипептидом, индуцирующим синтез ингибиторов протеиназ, является системин, выделенный из плодов
Другим полипептидом, индуцирующим синтез ингибиторов протеиназ, является системин, выделенный из плодов
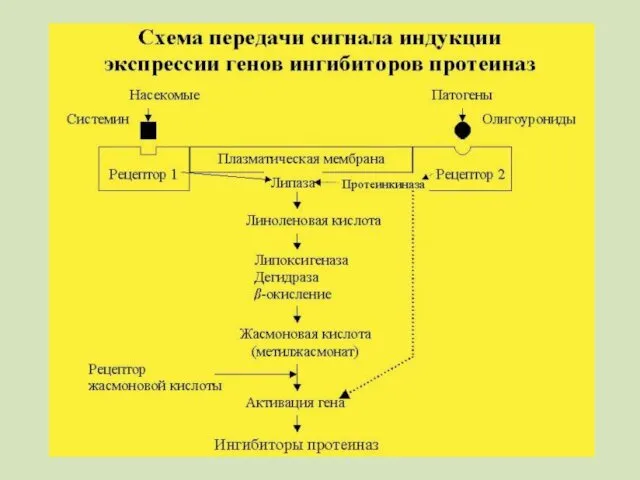

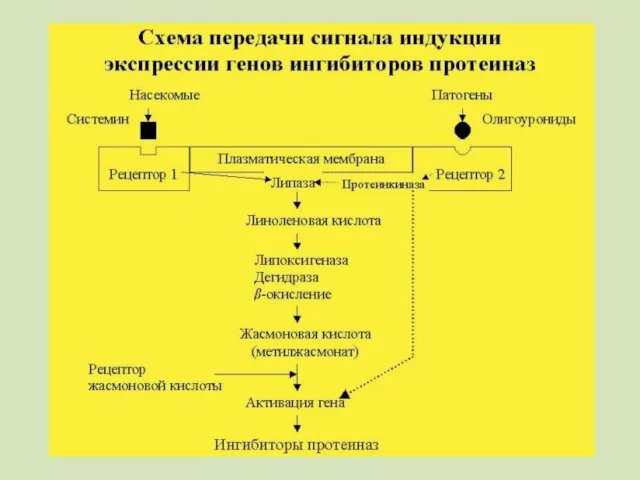
Предполагается, что существуют по крайней мере два различных пути распространения сигнала.
Предполагается, что существуют по крайней мере два различных пути распространения сигнала.

Второй путь реализуется при действии на растение фитoпaтoгeнныx микроорганизмов
Индукция олигоуронидами,
Второй путь реализуется при действии на растение фитoпaтoгeнныx микроорганизмов
Индукция олигоуронидами,

Участие ингибиторов протеиназ в регуляции активности внутриклеточных ферментов растений
Ингибиторы протеолитических ферментов
Участие ингибиторов протеиназ в регуляции активности внутриклеточных ферментов растений
Ингибиторы протеолитических ферментов

Во многих случаях ингибиторы, действующие на эндогенные протеиназы растений, отличаются от
Во многих случаях ингибиторы, действующие на эндогенные протеиназы растений, отличаются от

Ингибиторы протеиназ являются полифункциональными
Классический ингибитор сериновых протеиназ сои STI способен подавлять
Ингибиторы протеиназ являются полифункциональными
Классический ингибитор сериновых протеиназ сои STI способен подавлять

Ингибиторы протеиназ в системе защиты растений от биотических стрессов
Механизмы действия ингибиторов
Ингибиторы протеиназ в системе защиты растений от биотических стрессов
Механизмы действия ингибиторов

Ингибиторы протеиназ в системе защиты растений от насекомых
В середине 60-х годов
Ингибиторы протеиназ в системе защиты растений от насекомых
В середине 60-х годов

Многие белки растительного происхождения, первоначально описанные как ингибиторы трипсина и химотрипсина,
Многие белки растительного происхождения, первоначально описанные как ингибиторы трипсина и химотрипсина,

Модифицированный ген ингибитора ингибиторе трипсина из коровьего гороха (Vigna unguiculata) был
Модифицированный ген ингибитора ингибиторе трипсина из коровьего гороха (Vigna unguiculata) был

Ингибиторы протеиназ в системе защиты растений от патогенных микроорганизмов
Многие белки-ингибиторы протеаз
Ингибиторы протеиназ в системе защиты растений от патогенных микроорганизмов
Многие белки-ингибиторы протеаз

Подавление протеолитической активности может приводить к нарушению способности утилизировать растительные белки
Подавление протеолитической активности может приводить к нарушению способности утилизировать растительные белки

Белковые ингибиторы протеиназ из семян и листьев разных видов люпина подавляли
Белковые ингибиторы протеиназ из семян и листьев разных видов люпина подавляли

В тканях картофеля и томатов присутствуют ингибитор I и ингибитор II,
В тканях картофеля и томатов присутствуют ингибитор I и ингибитор II,

У растений обнаружены белки, обладающие специфической ингибиторной активностью против протеиназ микроорганизмов:
У растений обнаружены белки, обладающие специфической ингибиторной активностью против протеиназ микроорганизмов:

Белок-ингибитор, действующий на микробные протеиназы и не активный по отношению к
Белок-ингибитор, действующий на микробные протеиназы и не активный по отношению к

Ингибиторы цистеиновых протеиназ из растений также способны подавлять рост и развитие
Ингибиторы цистеиновых протеиназ из растений также способны подавлять рост и развитие

Индукция синтеза ингибиторов протеиназ наблюдалась и при бактериальной инфекции.
Инокуляция листьев
Индукция синтеза ингибиторов протеиназ наблюдалась и при бактериальной инфекции.
Инокуляция листьев

Из семян одной из разновидностей проса (Pennisetum glaucum) был выделен ингибитор
Из семян одной из разновидностей проса (Pennisetum glaucum) был выделен ингибитор

В последние годы появились данные, указывающие на то, что ингибиторы протеиназ
В последние годы появились данные, указывающие на то, что ингибиторы протеиназ

Характеристика белковых ингибиторов сериновых протеиназ гречихи
Большинство известных и исследованных в настоящее
Характеристика белковых ингибиторов сериновых протеиназ гречихи
Большинство известных и исследованных в настоящее

В экспериментах установлено, что аминокислотный составы трех очищенных анионных ингибиторов протеиназ
В экспериментах установлено, что аминокислотный составы трех очищенных анионных ингибиторов протеиназ

Анализ аминокислотной последовательности ингибитора BWI-1a показал, что она
на 65 %
Анализ аминокислотной последовательности ингибитора BWI-1a показал, что она
на 65 %
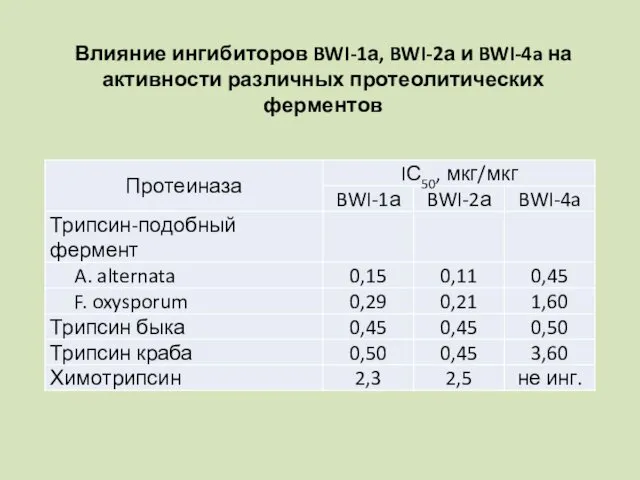
Влияние ингибиторов BWI-1а, BWI-2а и BWI-4a на активности различных протеолитических ферментов
Влияние ингибиторов BWI-1а, BWI-2а и BWI-4a на активности различных протеолитических ферментов

Показано, что BWI-1а и BWI-2а кроме активности трипсина ингибируют также активность
Показано, что BWI-1а и BWI-2а кроме активности трипсина ингибируют также активность
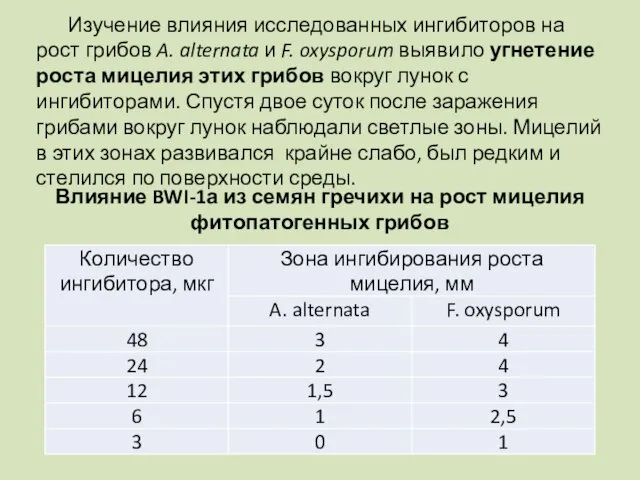
Влияние BWI-1а из семян гречихи на рост мицелия фитопатогенных грибов
Изучение влияния
Влияние BWI-1а из семян гречихи на рост мицелия фитопатогенных грибов
Изучение влияния

Сравнение констант ингибирования сериновых протеиназ ингибиторами протеаз из семян гречихи и
Сравнение констант ингибирования сериновых протеиназ ингибиторами протеаз из семян гречихи и

ДЕФЕНЗИНЫ РАСТЕНИЙ
Растительные дефензины принадлежат к большому семейству антимикробных пептидов с цистеин-стабилизированным
ДЕФЕНЗИНЫ РАСТЕНИЙ
Растительные дефензины принадлежат к большому семейству антимикробных пептидов с цистеин-стабилизированным

Структура дефензинов растений
Дефензины растений – это семейство небольших (около 5
Структура дефензинов растений
Дефензины растений – это семейство небольших (около 5

Сравнение последовательностей растительных дефензинов и растительных дефензин-подобных протеинов. Цистеины обозначены рамкой.
Сравнение последовательностей растительных дефензинов и растительных дефензин-подобных протеинов. Цистеины обозначены рамкой.

При этом были выявлены консервативные положения восьми цистеинов, двух глицинов в
При этом были выявлены консервативные положения восьми цистеинов, двух глицинов в

Локализация и предполагаемый механизм действия дефензинов растений
Было показано, что дефензины растений
Локализация и предполагаемый механизм действия дефензинов растений
Было показано, что дефензины растений

Методами иммунологической или in situ-гибридизации была показана локализация и аккумуляция конститутивно
Методами иммунологической или in situ-гибридизации была показана локализация и аккумуляция конститутивно

По крайней мере, на четырех видах растений, а именно в горохе,
По крайней мере, на четырех видах растений, а именно в горохе,

Было установлено, что экспрессию дефензина арабидопсиса PDF1.2 индуцирует экзогенное применение или
Было установлено, что экспрессию дефензина арабидопсиса PDF1.2 индуцирует экзогенное применение или

Дефензины редьки Rs-AFP2 и георгина Dm-AMP1 не образовывали ион-проникающих пор в
Дефензины редьки Rs-AFP2 и георгина Dm-AMP1 не образовывали ион-проникающих пор в

Один из предполагаемых механизмов действия состоит в том, что мембранный рецептор
Один из предполагаемых механизмов действия состоит в том, что мембранный рецептор

Классификация дефензинов растений
Морфогенные дефензины растений приводят к редукции роста гифов и
Классификация дефензинов растений
Морфогенные дефензины растений приводят к редукции роста гифов и

Напротив, дефензины, выделенные из Dahlia merckii, Clitoria ternatea, Aesculus hippocastanum, относящиеся
Напротив, дефензины, выделенные из Dahlia merckii, Clitoria ternatea, Aesculus hippocastanum, относящиеся

Фунгицидная активность дефензинов растений
Анализ антимикробной активности различных дефензинов показал, что область
Фунгицидная активность дефензинов растений
Анализ антимикробной активности различных дефензинов показал, что область

При использовании точечного мутагенеза было показано, что замена нейтральных остатков gly-9
При использовании точечного мутагенеза было показано, что замена нейтральных остатков gly-9

Фунгицидная активность как морфогенных, так и неморфогенных дефензинов растений уменьшается с
Фунгицидная активность как морфогенных, так и неморфогенных дефензинов растений уменьшается с
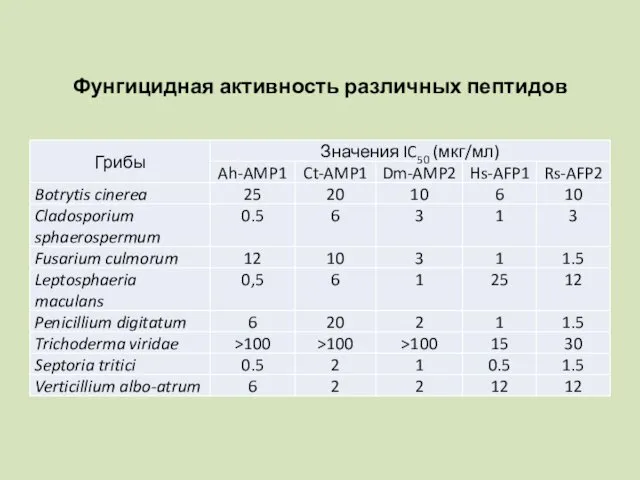
Фунгицидная активность различных пептидов
Фунгицидная активность различных пептидов

Бактерицидная активность дефензинов растений
Антибактериальные активности пептидов были измерены на четырех
Бактерицидная активность дефензинов растений
Антибактериальные активности пептидов были измерены на четырех

Вклад дефензинов в защиту растения-хозяина
Дефензины играют важную роль в защите семян
Вклад дефензинов в защиту растения-хозяина
Дефензины играют важную роль в защите семян

Растения табака были протрансформированы химерной генной конструкцией, включающей кодирующую область гена
Растения табака были протрансформированы химерной генной конструкцией, включающей кодирующую область гена

Болезни картофеля
Картофель относится к числу культур, в сильной степени поражаемых болезнями
Болезни картофеля
Картофель относится к числу культур, в сильной степени поражаемых болезнями

Фитофтороз
Возбудитель - гриб Phytophthora infestans D.В. Первые признаки - на нижних
Фитофтороз
Возбудитель - гриб Phytophthora infestans D.В. Первые признаки - на нижних

Ризоктониоз (белая ножка, черная парша клубней)
Возбудитель - гриб Rhizoctonia solani Kuhn.
Ризоктониоз (белая ножка, черная парша клубней)
Возбудитель - гриб Rhizoctonia solani Kuhn.

Фузариозное увядание, сухая гниль
Возбудитель - гриб Fusarium spp. На растениях нижняя
Фузариозное увядание, сухая гниль
Возбудитель - гриб Fusarium spp. На растениях нижняя

Парша обыкновенная
Возбудитель - актиномицеты Streptomyces scabies Waks et Henr. и другие
Парша обыкновенная
Возбудитель - актиномицеты Streptomyces scabies Waks et Henr. и другие

Рак картофеля
Возбудитель - гриб Sinchytrium endobioticum (Shilb). Проявляется в виде различного
Рак картофеля
Возбудитель - гриб Sinchytrium endobioticum (Shilb). Проявляется в виде различного

Черная ножка и бактериальная мягкая (мокрая) гниль
Возбудитель - бактерии Erwinia caratovora.
Черная ножка и бактериальная мягкая (мокрая) гниль
Возбудитель - бактерии Erwinia caratovora.

Мягкая бактериальная гниль поражает клубни. В месте прикрепления клубня к столону
Мягкая бактериальная гниль поражает клубни. В месте прикрепления клубня к столону

Кольцевая гниль
Возбудитель - бактерии Clavibacter michiganensis ssp. sepedonicus. Поражаются растения и
Кольцевая гниль
Возбудитель - бактерии Clavibacter michiganensis ssp. sepedonicus. Поражаются растения и

Крапчатая (обыкновенная, мягкая) мозаика картофеля
Возбудитель - Х-вирус картофеля. Проявляется в виде
Крапчатая (обыкновенная, мягкая) мозаика картофеля
Возбудитель - Х-вирус картофеля. Проявляется в виде

Морщинистая и полосчатая мозаики. Возбудитель - Y-вирус картофеля. Различают три основных
Морщинистая и полосчатая мозаики. Возбудитель - Y-вирус картофеля. Различают три основных

Скручивание листьев картофеля
Возбудитель - L-вирус картофеля (Вирус скручивания листьев картофеля -
Скручивание листьев картофеля
Возбудитель - L-вирус картофеля (Вирус скручивания листьев картофеля -

Ведьмина метла картофеля
Возбудитель - фитоплазма. Куст становится низкорослым, с большим количеством
Ведьмина метла картофеля
Возбудитель - фитоплазма. Куст становится низкорослым, с большим количеством

Бронзовость листьев
Возникает при недостаточном поступлении калия в растения. Листья покрываются мелкими
Бронзовость листьев
Возникает при недостаточном поступлении калия в растения. Листья покрываются мелкими

Коричневая пятнистость стеблей и клубней
Причина возникновения - избыточное поступление в растение
Коричневая пятнистость стеблей и клубней
Причина возникновения - избыточное поступление в растение

Картофельная нематода (бледная и золотистая)
Globodera rostochiensis и Globodera pallida. На пораженных
Картофельная нематода (бледная и золотистая)
Globodera rostochiensis и Globodera pallida. На пораженных

Стеблевая нематода
Ditylenchus destructor. Видимые признаки в период вегетации отсутствуют. Может вызывать
Стеблевая нематода
Ditylenchus destructor. Видимые признаки в период вегетации отсутствуют. Может вызывать

Галловая нематода
Meloidogyne marioni. Пораженные растения отстают в росте и увядают. На
Галловая нематода
Meloidogyne marioni. Пораженные растения отстают в росте и увядают. На

Генетическая инженерия картофеля
В сочетании с современными технологиями возделывания различные виды пестицидов
Генетическая инженерия картофеля
В сочетании с современными технологиями возделывания различные виды пестицидов

Картофель, наряду с другим известным представителем семейства Solanaceae Nicotiana tabacum был
Картофель, наряду с другим известным представителем семейства Solanaceae Nicotiana tabacum был

Основные факторы, влияющие на эффективность генетической модификации растений
Основные факторы, влияющие на эффективность генетической модификации растений

Основным фактором, определяющим эффективность получения трансгенных растений, является восприимчивость эксплантов конкретного
Основным фактором, определяющим эффективность получения трансгенных растений, является восприимчивость эксплантов конкретного

При селекции картофеля в полевых условиях необходимо провести оценку 50-100 тысяч
При селекции картофеля в полевых условиях необходимо провести оценку 50-100 тысяч
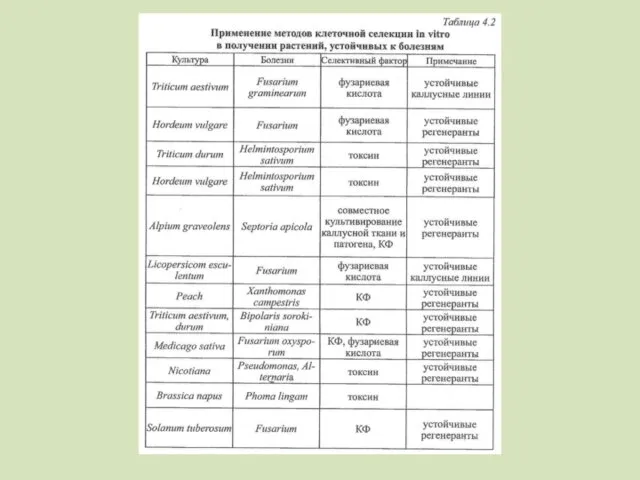

Использование патогенов в клеточной селекции
Непосредственное культивирование в присутствии патогена, если мало
Использование патогенов в клеточной селекции
Непосредственное культивирование в присутствии патогена, если мало

Некоторые грибы растут очень быстро, что не позволяет идентифицировать устойчивые клетки
С
Некоторые грибы растут очень быстро, что не позволяет идентифицировать устойчивые клетки
С

Использование патотоксинов в клеточной селекции
Большинство штаммов грибов производит микотоксины и поражает
Использование патотоксинов в клеточной селекции
Большинство штаммов грибов производит микотоксины и поражает

Патотоксины:
1 категория – токсины, не являющиеся определяющими в заболевании, но обладающие
Патотоксины:
1 категория – токсины, не являющиеся определяющими в заболевании, но обладающие
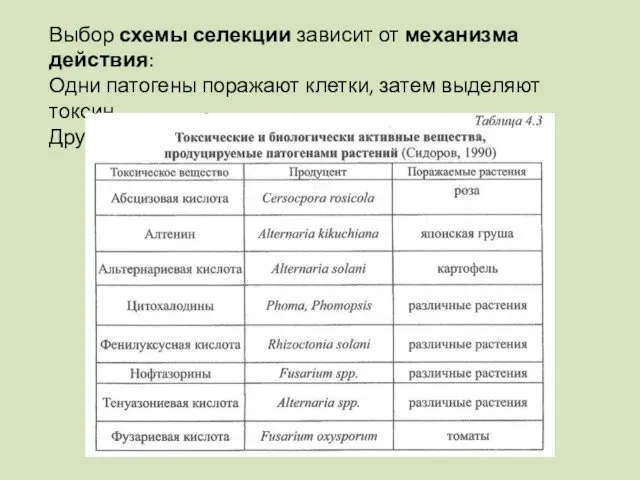
Выбор схемы селекции зависит от механизма действия:
Одни патогены поражают клетки, затем
Выбор схемы селекции зависит от механизма действия:
Одни патогены поражают клетки, затем
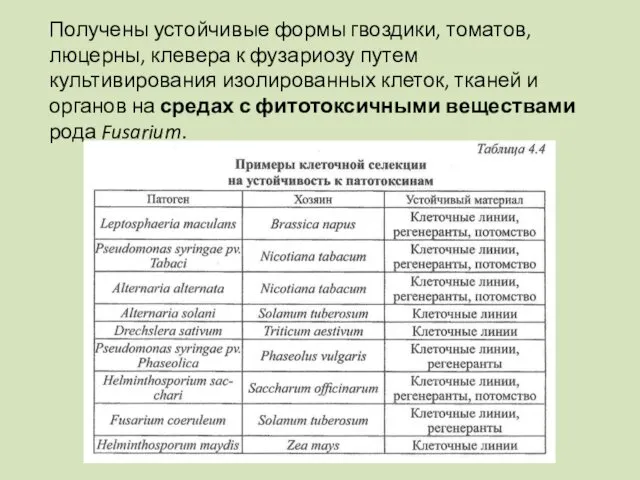
Получены устойчивые формы гвоздики, томатов, люцерны, клевера к фузариозу путем культивирования
Получены устойчивые формы гвоздики, томатов, люцерны, клевера к фузариозу путем культивирования

Для ряда культур отбор с использованием селективного фактора фузариевой кислоты (5-н-бутил-2-пиридин-карбоновая
Для ряда культур отбор с использованием селективного фактора фузариевой кислоты (5-н-бутил-2-пиридин-карбоновая

ВНИИ льна – лен-долгунец – изолированные пыльники и каллусная ткань –
ВНИИ льна – лен-долгунец – изолированные пыльники и каллусная ткань –

Использование культурального фильтрата патогена в клеточной селекции
Следует учитывать:
- способ получения КФ
-
Использование культурального фильтрата патогена в клеточной селекции
Следует учитывать:
- способ получения КФ
-

Фузариоз на томатах
ступенчатый отбор – 2n=48, 2n=64,
прямой отбор – 2n=32
Клевер,
Фузариоз на томатах
ступенчатый отбор – 2n=48, 2n=64,
прямой отбор – 2n=32
Клевер,

Сельдерей – септориоз – каллусные и суспензионные культуры из молодых черешков
Важно
Сельдерей – септориоз – каллусные и суспензионные культуры из молодых черешков
Важно

Участие фитогормонов в клеточной селекции
Под действием факторов биотической природы происходит быстрый
Участие фитогормонов в клеточной селекции
Под действием факторов биотической природы происходит быстрый
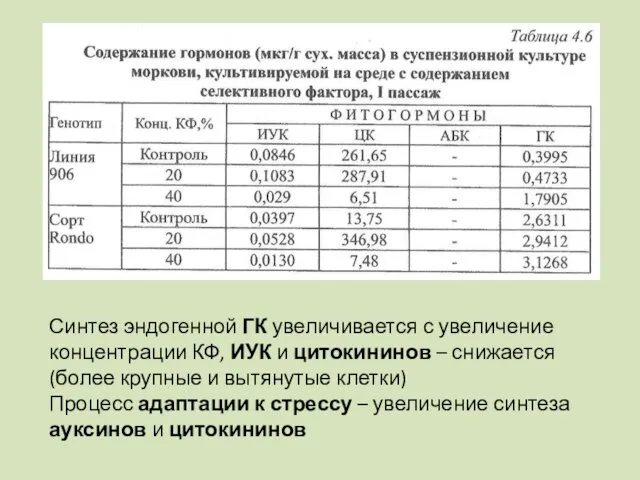
Синтез эндогенной ГК увеличивается с увеличение концентрации КФ, ИУК и цитокининов
Синтез эндогенной ГК увеличивается с увеличение концентрации КФ, ИУК и цитокининов
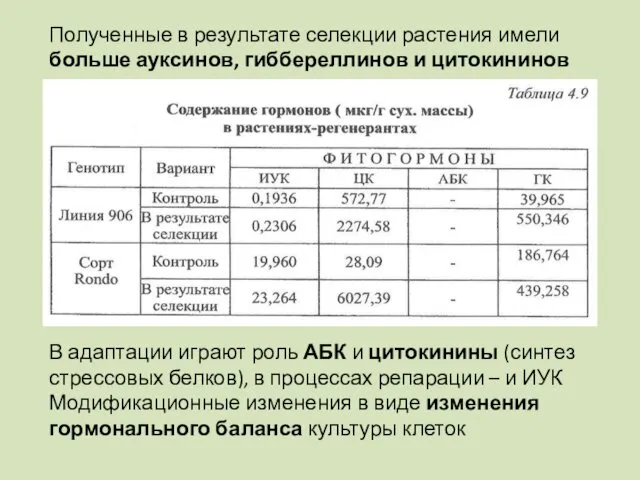
Полученные в результате селекции растения имели больше ауксинов, гиббереллинов и цитокининов
В
Полученные в результате селекции растения имели больше ауксинов, гиббереллинов и цитокининов
В

Устойчивость против проникновения фитопатогенов
Накопление и отложение лигнина в местах внедрения патогена.
Устойчивость против проникновения фитопатогенов
Накопление и отложение лигнина в местах внедрения патогена.
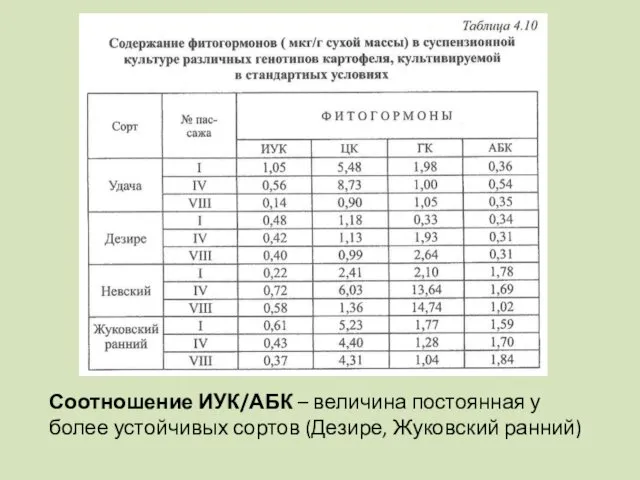
Соотношение ИУК/АБК – величина постоянная у более устойчивых сортов (Дезире, Жуковский
Соотношение ИУК/АБК – величина постоянная у более устойчивых сортов (Дезире, Жуковский
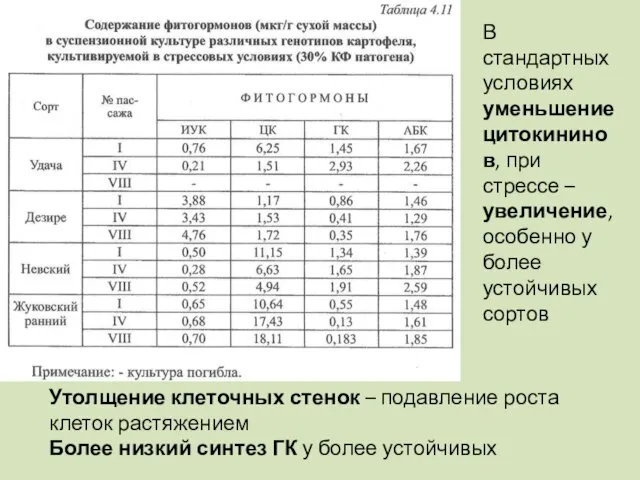
В стандартных условиях уменьшение цитокининов, при стрессе – увеличение, особенно у
В стандартных условиях уменьшение цитокининов, при стрессе – увеличение, особенно у

Участие фенольных соединений
Механизм защиты и адаптации
Поражение патогенами активизирует синтез фенольных соединений,
Участие фенольных соединений
Механизм защиты и адаптации
Поражение патогенами активизирует синтез фенольных соединений,
 Познавательно – игровая программа: Благодарное слово.
Познавательно – игровая программа: Благодарное слово. Будущие защитники
Будущие защитники Невроз навязчивых состояний
Невроз навязчивых состояний Русский фольклор
Русский фольклор Профилактика профессиональных заболеваний
Профилактика профессиональных заболеваний INDIVIDUAL_NYJ_PROEKT_pptx_Mashi (2)
INDIVIDUAL_NYJ_PROEKT_pptx_Mashi (2) Особенности организации проектной деятельности в образовательных организациях РФ
Особенности организации проектной деятельности в образовательных организациях РФ Автоматизация звука [Ль]
Автоматизация звука [Ль] Задачи на разрезание. Занятие 5
Задачи на разрезание. Занятие 5 Анкетирование Как повысить успеваемость по химии?
Анкетирование Как повысить успеваемость по химии? Инновационный менеджмент
Инновационный менеджмент Проект в старшей группе Профессия Нефтяник
Проект в старшей группе Профессия Нефтяник Презентация Азбука ч.3 Диск
Презентация Азбука ч.3 Диск Строевая подготовка
Строевая подготовка Амплипульстерапия. Интерференцтерапия. Флюктуоризация
Амплипульстерапия. Интерференцтерапия. Флюктуоризация Вся жизнь-полет
Вся жизнь-полет Выставка детских работ Открытка для мамы
Выставка детских работ Открытка для мамы Залесские земли.
Залесские земли. Программа работы с родителями
Программа работы с родителями Выпрямители
Выпрямители Покажи свое настроение. Прививка
Покажи свое настроение. Прививка Специфика восприятия пространства и времени в разных культурных группах
Специфика восприятия пространства и времени в разных культурных группах Культура и духовная сфера жизни
Культура и духовная сфера жизни Острый живот
Острый живот Физкультурно-оздоровительный комплекс в Санкт-Петербурге
Физкультурно-оздоровительный комплекс в Санкт-Петербурге Беніто Муссоліні ( 1883 – 1945)
Беніто Муссоліні ( 1883 – 1945)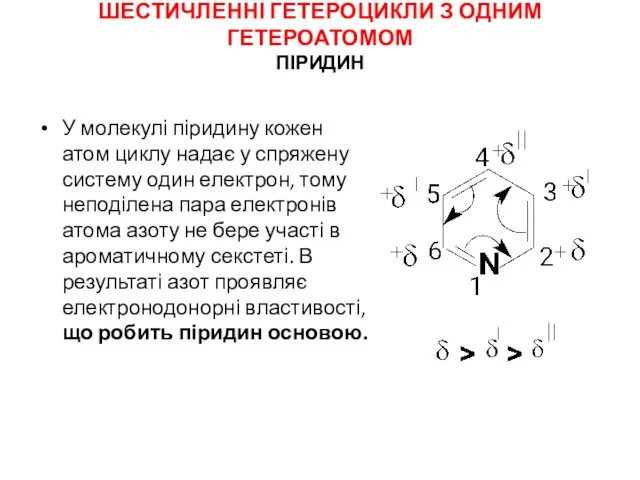 Шестичленні гетероцикли з одним гетероатомом. Піридин
Шестичленні гетероцикли з одним гетероатомом. Піридин Все народы воспевают материнство
Все народы воспевают материнство