- Генетическая инженерия

Содержание
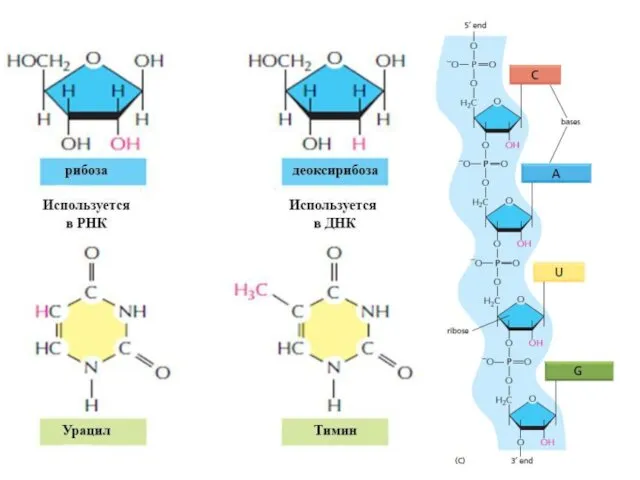
- 2. Аденин (А) Гуанин (Г, G) Тимин (Т) Урацил (У, U) Цитозин (Ц, С)
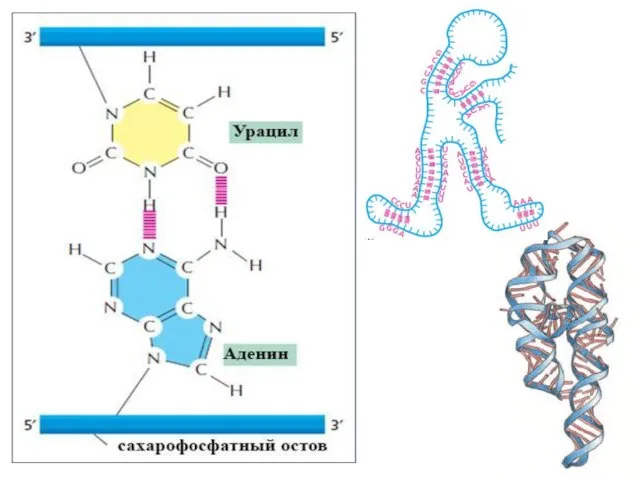
- 4. 5’ 3’ 5’ 3’ 5’ 5’ 3’ 3’
- 6. Репликация ДНК
- 7. Репликация ДНК
- 8. Репликация ДНК
- 10. ttctcatgtttgacagcttatcatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatcctctacgccggacgcatcgtggccggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgaccgatgcccttgagagccttcaacccagtcagctccttccggtgggcgcggggcatgactatcgtcgccgcacttatgactgtcttctttatcatgcaactcgtaggacaggtgccggcagcgctctgggtcattttcggcgaggaccgctttcgctggagcgcgacgatgatcggcctgtcgcttgcggtattcggaatcttgcacgccctcgctcaagccttcgtcactggtcccgccaccaaacgtttcggcgagaagcaggccattatcgccggcatggcggccgacgcgctgggctacgtcttgctggcgttcgcgacgcgaggctggatggccttccccattatgattcttctcgcttccggcggcatcgggatgcccgcgttgcaggccatgctgtccaggcaggtagatgacgaccatcagggacagcttcaaggatcgctcgcggctcttaccagcctaacttcgatcactggaccgctgatcgtcacggcgatttatgccgcctcggcgagcacatggaacgggttggcatggattgtaggcgccgccctataccttgtctgcctccccgcgttgcgtcgcggtgcatggagccgggccacctcgacctgaatggaagccggcggcacctcgctaacggattcaccactccaagaattggagccaatcaattcttgcggagaactgtgaatgcgcaaaccaacccttggcagaacatatccatcgcgtccgccatctccagcagccgcacgcggcgcatctcgggcagcgttgggtcctggccacgggtgcgcatgatcgtgctcctgtcgttgaggacccggctaggctggcggggttgccttactggttagcagaatgaatcaccgatacgcgagcgaacgtgaagcgactgctgctgcaaaacgtctgcgacctgagcaacaacatgaatggtcttcggtttccgtgtttcgtaaagtctggaaacgcggaagtcagcgccctgcaccattatgttccggatctgcatcgcaggatgctgctggctaccctgtggaacacctacatctgtattaacgaagcgctggcattgaccctgagtgatttttctctggtcccgccgcatccataccgccagttgtttaccctcacaacgttccagtaaccgggcatgttcatcatcagtaacccgtatcgtgagcatcctctctcgtttcatcggtatcattacccccatgaacagaaatcccccttacacggaggcatcagtgaccaaacaggaaaaaaccgcccttaacatggcccgctttatcagaagccagacattaacgcttctggagaaactcaacgagctggacgcggatgaacaggcagacatctgtgaatcgcttcacgaccacgctgatgagctttaccgcagctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaa
- 11. Современная концепция гена ПРОМОТОР СТРУКТУРНАЯ ЧАСТЬ ГЕНА ТЕРМИНАТОР
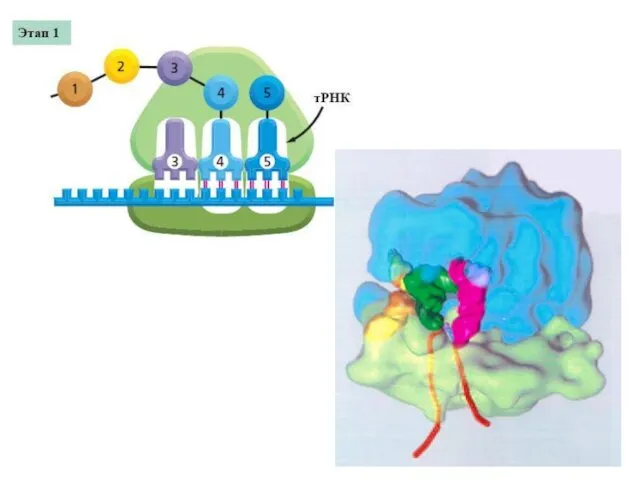
- 17. Трансляция
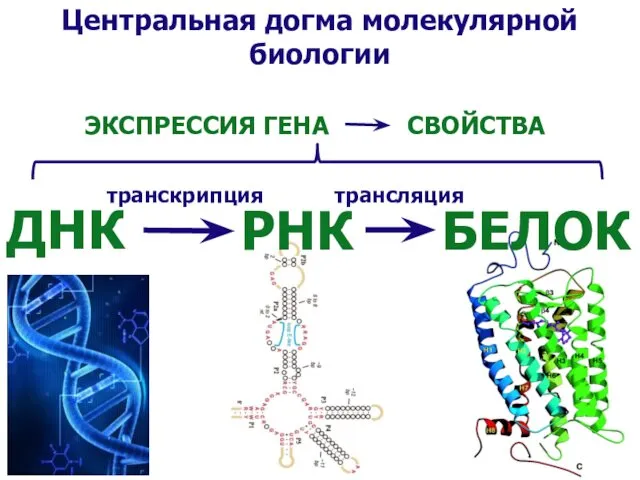
- 24. Центральная догма молекулярной биологии ДНК РНК БЕЛОК транскрипция трансляция ЭКСПРЕССИЯ ГЕНА СВОЙСТВА
- 25. Биотехнология в современном мире
- 26. Предпосылки возникновения генной инженерии и история развития
- 27. Рекомбинантная ДНК
- 28. Ферменты, применяемые в генетической инженерии
- 29. I. РЕСТРИКТАЗЫ (ЕС 3.1.21.) ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ)
- 30. РЕСТРИКТАЗЫ II типа HindIII (Haemophilus influenzae) 4 – 14 нуклеотидов
- 31. ПАЛИНДРОМЫ А РОЗА УПАЛА НА ЛАПУ АЗОРА По обеим цепям ДНК в одном направлении читаются одинаково
- 32. Рестриктазы в генной инженерии EcoRI (E. coli RY13) NdeI (Neisseria denitrificans) BamHI (Bacillus amyloliquefaciens H) BglII
- 33. Restriction fragment length polymorphism (RFLP-analysis)
- 34. II. ЛИГАЗА (EC 6.5.1.1)
- 35. Механизм действия
- 36. ДНК-лигаза фага Т4 Единица активности лигазы соответствует количеству фермента, необходимого для лигирования фрагментов ДНК фага λ
- 37. Лигирование по липким и тупым концам
- 38. V. Нуклеазы – гидролитические ферменты, расщепляющие фосфодиэфирные связи в нуклеиновых кислотах
- 39. Нуклеазы Экзонуклеазы Эндонуклеазы Экзонуклеаза III E. coli (катализирует последовательное отщепление нуклеотидов из дцДНК в направлении 3’----5’
- 40. V. ДНК-полимеразы
- 41. Репликация ДНК
- 42. матрица праймер Фрагмент Кленова ДНК-полимераза фага Т4 ДНК полимераза Thermus aquaticus
- 43. ДНК-полимеразы термофильных архебактерий
- 44. История открытия Апрель 1983 г – идея ПЦР Декабрь 1983 г – осуществление ПЦР 1993 г
- 45. Что такое ПЦР? По сути, это упрощенная версия репликации бактерий, при которой возрастает количество копий специфической
- 46. Что такое ПЦР?
- 47. Сравнение ПЦР и репликации in vivo (1)
- 48. Сравнение ПЦР и репликации in vivo (2)
- 49. Компоненты реакции ПЦР Master Mix (MM)
- 50. 72 - 75 0C амплификация 90 - 99 0C денатурация Temperature, 0C 40 – 70 0C
- 51. Денатурация 3’ 5’ 5’ 3’ 5’ 3’ 3’ 5’
- 52. Отжиг праймеров 3’ 5’ 5’ 3’ • Праймеры комплементарно связываются с ДНК, согласно их температуре плавления
- 53. Температура плавления (T melting, Tm) Под температурой плавления понимают температуру, при которой половина молекул гибридизована (находится
- 54. Расчет оптимальной температуры денатурации ампликона Td=Tm+3-40С
- 55. Температура отжига (T annealing, Ta) Ta=Tm – 50С (по Ребрикову) Ta=Tm – 100С (по Патрушеву) Под
- 56. Задача №1 Определите температуры плавления и отжига следующего олигонуклеотида: 5’ GGCATTTAGCTTAGGC 3’ Решение: N=16: А=3, Т=5,
- 57. Задача №2 Определите температуры плавления и отжига следующего олигонуклеотида: 5’ GGCATTTAGCTTAGGCTTAAGGCGGGAG 3’ Решение: N=28: А=6, Т=7,
- 58. Термодинамический расчет температуры отжига
- 59. Конструирование (дизайн) праймеров: При подборе праймеров зачастую удобно использовать програмное обеспечение, упрощающее поиск подходящих регионов или
- 60. Эффективность ПЦР Y=X×2n
- 61. ПЦР в реальном времени
- 62. 2a. Фильтр возбуждения 2b. Фильтр эмиссии 1. Источник излучения 4. образцы 3. Усилитель 5. Полупроводниковый детектор
- 63. Amplification Plot of real-time PCR DNA copy number (log) PCR cycle (Ct)
- 64. Секвенирование по Сэнглеру В 1977 г. автор способ ферментативного секвенирования, получивший название метода терминирующих аналогов трифосфатов.
- 65. Секвенирование ДНК Использование электрофореза в капиллярах, автоматическое считывание флуоресценции на границе выхода из геля
- 66. Обратная транскриптаза (КФ 2.7.7.49) (ревертаза)
- 67. Функции обратных транскриптаз Ферменты катализируют синтез ДНК на матрице РНК в процессе обратной транскрипции. Обладает: РНК-зависимой
- 68. Обратные транскриптазы в генной инженерии Обратная транскриптаза вируса миелобластоза птиц (Avian Myeloblastosis Virus, AMV) Помимо 5’-
- 69. Процессинг РНК
- 70. Реакция ОТ Рабочая температура: 370С; Время реакции: 1 час; Ионы магния; Чистая матрица (обработка ДНКазой); В
- 71. ДНК-полимераза Tth Обратная транскрипция и ПЦР в одной пробирке !!!
- 72. ВЕКТОРЫ
- 73. Что такое вектор? Вектор – молекула ДНК, используемая в генетической инженерии для передачи генетического материала другой
- 74. Свойства векторов По функциям: Векторы для клонирования Векторы для экспрессии Векторы для трансформации По месту применения:
- 75. Плазмидная ДНК бактерий Способность к автономной репликации; (ориджин репликации, ori) Емкость вектора; (до размеров самой плазмиды,
- 76. Локусы контроля репликации плазмидной ДНК
- 77. Создание плазмид с повышенной копийностью _____________________________________________ Плазмида Ori Копийность Классификация _____________________________________________ pUC pMB1* 500 – 700
- 78. Селективные маркеры
- 79. Емкость плазмиды
- 80. Отличие векторов и плазмид Уникальные сайты, емкость плазмид до 10 kb!!!
- 81. Полилинкер (MCS) небольшой, искусственно синтезированный фрагмент ДНК, который представляет собой последовательность, содержащую сайты рестрикции для наиболее
- 82. Серии векторов
- 83. Бело-голубая селекция
- 84. Бело-голубая селекция ИПТГ – индуктор lac-оперона X-Gal – хромогенный субстрат
- 85. Космиды Это гибридный тип векторов, совмещающий в себе свойства плазмиды и фага λ. Они конструируются на
- 86. Фазмиды (фасмиды, фагмиды) Это гибридный тип векторов, совмещающий в себе свойства плазмиды и нитчатых фагов (f1,
- 87. Искусственные хромосомы
- 88. Искусственные хромосомы дрожжей (yeast artificial chromosomes, YACs) Cen4 – центромера дрожжей; ORI – ориджин бактерий; Telomere
- 89. Создание геномных библиотек
- 90. Геномные библиотеки (банки генов) Геномная библиотека – фрагменты генома, клонированные в фаге λ (фаговые библиотеки) или
- 91. Принцип конструирования геномных библиотек
- 92. Перекрывающиеся последовательности Перекрытие – в скольких фрагментах у вас будет 1 участок ДНК
- 93. II Этап. Создание геномной библиотеки Левое и правое плечо фага с адапторами HindIII
- 94. III Этап. Клонирование геномной библиотеки
- 95. Расчет количества клонов, необходимых для получения геномной библиотеки Где N – количество клонов, P – вероятность
- 96. BAC, PAC библиотеки Сложно выделять ДНК (фрагментация редкощепящими рестриктазами или не режут вообще)
- 97. Поиск (скрининг) клонов в библиотеке Гибридизация – отжиг цепи ДНК на комплементарной ей цепи ДНК
- 98. Принцип проведения скрининга
- 99. Гибридизация: получение реплики Чашка Петри Нитроцеллюлозная (нейлоновая) мембрана
- 100. Блоттинг (по Саузерну)
- 101. Прогулка по хромосомам
- 102. Библиотеки кДНК
- 103. ТРАНСГЕННЫЕ РАСТЕНИЯ
- 104. 50-е годы XX века Получение культур in vitro - растений «в пробирке»
- 105. 1983 год, первое трансгенное растение Растение табака, устойчивое к канамицину
- 106. 1990 год, первое трансгенное растение, пошедшее «в поля» Растения хлопка, устойчивые к насекомым
- 107. 1994 год, томаты «flavr savr» Устойчивость к бактериальным гнилям
- 108. 1995 год, Monsanto Соя, устойчивая к гербицидам
- 109. Картофель, устойчивый к колородскому жуку Кукуруза, устойчивая к кукурузной огневке, гербицидам Папайя, устойчивая к вирусу кольцевой
- 110. Suntory – голубая трансгенная роза (дигидрокверцитин 5’-гидролаза)
- 111. - трансформация клеток Как получают трансгенные растения? - Получение культур и регенерация культур in vitro
- 112. Агробактериальная трансформация Agrobacterium tumefaciens (A. tumefaciens)
- 113. A. tumefaciens вызывает болезнь корончатых галлов
- 114. Корончатые галлы состоят из дедифференцированных клеток
- 115. A. tumefaciens – плазмидосодержащая бактерия
- 116. Опины – источники азота и энергии агробактерии Нопалины Октопины Агропины
- 117. (локус shi) (локус roi)
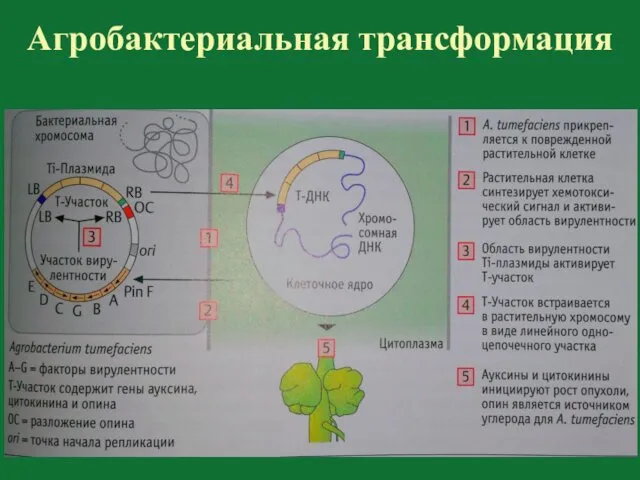
- 118. Агробактериальная трансформация
- 119. клеточная стенка растения оболочка клетки агробактерии Vir A сигнальные молекулы растения P Vir G Vir G
- 120. Влияние растительных гормонов на клетку
- 122. Оценка устойчивости трансгенных линий сахарной свеклы к действию гербицида «Баста» КОНТРОЛЬ ТРАНСГЕННЫЕ ЛИНИИ ТРАНСГЕННЫЕ ЛИНИИ ТРАНСГЕННЫЕ
- 123. Получение трансгенного картофеля
- 124. Участок, зарегистрированный МВК ГИД, Краснодар, ВНИИБЗР Контрольные растения ГМ сорт Невский ГМ сорт Луговской
- 125. Контрольные растения, 150 mM NaCl Трансгенные растения, 150 mM NaCl Контрольные растения Трансгенные растения NaCl 25
- 126. РИС (Oryza sativa ) Основной пищевой продукт, производимый в мире Низкое содержание витамина А и железа
- 127. “Золотой” рис с повышенным содержанием β -каротина
- 128. Получение лекарственных препаратов в растениях РАСТЕНИЯ – БИОФАРМАЦЕВТИКИ
- 130. Скачать презентацию
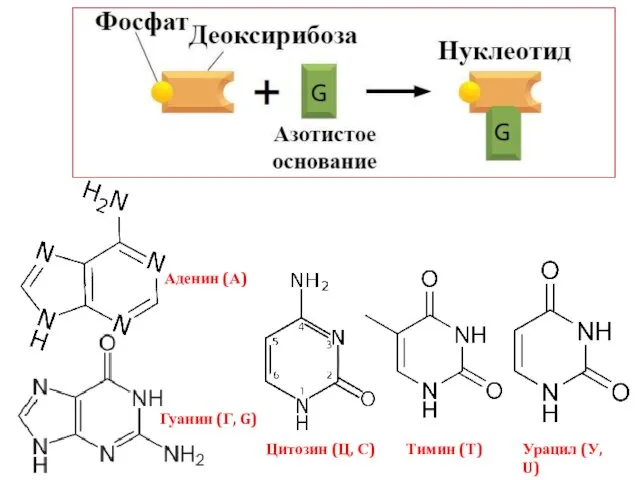
Аденин (А)
Гуанин (Г, G)
Тимин (Т)
Урацил (У, U)
Цитозин (Ц, С)
Аденин (А)
Гуанин (Г, G)
Тимин (Т)
Урацил (У, U)
Цитозин (Ц, С)
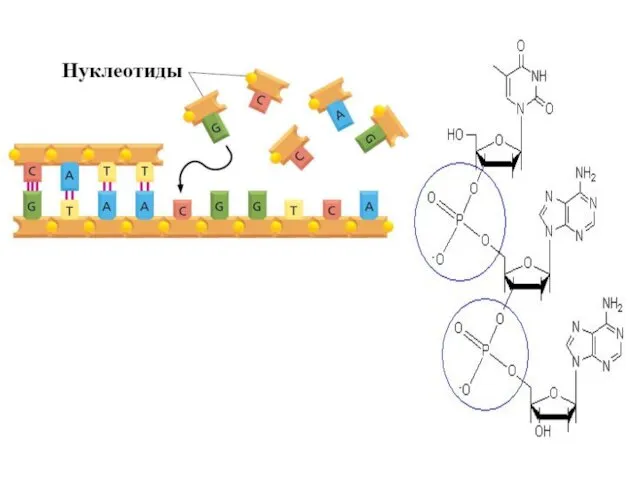
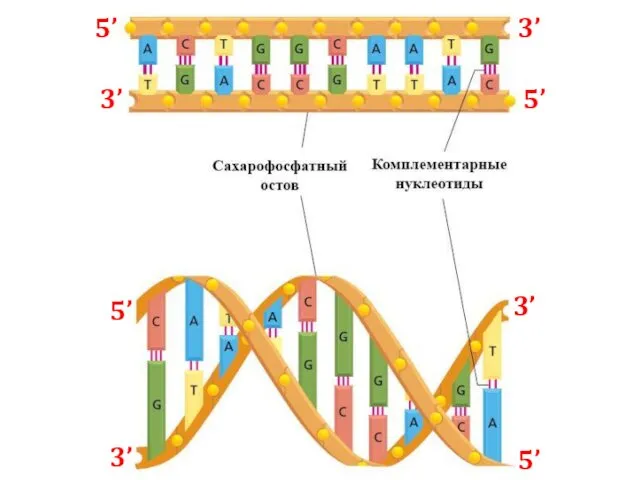
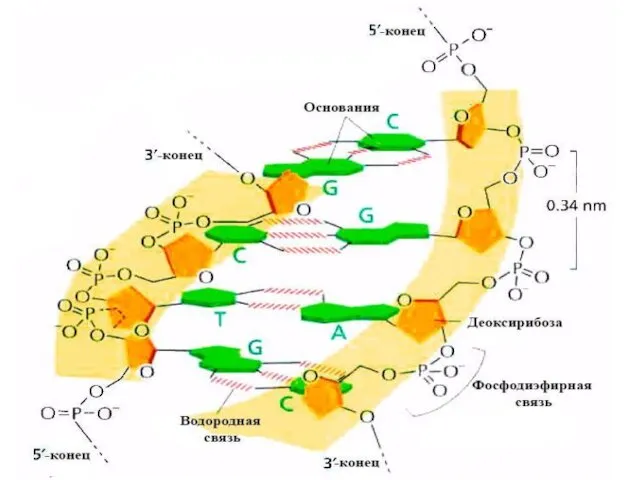
5’
3’
5’
3’
5’
5’
3’
3’
5’
3’
5’
3’
5’
5’
3’
3’
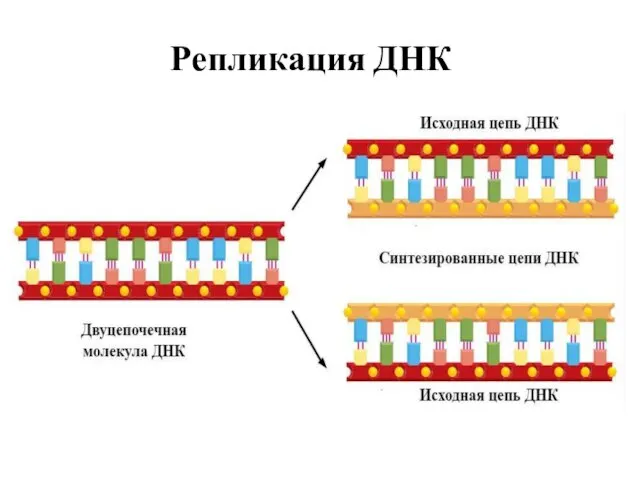
Репликация ДНК
Репликация ДНК
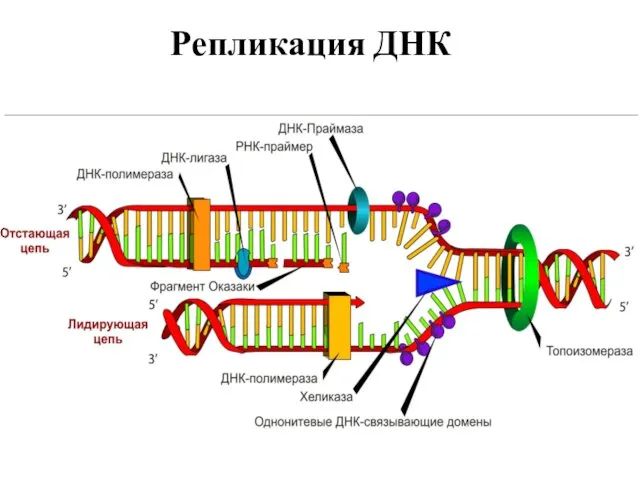
Репликация ДНК
Репликация ДНК
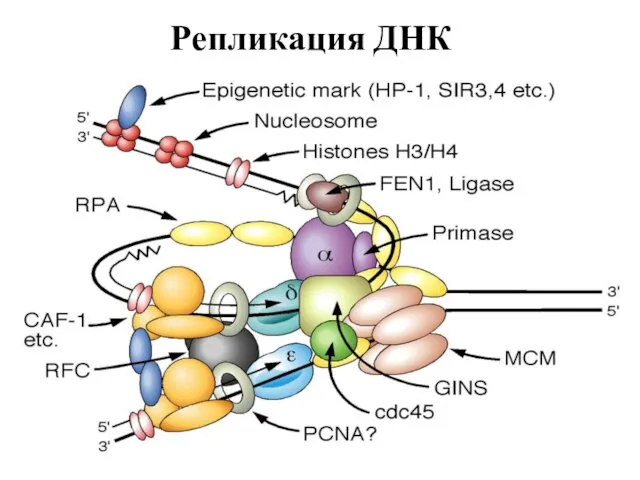
Репликация ДНК
Репликация ДНК

ttctcatgtttgacagcttatcatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatcctctacgccggacgcatcgtggccggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgaccgatgcccttgagagccttcaacccagtcagctccttccggtgggcgcggggcatgactatcgtcgccgcacttatgactgtcttctttatcatgcaactcgtaggacaggtgccggcagcgctctgggtcattttcggcgaggaccgctttcgctggagcgcgacgatgatcggcctgtcgcttgcggtattcggaatcttgcacgccctcgctcaagccttcgtcactggtcccgccaccaaacgtttcggcgagaagcaggccattatcgccggcatggcggccgacgcgctgggctacgtcttgctggcgttcgcgacgcgaggctggatggccttccccattatgattcttctcgcttccggcggcatcgggatgcccgcgttgcaggccatgctgtccaggcaggtagatgacgaccatcagggacagcttcaaggatcgctcgcggctcttaccagcctaacttcgatcactggaccgctgatcgtcacggcgatttatgccgcctcggcgagcacatggaacgggttggcatggattgtaggcgccgccctataccttgtctgcctccccgcgttgcgtcgcggtgcatggagccgggccacctcgacctgaatggaagccggcggcacctcgctaacggattcaccactccaagaattggagccaatcaattcttgcggagaactgtgaatgcgcaaaccaacccttggcagaacatatccatcgcgtccgccatctccagcagccgcacgcggcgcatctcgggcagcgttgggtcctggccacgggtgcgcatgatcgtgctcctgtcgttgaggacccggctaggctggcggggttgccttactggttagcagaatgaatcaccgatacgcgagcgaacgtgaagcgactgctgctgcaaaacgtctgcgacctgagcaacaacatgaatggtcttcggtttccgtgtttcgtaaagtctggaaacgcggaagtcagcgccctgcaccattatgttccggatctgcatcgcaggatgctgctggctaccctgtggaacacctacatctgtattaacgaagcgctggcattgaccctgagtgatttttctctggtcccgccgcatccataccgccagttgtttaccctcacaacgttccagtaaccgggcatgttcatcatcagtaacccgtatcgtgagcatcctctctcgtttcatcggtatcattacccccatgaacagaaatcccccttacacggaggcatcagtgaccaaacaggaaaaaaccgcccttaacatggcccgctttatcagaagccagacattaacgcttctggagaaactcaacgagctggacgcggatgaacaggcagacatctgtgaatcgcttcacgaccacgctgatgagctttaccgcagctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaa
ttctcatgtttgacagcttatcatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatcctctacgccggacgcatcgtggccggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgaccgatgcccttgagagccttcaacccagtcagctccttccggtgggcgcggggcatgactatcgtcgccgcacttatgactgtcttctttatcatgcaactcgtaggacaggtgccggcagcgctctgggtcattttcggcgaggaccgctttcgctggagcgcgacgatgatcggcctgtcgcttgcggtattcggaatcttgcacgccctcgctcaagccttcgtcactggtcccgccaccaaacgtttcggcgagaagcaggccattatcgccggcatggcggccgacgcgctgggctacgtcttgctggcgttcgcgacgcgaggctggatggccttccccattatgattcttctcgcttccggcggcatcgggatgcccgcgttgcaggccatgctgtccaggcaggtagatgacgaccatcagggacagcttcaaggatcgctcgcggctcttaccagcctaacttcgatcactggaccgctgatcgtcacggcgatttatgccgcctcggcgagcacatggaacgggttggcatggattgtaggcgccgccctataccttgtctgcctccccgcgttgcgtcgcggtgcatggagccgggccacctcgacctgaatggaagccggcggcacctcgctaacggattcaccactccaagaattggagccaatcaattcttgcggagaactgtgaatgcgcaaaccaacccttggcagaacatatccatcgcgtccgccatctccagcagccgcacgcggcgcatctcgggcagcgttgggtcctggccacgggtgcgcatgatcgtgctcctgtcgttgaggacccggctaggctggcggggttgccttactggttagcagaatgaatcaccgatacgcgagcgaacgtgaagcgactgctgctgcaaaacgtctgcgacctgagcaacaacatgaatggtcttcggtttccgtgtttcgtaaagtctggaaacgcggaagtcagcgccctgcaccattatgttccggatctgcatcgcaggatgctgctggctaccctgtggaacacctacatctgtattaacgaagcgctggcattgaccctgagtgatttttctctggtcccgccgcatccataccgccagttgtttaccctcacaacgttccagtaaccgggcatgttcatcatcagtaacccgtatcgtgagcatcctctctcgtttcatcggtatcattacccccatgaacagaaatcccccttacacggaggcatcagtgaccaaacaggaaaaaaccgcccttaacatggcccgctttatcagaagccagacattaacgcttctggagaaactcaacgagctggacgcggatgaacaggcagacatctgtgaatcgcttcacgaccacgctgatgagctttaccgcagctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaa
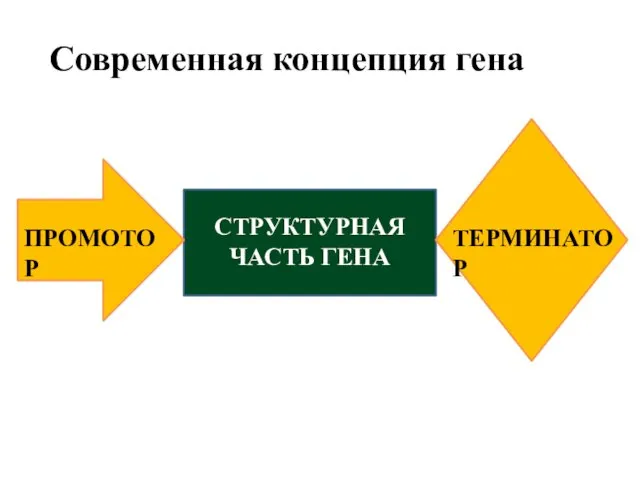
Современная концепция гена
ПРОМОТОР
СТРУКТУРНАЯ ЧАСТЬ ГЕНА
ТЕРМИНАТОР
Современная концепция гена
ПРОМОТОР
СТРУКТУРНАЯ ЧАСТЬ ГЕНА
ТЕРМИНАТОР
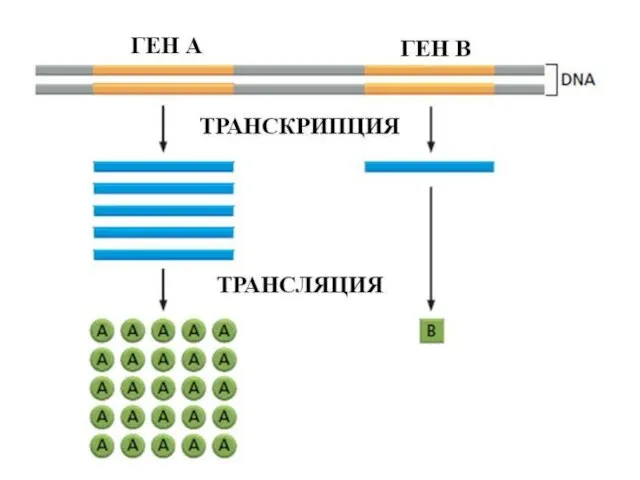
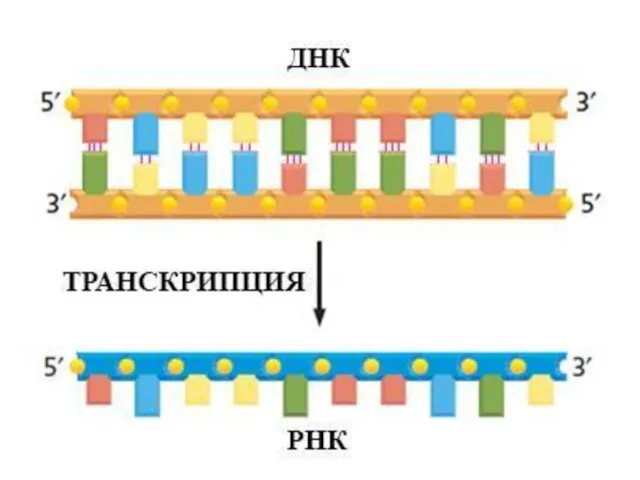
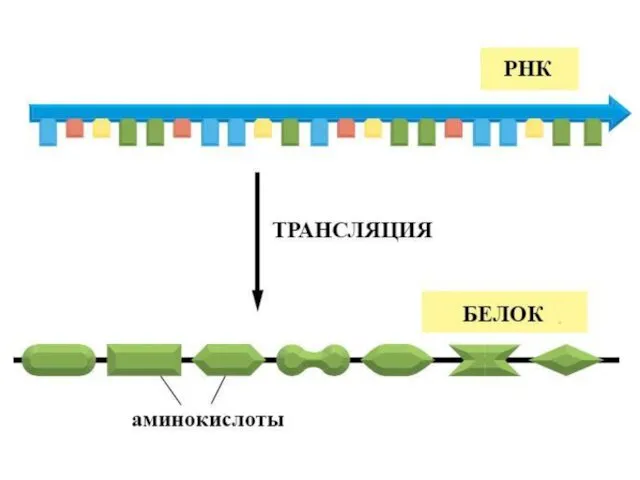
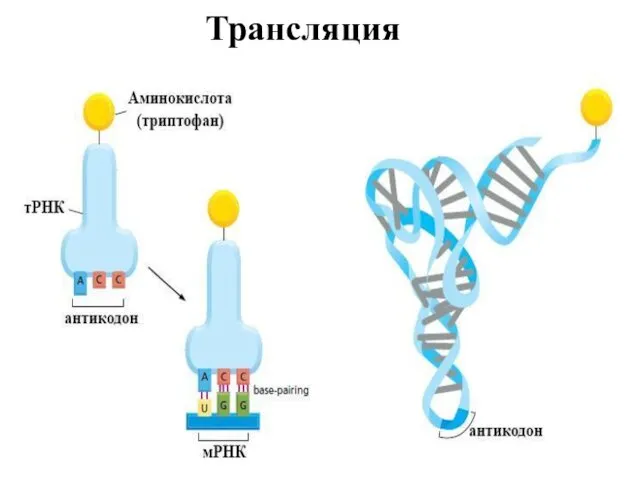
Трансляция
Трансляция
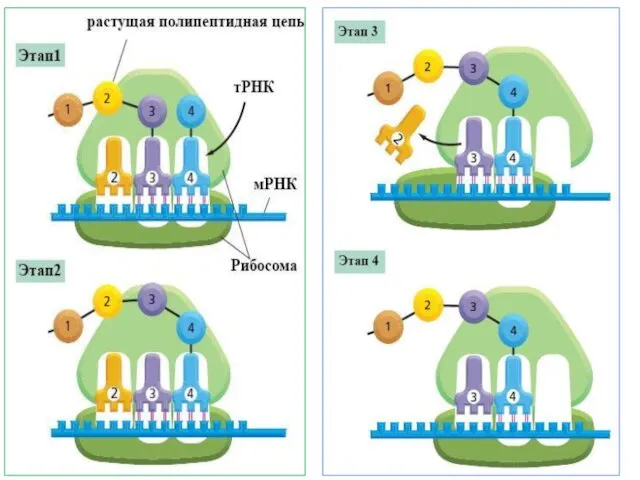
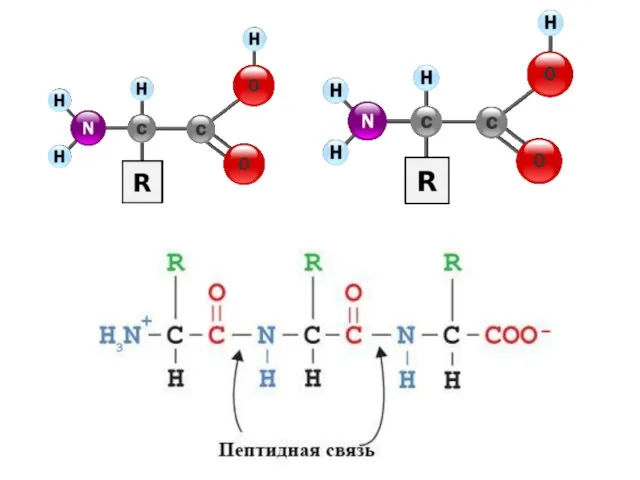
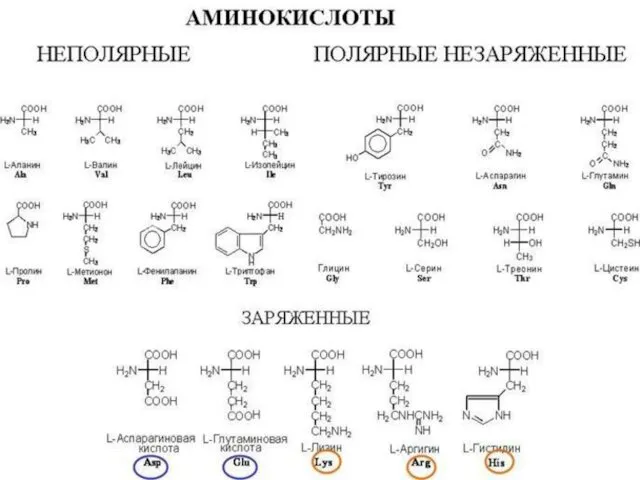
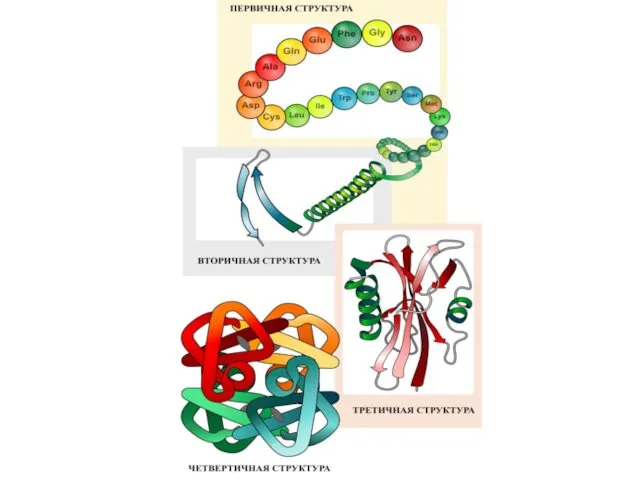
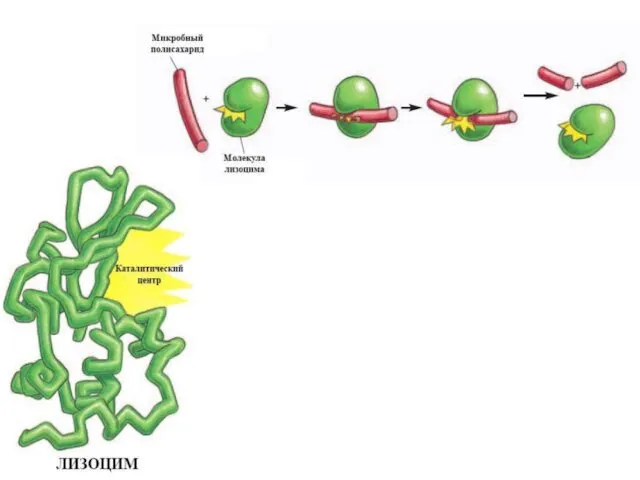
Центральная догма молекулярной биологии
ДНК
РНК
БЕЛОК
транскрипция
трансляция
ЭКСПРЕССИЯ ГЕНА
СВОЙСТВА
Центральная догма молекулярной биологии
ДНК
РНК
БЕЛОК
транскрипция
трансляция
ЭКСПРЕССИЯ ГЕНА
СВОЙСТВА
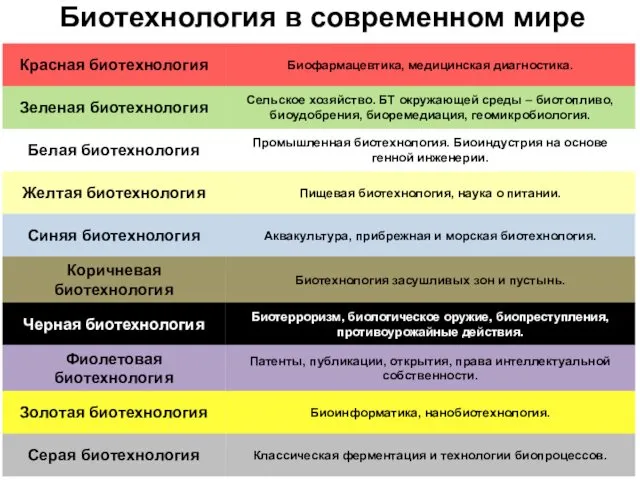
Биотехнология в современном мире
Биотехнология в современном мире

Предпосылки возникновения генной инженерии и история развития
Предпосылки возникновения генной инженерии и история развития
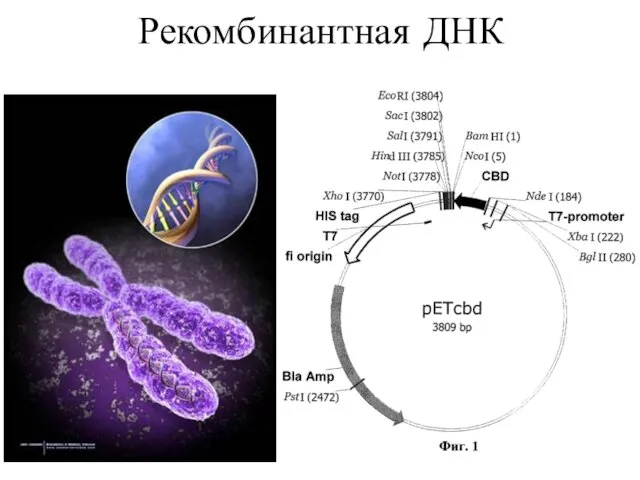
Рекомбинантная ДНК
Рекомбинантная ДНК
Ферменты, применяемые в генетической инженерии
Ферменты, применяемые в генетической инженерии

I. РЕСТРИКТАЗЫ (ЕС 3.1.21.) ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ)
I. РЕСТРИКТАЗЫ (ЕС 3.1.21.) ЭНДОНУКЛЕАЗЫ РЕСТРИКЦИИ)
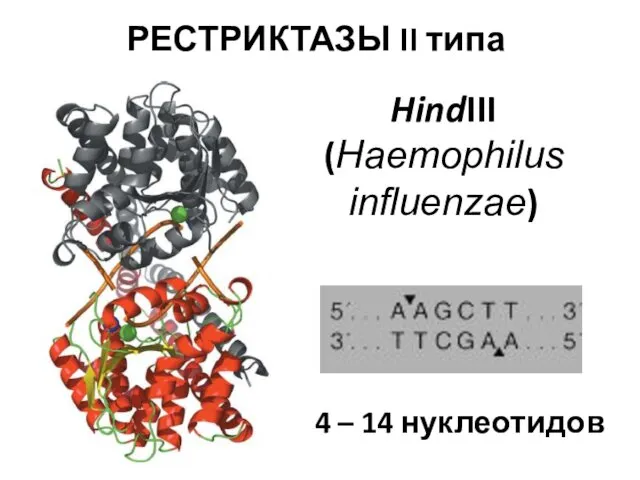
РЕСТРИКТАЗЫ II типа
HindIII
(Haemophilus influenzae)
4 – 14 нуклеотидов
РЕСТРИКТАЗЫ II типа
HindIII
(Haemophilus influenzae)
4 – 14 нуклеотидов

ПАЛИНДРОМЫ
А РОЗА УПАЛА НА ЛАПУ АЗОРА
По обеим цепям ДНК в одном
ПАЛИНДРОМЫ
А РОЗА УПАЛА НА ЛАПУ АЗОРА
По обеим цепям ДНК в одном

Рестриктазы в генной инженерии
EcoRI (E. coli RY13)
NdeI (Neisseria denitrificans)
BamHI (Bacillus amyloliquefaciens
Рестриктазы в генной инженерии
EcoRI (E. coli RY13)
NdeI (Neisseria denitrificans)
BamHI (Bacillus amyloliquefaciens
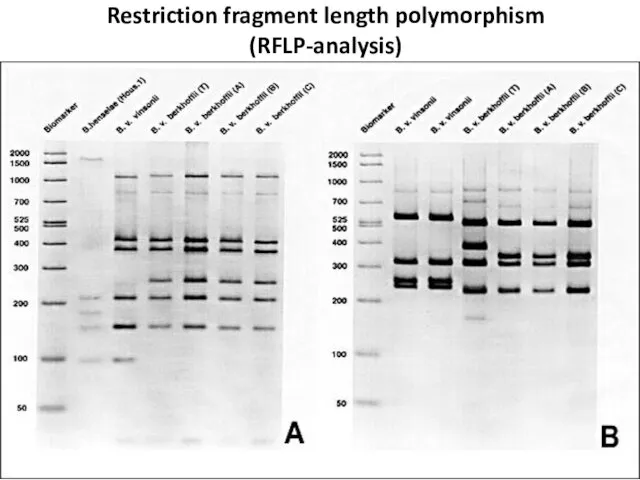
Restriction fragment length polymorphism (RFLP-analysis)
Restriction fragment length polymorphism (RFLP-analysis)

II. ЛИГАЗА (EC 6.5.1.1)
II. ЛИГАЗА (EC 6.5.1.1)
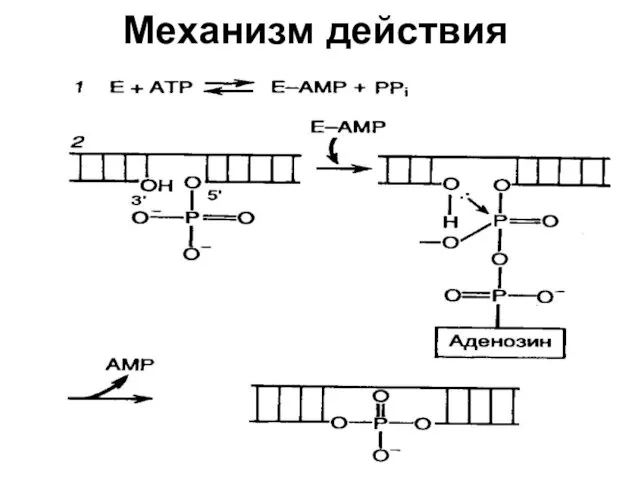
Механизм действия
Механизм действия
ДНК-лигаза фага Т4
Единица активности лигазы соответствует количеству фермента, необходимого для лигирования
ДНК-лигаза фага Т4
Единица активности лигазы соответствует количеству фермента, необходимого для лигирования
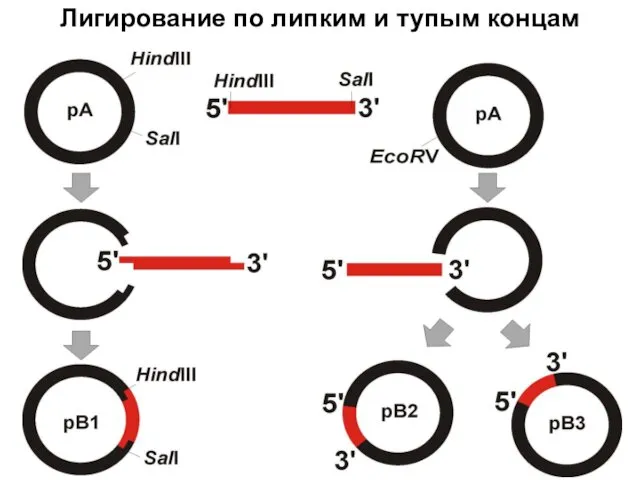
Лигирование по липким и тупым концам
Лигирование по липким и тупым концам

V. Нуклеазы – гидролитические ферменты, расщепляющие фосфодиэфирные связи в нуклеиновых кислотах
V. Нуклеазы – гидролитические ферменты, расщепляющие фосфодиэфирные связи в нуклеиновых кислотах
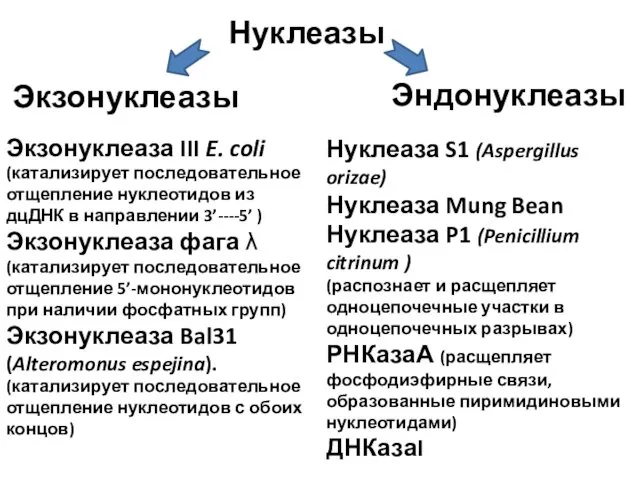
Нуклеазы
Экзонуклеазы
Эндонуклеазы
Экзонуклеаза III E. coli
(катализирует последовательное отщепление нуклеотидов
Нуклеазы
Экзонуклеазы
Эндонуклеазы
Экзонуклеаза III E. coli
(катализирует последовательное отщепление нуклеотидов
V. ДНК-полимеразы
V. ДНК-полимеразы
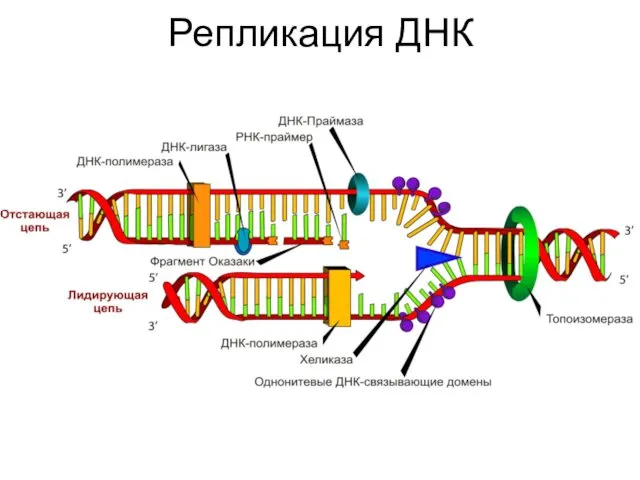
Репликация ДНК
Репликация ДНК
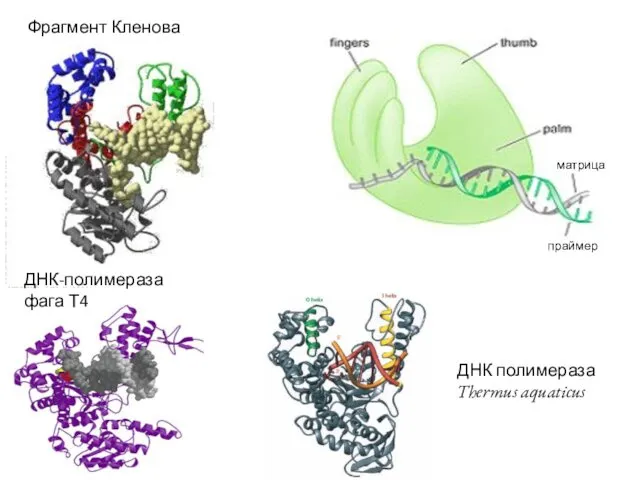
матрица
праймер
Фрагмент Кленова
ДНК-полимераза
фага Т4
ДНК полимераза
Thermus aquaticus
матрица
праймер
Фрагмент Кленова
ДНК-полимераза
фага Т4
ДНК полимераза
Thermus aquaticus

ДНК-полимеразы термофильных архебактерий
ДНК-полимеразы термофильных архебактерий

История открытия
Апрель 1983 г – идея ПЦР
Декабрь 1983 г – осуществление
История открытия
Апрель 1983 г – идея ПЦР
Декабрь 1983 г – осуществление

Что такое ПЦР?
По сути, это упрощенная версия репликации бактерий, при которой
Что такое ПЦР?
По сути, это упрощенная версия репликации бактерий, при которой
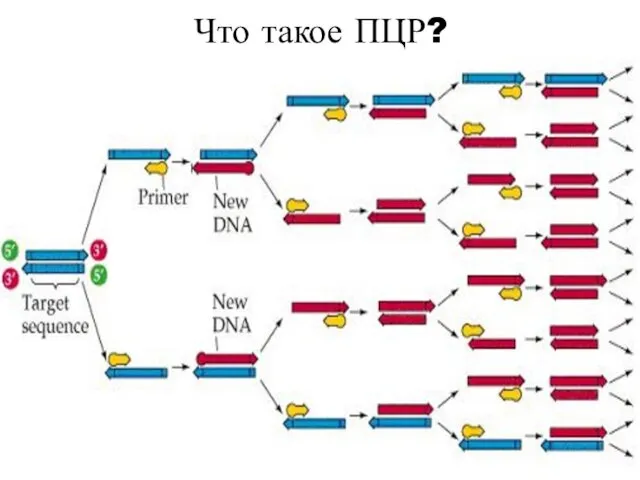
Что такое ПЦР?
Что такое ПЦР?

Сравнение ПЦР и репликации in vivo (1)
Сравнение ПЦР и репликации in vivo (1)

Сравнение ПЦР и репликации in vivo (2)
Сравнение ПЦР и репликации in vivo (2)
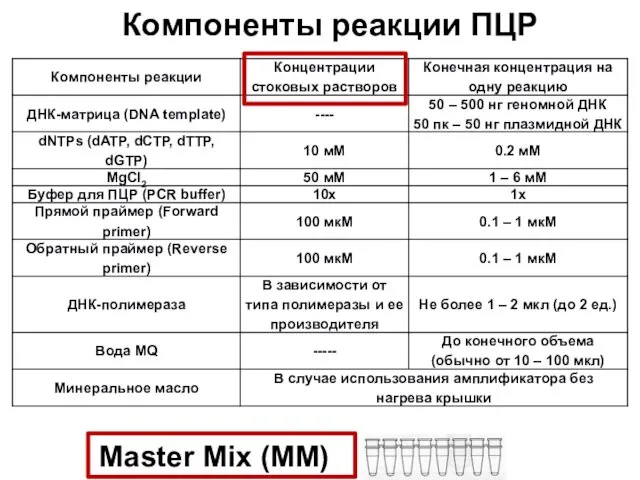
Компоненты реакции ПЦР
Master Mix (MM)
Компоненты реакции ПЦР
Master Mix (MM)
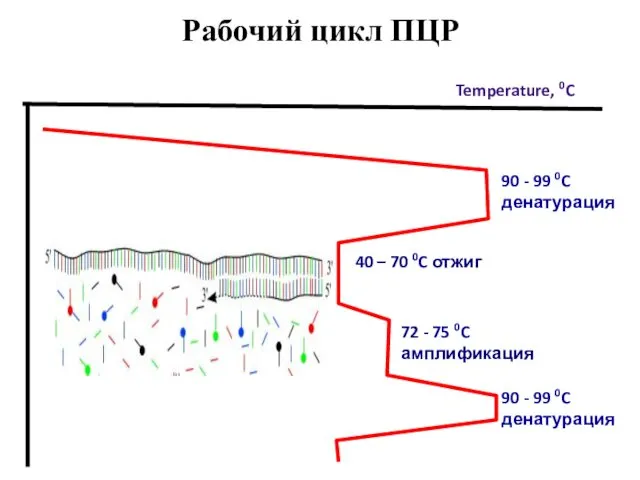
72 - 75 0C амплификация
90 - 99 0C
денатурация
Temperature, 0C
40 – 70
72 - 75 0C амплификация
90 - 99 0C
денатурация
Temperature, 0C
40 – 70
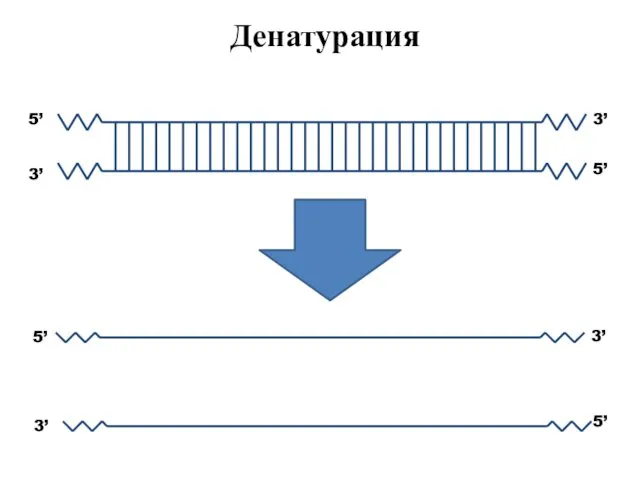
Денатурация
3’
5’
5’
3’
5’
3’
3’
5’
Денатурация
3’
5’
5’
3’
5’
3’
3’
5’
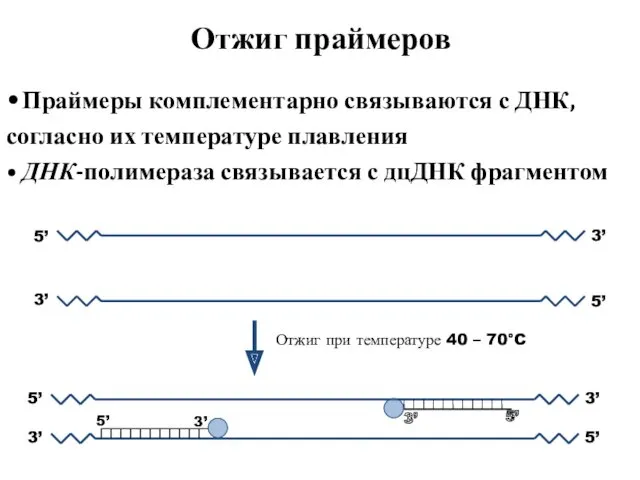
Отжиг праймеров
3’
5’
5’
3’
• Праймеры комплементарно связываются с ДНК,
согласно их температуре плавления
• ДНК-полимераза связывается
Отжиг праймеров
3’
5’
5’
3’
• Праймеры комплементарно связываются с ДНК,
согласно их температуре плавления
• ДНК-полимераза связывается

Температура плавления (T melting, Tm)
Под температурой плавления понимают температуру, при которой
Температура плавления (T melting, Tm)
Под температурой плавления понимают температуру, при которой

Расчет оптимальной температуры денатурации ампликона
Td=Tm+3-40С
Расчет оптимальной температуры денатурации ампликона
Td=Tm+3-40С

Температура отжига (T annealing, Ta)
Ta=Tm – 50С (по Ребрикову)
Ta=Tm – 100С
Температура отжига (T annealing, Ta)
Ta=Tm – 50С (по Ребрикову)
Ta=Tm – 100С

Задача №1
Определите температуры плавления и отжига следующего олигонуклеотида:
5’ GGCATTTAGCTTAGGC 3’
Решение:
N=16: А=3,
Задача №1
Определите температуры плавления и отжига следующего олигонуклеотида:
5’ GGCATTTAGCTTAGGC 3’
Решение:
N=16: А=3,

Задача №2
Определите температуры плавления и отжига следующего олигонуклеотида:
5’ GGCATTTAGCTTAGGCTTAAGGCGGGAG 3’
Решение:
N=28: А=6,
Задача №2
Определите температуры плавления и отжига следующего олигонуклеотида:
5’ GGCATTTAGCTTAGGCTTAAGGCGGGAG 3’
Решение:
N=28: А=6,
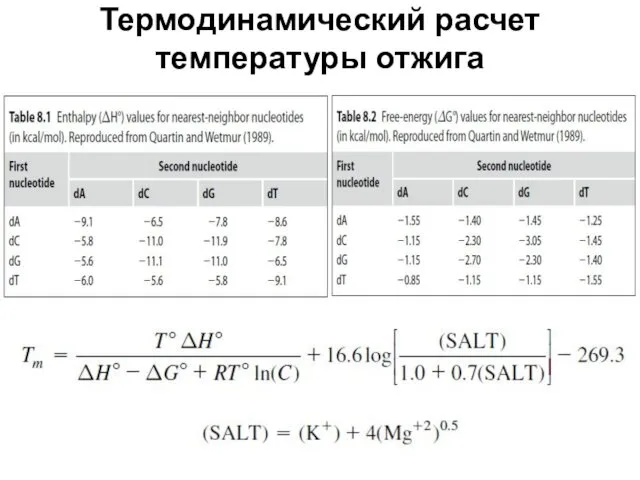
Термодинамический расчет температуры отжига
Термодинамический расчет температуры отжига

Конструирование (дизайн) праймеров:
При подборе праймеров зачастую удобно использовать програмное обеспечение,
Конструирование (дизайн) праймеров:
При подборе праймеров зачастую удобно использовать програмное обеспечение,
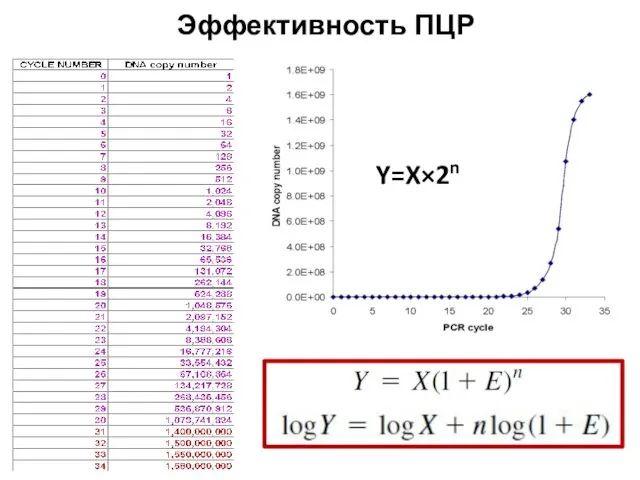
Эффективность ПЦР
Y=X×2n
Эффективность ПЦР
Y=X×2n

ПЦР в реальном времени
ПЦР в реальном времени
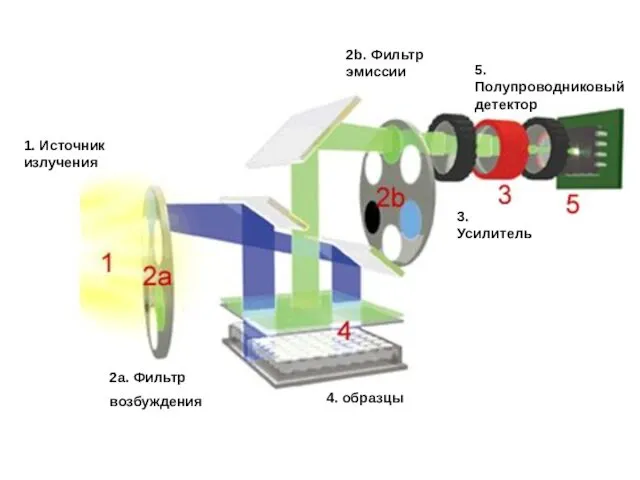
2a. Фильтр
возбуждения
2b. Фильтр эмиссии
1. Источник излучения
4. образцы
3. Усилитель
5. Полупроводниковый детектор
2a. Фильтр
возбуждения
2b. Фильтр эмиссии
1. Источник излучения
4. образцы
3. Усилитель
5. Полупроводниковый детектор
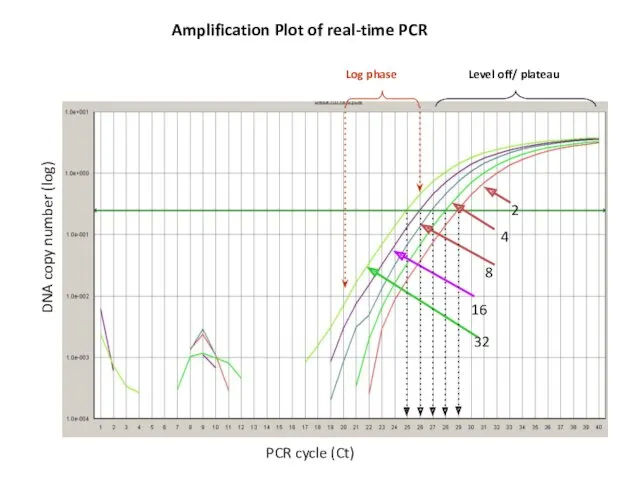
Amplification Plot of real-time PCR
DNA copy number (log)
PCR cycle (Ct)
Amplification Plot of real-time PCR
DNA copy number (log)
PCR cycle (Ct)
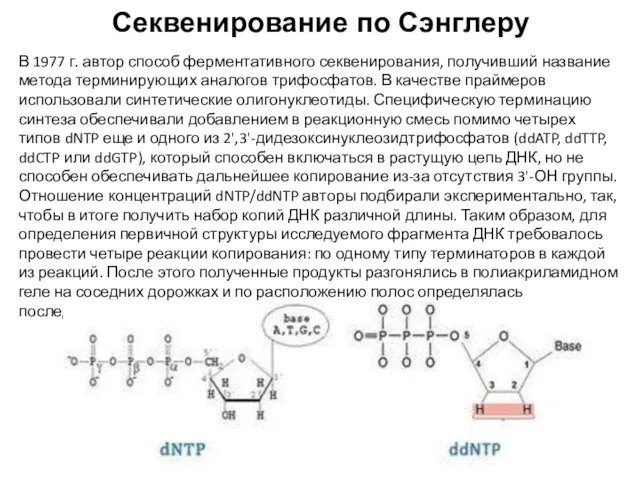
Секвенирование по Сэнглеру
В 1977 г. автор способ ферментативного секвенирования, получивший название
Секвенирование по Сэнглеру
В 1977 г. автор способ ферментативного секвенирования, получивший название
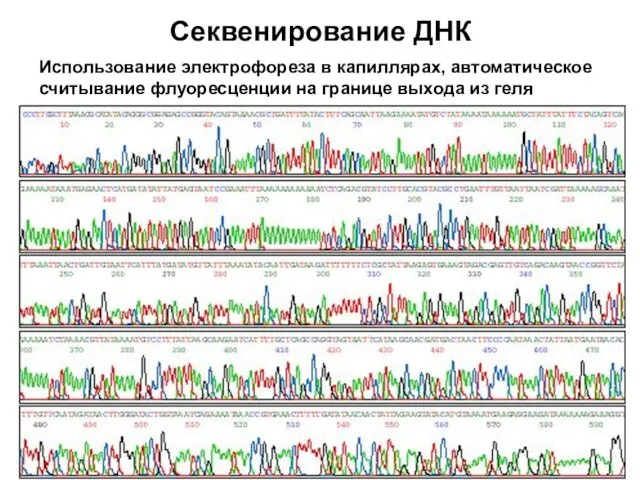
Секвенирование ДНК
Использование электрофореза в капиллярах, автоматическое считывание флуоресценции на границе выхода
Секвенирование ДНК
Использование электрофореза в капиллярах, автоматическое считывание флуоресценции на границе выхода
Обратная транскриптаза (КФ 2.7.7.49) (ревертаза)
Обратная транскриптаза (КФ 2.7.7.49) (ревертаза)
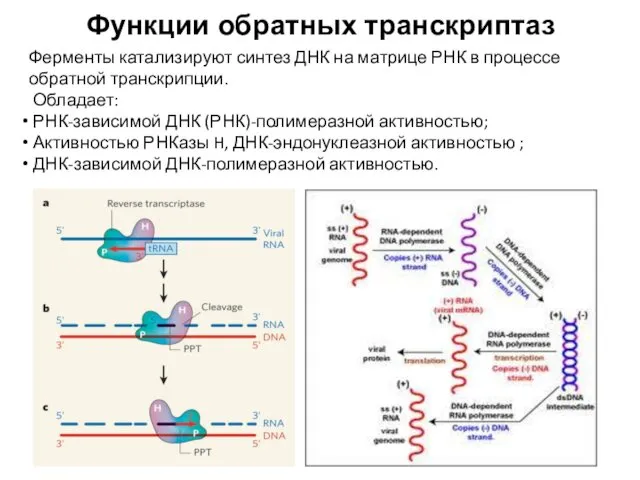
Функции обратных транскриптаз
Ферменты катализируют синтез ДНК на матрице РНК в процессе
Функции обратных транскриптаз
Ферменты катализируют синтез ДНК на матрице РНК в процессе

Обратные транскриптазы в генной инженерии
Обратная транскриптаза вируса миелобластоза птиц (Avian Myeloblastosis
Обратные транскриптазы в генной инженерии
Обратная транскриптаза вируса миелобластоза птиц (Avian Myeloblastosis
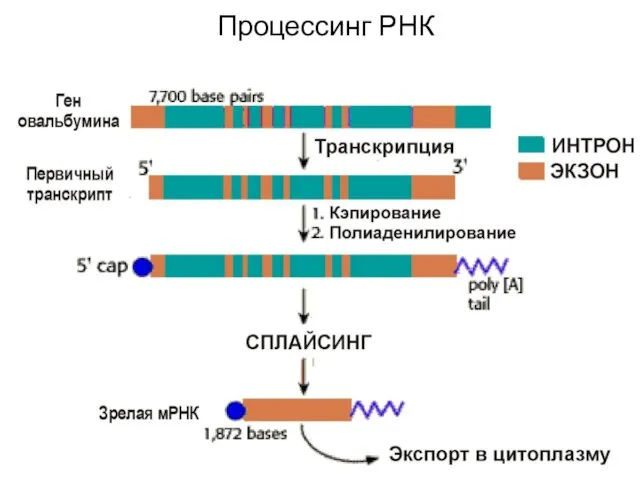
Процессинг РНК
Процессинг РНК
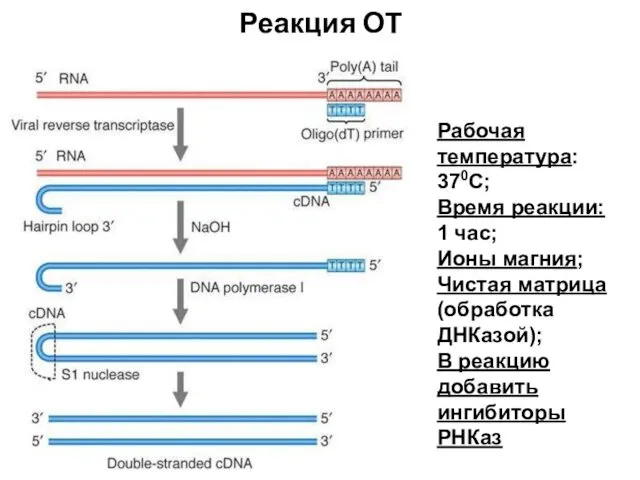
Реакция ОТ
Рабочая температура:
370С;
Время реакции:
1 час;
Ионы магния;
Чистая матрица
(обработка ДНКазой);
В реакцию добавить
ингибиторы РНКаз
Реакция ОТ
Рабочая температура:
370С;
Время реакции:
1 час;
Ионы магния;
Чистая матрица
(обработка ДНКазой);
В реакцию добавить
ингибиторы РНКаз

ДНК-полимераза Tth
Обратная транскрипция и ПЦР в одной пробирке !!!
ДНК-полимераза Tth
Обратная транскрипция и ПЦР в одной пробирке !!!
ВЕКТОРЫ
ВЕКТОРЫ

Что такое вектор?
Вектор – молекула ДНК, используемая в генетической инженерии для
Что такое вектор?
Вектор – молекула ДНК, используемая в генетической инженерии для

Свойства векторов
По функциям:
Векторы для клонирования
Векторы для экспрессии
Векторы для трансформации
По месту применения:
Бактериальные
Эукариотические
Челночные
Свойства векторов
По функциям:
Векторы для клонирования
Векторы для экспрессии
Векторы для трансформации
По месту применения:
Бактериальные
Эукариотические
Челночные
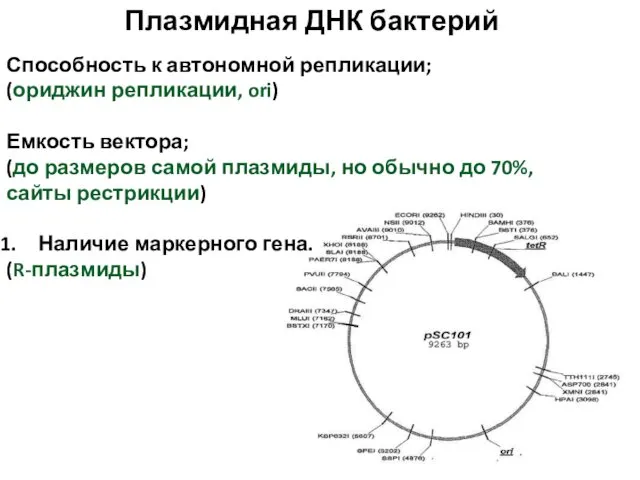
Плазмидная ДНК бактерий
Способность к автономной репликации;
(ориджин репликации, ori)
Емкость вектора;
(до размеров самой
Плазмидная ДНК бактерий
Способность к автономной репликации;
(ориджин репликации, ori)
Емкость вектора;
(до размеров самой
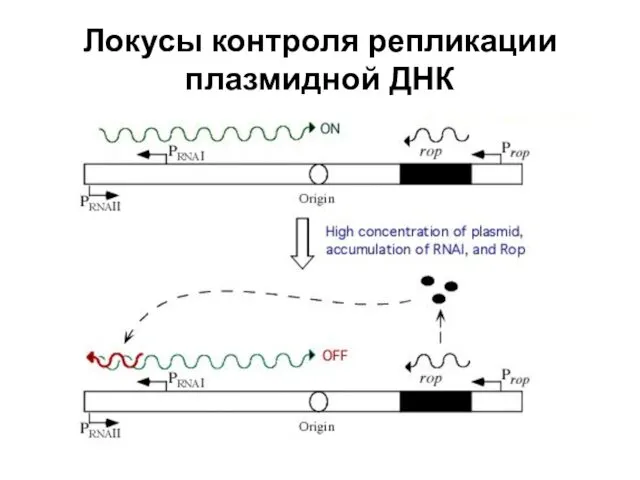
Локусы контроля репликации плазмидной ДНК
Локусы контроля репликации плазмидной ДНК

Создание плазмид с повышенной копийностью
_____________________________________________
Плазмида Ori Копийность Классификация
_____________________________________________
pUC pMB1* 500 – 700 высококопийная
pBluescript ColE1 300
Создание плазмид с повышенной копийностью
_____________________________________________
Плазмида Ori Копийность Классификация
_____________________________________________
pUC pMB1* 500 – 700 высококопийная
pBluescript ColE1 300
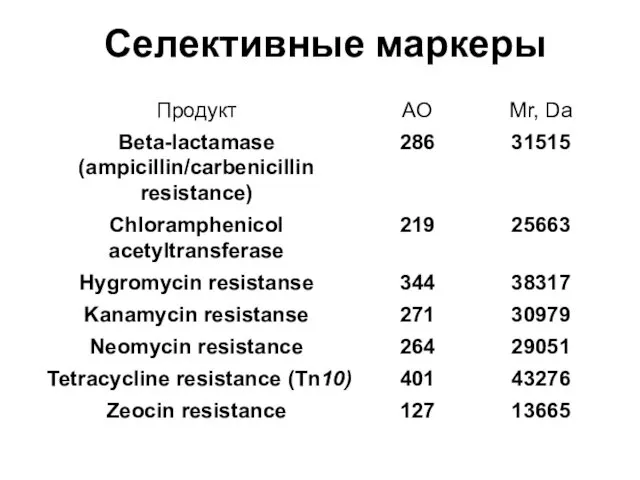
Селективные маркеры
Селективные маркеры
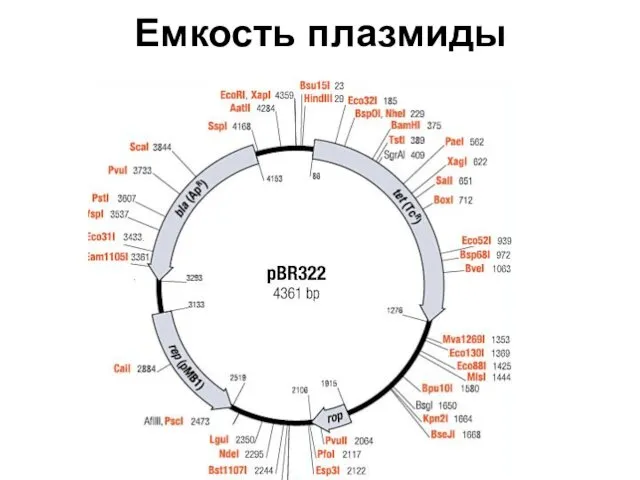
Емкость плазмиды
Емкость плазмиды
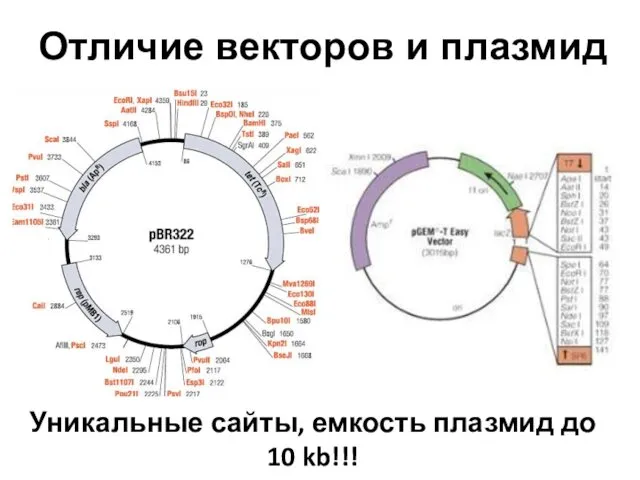
Отличие векторов и плазмид
Уникальные сайты, емкость плазмид до 10 kb!!!
Отличие векторов и плазмид
Уникальные сайты, емкость плазмид до 10 kb!!!
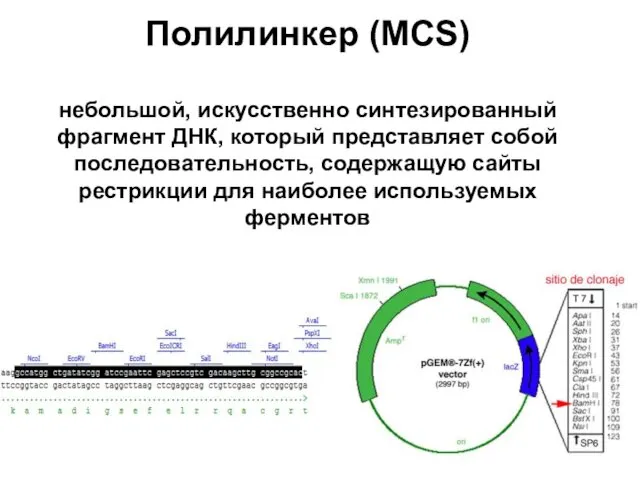
Полилинкер (MCS)
небольшой, искусственно синтезированный фрагмент ДНК, который представляет собой последовательность, содержащую
Полилинкер (MCS) небольшой, искусственно синтезированный фрагмент ДНК, который представляет собой последовательность, содержащую
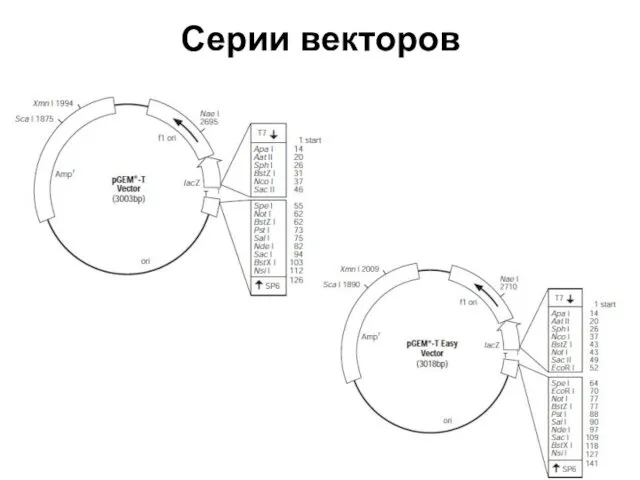
Серии векторов
Серии векторов
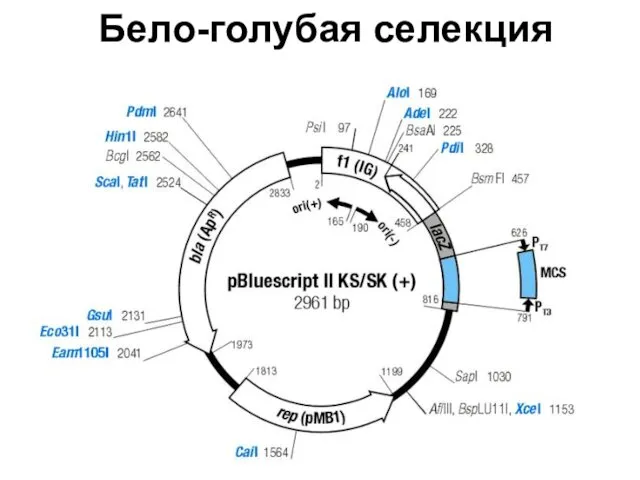
Бело-голубая селекция
Бело-голубая селекция
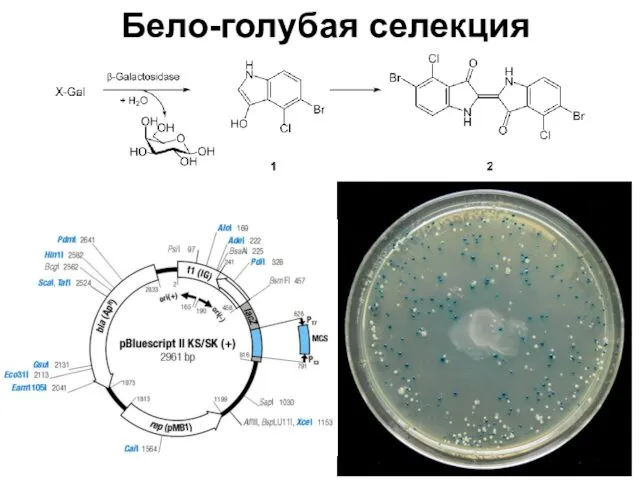
Бело-голубая селекция
ИПТГ – индуктор lac-оперона
X-Gal – хромогенный субстрат
Бело-голубая селекция
ИПТГ – индуктор lac-оперона
X-Gal – хромогенный субстрат
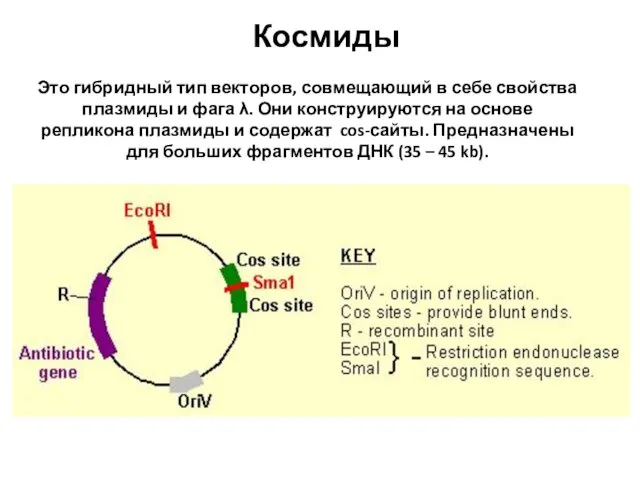
Космиды
Это гибридный тип векторов, совмещающий в себе свойства плазмиды и
Космиды
Это гибридный тип векторов, совмещающий в себе свойства плазмиды и
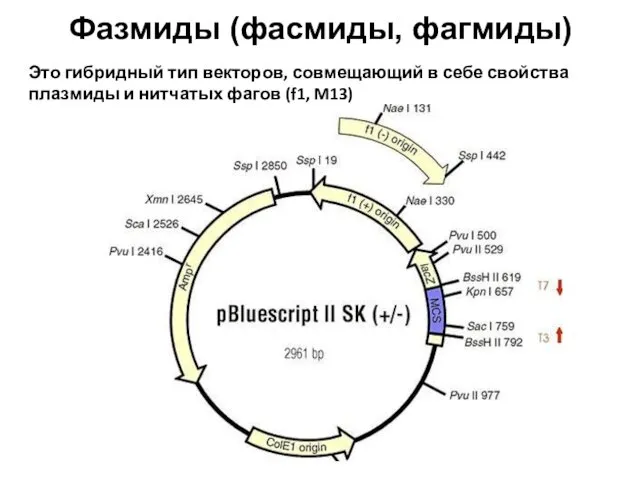
Фазмиды (фасмиды, фагмиды)
Это гибридный тип векторов, совмещающий в себе свойства плазмиды
Фазмиды (фасмиды, фагмиды)
Это гибридный тип векторов, совмещающий в себе свойства плазмиды
Искусственные хромосомы
Искусственные хромосомы
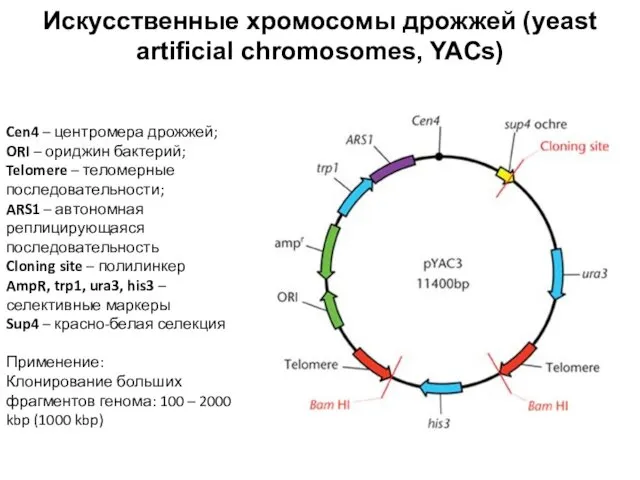
Искусственные хромосомы дрожжей (yeast artificial chromosomes, YACs)
Cen4 – центромера дрожжей;
ORI –
Искусственные хромосомы дрожжей (yeast artificial chromosomes, YACs)
Cen4 – центромера дрожжей;
ORI –

Создание геномных библиотек
Создание геномных библиотек

Геномные библиотеки (банки генов)
Геномная библиотека – фрагменты генома, клонированные в фаге
Геномные библиотеки (банки генов)
Геномная библиотека – фрагменты генома, клонированные в фаге

Принцип конструирования геномных библиотек
Принцип конструирования геномных библиотек
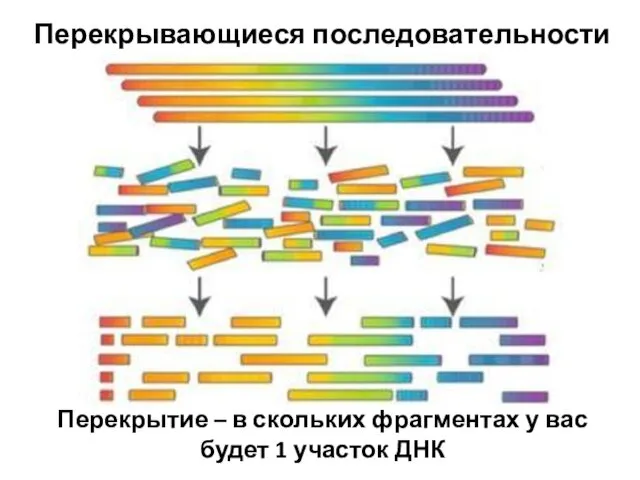
Перекрывающиеся последовательности
Перекрытие – в скольких фрагментах у вас будет 1
Перекрывающиеся последовательности
Перекрытие – в скольких фрагментах у вас будет 1
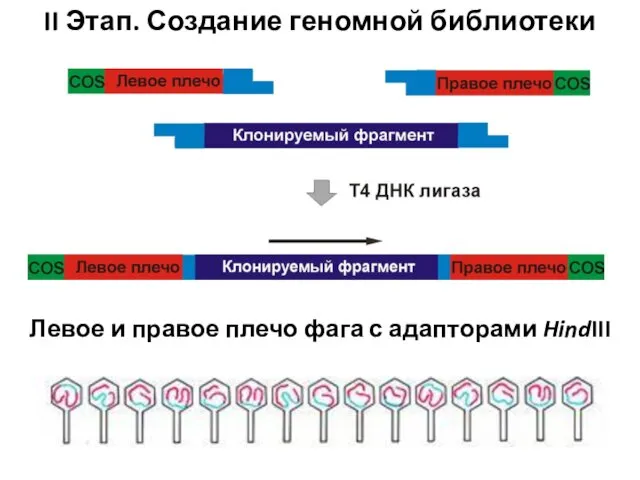
II Этап. Создание геномной библиотеки
Левое и правое плечо фага с адапторами
II Этап. Создание геномной библиотеки
Левое и правое плечо фага с адапторами
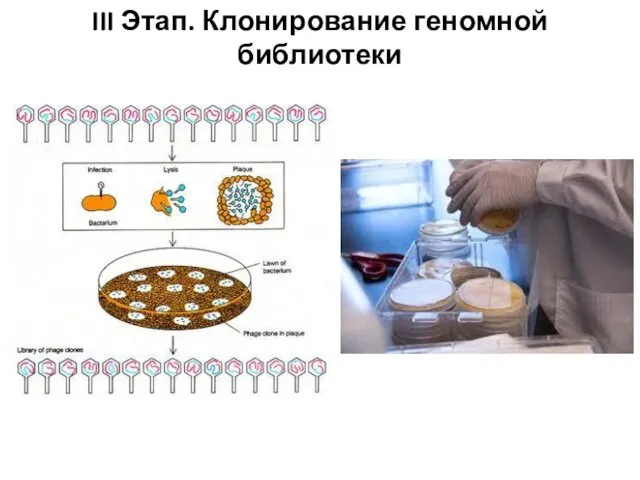
III Этап. Клонирование геномной библиотеки
III Этап. Клонирование геномной библиотеки

Расчет количества клонов, необходимых для получения геномной библиотеки
Где N – количество
Расчет количества клонов, необходимых для получения геномной библиотеки
Где N – количество
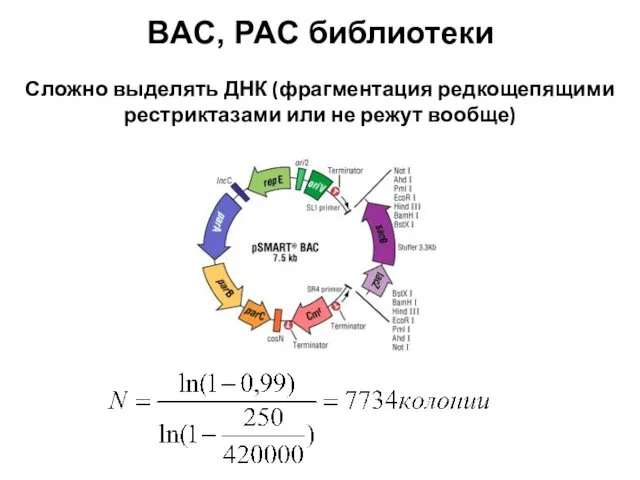
BAC, PAC библиотеки
Сложно выделять ДНК (фрагментация редкощепящими рестриктазами или не режут
BAC, PAC библиотеки
Сложно выделять ДНК (фрагментация редкощепящими рестриктазами или не режут
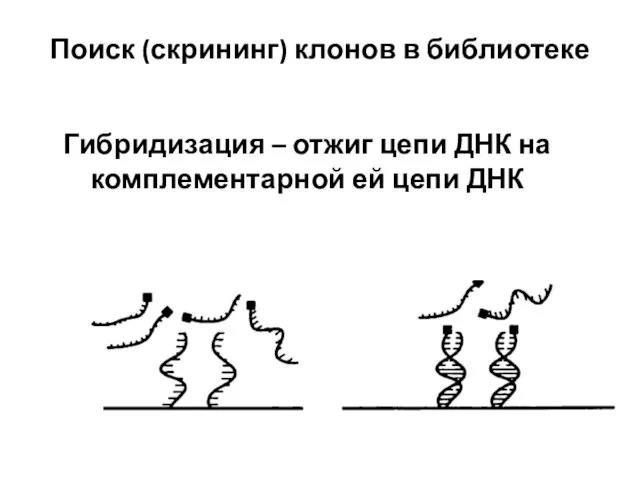
Поиск (скрининг) клонов в библиотеке
Гибридизация – отжиг цепи ДНК на комплементарной
Поиск (скрининг) клонов в библиотеке
Гибридизация – отжиг цепи ДНК на комплементарной
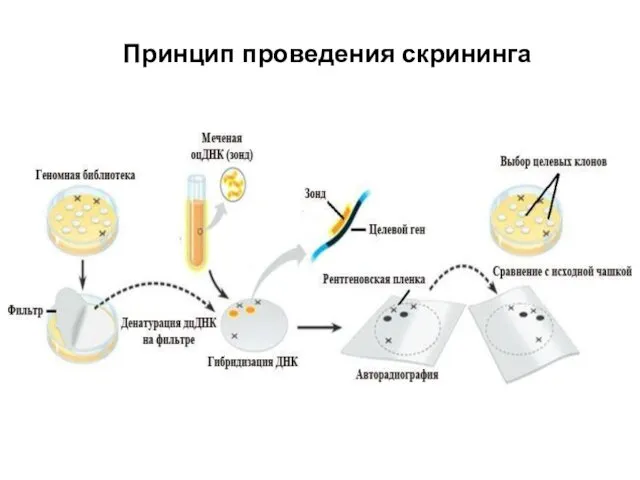
Принцип проведения скрининга
Принцип проведения скрининга
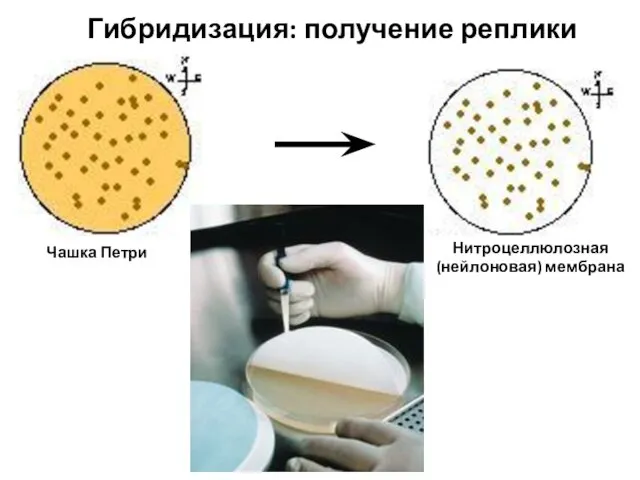
Гибридизация: получение реплики
Чашка Петри
Нитроцеллюлозная
(нейлоновая) мембрана
Гибридизация: получение реплики
Чашка Петри
Нитроцеллюлозная
(нейлоновая) мембрана
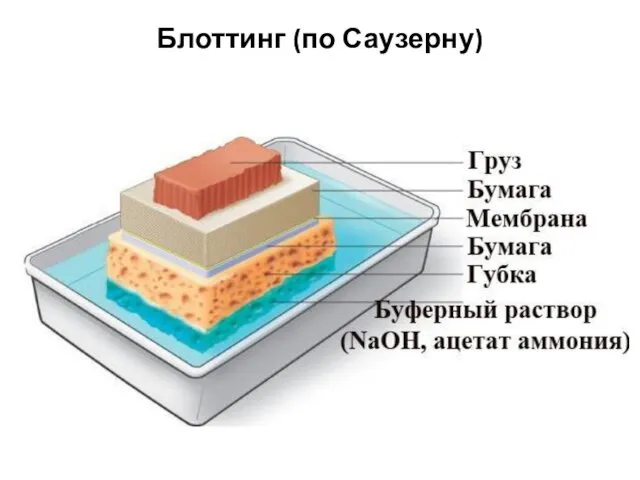
Блоттинг (по Саузерну)
Блоттинг (по Саузерну)
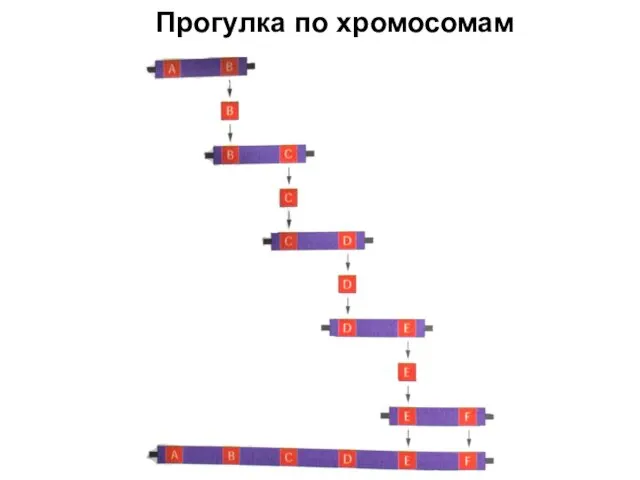
Прогулка по хромосомам
Прогулка по хромосомам
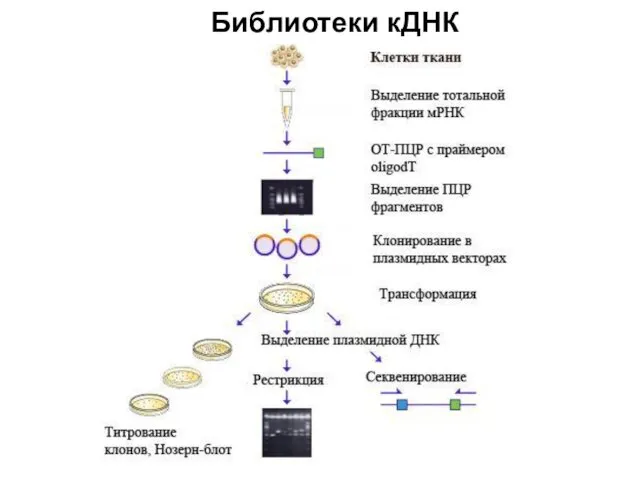
Библиотеки кДНК
Библиотеки кДНК

ТРАНСГЕННЫЕ РАСТЕНИЯ
ТРАНСГЕННЫЕ РАСТЕНИЯ

50-е годы XX века
Получение культур in vitro
- растений «в пробирке»
50-е годы XX века
Получение культур in vitro
- растений «в пробирке»

1983 год, первое трансгенное растение
Растение табака, устойчивое к канамицину
1983 год, первое трансгенное растение
Растение табака, устойчивое к канамицину

1990 год, первое трансгенное растение, пошедшее «в поля»
Растения хлопка, устойчивые к
1990 год, первое трансгенное растение, пошедшее «в поля»
Растения хлопка, устойчивые к

1994 год, томаты «flavr savr»
Устойчивость к
бактериальным гнилям
1994 год, томаты «flavr savr»
Устойчивость к
бактериальным гнилям

1995 год, Monsanto
Соя, устойчивая к гербицидам
1995 год, Monsanto
Соя, устойчивая к гербицидам

Картофель, устойчивый к колородскому жуку
Кукуруза, устойчивая к кукурузной огневке, гербицидам
Картофель, устойчивый к колородскому жуку
Кукуруза, устойчивая к кукурузной огневке, гербицидам

Suntory – голубая трансгенная роза (дигидрокверцитин 5’-гидролаза)
Suntory – голубая трансгенная роза (дигидрокверцитин 5’-гидролаза)

- трансформация клеток
Как получают трансгенные растения?
- Получение культур
- трансформация клеток
Как получают трансгенные растения?
- Получение культур
Агробактериальная трансформация
Agrobacterium tumefaciens
(A. tumefaciens)
Агробактериальная трансформация
Agrobacterium tumefaciens
(A. tumefaciens)

A. tumefaciens вызывает болезнь корончатых галлов
A. tumefaciens вызывает болезнь корончатых галлов

Корончатые галлы состоят из
дедифференцированных клеток
Корончатые галлы состоят из
дедифференцированных клеток
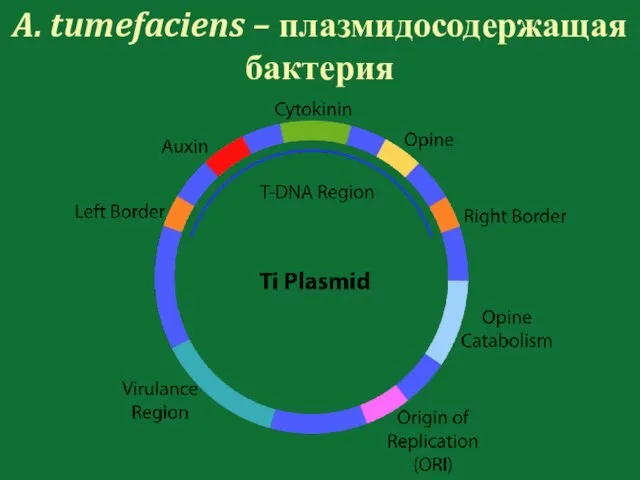
A. tumefaciens – плазмидосодержащая бактерия
A. tumefaciens – плазмидосодержащая бактерия
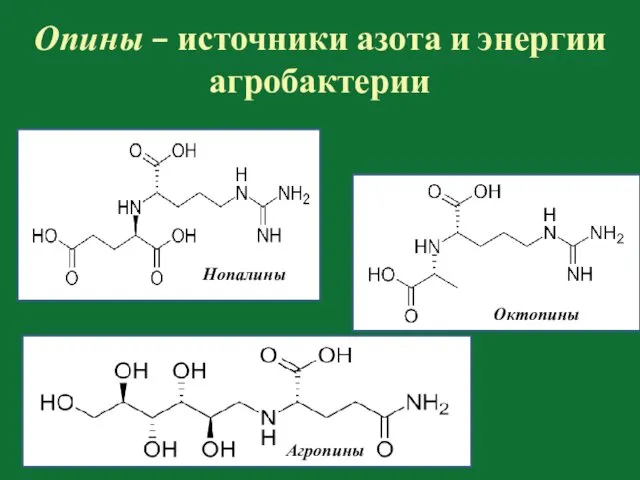
Опины – источники азота и энергии агробактерии
Нопалины
Октопины
Агропины
Опины – источники азота и энергии агробактерии
Нопалины
Октопины
Агропины
(локус shi)
(локус roi)
(локус shi)
(локус roi)
Агробактериальная трансформация
Агробактериальная трансформация
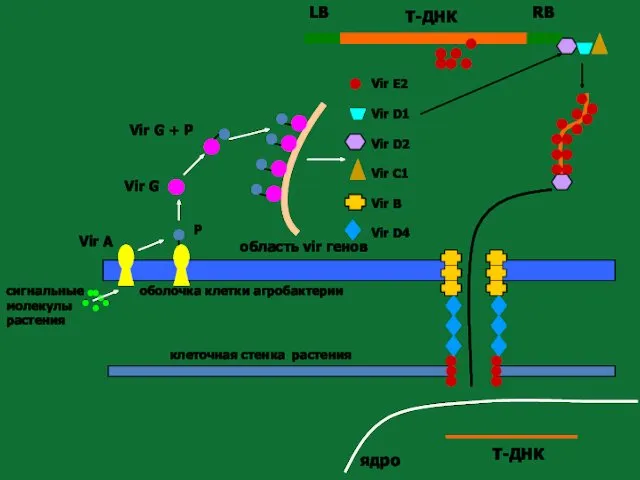
клеточная стенка растения
оболочка клетки агробактерии
Vir A
сигнальные молекулы растения
P
Vir G
Vir G +
клеточная стенка растения
оболочка клетки агробактерии
Vir A
сигнальные молекулы растения
P
Vir G
Vir G +

Влияние растительных гормонов на клетку
Влияние растительных гормонов на клетку


Оценка устойчивости трансгенных линий сахарной свеклы к действию гербицида «Баста»
КОНТРОЛЬ
ТРАНСГЕННЫЕ ЛИНИИ
ТРАНСГЕННЫЕ
ЛИНИИ
ТРАНСГЕННЫЕ
Оценка устойчивости трансгенных линий сахарной свеклы к действию гербицида «Баста»
КОНТРОЛЬ
ТРАНСГЕННЫЕ ЛИНИИ
ТРАНСГЕННЫЕ
ЛИНИИ
ТРАНСГЕННЫЕ

Получение трансгенного картофеля
Получение трансгенного картофеля

Участок, зарегистрированный МВК ГИД, Краснодар, ВНИИБЗР
Контрольные растения
ГМ сорт
Невский
ГМ сорт Луговской
Участок, зарегистрированный МВК ГИД, Краснодар, ВНИИБЗР
Контрольные растения
ГМ сорт
Невский
ГМ сорт Луговской

Контрольные растения, 150 mM NaCl
Трансгенные растения, 150 mM NaCl
Контрольные растения
Трансгенные
Контрольные растения, 150 mM NaCl
Трансгенные растения, 150 mM NaCl
Контрольные растения
Трансгенные

РИС
(Oryza sativa )
Основной пищевой продукт, производимый в мире
Низкое содержание витамина
РИС
(Oryza sativa )
Основной пищевой продукт, производимый в мире
Низкое содержание витамина

“Золотой” рис с повышенным
содержанием β -каротина
“Золотой” рис с повышенным
содержанием β -каротина
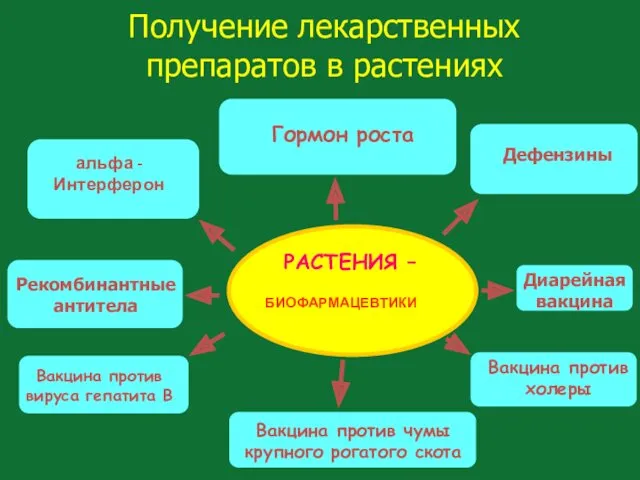
Получение лекарственных препаратов в растениях
РАСТЕНИЯ –
БИОФАРМАЦЕВТИКИ
Получение лекарственных препаратов в растениях
РАСТЕНИЯ –
БИОФАРМАЦЕВТИКИ
 Cargo transportation by road transport
Cargo transportation by road transport Бензоэлектрический агрегат
Бензоэлектрический агрегат Правила конструктивного общения
Правила конструктивного общения Презентация Предметно-развивающая среда группы раннего возраста
Презентация Предметно-развивающая среда группы раннего возраста Патология органов речи
Патология органов речи Словосочетание. Виды словосочетаний
Словосочетание. Виды словосочетаний Топографическая анатомия и оперативная хирургия желудка
Топографическая анатомия и оперативная хирургия желудка Игра Автоматизация звука [Л]
Игра Автоматизация звука [Л] Специфика труда и рынка труда в современном обществе
Специфика труда и рынка труда в современном обществе Публицистический стиль. Функции языка, реализуемые в публицистических текстах. Лексика и фразеология
Публицистический стиль. Функции языка, реализуемые в публицистических текстах. Лексика и фразеология Монастыри Православной Церкви в Америке
Монастыри Православной Церкви в Америке Теория производства
Теория производства Создание Web-страниц
Создание Web-страниц Изобретения в XX-XXI веках
Изобретения в XX-XXI веках Шаблоны презентаций - 9
Шаблоны презентаций - 9 Проект Костюм рыцаря
Проект Костюм рыцаря Теория социального характера Д. Рисмена
Теория социального характера Д. Рисмена Різдвяний вертеп
Різдвяний вертеп Е. Чарушин Теремок
Е. Чарушин Теремок Таблица сложения. Тренажёр Игра в футбол
Таблица сложения. Тренажёр Игра в футбол Возможности электронного журнала
Возможности электронного журнала Символы России. 5 класс
Символы России. 5 класс Архитектура ПК
Архитектура ПК Конструкции котлов. Лекция 4
Конструкции котлов. Лекция 4 Презентация Конкурс пословиц о матери
Презентация Конкурс пословиц о матери Вальдорфская педагогика раннего развития
Вальдорфская педагогика раннего развития Внешняя политика СССР в 30-е годы ХХ века
Внешняя политика СССР в 30-е годы ХХ века Методы измерения и приборное обеспечение радиационно-экологического мониторинга. (Лекция 6)
Методы измерения и приборное обеспечение радиационно-экологического мониторинга. (Лекция 6)