- Клеточный цикл, репарация и рекомбинация. Лекция 2

Содержание
- 2. Каждая клетка происходит от родительской клетки. Клетки воспроизводятся, проходя через ряд упорядоченных событий, в результате которых
- 4. G1 G2
- 5. Перенос цитоплазмы из митотической клетки в интерфазную стимулирует вхождение интерфазной клетки в митоз MPF - Maturation
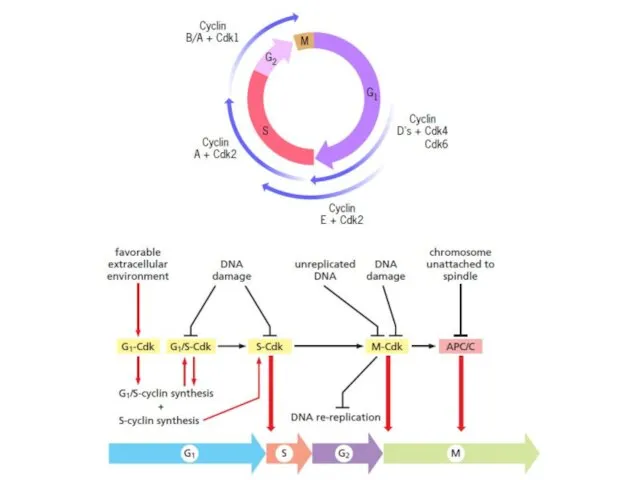
- 6. Cell Cycle Checkpoints… Gap 1 Gap 2 Продвижение по клеточному циклу регулируется на разных стадиях.
- 7. Cdk - cyclin-dependent kinases Saccharomyces cerevisiae (budding yeast) Schizosaccharomyces pombe (fission yeast) Leland Hartwell (University of
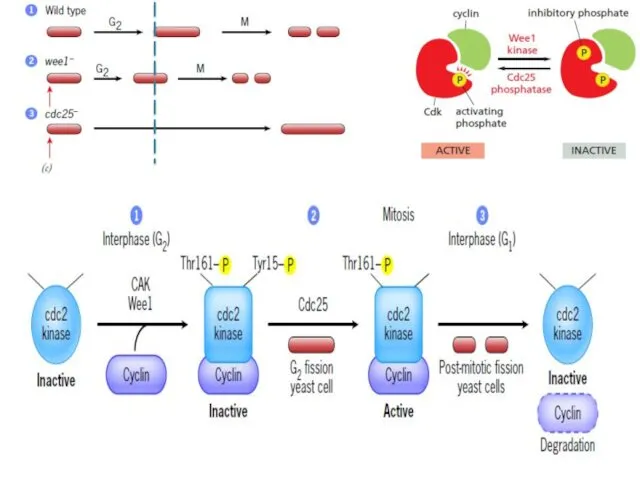
- 8. (Restriction point R1) A simplified model for cell cycle regulation in fission yeast
- 10. Cdk Phosphorylation/dephosphorylation Cdk2 and cyclin A
- 12. The inhibition of a cyclin–Cdk complex by a CKI (Cdk inhibitor proteins) Сyclin A–Cdk2 complex связанный
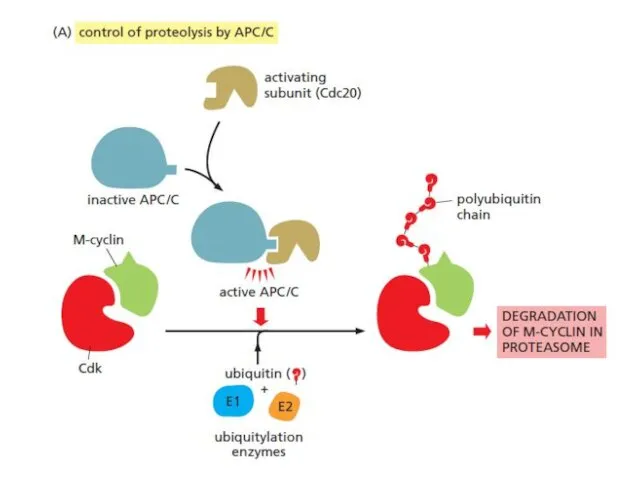
- 13. metaphase-to-anaphase transition Anaphase promoting complex, or cyclosome (APC/C) = ubiquitin ligase Продвижение через R1 (START) и
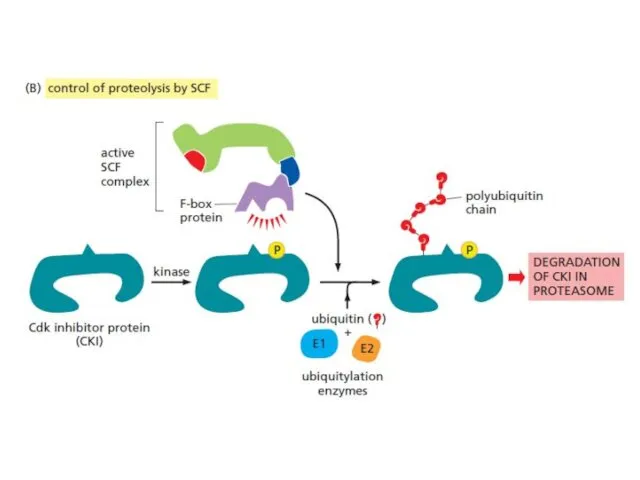
- 14. SCF убиквитинирует: Определенные CKI в поздней G1, что позволяет активировать S-Cdk’s и запустить репликацию DNA. SCF
- 18. S PHASE Geminin = inhibitor of cdt1
- 19. S PHASE (continuation)
- 20. Cohesin SMC proteins (Structural Maintenance of Chromosomes) Scc1 и Scc3 – дополнительные субъединицы, связывающие ATPase-ные домены
- 21. MITOSIS Cyclin B in vertebrate
- 22. MITOSIS: Condensins Фосфориллирование конденсинов комплексом Cyclin B – Cdk активирует их и стимулирует образование петлевых структур
- 23. Mitotic spindle
- 24. centrosome
- 25. Major motor proteins of the spindle
- 26. Полуконсервативный механизм дупликации центросом
- 27. Kinetochore polymerization and depolymerization of the microtubule
- 28. Kinetochore Вначале происходит латеральный захват микротрубочек, а затем при содействии кинезиновых моторов микротрубочки ориентируются в правильное
- 29. Kinetochore: Bi-orientation Low tension Low tension High tension
- 30. Kinetochore: Bi-orientation Ингибиторный сигнал ослабляет захват микротрубочек Ингибиторного сигнала нет, захват микротрубочек усиливается, привлекаются новые микротрубочки.
- 31. Сила, направляющая кинетохор к полюсу (разборка микротрубочек в месте прикрепления к кинетохору). К полюсу Движение самих
- 32. Флуоресцентно меченый тубулин Microtubule flux
- 33. polar ejection force A. Эксперимент с разрезанием лазером B. Разъясняющая «полюсный ветер» модель
- 34. Расхождение сестринских хроматид Активный APC/C комплекс так же разрушает и М-циклины (отрицательная обратная связь)
- 35. Mad2 белок является проверочным механизмом присоединения к кинетохору трубочек. Если напряжения на микротрубочках, связанных с кинетохором
- 36. Разборка веретена; Фрагменты ядерной мембраны ассоциируются с отдельными хромосомами и затем соединяются друг с другом; комплекс
- 37. CYTOKINESIS Обычно митоз заканчивается цитокинезом. Некоторые клетки (ранние эмбрионы дрозофилы, гепатоциты и кардиомиоциты) проходят митоз без
- 38. CYTOKINESIS a cleaving frog egg
- 39. The MIDBODY
- 40. Активация RhoA включает сборку и сокращение контрактильного кольца
- 41. Микротрубочки веретена деления определяют время и место разделения клетки Три модели, объясняющие, как микротрубочки веретена дают
- 42. Некоторые клетки делятся ассиметрично, что приводит к их развитию по разным путям. Материнская клетка перед делением
- 43. Многоядерные клетки Многоядерная клетка называется синцитиум. У человека мегакариоциты (предшественники тромбоцитов), некоторые гепатоциты, а так же
- 44. М-cdk активирует cdc20-APC/C, он в свою очередь дезактивирует М-циклины, а это ведет к дезактивации самого APC/C,
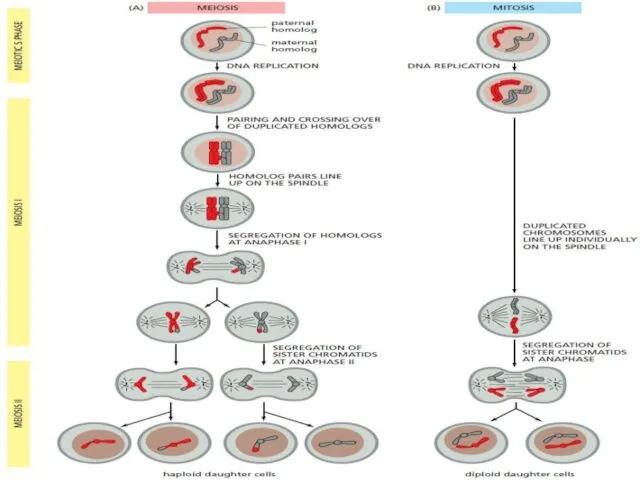
- 45. MEIOSIS
- 47. Спаривание хромосом в профазе мейоза 1 Кроссинговер в профазе мейоза 1
- 48. Synaptonemal Complex presynaptic alignment synapsis ds DNA breaks recombination complex
- 49. Prophase I
- 50. После разборки синаптонемального комплекса в диплотене и следующей за ним конденсации хроматид могут быть увидены перекрёсты.
- 51. 1. Оба сестринских кинетохора в гомологе должны быть соединены с ОДНИМ и тем же полюсом веретена.
- 53. Кроссинговер тщательно регулируется: на каждую гомологичную пару хромосом приходится не менее одного (поскольку гомологи должны удерживаться
- 54. Контроль клеточного деления и роста Внеклеточные сигналы, регулирующие клеточный рост, деление и «выживание», могут быть разделены
- 55. PDGF (platelet-derived growth factor) – ростовой фактор с широкой специфичностью; EGF (epidermal growth factor); Erythropoietin –
- 56. Митогены действуют в основном в G1
- 57. Retinoblastoma protein
- 60. Клетки имеют ограниченное число делений (предел Хейфлика). После этого предела клетки входят в неделящееся состояние, из
- 61. или Ras
- 63. Скачать презентацию
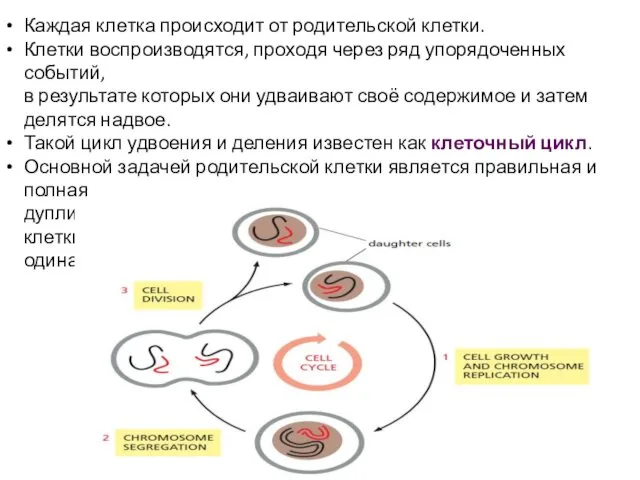
Каждая клетка происходит от родительской клетки.
Клетки воспроизводятся, проходя через ряд упорядоченных
Каждая клетка происходит от родительской клетки.
Клетки воспроизводятся, проходя через ряд упорядоченных
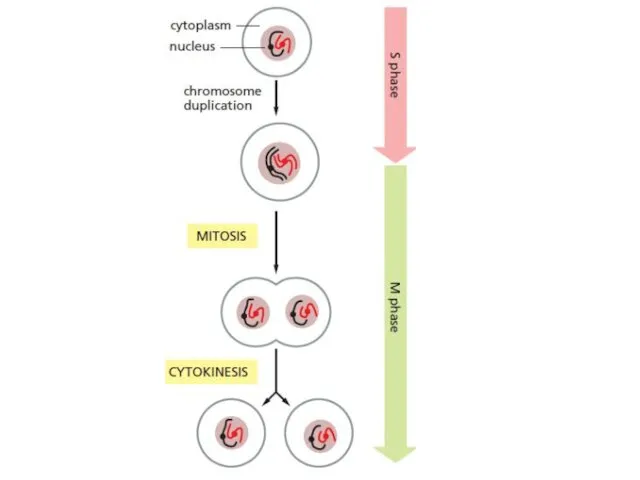
G1
G2
G1
G2
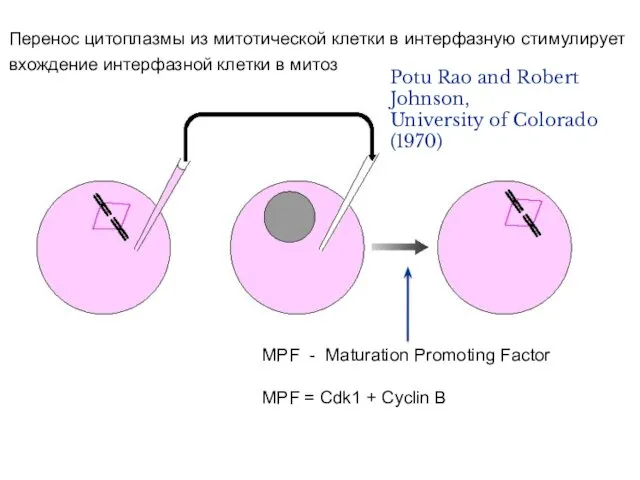
Перенос цитоплазмы из митотической клетки в интерфазную стимулирует
вхождение интерфазной клетки в
Перенос цитоплазмы из митотической клетки в интерфазную стимулирует
вхождение интерфазной клетки в
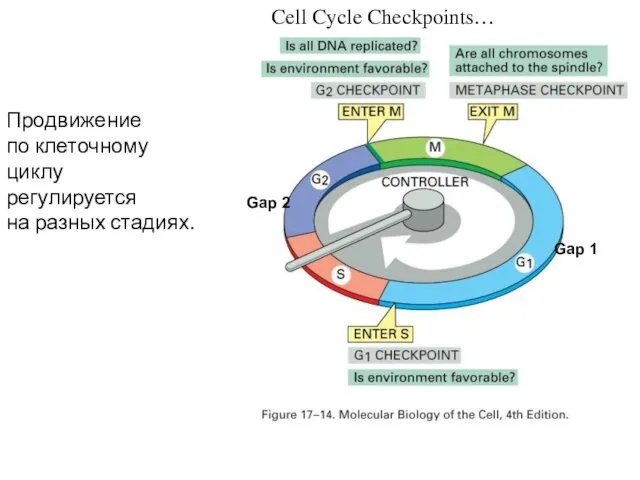
Cell Cycle Checkpoints…
Gap 1
Gap 2
Продвижение
по клеточному циклу
регулируется
на разных
Cell Cycle Checkpoints…
Gap 1
Gap 2
Продвижение по клеточному циклу регулируется на разных

Cdk - cyclin-dependent kinases
Saccharomyces cerevisiae (budding yeast)
Schizosaccharomyces pombe (fission yeast)
Leland Hartwell
Cdk - cyclin-dependent kinases
Saccharomyces cerevisiae (budding yeast)
Schizosaccharomyces pombe (fission yeast)
Leland Hartwell
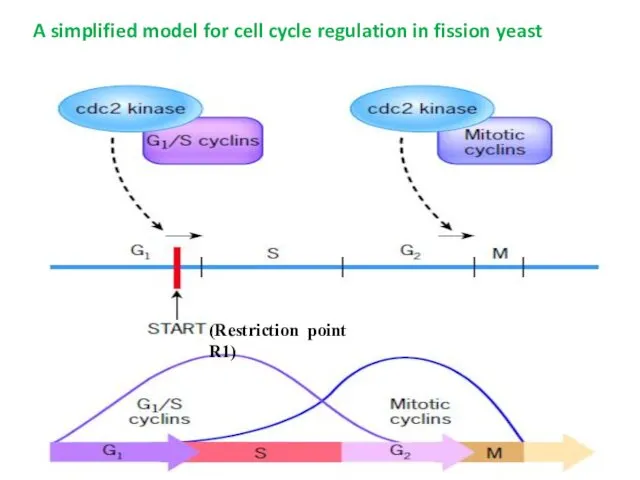
(Restriction point R1)
A simplified model for cell cycle regulation in fission
(Restriction point R1)
A simplified model for cell cycle regulation in fission
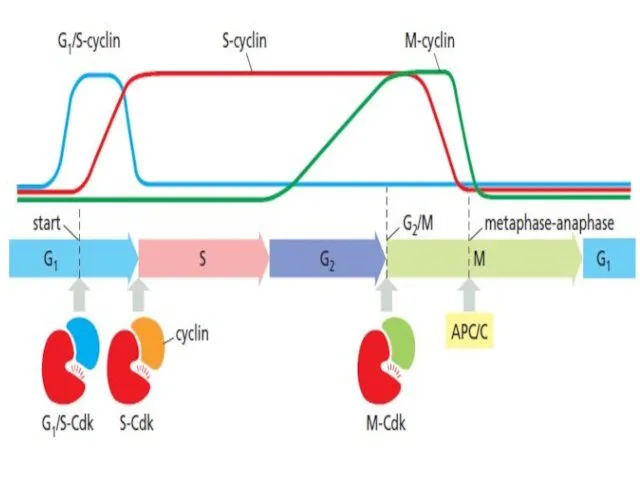
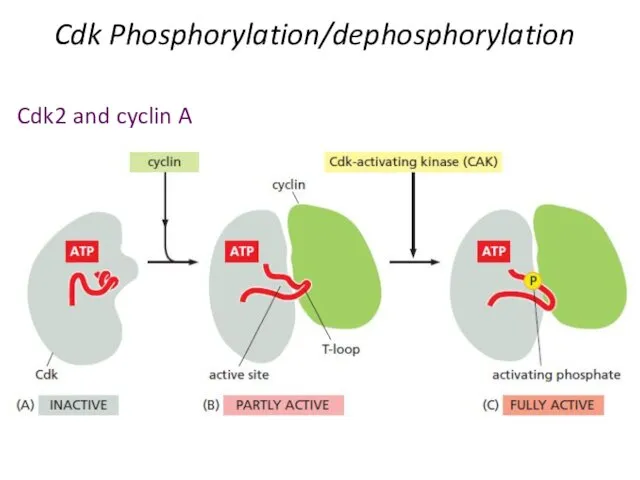
Cdk Phosphorylation/dephosphorylation
Cdk2 and cyclin A
Cdk Phosphorylation/dephosphorylation
Cdk2 and cyclin A
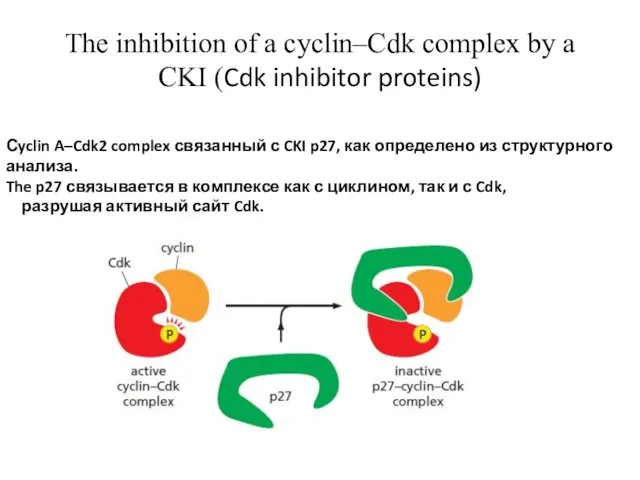
The inhibition of a cyclin–Cdk complex by a CKI (Cdk inhibitor
The inhibition of a cyclin–Cdk complex by a CKI (Cdk inhibitor

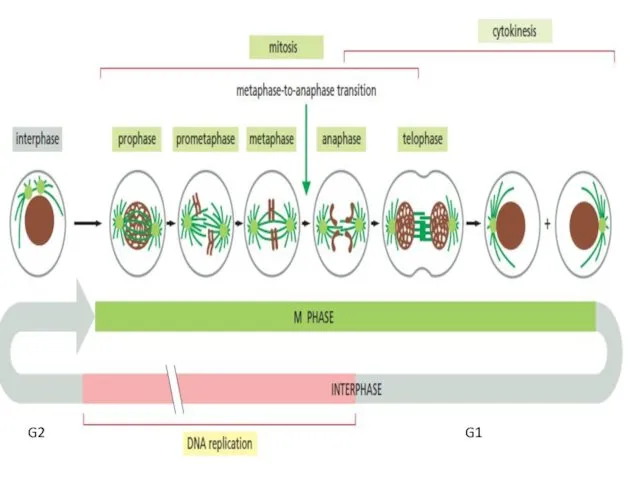
metaphase-to-anaphase transition
Anaphase promoting complex, or cyclosome (APC/C) = ubiquitin ligase
Продвижение через
metaphase-to-anaphase transition
Anaphase promoting complex, or cyclosome (APC/C) = ubiquitin ligase
Продвижение через

SCF убиквитинирует:
Определенные CKI в поздней G1, что позволяет активировать S-Cdk’s и
SCF убиквитинирует:
Определенные CKI в поздней G1, что позволяет активировать S-Cdk’s и
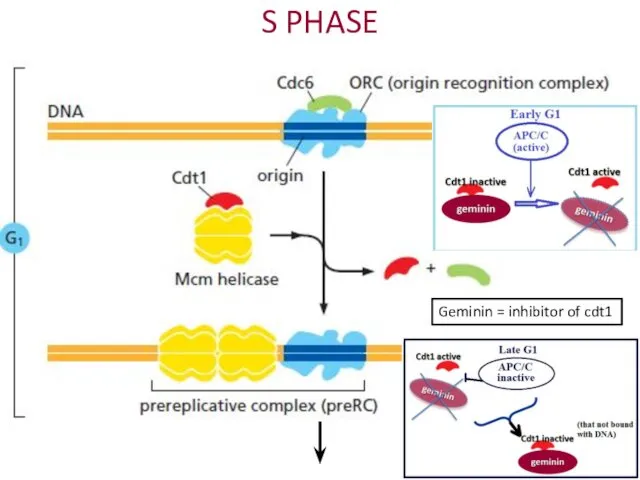
S PHASE
Geminin = inhibitor of cdt1
S PHASE
Geminin = inhibitor of cdt1
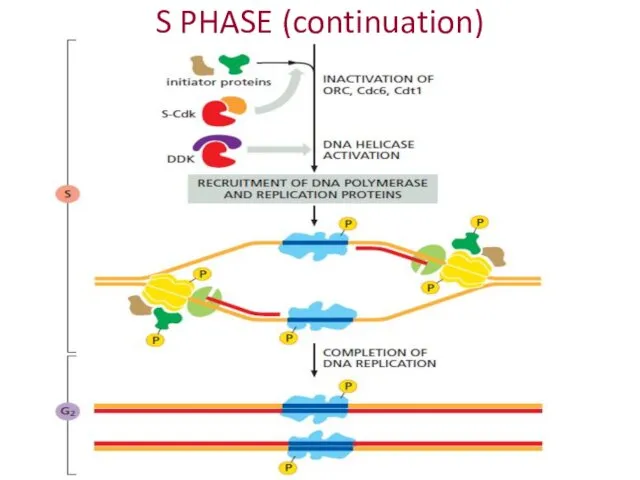
S PHASE (continuation)
S PHASE (continuation)
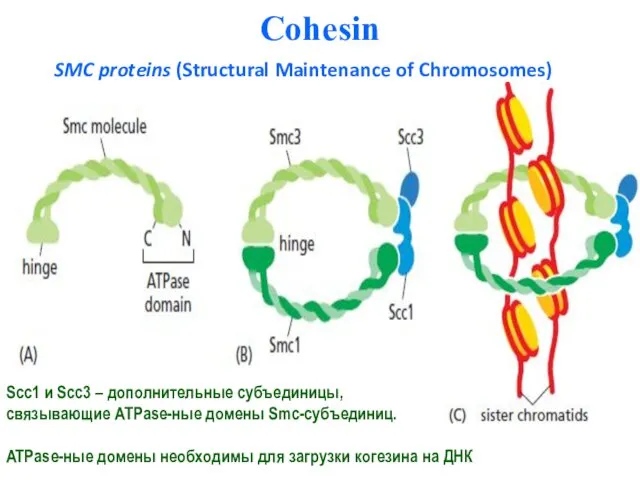
Cohesin
SMC proteins (Structural Maintenance of Chromosomes)
Scc1 и Scc3 – дополнительные субъединицы,
Cohesin
SMC proteins (Structural Maintenance of Chromosomes)
Scc1 и Scc3 – дополнительные субъединицы,
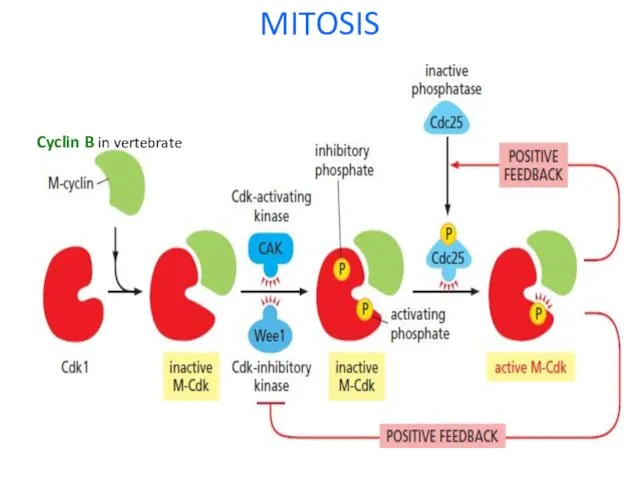
MITOSIS
Cyclin B in vertebrate
MITOSIS
Cyclin B in vertebrate
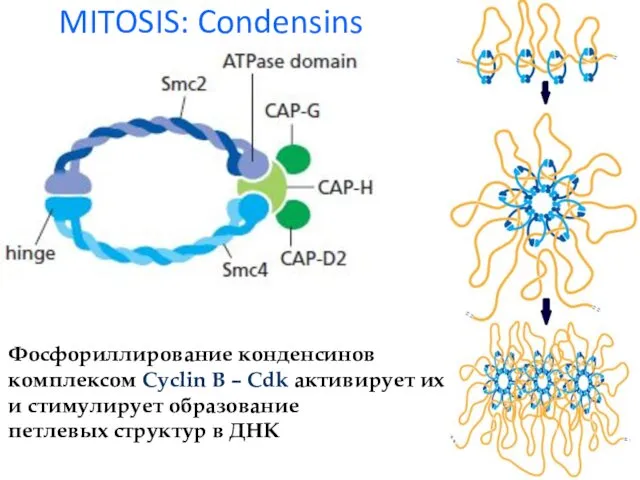
MITOSIS: Condensins
Фосфориллирование конденсинов
комплексом Cyclin B – Cdk активирует их
и
MITOSIS: Condensins
Фосфориллирование конденсинов
комплексом Cyclin B – Cdk активирует их
и
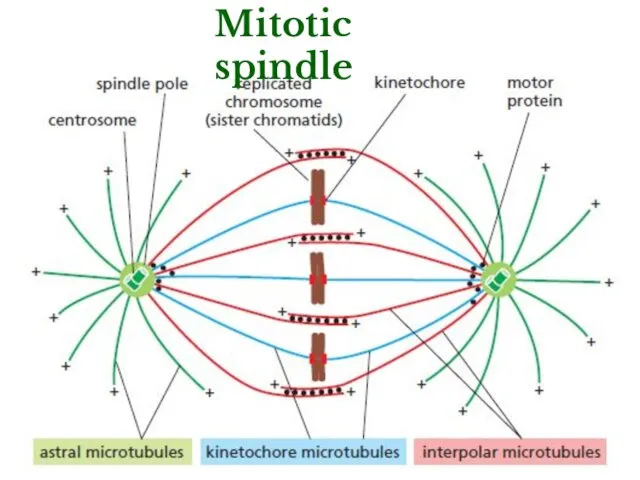
Mitotic spindle
Mitotic spindle
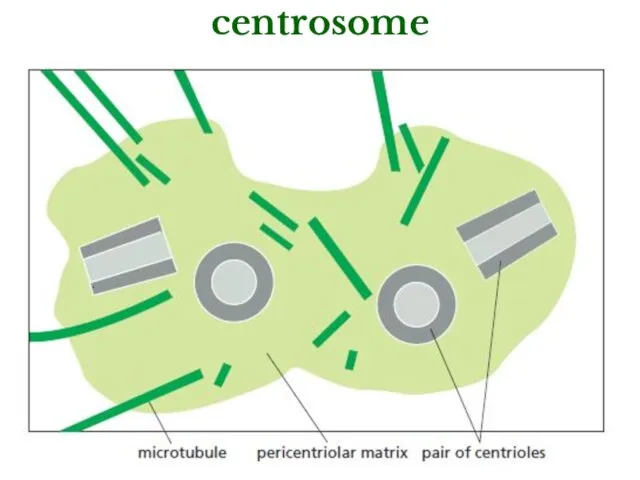
centrosome
centrosome
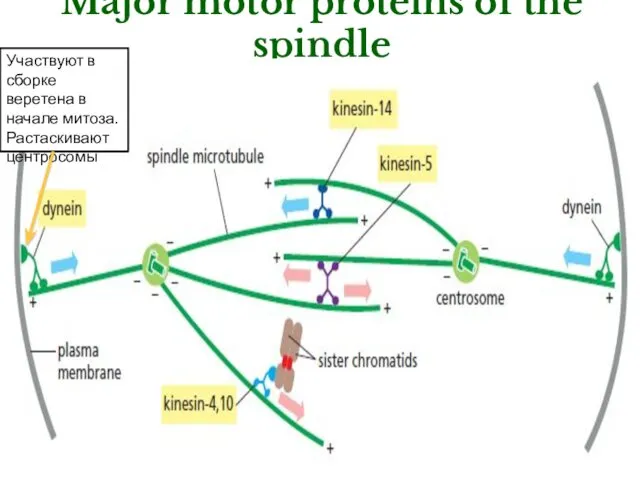
Major motor proteins of the spindle
Major motor proteins of the spindle
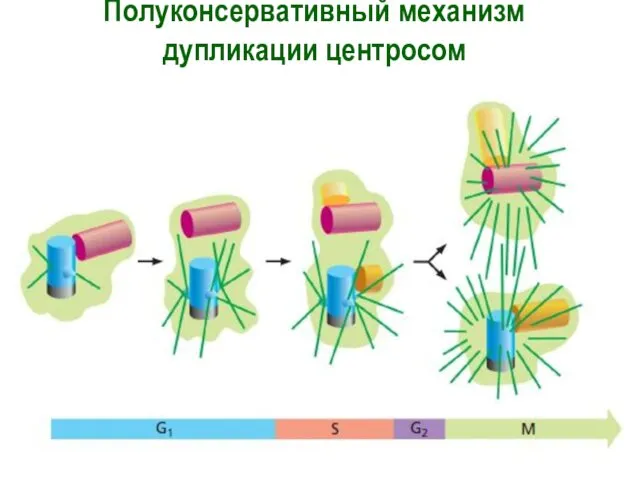
Полуконсервативный механизм дупликации центросом
Полуконсервативный механизм дупликации центросом
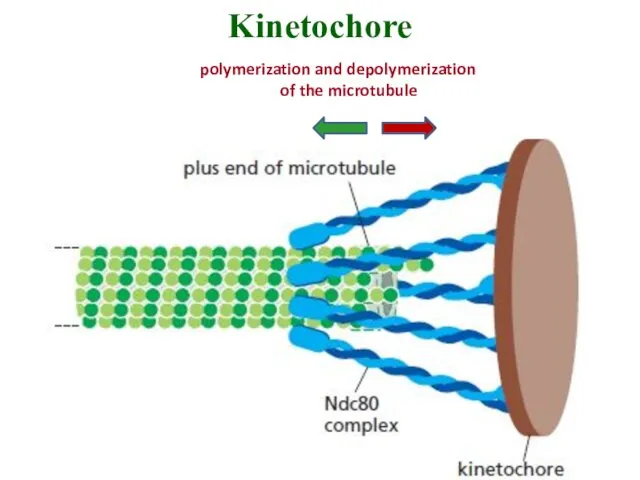
Kinetochore
polymerization and depolymerization
of the microtubule
Kinetochore
polymerization and depolymerization
of the microtubule
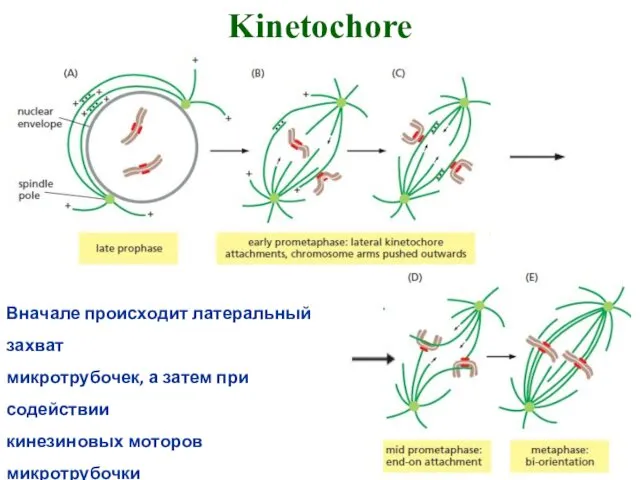
Kinetochore
Вначале происходит латеральный захват
микротрубочек, а затем при содействии
кинезиновых моторов микротрубочки
ориентируются
Kinetochore
Вначале происходит латеральный захват микротрубочек, а затем при содействии кинезиновых моторов микротрубочки ориентируются
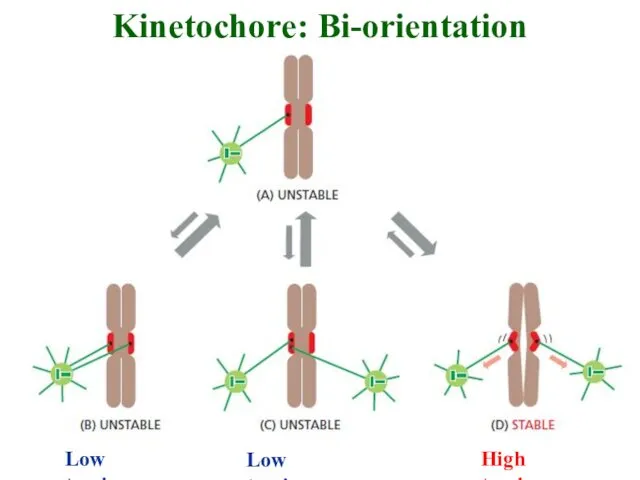
Kinetochore: Bi-orientation
Low tension
Low tension
High tension
Kinetochore: Bi-orientation
Low tension
Low tension
High tension
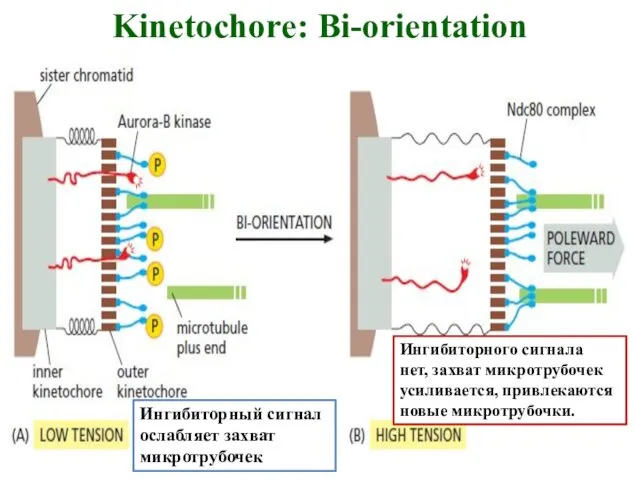
Kinetochore: Bi-orientation
Ингибиторный сигнал
ослабляет захват
микротрубочек
Ингибиторного сигнала
нет, захват микротрубочек
усиливается, привлекаются
Kinetochore: Bi-orientation
Ингибиторный сигнал
ослабляет захват
микротрубочек
Ингибиторного сигнала нет, захват микротрубочек усиливается, привлекаются

Сила, направляющая кинетохор к полюсу (разборка микротрубочек в месте прикрепления к
Сила, направляющая кинетохор к полюсу (разборка микротрубочек в месте прикрепления к
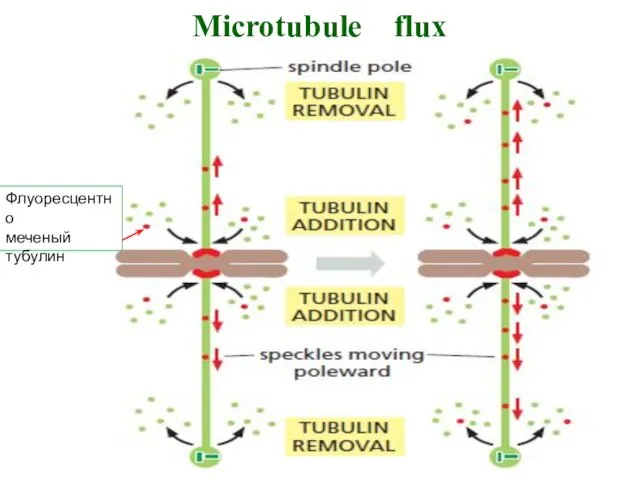
Флуоресцентно
меченый
тубулин
Microtubule flux
Флуоресцентно
меченый
тубулин
Microtubule flux
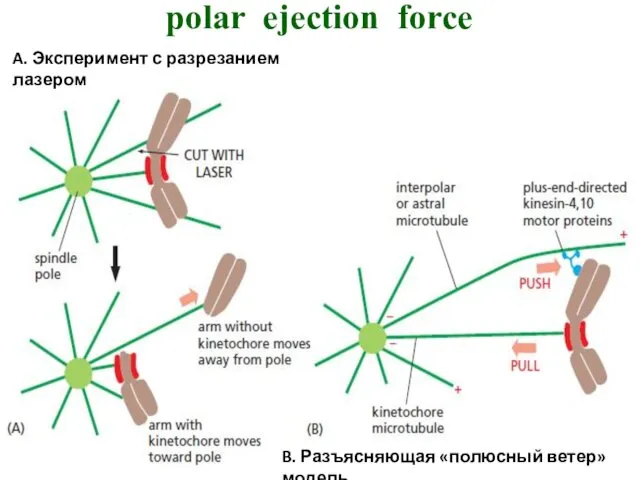
polar ejection force
A. Эксперимент с разрезанием лазером
B. Разъясняющая «полюсный ветер» модель
polar ejection force
A. Эксперимент с разрезанием лазером
B. Разъясняющая «полюсный ветер» модель
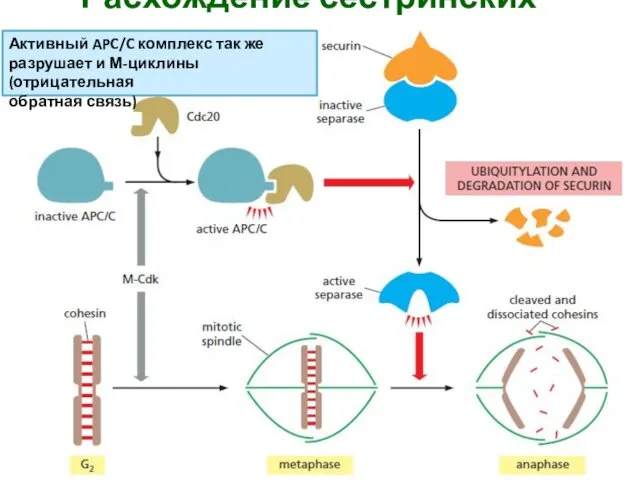
Расхождение сестринских хроматид
Активный APC/C комплекс так же разрушает и М-циклины (отрицательная
обратная
Расхождение сестринских хроматид
Активный APC/C комплекс так же разрушает и М-циклины (отрицательная обратная

Mad2 белок является проверочным механизмом присоединения к кинетохору трубочек. Если напряжения
Mad2 белок является проверочным механизмом присоединения к кинетохору трубочек. Если напряжения

Разборка веретена;
Фрагменты ядерной мембраны ассоциируются с отдельными хромосомами и затем
Разборка веретена;
Фрагменты ядерной мембраны ассоциируются с отдельными хромосомами и затем

CYTOKINESIS
Обычно митоз заканчивается цитокинезом.
Некоторые клетки (ранние эмбрионы дрозофилы, гепатоциты и кардиомиоциты)
CYTOKINESIS
Обычно митоз заканчивается цитокинезом.
Некоторые клетки (ранние эмбрионы дрозофилы, гепатоциты и кардиомиоциты)
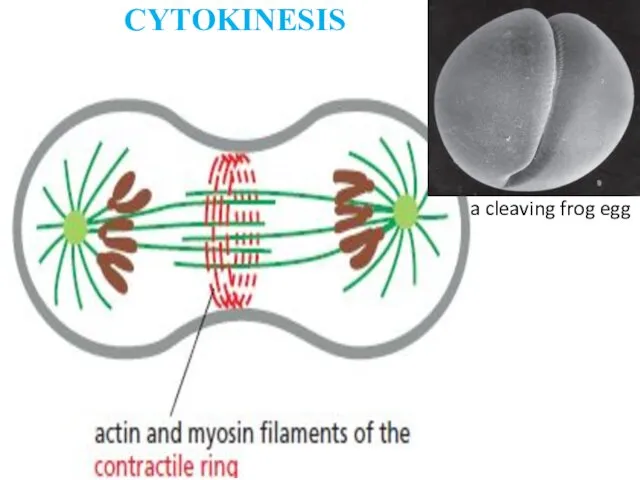
CYTOKINESIS
a cleaving frog egg
CYTOKINESIS
a cleaving frog egg
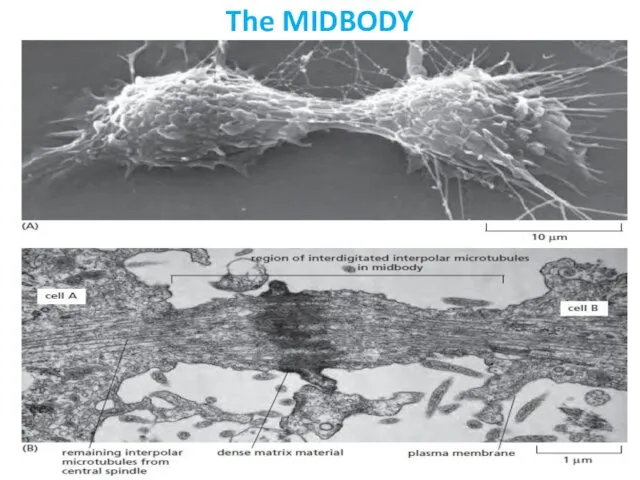
The MIDBODY
The MIDBODY
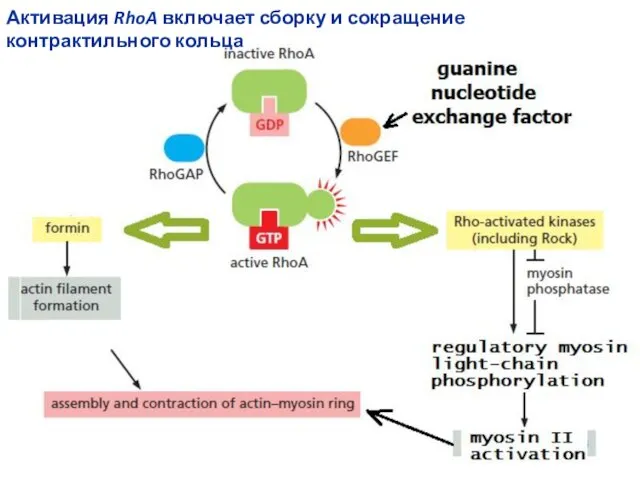
Активация RhoA включает сборку и сокращение
контрактильного кольца
Активация RhoA включает сборку и сокращение
контрактильного кольца
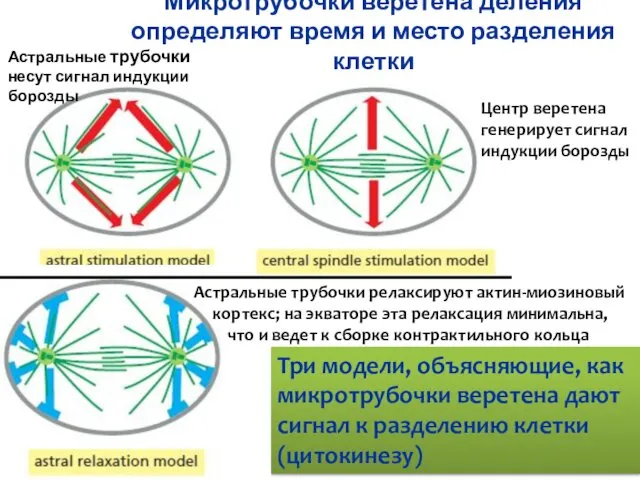
Микротрубочки веретена деления определяют время и место разделения клетки
Три модели, объясняющие,
Микротрубочки веретена деления определяют время и место разделения клетки
Три модели, объясняющие,
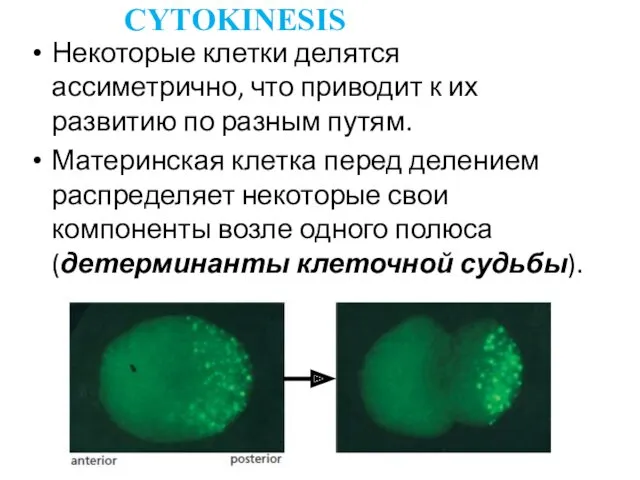
Некоторые клетки делятся ассиметрично, что приводит к их развитию по разным
Некоторые клетки делятся ассиметрично, что приводит к их развитию по разным
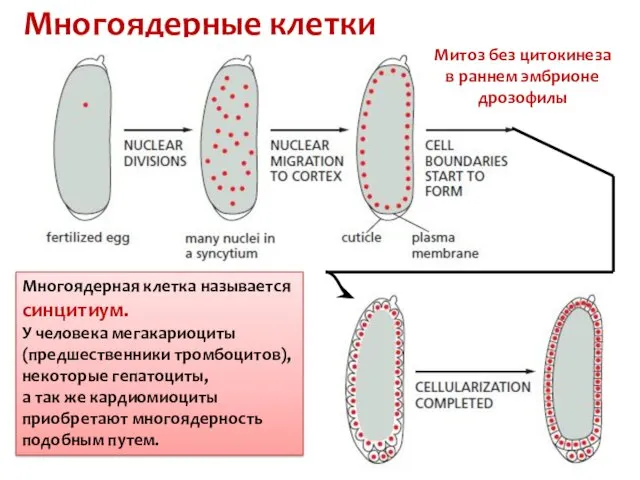
Многоядерные клетки
Многоядерная клетка называется
синцитиум.
У человека мегакариоциты
(предшественники тромбоцитов),
некоторые гепатоциты,
а
Многоядерные клетки
Многоядерная клетка называется
синцитиум.
У человека мегакариоциты
(предшественники тромбоцитов),
некоторые гепатоциты,
а
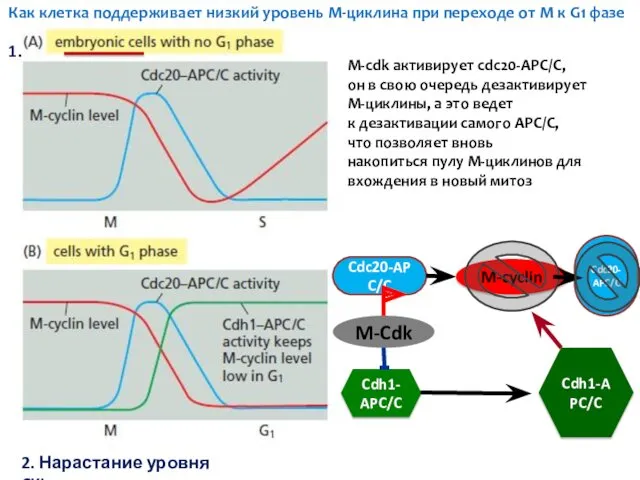
М-cdk активирует cdc20-APC/C,
он в свою очередь дезактивирует
М-циклины, а это ведет
М-cdk активирует cdc20-APC/C, он в свою очередь дезактивирует М-циклины, а это ведет

MEIOSIS
MEIOSIS
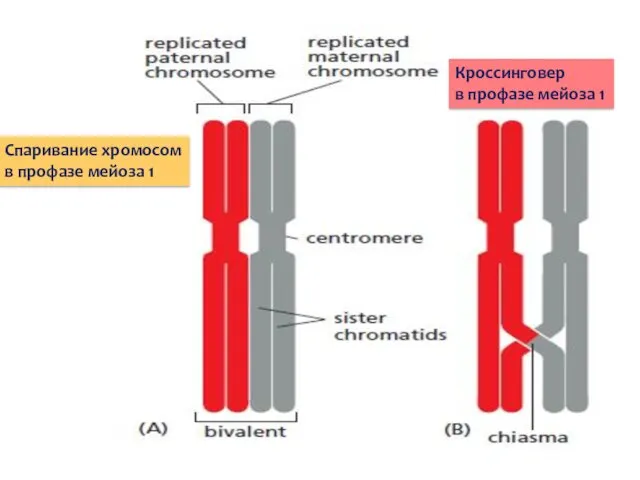
Спаривание хромосом
в профазе мейоза 1
Кроссинговер
в профазе мейоза 1
Спаривание хромосом
в профазе мейоза 1
Кроссинговер
в профазе мейоза 1
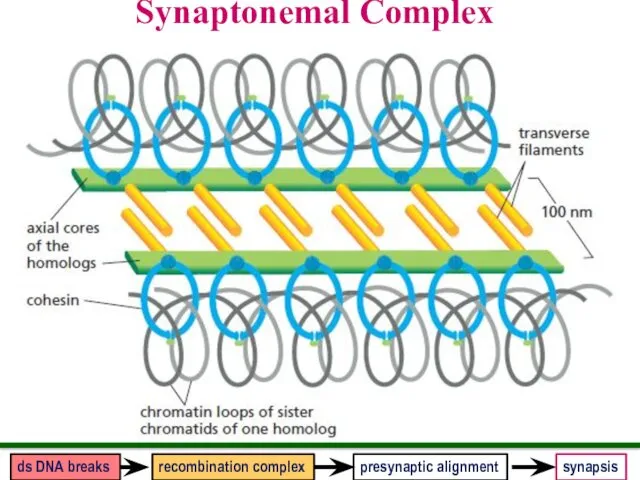
Synaptonemal Complex
presynaptic alignment
synapsis
ds DNA breaks
recombination complex
Synaptonemal Complex
presynaptic alignment
synapsis
ds DNA breaks
recombination complex
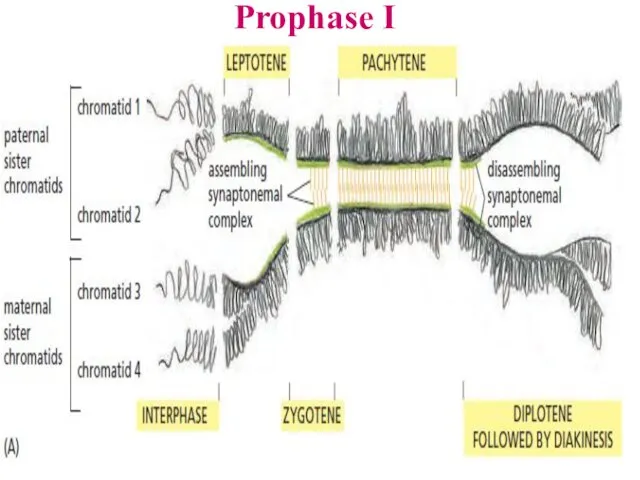
Prophase I
Prophase I
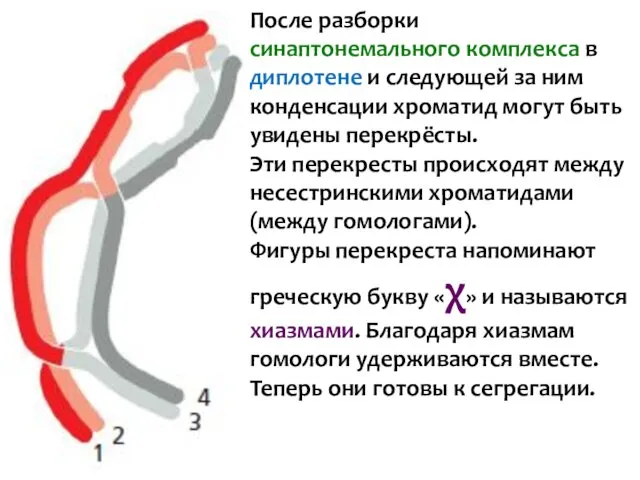
После разборки синаптонемального комплекса в диплотене и следующей за ним конденсации
После разборки синаптонемального комплекса в диплотене и следующей за ним конденсации

1. Оба сестринских кинетохора в гомологе должны быть соединены с ОДНИМ
1. Оба сестринских кинетохора в гомологе должны быть соединены с ОДНИМ
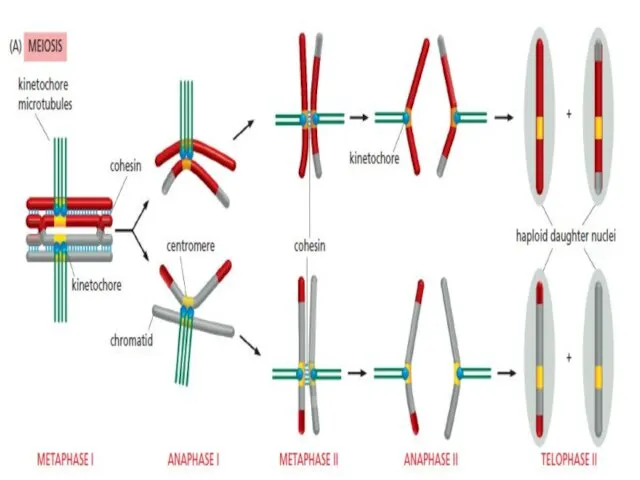

Кроссинговер тщательно регулируется: на каждую гомологичную пару хромосом приходится не менее
Кроссинговер тщательно регулируется: на каждую гомологичную пару хромосом приходится не менее

Контроль клеточного деления и роста
Внеклеточные сигналы, регулирующие клеточный рост,
деление и
Контроль клеточного деления и роста
Внеклеточные сигналы, регулирующие клеточный рост, деление и
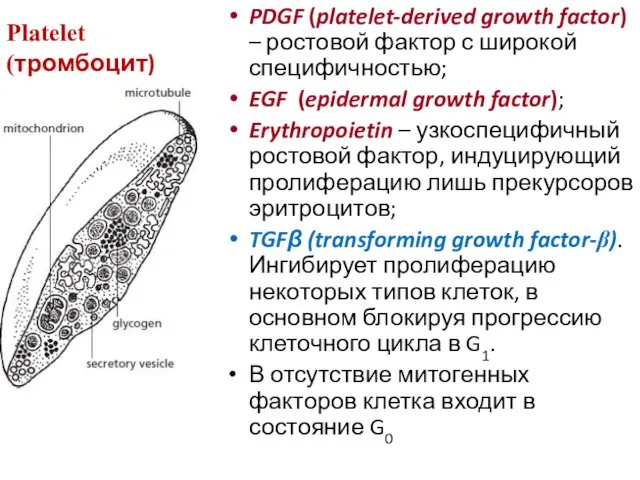
PDGF (platelet-derived growth factor) – ростовой фактор с широкой специфичностью;
EGF (epidermal
PDGF (platelet-derived growth factor) – ростовой фактор с широкой специфичностью;
EGF (epidermal
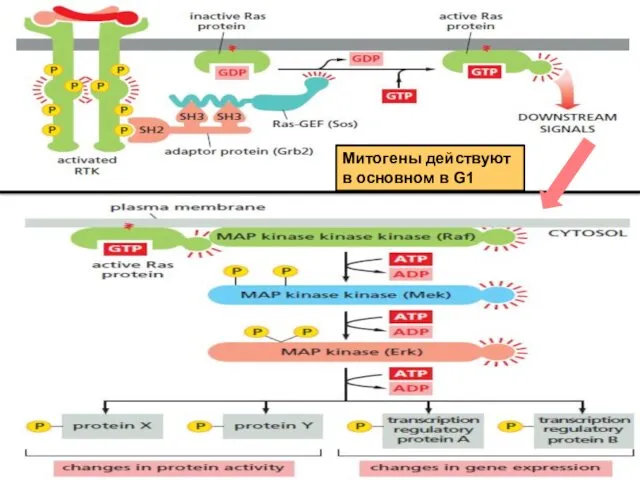
Митогены действуют
в основном в G1
Митогены действуют
в основном в G1
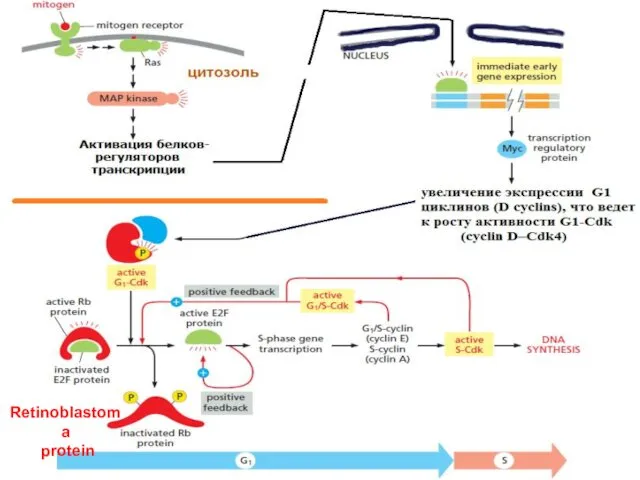
Retinoblastoma
protein
Retinoblastoma
protein
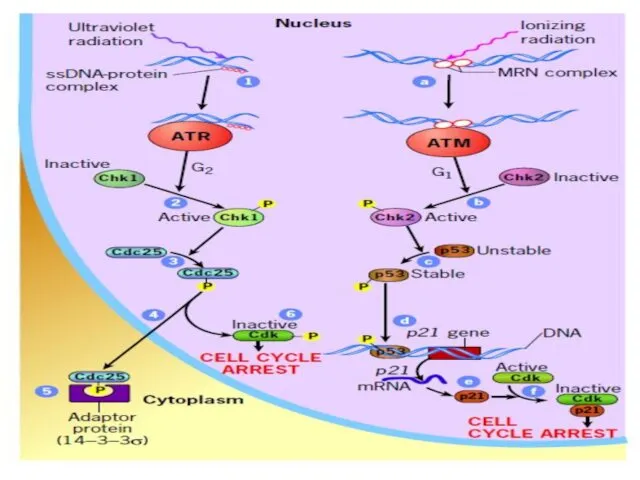
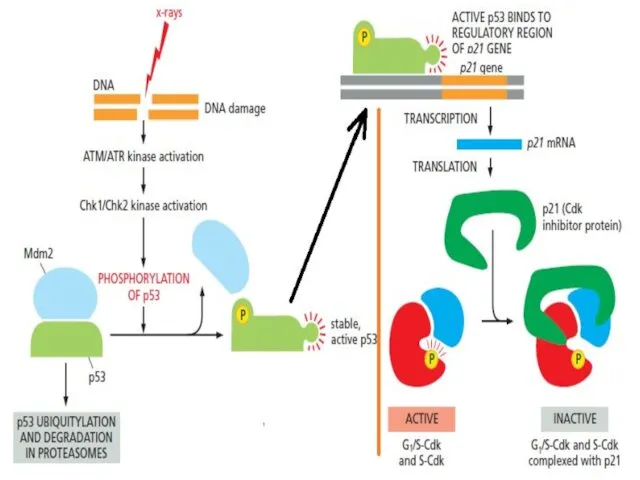

Клетки имеют ограниченное число делений (предел Хейфлика).
После этого предела клетки входят
Клетки имеют ограниченное число делений (предел Хейфлика).
После этого предела клетки входят
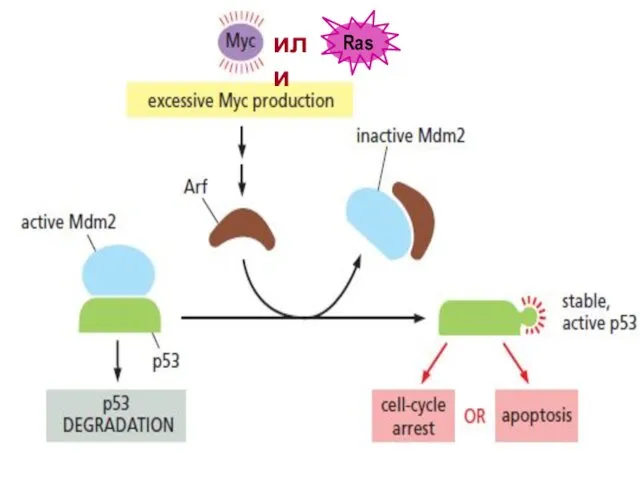
или
Ras
или
Ras
 Презентация Коллективная разработка ПО
Презентация Коллективная разработка ПО Невынашивание беременности
Невынашивание беременности Мікропроцесорна техніка (лекція 7)
Мікропроцесорна техніка (лекція 7) 7 класс: Оледенение С.Америки
7 класс: Оледенение С.Америки Чем археолог отличается от криминалиста. Социальные ландшафты: вещи в динамике прошлого
Чем археолог отличается от криминалиста. Социальные ландшафты: вещи в динамике прошлого Functional styles of the english language: newspaper style
Functional styles of the english language: newspaper style Статус - отчет по ремонту Сормовский 3060
Статус - отчет по ремонту Сормовский 3060 К юбилею В.Г. Распутина, Героя Социалистического Труда, дважды лауреата Государственной премии СССР
К юбилею В.Г. Распутина, Героя Социалистического Труда, дважды лауреата Государственной премии СССР Планиметрия. Повторение. Часть 1
Планиметрия. Повторение. Часть 1 Мартеновский способ производства стали. Выплавка стали в электропечах
Мартеновский способ производства стали. Выплавка стали в электропечах How to present in English
How to present in English Интеллектуальные роботы состояние и перспективы
Интеллектуальные роботы состояние и перспективы Презентация С правом по жизни.
Презентация С правом по жизни. особенности изображения Земли на глобусе и на карте
особенности изображения Земли на глобусе и на карте Декоративные злаки
Декоративные злаки Мы вместе. Часть 2
Мы вместе. Часть 2 Рисуем пластилином
Рисуем пластилином Монтаж и эксплуатация ПВО. Противовыбросовое оборудование иностранных производителей
Монтаж и эксплуатация ПВО. Противовыбросовое оборудование иностранных производителей Философия Августина Блаженного
Философия Августина Блаженного Классный час 1сентября Эхо Бесланской печали
Классный час 1сентября Эхо Бесланской печали Конспект урока географии в 6 классе Тайны подземных бурь.
Конспект урока географии в 6 классе Тайны подземных бурь. Закрепление изученного в 4 классе
Закрепление изученного в 4 классе Стандартизация свойств. Физические, механические, физико-химические свойства СМ. Долговечность и надежность
Стандартизация свойств. Физические, механические, физико-химические свойства СМ. Долговечность и надежность презентация
презентация Биосфера Земли. Тест
Биосфера Земли. Тест Презентация Автоматизация звука Л в словах
Презентация Автоматизация звука Л в словах Священная Библейская история Ветхого Завета. Грехопадение и его последствия (Быт.3)
Священная Библейская история Ветхого Завета. Грехопадение и его последствия (Быт.3) Нарушения обмена хромопротеидов
Нарушения обмена хромопротеидов