- Матричная РНК. (Лекция 7)

Содержание
- 2. Размеры синтезированной мРНК у бактерий практически совпадают с размерами гена – средний размер бактериальной мРНК –
- 3. Интересной особенностью бактерий является то, что некоторые гены у них сгруппированы в опероны. Оперон – это
- 4. По своим свойствам мРНК про- и эукариот существенно различаются. Бактериальные мРНК очень нестабильны – период их
- 5. Сопряжение транскрипции и трансляции у прокариот Направление транскрипции Рибосомы ДНК мРНК
- 6. Сопряжение транскрипции и трансляции у прокариот РНК полимераза Рибосома мРНК
- 7. У эукариотических организмов процессы транскрипции и трансляции разобщены. Транскрипция и созревание РНК идет в ядре, а
- 8. Структура мРНК прокариот Лидер - это 5’ не транслируемый участок - 5’ UTR (UnTranslated Region) Трейлер
- 9. Строение мРНК эукариот 5’-кэп- 7метил-гуанозин Лидер – 5’ нетранслируемый участок - 5’ UTR Кодирующая последовательность Трейлер
- 10. Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК). 2. Созревание 3‘-конца пре-мРНК. Присоединение
- 11. Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина, а также метилирование остатков рибозы двух первых нуклеотидов.
- 12. Функции кэпа: 1. Необходим для узнавания мРНК 40S-субчастицей рибосомы при инициации трансляции. 2. Для защиты 5'-конца
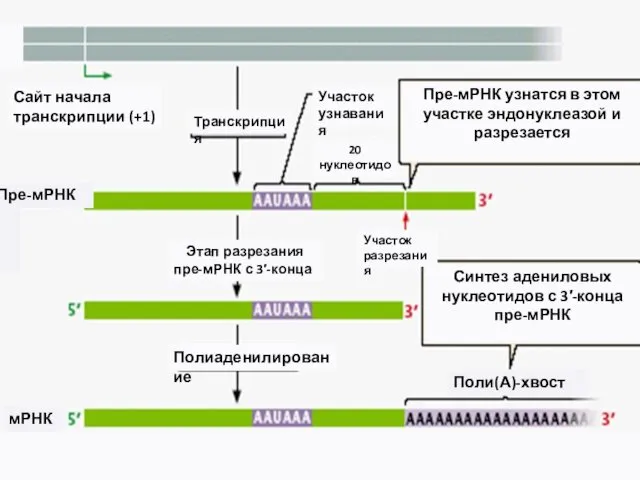
- 13. 2. Присоединение полиА-хвоста к 3`-концу пре-мРНК Молекула ДНК Пре-мРНК 5‘-КЭП Сигнал полиаденилирования Специальная эндонуклеаза узнает AAUAAA-участок
- 15. Функции полиА-хвоста 1. Способствуют экспорту зрелых мРНК из ядра. 2. Защищают мРНК от действия нуклеаз в
- 16. 3. Вырезание интронов из пре-мРНК В 1978 г. Филипп Шарп (Массачусетский технологический институт) открыл явление сплайсинга
- 17. Для мРНК высших организмов существуют обязательные правила сплайсинга: Правило 1. 5' и 3' концы интрона консервативны:
- 19. 5 типов мяРНК (U1,U2,U3,U4,U5) 150 белков
- 21. Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК). 2. Созревание 3‘-конца пре-мРНК. Присоединение
- 22. Схема созревания пре-мРНК у эукариотических организмов
- 23. Альтернативный сплайсинг
- 25. А
- 26. Альтернативный сплайсинг Альтернативный сплайсинг - форма сплайсинга, обеспечивающая кодирование одним геном структурно и функционально различающихся полипептидов.
- 27. После трансляции дает белок – гормон щитовидной железы кальцитоцин мРНК После трансляции дает нейропептид головного мозга
- 28. У человека есть ген slo. Он «работает» во внутреннем ухе, в частности, этот белок присутствует в
- 29. Нарушения альтернативного сплайсинга повинны в возникновении заболеваний человека, например, нейродегенеративных заболеваний, к которым относится болезнь Альцгеймера,
- 30. Редактирование мРНК Редактирование РНК (англ. RNA editing) — процесс, в ходе которого информация, содержащаяся в молекуле
- 31. Примеры редактирования хлоропластных и митохондриальных РНК растений А. Конструирование АУГ кодона Б. Замена аминокислоты В. Конструирование
- 32. Ген аполипопротеина ДНК Пре-мРНК кишечник печень без редактирования редактирование: ЦАА ? УАА Аполипопротеин В48 Аполипопротеин В100
- 33. Механизм редактирования с помощью гРНК гРНК (guide RNA, gRNA— небольшая РНК, которая функционирует в качестве матрицы
- 34. Биологические последствия редактирования мРНК Исправление ошибок в мРНК. Образование пригодной для трансляции мРНК из транскриптов, которые
- 35. Функциональное строение мРНК
- 36. ПРОКАРИОТЫ промотор ген А ген B ген C транскрипция старт стоп старт стоп старт стоп полицистронная
- 37. Область Шайн-Дальгарно
- 38. Правее точки «+1» (5'-конец мРНК) на небольшом расстоянии находится стартовый AUG-кодон, с которого рибосома начинает синтез
- 39. Точность посадки рибосомы контролируется самой мРНК – на участке между точкой «+1» и AUG-кодоном на молекуле
- 40. Далее между АUG-кодоном и стоп-кодоном находится транслируемая рибосомой область. Стоп-кодон (их известно три) – UGA, UAG
- 41. Строение мРНК эукариот 5’-кэп- 7метил-гуанозин. Лидер – 5’ нетранслируемый участок - 5’ UTR (UnTranslated Region) или
- 42. Основные типы РНК
- 43. Один из классов малых некодирующих РНК – микроРНК (miRNA) эндогенного происхождения, выполняющих в клетках функцию регуляции
- 44. Количество генов, кодирующих микроРНК, у различных организмов значительно варьирует. Например, у нематоды их 90-120, у человека
- 45. Исследование механизмов образования микроРНК в клетках животных показало, что этот процесс происходит в два этапа. Вначале
- 46. В настоящее время установлено, что микроРНК могут регулировать процесс трансляции. Регуляция осуществляется путем комплементарного связывания микроРНК
- 47. Один тип микроРНК может регулировать трансляцию мРНК более 100 различных генов. Степень подавления работы генов зависит
- 48. Функции микроРНК Регуляция синтеза РНК на уровне трансляции. Метилирование гетерохроматина. Участвует в дифференцировке тканей. Участвует в
- 49. Еще не изучив до конца функции микроРНК и способы их действия, ученые обнаружили другой интереснейший факт
- 50. Подобные работы были проведены сотрудниками Университета штата Огайо на опухолевых клетках поджелудочной железы. Было показано, что
- 51. Несколько иного плана работы были проведены в отношении другого онкологического заболевания – рака легких. Исследователи сосредоточили
- 52. Таким образом, сегодня становиться очевидным, что функции микроРНК более разнообразны, чем нам известно сегодня. Есть предположение,
- 53. Количество разновидностей микроРНК у высших организмов ещё до конца не установлено, по некоторым данным, оно превосходит
- 54. Транскриция При-микроРНК Обрезание шпильки Ядро Цитоплазма Дальнейшее созревание
- 55. 1 2 3 4 5 1 Предшественник микроРНК транскрибируется с обеих цепей геномной ДНК РНК-полимеразой II,
- 56. 2 Эндонуклеаза Drosha отрезает от при-микроРНК одноцепочечные «хвосты». После чего структура транспортируется в цитоплазму. 3 В
- 58. Функции микроРНК 1. МикроРНК - небелковый регулятор онтогенеза. На различных этапах жизни организмов включаются и выключаются
- 59. siРНК – малые интерферирующие РНК (даем латинское обозначение, русская аббревиатура пока не является общепринятой) – это
- 60. В нормальной клетке каждый ген выполняет свою собственную, строго определенную функцию, например, кодирует синтез мРНК, которая
- 61. Образуются из длинных двуцепочечных РНК, источником которых являются: РНК-содержащие вирусы, попавшие в клетки эукариот. РНК, считываемая
- 62. Функции siРНК Защита от вирусов. Репрессия трансгенов. Подавление экспрессии мобильных элементов. Подавление работы ряда генов в
- 63. Почему же именно siРНК вызвала столь бурный всплеск интереса к себе? Ведь в распоряжении биологов имеется
- 64. «Интерферирующие» короткие молекулы двухцепочечной РНК (дцРНК) попадают в клетку извне двумя путями: в готовом виде, либо
- 65. Комплексы Dicer и RISC могут использовать в качестве «затравки» не только дцРНК, но и одноцепочечную РНК,
- 66. Функции киРНК (siРНК) Это иммунная защита в клетках беспозвоночных. «Традиционная» иммунная система (лимфоциты + лейкоциты +
- 67. Практическое использование киРНК На основе киРНК разработана технология «выключения» (или нокдауна) отдельных генов in vitro (на
- 68. МикроРНК закодированны в уникальных генах геномов многоклеточных организмов (>1% от всех генов у человека); мРНК может
- 69. siРНК в настоящее время широко используется для «нокаутирования» генов (их выключения) в ходе анализа функций различных
- 70. Проводятся эксперименты на модельных животных по использованию siРНК в качестве инструмента выключения генов с целью лечения
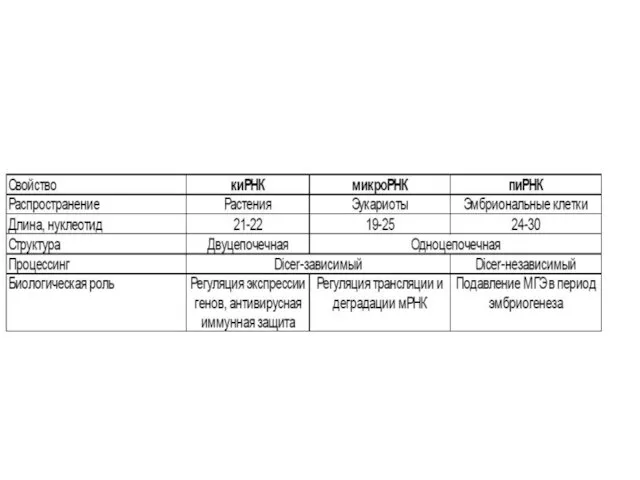
- 71. Следует отметить, что спектр исследований в мире в области РНК-интерференции расширяется с каждым днем. Например, делаются
- 72. Созданы библиотеки коротких siРНК и ДНК-векторов, кодирующих короткие РНК, мишенями которых является около 8000 генов генома
- 73. Малые ядерные РНК - мяРНК (snRNA) Обнаруживаются в ядре, всегда связаны с белками, формируя малые ядерные
- 74. мякРНК имеют длину до 70 нуклеотидов. мякРНК – это многочисленная группа не кодирующих белок РНК. В
- 75. тмРНК – это бактериальная РНК с двойной функцией – имеет участок тРНК и участок мРНК. тмРНК
- 76. tmРНК Рибосома мРНК мРНК tmРНК способна присоединять аминокислоту аланин или нескольких аланиновых остатоков к белковой молекуле
- 77. Pi-РНК - специфический агент против транспозиции пиРНК (pi RNA) — короткие молекулы длиной в 24–30 нуклеотидов,
- 78. Гены пиРНК активны только в зародышевых клетках (во время эмбриогенеза) и окружающих их эндотелиальных клетках. Важно!
- 79. Фотография половой клетки после мейоза, в которой отсутствуют piРНК. Зелёное светящееся вещество указывает на серьёзные повреждения
- 80. На сегодняшний день идентифицированы уже тысячи коротких регуляторных РНК, а механизм РНК-интерференции изучен очень подробно, однако
- 81. «Портрет» пиРНК пиРНК — короткие молекулы длиной в 24–30 нуклеотидов, закодированные в центромерных и теломерных областях
- 84. Скачать презентацию

Размеры синтезированной мРНК у бактерий практически совпадают с размерами гена –
Размеры синтезированной мРНК у бактерий практически совпадают с размерами гена –

Интересной особенностью бактерий является то, что некоторые гены у них сгруппированы
Интересной особенностью бактерий является то, что некоторые гены у них сгруппированы

По своим свойствам мРНК про- и эукариот существенно различаются.
Бактериальные мРНК
По своим свойствам мРНК про- и эукариот существенно различаются.
Бактериальные мРНК

Сопряжение транскрипции и трансляции у прокариот
Направление транскрипции
Рибосомы
ДНК
мРНК
Сопряжение транскрипции и трансляции у прокариот
Направление транскрипции
Рибосомы
ДНК
мРНК
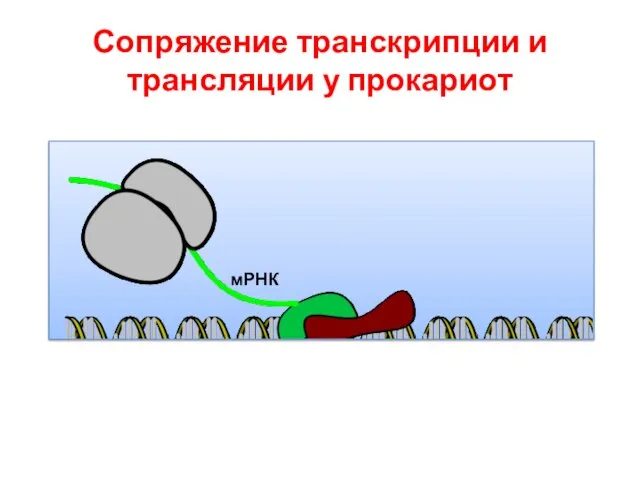
Сопряжение транскрипции и трансляции у прокариот
РНК полимераза
Рибосома
мРНК
Сопряжение транскрипции и трансляции у прокариот
РНК полимераза
Рибосома
мРНК
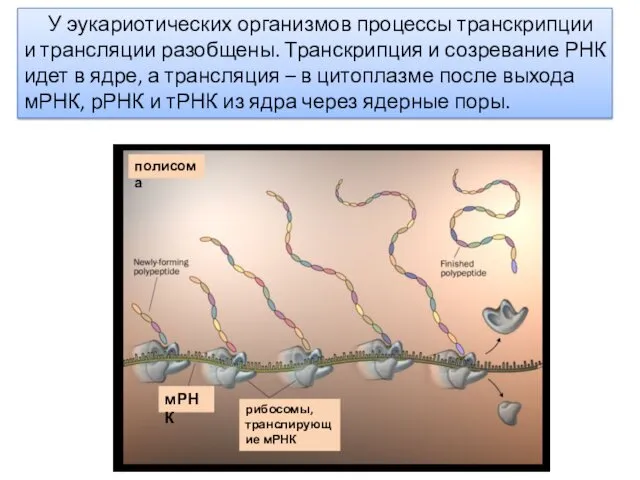
У эукариотических организмов процессы транскрипции и трансляции разобщены. Транскрипция и созревание
У эукариотических организмов процессы транскрипции и трансляции разобщены. Транскрипция и созревание
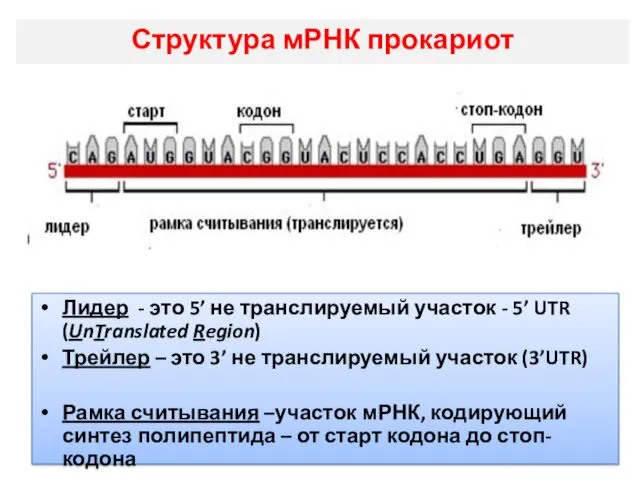
Структура мРНК прокариот
Лидер - это 5’ не транслируемый участок - 5’
Структура мРНК прокариот
Лидер - это 5’ не транслируемый участок - 5’
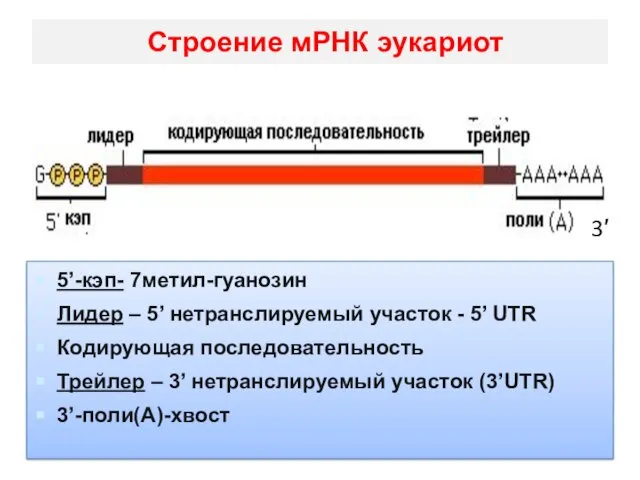
Строение мРНК эукариот
5’-кэп- 7метил-гуанозин
Лидер – 5’ нетранслируемый участок - 5’ UTR
Строение мРНК эукариот
5’-кэп- 7метил-гуанозин
Лидер – 5’ нетранслируемый участок - 5’ UTR

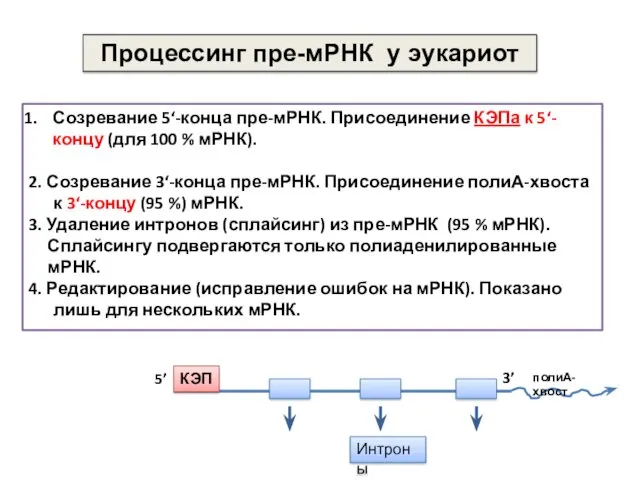
Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК).
2.
Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК).
2.
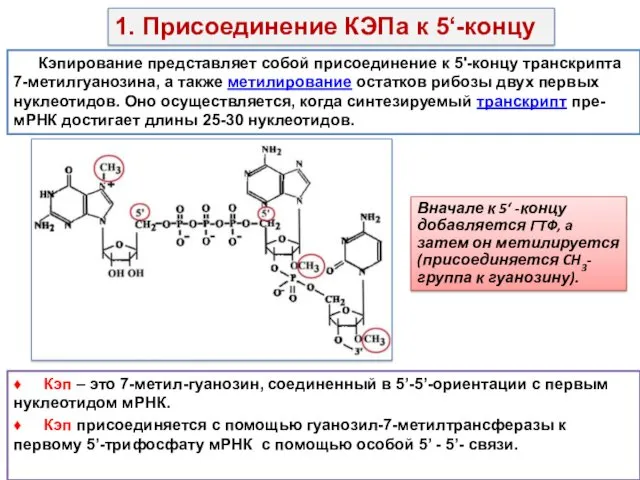
Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина, а также метилирование
Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина, а также метилирование

Функции кэпа:
1. Необходим для узнавания мРНК 40S-субчастицей рибосомы при инициации трансляции.
Функции кэпа:
1. Необходим для узнавания мРНК 40S-субчастицей рибосомы при инициации трансляции.
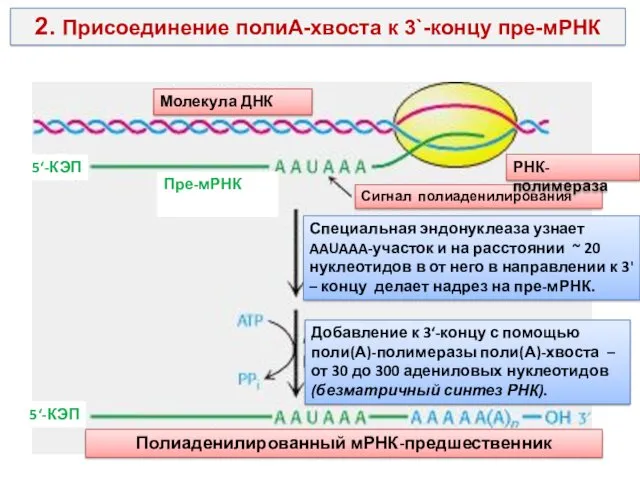
2. Присоединение полиА-хвоста к 3`-концу пре-мРНК
Молекула ДНК
Пре-мРНК
5‘-КЭП
Сигнал полиаденилирования
Специальная эндонуклеаза узнает AAUAAA-участок
2. Присоединение полиА-хвоста к 3`-концу пре-мРНК
Молекула ДНК
Пре-мРНК
5‘-КЭП
Сигнал полиаденилирования
Специальная эндонуклеаза узнает AAUAAA-участок

Функции полиА-хвоста
1. Способствуют экспорту зрелых мРНК из ядра.
2. Защищают мРНК
Функции полиА-хвоста
1. Способствуют экспорту зрелых мРНК из ядра.
2. Защищают мРНК
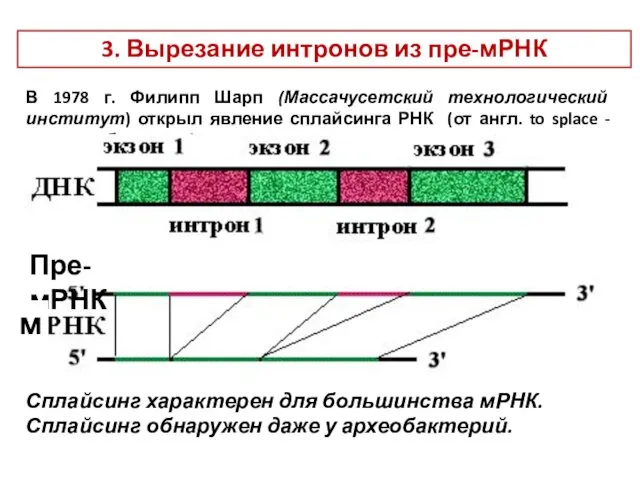
3. Вырезание интронов из пре-мРНК
В 1978 г. Филипп Шарп (Массачусетский технологический
3. Вырезание интронов из пре-мРНК
В 1978 г. Филипп Шарп (Массачусетский технологический
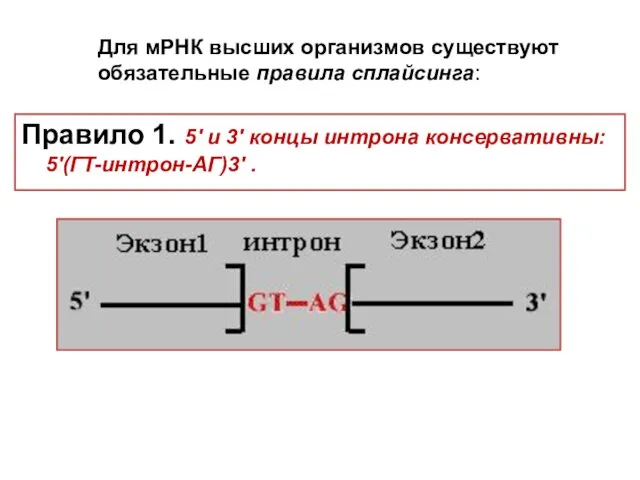
Для мРНК высших организмов существуют обязательные правила сплайсинга:
Правило 1. 5'
Для мРНК высших организмов существуют обязательные правила сплайсинга:
Правило 1. 5'
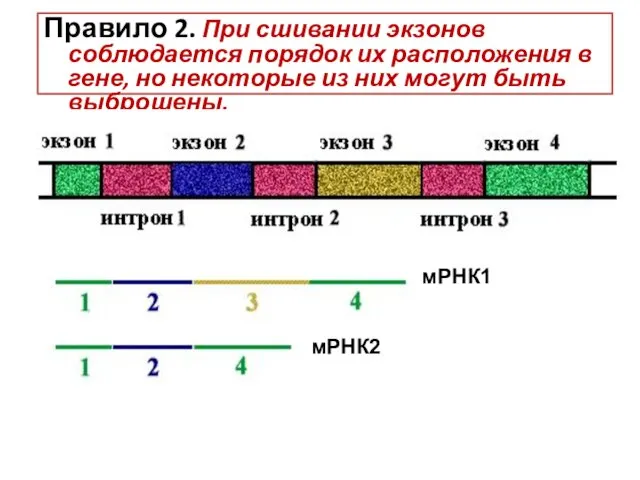
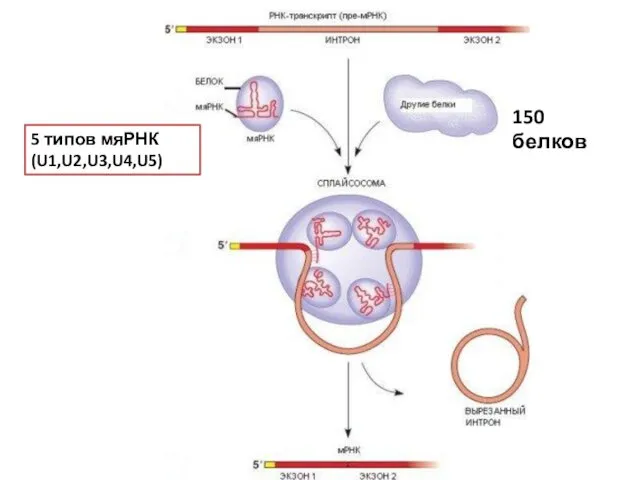
5 типов мяРНК (U1,U2,U3,U4,U5)
150 белков
5 типов мяРНК (U1,U2,U3,U4,U5)
150 белков
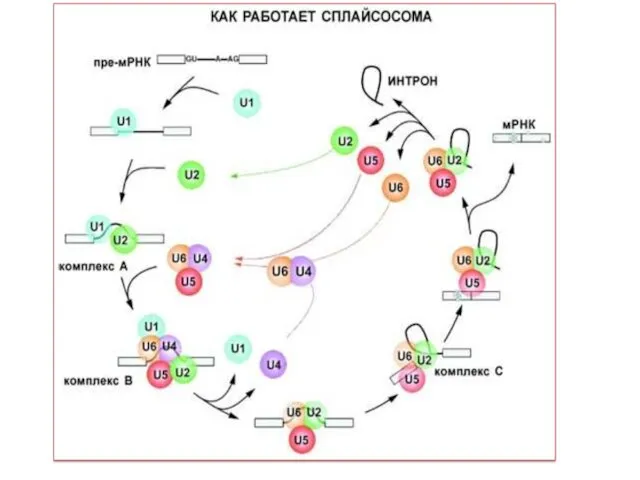
Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК).
2.
Созревание 5‘-конца пре-мРНК. Присоединение КЭПа к 5‘-концу (для 100 % мРНК).
2.
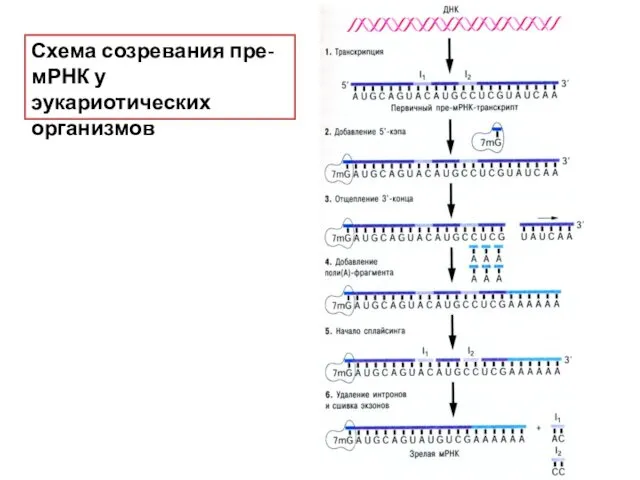
Схема созревания пре-мРНК у эукариотических организмов
Схема созревания пре-мРНК у эукариотических организмов

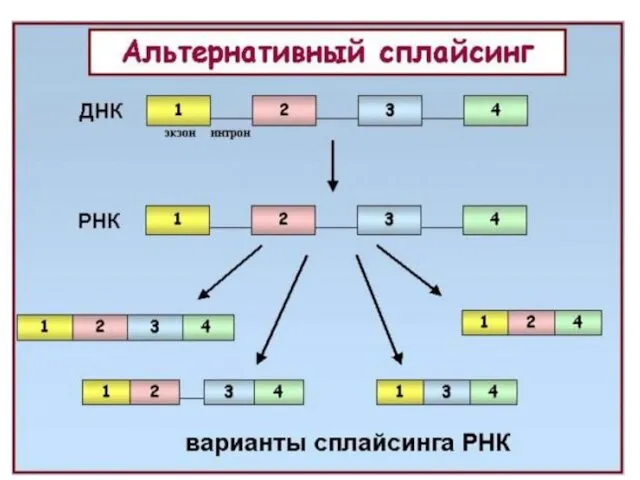
Альтернативный сплайсинг
Альтернативный сплайсинг
А
А

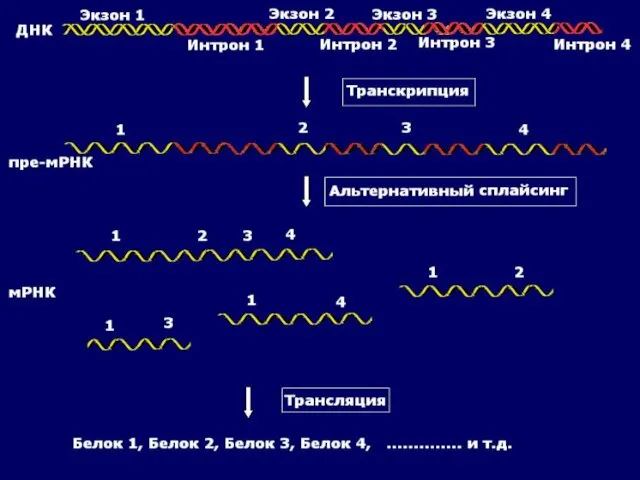
Альтернативный сплайсинг
Альтернативный сплайсинг - форма сплайсинга, обеспечивающая кодирование одним геном структурно
Альтернативный сплайсинг
Альтернативный сплайсинг - форма сплайсинга, обеспечивающая кодирование одним геном структурно
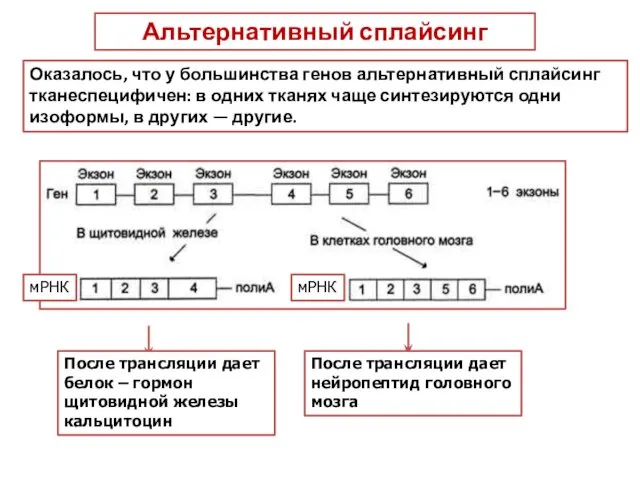
После трансляции дает белок – гормон щитовидной железы кальцитоцин
мРНК
После трансляции дает
После трансляции дает белок – гормон щитовидной железы кальцитоцин
мРНК
После трансляции дает

У человека есть ген slo. Он «работает» во внутреннем ухе, в частности,
У человека есть ген slo. Он «работает» во внутреннем ухе, в частности,

Нарушения альтернативного сплайсинга повинны в возникновении заболеваний человека, например, нейродегенеративных заболеваний,
Нарушения альтернативного сплайсинга повинны в возникновении заболеваний человека, например, нейродегенеративных заболеваний,

Редактирование мРНК
Редактирование РНК (англ. RNA editing) — процесс, в ходе которого информация, содержащаяся
Редактирование мРНК
Редактирование РНК (англ. RNA editing) — процесс, в ходе которого информация, содержащаяся
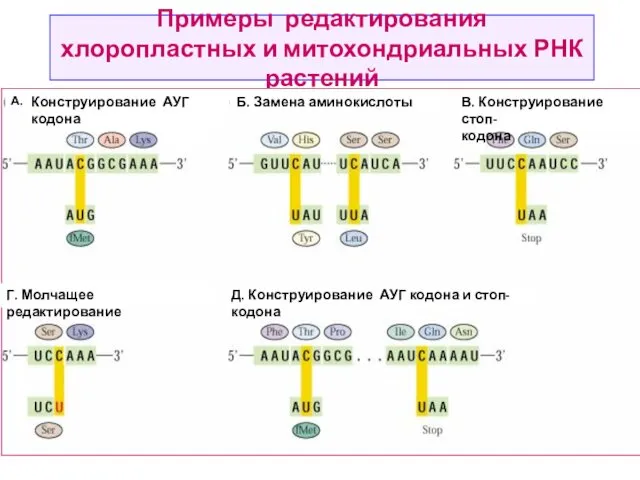
Примеры редактирования хлоропластных и митохондриальных РНК растений
А. Конструирование АУГ кодона
Б. Замена
А. Конструирование АУГ кодона
Б. Замена
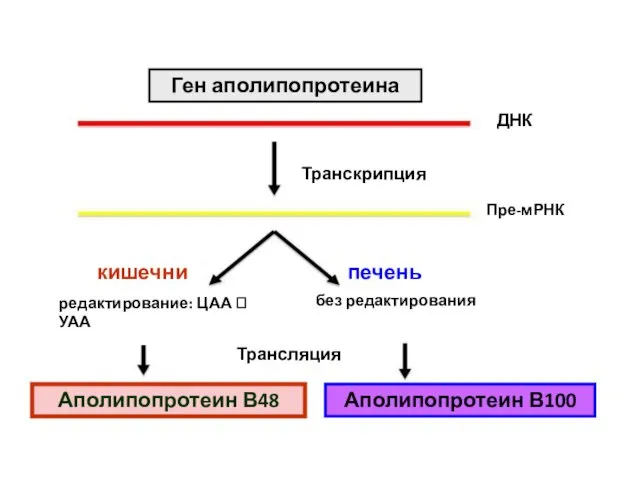
Ген аполипопротеина
ДНК
Пре-мРНК
кишечник
печень
без редактирования
редактирование: ЦАА ? УАА
Аполипопротеин В48
Аполипопротеин В100
Транскрипция
Трансляция
Ген аполипопротеина
ДНК
Пре-мРНК
кишечник
печень
без редактирования
редактирование: ЦАА ? УАА
Аполипопротеин В48
Аполипопротеин В100
Транскрипция
Трансляция
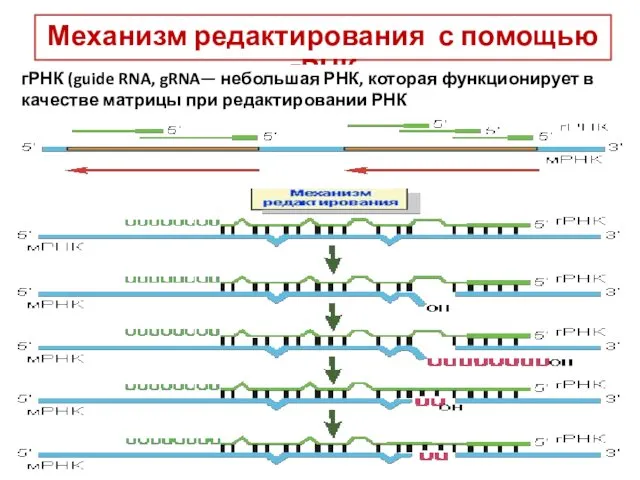
Механизм редактирования с помощью гРНК
гРНК (guide RNA, gRNA— небольшая РНК, которая
Механизм редактирования с помощью гРНК
гРНК (guide RNA, gRNA— небольшая РНК, которая

Биологические последствия редактирования мРНК
Исправление ошибок в мРНК.
Образование пригодной для трансляции
Биологические последствия редактирования мРНК
Исправление ошибок в мРНК.
Образование пригодной для трансляции

Функциональное строение мРНК
Функциональное строение мРНК
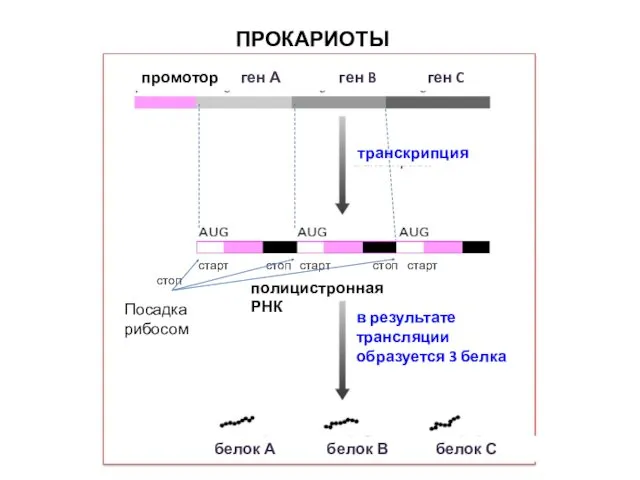
ПРОКАРИОТЫ
промотор ген А ген B ген C
транскрипция
старт стоп старт
ПРОКАРИОТЫ
промотор ген А ген B ген C
транскрипция
старт стоп старт
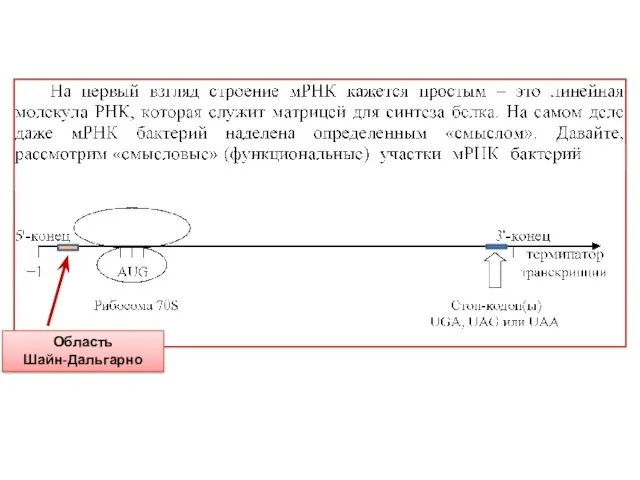
Область
Шайн-Дальгарно
Область
Шайн-Дальгарно

Правее точки «+1» (5'-конец мРНК) на небольшом расстоянии находится стартовый AUG-кодон,
Правее точки «+1» (5'-конец мРНК) на небольшом расстоянии находится стартовый AUG-кодон,

Точность посадки рибосомы контролируется самой мРНК – на участке между точкой
Точность посадки рибосомы контролируется самой мРНК – на участке между точкой

Далее между АUG-кодоном и стоп-кодоном находится транслируемая рибосомой область.
Стоп-кодон (их
Далее между АUG-кодоном и стоп-кодоном находится транслируемая рибосомой область.
Стоп-кодон (их
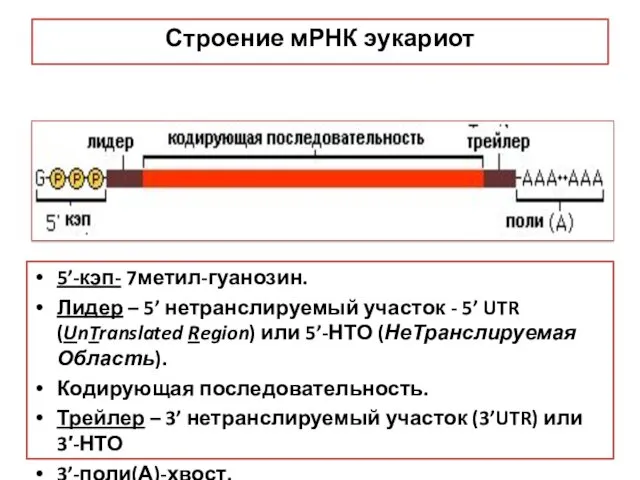
Строение мРНК эукариот
5’-кэп- 7метил-гуанозин.
Лидер – 5’ нетранслируемый участок - 5’ UTR
Строение мРНК эукариот
5’-кэп- 7метил-гуанозин.
Лидер – 5’ нетранслируемый участок - 5’ UTR
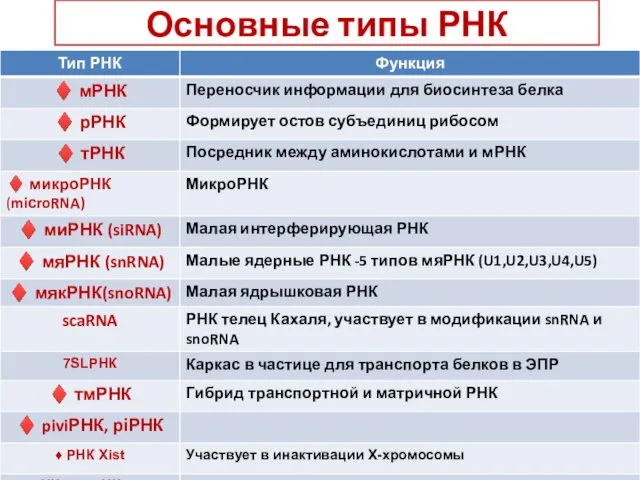
Основные типы РНК
Основные типы РНК

Один из классов малых некодирующих РНК – микроРНК (miRNA) эндогенного происхождения,
Один из классов малых некодирующих РНК – микроРНК (miRNA) эндогенного происхождения,

Количество генов, кодирующих микроРНК, у различных организмов значительно варьирует. Например, у
Количество генов, кодирующих микроРНК, у различных организмов значительно варьирует. Например, у

Исследование механизмов образования микроРНК в клетках животных показало, что этот процесс
Исследование механизмов образования микроРНК в клетках животных показало, что этот процесс

В настоящее время установлено, что микроРНК могут регулировать процесс трансляции. Регуляция
В настоящее время установлено, что микроРНК могут регулировать процесс трансляции. Регуляция

Один тип микроРНК может регулировать трансляцию мРНК более 100 различных
Один тип микроРНК может регулировать трансляцию мРНК более 100 различных

Функции микроРНК
Регуляция синтеза РНК на уровне трансляции.
Метилирование гетерохроматина.
Участвует в дифференцировке тканей.
Функции микроРНК
Регуляция синтеза РНК на уровне трансляции.
Метилирование гетерохроматина.
Участвует в дифференцировке тканей.

Еще не изучив до конца функции микроРНК и способы их действия,
Еще не изучив до конца функции микроРНК и способы их действия,

Подобные работы были проведены сотрудниками Университета штата Огайо на опухолевых клетках
Подобные работы были проведены сотрудниками Университета штата Огайо на опухолевых клетках

Несколько иного плана работы были проведены в отношении другого онкологического заболевания
Несколько иного плана работы были проведены в отношении другого онкологического заболевания

Таким образом, сегодня становиться очевидным, что функции микроРНК более разнообразны,
Таким образом, сегодня становиться очевидным, что функции микроРНК более разнообразны,

Количество разновидностей микроРНК у высших организмов ещё до конца не установлено, по некоторым данным, оно
Количество разновидностей микроРНК у высших организмов ещё до конца не установлено, по некоторым данным, оно
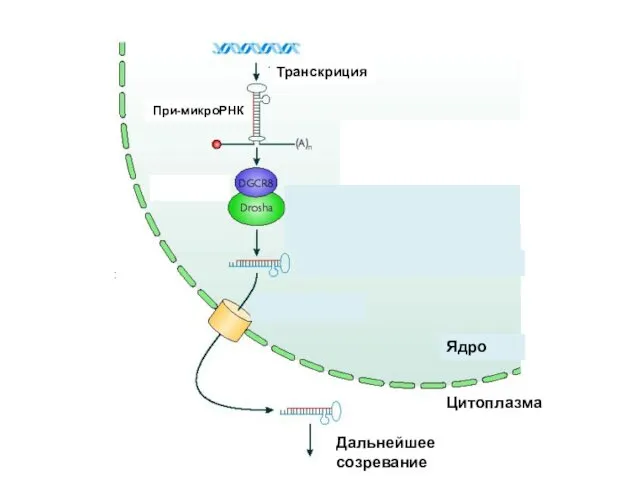
Транскриция
При-микроРНК
Обрезание шпильки
Ядро
Цитоплазма
Дальнейшее созревание
Транскриция
При-микроРНК
Обрезание шпильки
Ядро
Цитоплазма
Дальнейшее созревание
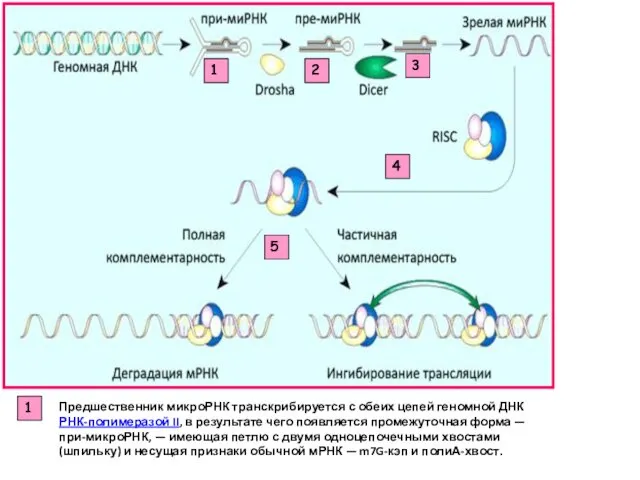
1
2
3
4
5
1
Предшественник микроРНК транскрибируется с обеих цепей геномной ДНК РНК-полимеразой II, в результате чего
1
2
3
4
5
1
Предшественник микроРНК транскрибируется с обеих цепей геномной ДНК РНК-полимеразой II, в результате чего

2
Эндонуклеаза Drosha отрезает от при-микроРНК одноцепочечные «хвосты». После чего структура транспортируется в
2
Эндонуклеаза Drosha отрезает от при-микроРНК одноцепочечные «хвосты». После чего структура транспортируется в

Функции микроРНК
1. МикроРНК - небелковый регулятор онтогенеза. На различных этапах жизни организмов
Функции микроРНК
1. МикроРНК - небелковый регулятор онтогенеза. На различных этапах жизни организмов

siРНК – малые интерферирующие РНК (даем латинское обозначение, русская аббревиатура пока
siРНК – малые интерферирующие РНК (даем латинское обозначение, русская аббревиатура пока

В нормальной клетке каждый ген выполняет свою собственную, строго определенную функцию,
В нормальной клетке каждый ген выполняет свою собственную, строго определенную функцию,

Образуются из длинных двуцепочечных РНК, источником которых являются:
РНК-содержащие вирусы,
Образуются из длинных двуцепочечных РНК, источником которых являются:
РНК-содержащие вирусы,

Функции siРНК
Защита от вирусов.
Репрессия трансгенов.
Подавление экспрессии мобильных элементов.
Функции siРНК
Защита от вирусов.
Репрессия трансгенов.
Подавление экспрессии мобильных элементов.

Почему же именно siРНК вызвала столь бурный всплеск интереса к себе?
Почему же именно siРНК вызвала столь бурный всплеск интереса к себе?
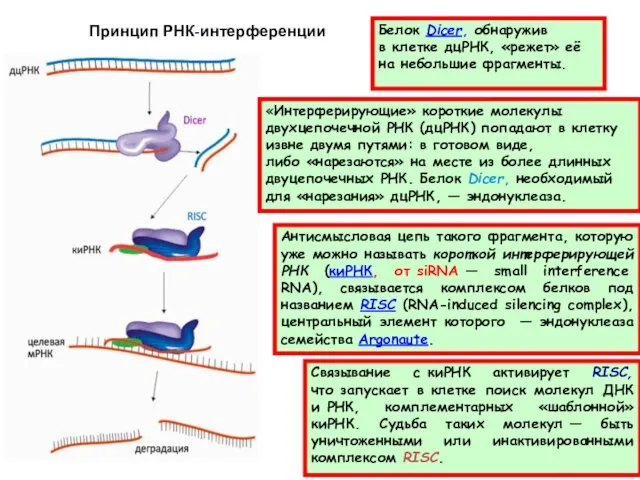
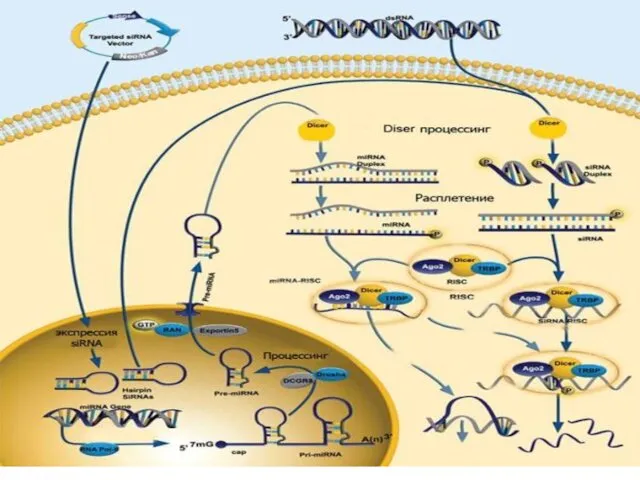
«Интерферирующие» короткие молекулы двухцепочечной РНК (дцРНК) попадают в клетку извне двумя путями:
«Интерферирующие» короткие молекулы двухцепочечной РНК (дцРНК) попадают в клетку извне двумя путями:

Комплексы Dicer и RISC могут использовать в качестве «затравки» не только дцРНК, но и одноцепочечную РНК,
Комплексы Dicer и RISC могут использовать в качестве «затравки» не только дцРНК, но и одноцепочечную РНК,

Функции киРНК (siРНК)
Это иммунная защита в клетках беспозвоночных. «Традиционная» иммунная система
Функции киРНК (siРНК)
Это иммунная защита в клетках беспозвоночных. «Традиционная» иммунная система

Практическое использование киРНК
На основе киРНК разработана технология «выключения» (или нокдауна) отдельных
Практическое использование киРНК
На основе киРНК разработана технология «выключения» (или нокдауна) отдельных

МикроРНК закодированны в уникальных генах геномов многоклеточных организмов (>1% от всех

siРНК в настоящее время широко используется для «нокаутирования» генов (их выключения)
siРНК в настоящее время широко используется для «нокаутирования» генов (их выключения)

Проводятся эксперименты на модельных животных по использованию siРНК в качестве инструмента
Проводятся эксперименты на модельных животных по использованию siРНК в качестве инструмента

Следует отметить, что спектр исследований в мире в области РНК-интерференции расширяется
Следует отметить, что спектр исследований в мире в области РНК-интерференции расширяется

Созданы библиотеки коротких siРНК и ДНК-векторов, кодирующих короткие РНК, мишенями
Созданы библиотеки коротких siРНК и ДНК-векторов, кодирующих короткие РНК, мишенями
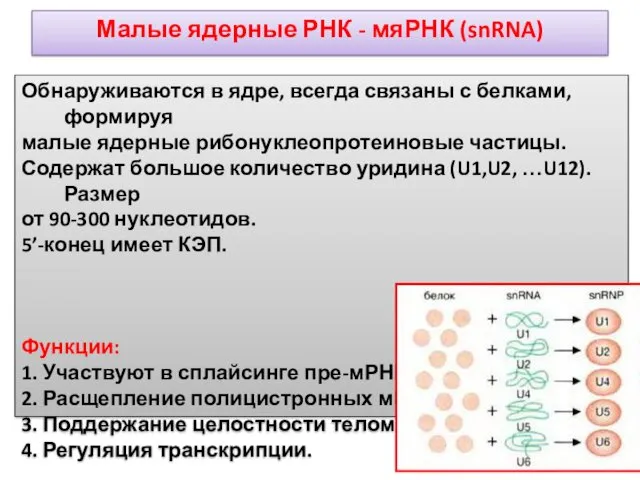
Малые ядерные РНК - мяРНК (snRNA)
Обнаруживаются в ядре, всегда связаны с
Малые ядерные РНК - мяРНК (snRNA)
Обнаруживаются в ядре, всегда связаны с

мякРНК имеют длину до 70 нуклеотидов.
мякРНК – это многочисленная группа
мякРНК имеют длину до 70 нуклеотидов.
мякРНК – это многочисленная группа
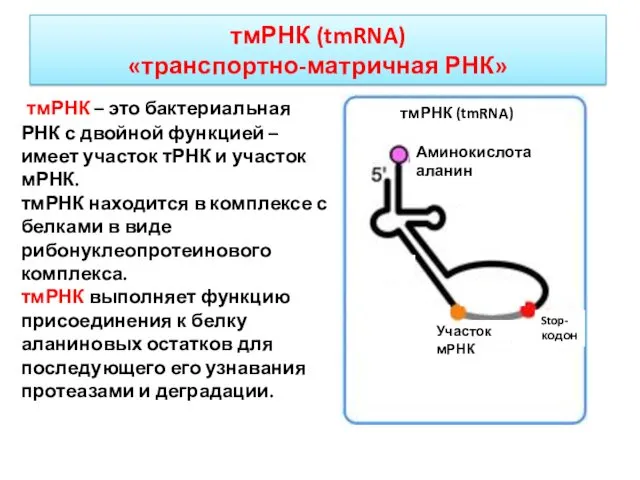
тмРНК – это бактериальная РНК с двойной функцией – имеет
тмРНК – это бактериальная РНК с двойной функцией – имеет
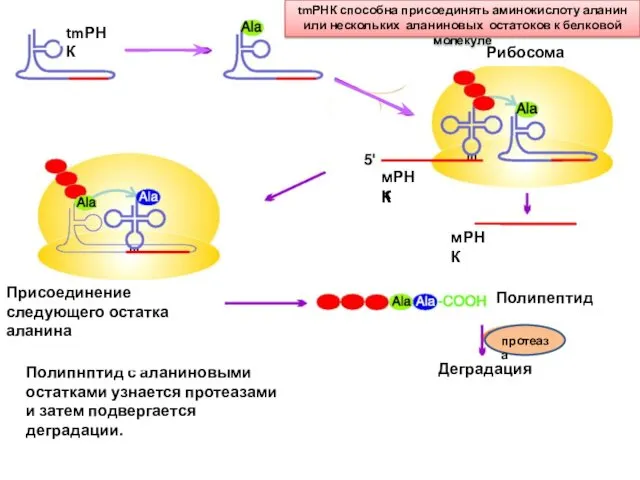
tmРНК
Рибосома
мРНК
мРНК
tmРНК способна присоединять аминокислоту аланин или нескольких аланиновых остатоков к белковой
tmРНК
Рибосома
мРНК
мРНК
tmРНК способна присоединять аминокислоту аланин или нескольких аланиновых остатоков к белковой

Pi-РНК - специфический агент против транспозиции
пиРНК (pi RNA) — короткие молекулы длиной в
Pi-РНК - специфический агент против транспозиции
пиРНК (pi RNA) — короткие молекулы длиной в

Гены пиРНК активны только в зародышевых клетках (во время эмбриогенеза) и окружающих их эндотелиальных клетках.
Важно!
Функции
Гены пиРНК активны только в зародышевых клетках (во время эмбриогенеза) и окружающих их эндотелиальных клетках.
Важно!
Функции
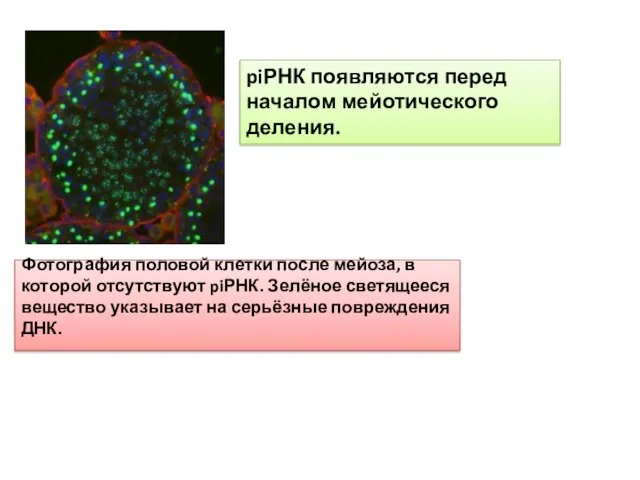
Фотография половой клетки после мейоза, в которой отсутствуют piРНК. Зелёное светящееся
Фотография половой клетки после мейоза, в которой отсутствуют piРНК. Зелёное светящееся

На сегодняшний день идентифицированы уже тысячи коротких регуляторных РНК, а механизм РНК-интерференции изучен
На сегодняшний день идентифицированы уже тысячи коротких регуляторных РНК, а механизм РНК-интерференции изучен

«Портрет» пиРНК
пиРНК — короткие молекулы длиной в 24–30 нуклеотидов, закодированные в центромерных и теломерных
«Портрет» пиРНК
пиРНК — короткие молекулы длиной в 24–30 нуклеотидов, закодированные в центромерных и теломерных
 Родники саратовской поэзии. Конкурс чтецов, посвященный 80-летию Саратовской области
Родники саратовской поэзии. Конкурс чтецов, посвященный 80-летию Саратовской области Функции современной семьи
Функции современной семьи Социально-экономические последствия безработицы и методы её преодоления
Социально-экономические последствия безработицы и методы её преодоления Презентация Александр Алексеевич Головачев - дважды герой Советского Союза
Презентация Александр Алексеевич Головачев - дважды герой Советского Союза Литературная игра-презентация Заверши стихотворение
Литературная игра-презентация Заверши стихотворение Фтор в профилактике кариеса. Роль и значение фторидов в профилактике кариеса. Механизм действия. (Лекция 10)
Фтор в профилактике кариеса. Роль и значение фторидов в профилактике кариеса. Механизм действия. (Лекция 10) Татарстан. Презентация.
Татарстан. Презентация. Синтоизм. Праздник Мацури
Синтоизм. Праздник Мацури Знатоки Правил дорожного движения
Знатоки Правил дорожного движения IKEA. Uczący się łańcuch dostaw (od drzewa w lesie do kasy w sklepie)
IKEA. Uczący się łańcuch dostaw (od drzewa w lesie do kasy w sklepie) Новые технологии в строительстве
Новые технологии в строительстве Трансакционные издержки. Теневая экономика
Трансакционные издержки. Теневая экономика Гидрология. Мировой океан
Гидрология. Мировой океан Поисковый проект Я помню! Я горжусь!
Поисковый проект Я помню! Я горжусь! Теория черт личности. Рэймонд Бернард Кэттелл
Теория черт личности. Рэймонд Бернард Кэттелл Сельское хозяйство. Растениеводство
Сельское хозяйство. Растениеводство Вазоренальная гипертензия
Вазоренальная гипертензия Роль и значение транспорта. Сухопутный транспорт
Роль и значение транспорта. Сухопутный транспорт Национальные парки
Национальные парки Планеты земной группы
Планеты земной группы Воспитательная программа Радуга
Воспитательная программа Радуга Определение уровня инновационного развития человеческого капитала: Чеченская республика
Определение уровня инновационного развития человеческого капитала: Чеченская республика Космические путешественники.
Космические путешественники. Организация Казахтелеком
Организация Казахтелеком Использование информационной технологии в работе с детьми с нарушением речи, как одно из альтернативных средств коммуникации.
Использование информационной технологии в работе с детьми с нарушением речи, как одно из альтернативных средств коммуникации. Графика
Графика Презентация Широка страна моя родная Диск
Презентация Широка страна моя родная Диск Персональный компьютер
Персональный компьютер