Неконтролируемое размножение генно-модифицированных деревьев - эпический провал биоинженеров презентация
- Неконтролируемое размножение генно-модифицированных деревьев - эпический провал биоинженеров

Содержание
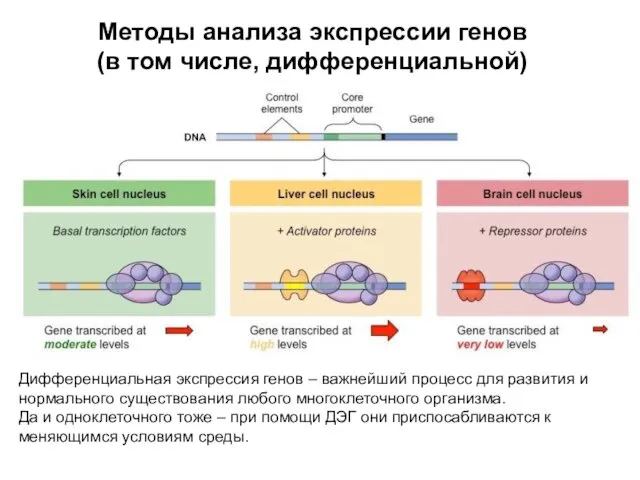
- 2. Методы анализа экспрессии генов (в том числе, дифференциальной) Дифференциальная экспрессия генов – важнейший процесс для развития
- 3. Как можно измерить экспрессию генов? По количеству РНК По количеству белка Какой подход вам нравится больше?
- 4. С одной стороны, по белку судить корректней – это же финальный продукт экспрессии генов. Но возникает
- 5. С РНК самой по себе дело иметь непросто: она не очень стабильная, и выделить ее из
- 6. ПЦР с обратной транскрипцией (RT-PCR) Используется для получения кДНК (комплементарная ДНК, cDNA), то есть ДНК, являющейся
- 7. ПРОБЛЕМЫ СИНТЕЗА СУММАРНОЙ кДНК • Для выделения полиА+ фракции нужно значительное количество РНК (>100 мкг). •
- 8. Метод выделения мРНК «oligo-dT» позволяет сильно обогатить вашу РНК молекулами матричной РНК
- 9. Метод «oligo-capping» решает проблему 5’-концевых последовательностей кДНК BAP – бактериальная щелочная фосфатаза, убирает фосфат с 5’-конца
- 10. Метод SMART решает все проблемы! Используется вирусная обратная транскриптаза, которая всегда навешивает несколько дополнительных нуклеотидов на
- 11. Хорошо. Выделили мРНК, амплифицировали кДНК. А мы вроде как собирались экспрессию генов смотреть. Этот человек как
- 12. EST (Expressed Sequence Tags) Метод массово использовался для анализа экспрессии генов до начала эпохи NGS. Подготовьте
- 13. Два методологических подхода к оценке дифференциальной экспрессии генов, которых мы еще не знаем 1. Радиоактивноcть А)
- 14. 1. Радиоактивноcть Б) Гибридизация ДНК на мембране с радиоактивно меченными олигонуклеотидами по принципу комплементарности. Саузерн-блот гибридизация
- 15. Вычитающая гибридизация денатурация То, что мы вычитаем То, из чего мы вычитаем
- 16. Вот такие замечательные результаты можно получать при помощи вычитающей гибридизации! 3 = 1 - 2
- 17. Дифференциальный дисплей позволяет сравнивать два препарата мРНК на геле
- 18. SAGE (Serial Analysis of Gene Expression) Выделите мРНК. Возьмите биотинилированный олигоТ-праймер и пришейте его к стрептавидин-сефарозе.
- 19. Прочное связывание стрептавидина с биотином (1:4) очень широко используется в молекулярной биологии
- 20. SAGE (Serial Analysis of Gene Expression) Проведите рестрикцию ферментом TE. Продукты рестрикции (а) не будут прикреплены
- 21. Результат секвенирования не только идентифицирует мРНК, но и покажет, сколько ее было в исходном образце!
- 22. Технологии ДНК-чипов Микрочип – это, как вы знаете, компьютерная микросхема. А ДНК-чип – это некая совокупность
- 23. DNA Macroarray (ДНК-макрочип) Да это же Саузерн-блот гибридизация!
- 24. 2. Флюоресценция Пометьте какой-нибудь нуклеотид этими молекулами и встройте в ДНК – вот и вся недолга!
- 25. Принцип технологии DNA-microarrays Выделите РНК откуда вам хочется. Проведите обратную транскрипцию. Включите меченные Cy3 или Cy5
- 26. Принцип технологии DNA-microarrays 5. Сканируйте красный и зеленый сигналы. 6. Наложите их друг на друга. 7.
- 27. Анализ экспрессии дрожжевых генов в зависимости от типа питания (аэробное / анаэробное)
- 28. Cy3 Cy5 Cy5 Cy3 log2 Анализ результатов использования ДНК-чипов Наложите друг на друга картинки с Cy3
- 29. В стандартной технологии ДНК-чипов в качестве находящегося на чипе материала могут использоваться: Фрагменты кДНК, ПЦР-продукты, Олигонуклеотиды.
- 30. Как олигонуклеотиды синтезируются на стекле? Изначально на стекло нанесены «точки связывания», и они закрыты «масками». Вторые
- 32. Скачать презентацию
Методы анализа экспрессии генов
(в том числе, дифференциальной)
Дифференциальная экспрессия генов – важнейший
Методы анализа экспрессии генов
(в том числе, дифференциальной)
Дифференциальная экспрессия генов – важнейший

Как можно измерить экспрессию генов?
По количеству РНК
По количеству белка
Какой подход вам
Как можно измерить экспрессию генов?
По количеству РНК
По количеству белка
Какой подход вам

С одной стороны, по белку судить корректней – это же финальный
С одной стороны, по белку судить корректней – это же финальный
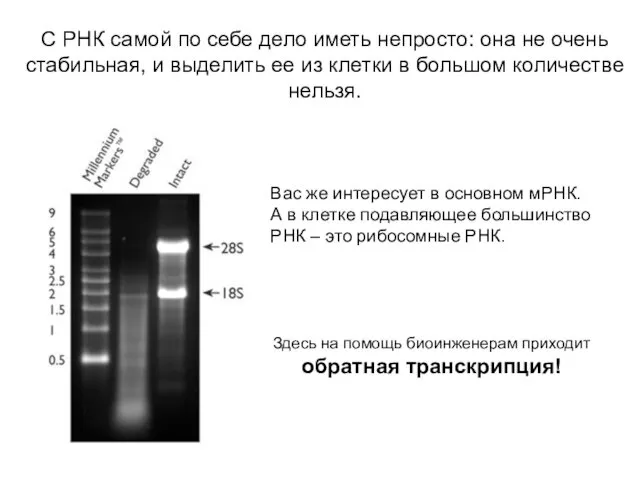
С РНК самой по себе дело иметь непросто: она не очень
С РНК самой по себе дело иметь непросто: она не очень
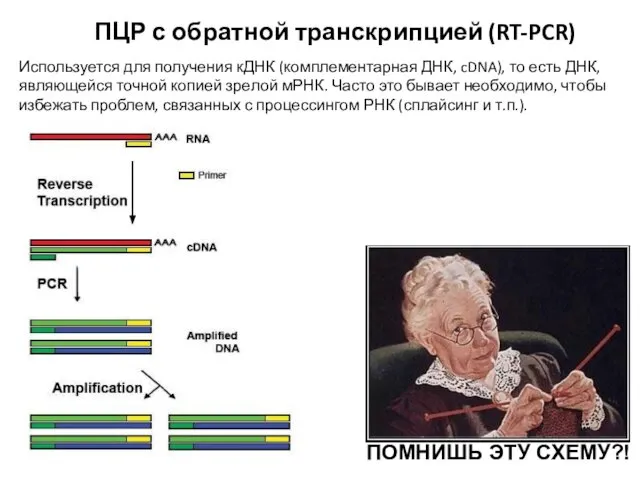
ПЦР с обратной транскрипцией (RT-PCR)
Используется для получения кДНК (комплементарная ДНК, cDNA),
ПЦР с обратной транскрипцией (RT-PCR)
Используется для получения кДНК (комплементарная ДНК, cDNA),

ПРОБЛЕМЫ СИНТЕЗА СУММАРНОЙ кДНК
• Для выделения полиА+ фракции нужно значительное количество
ПРОБЛЕМЫ СИНТЕЗА СУММАРНОЙ кДНК
• Для выделения полиА+ фракции нужно значительное количество
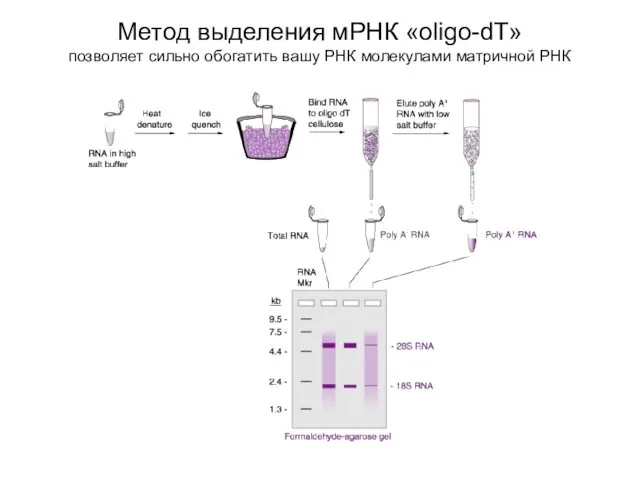
Метод выделения мРНК «oligo-dT»
позволяет сильно обогатить вашу РНК молекулами матричной
Метод выделения мРНК «oligo-dT»
позволяет сильно обогатить вашу РНК молекулами матричной
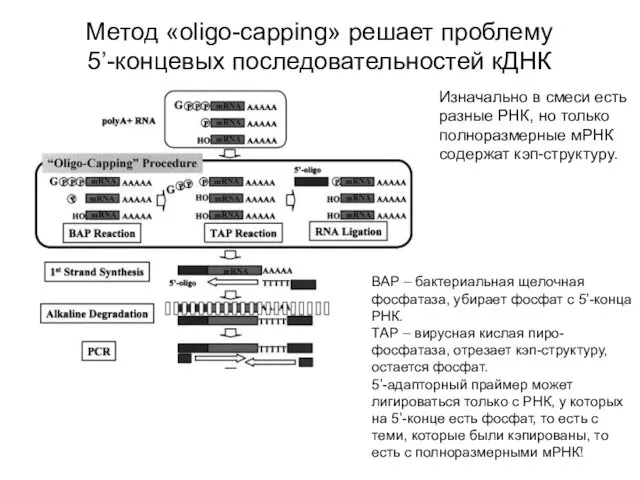
Метод «oligo-capping» решает проблему
5’-концевых последовательностей кДНК
BAP – бактериальная щелочная фосфатаза,
Метод «oligo-capping» решает проблему
5’-концевых последовательностей кДНК
BAP – бактериальная щелочная фосфатаза,
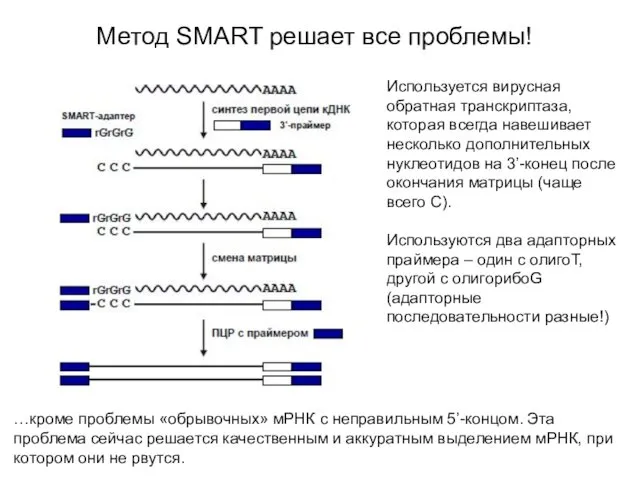
Метод SMART решает все проблемы!
Используется вирусная обратная транскриптаза, которая всегда навешивает
Метод SMART решает все проблемы!
Используется вирусная обратная транскриптаза, которая всегда навешивает

Хорошо. Выделили мРНК, амплифицировали кДНК.
А мы вроде как собирались экспрессию
Хорошо. Выделили мРНК, амплифицировали кДНК.
А мы вроде как собирались экспрессию

EST (Expressed Sequence Tags)
Метод массово использовался для анализа экспрессии генов до
EST (Expressed Sequence Tags)
Метод массово использовался для анализа экспрессии генов до
Два методологических подхода к оценке дифференциальной экспрессии генов,
которых мы еще не
Два методологических подхода к оценке дифференциальной экспрессии генов,
которых мы еще не
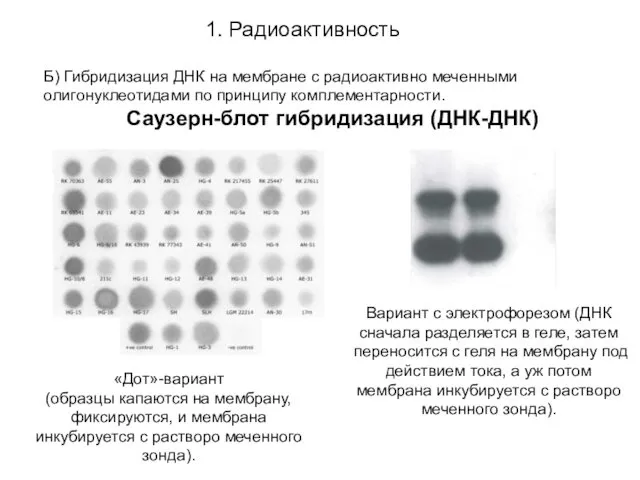
1. Радиоактивноcть
Б) Гибридизация ДНК на мембране с радиоактивно меченными олигонуклеотидами по
1. Радиоактивноcть
Б) Гибридизация ДНК на мембране с радиоактивно меченными олигонуклеотидами по
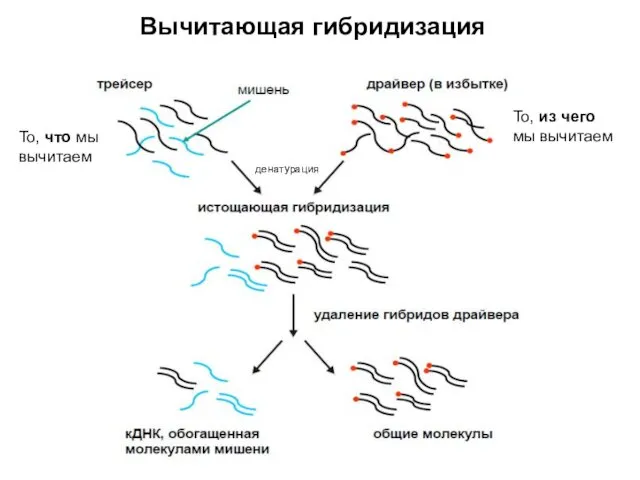
Вычитающая гибридизация
денатурация
То, что мы вычитаем
То, из чего мы вычитаем
Вычитающая гибридизация
денатурация
То, что мы вычитаем
То, из чего мы вычитаем
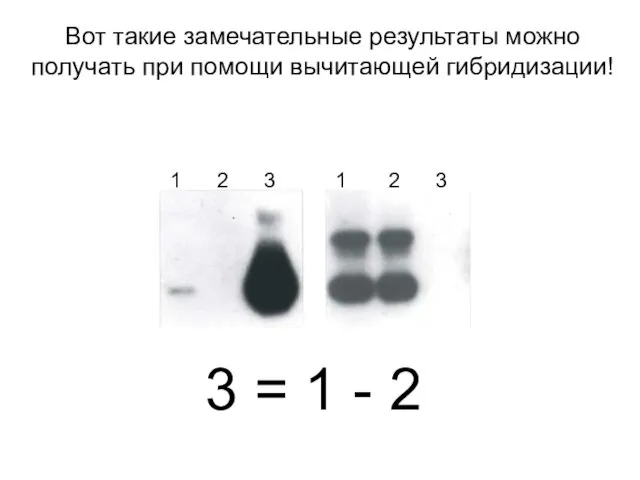
Вот такие замечательные результаты можно получать при помощи вычитающей гибридизации!
3 =
Вот такие замечательные результаты можно получать при помощи вычитающей гибридизации!
3 =
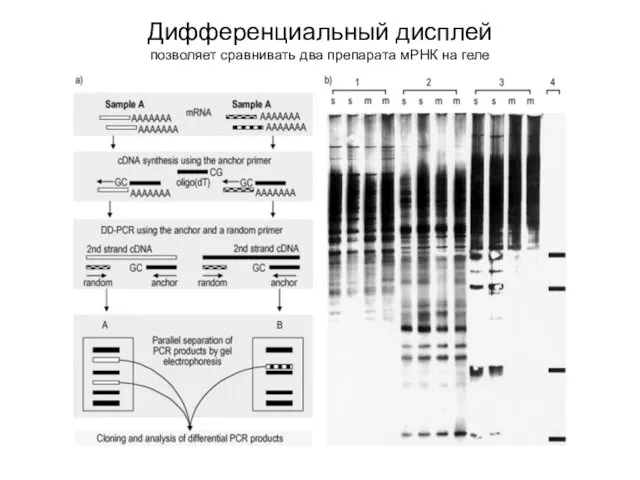
Дифференциальный дисплей
позволяет сравнивать два препарата мРНК на геле
Дифференциальный дисплей
позволяет сравнивать два препарата мРНК на геле
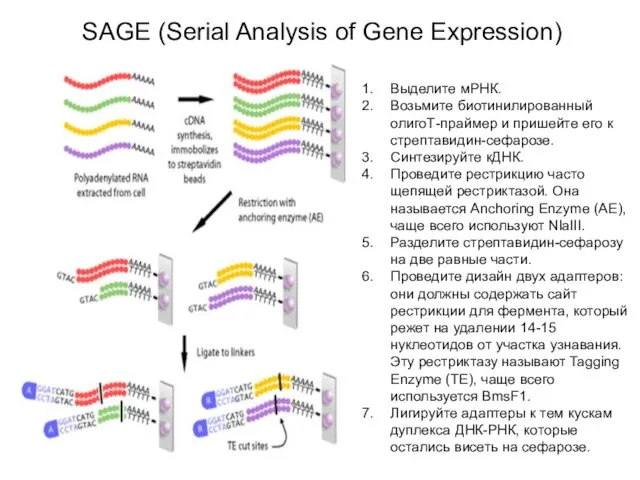
SAGE (Serial Analysis of Gene Expression)
Выделите мРНК.
Возьмите биотинилированный олигоТ-праймер и пришейте
SAGE (Serial Analysis of Gene Expression)
Выделите мРНК.
Возьмите биотинилированный олигоТ-праймер и пришейте
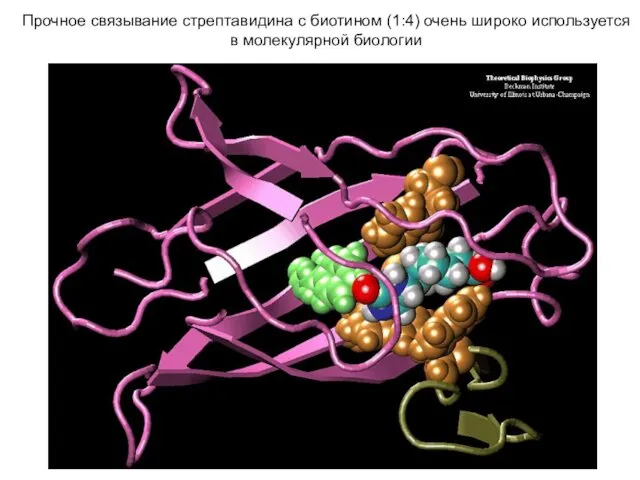
Прочное связывание стрептавидина с биотином (1:4) очень широко используется в молекулярной
Прочное связывание стрептавидина с биотином (1:4) очень широко используется в молекулярной
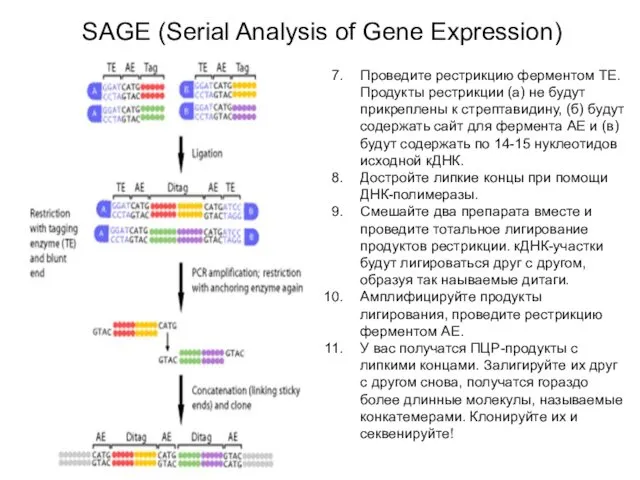
SAGE (Serial Analysis of Gene Expression)
Проведите рестрикцию ферментом TE. Продукты рестрикции
SAGE (Serial Analysis of Gene Expression)
Проведите рестрикцию ферментом TE. Продукты рестрикции
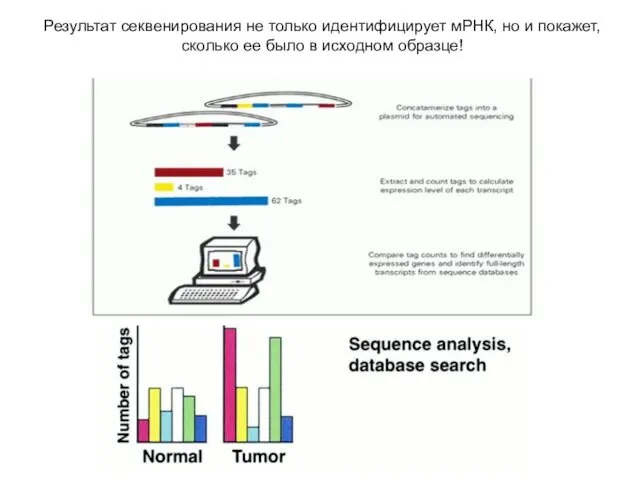
Результат секвенирования не только идентифицирует мРНК, но и покажет, сколько ее
Результат секвенирования не только идентифицирует мРНК, но и покажет, сколько ее

Технологии ДНК-чипов
Микрочип – это, как вы знаете, компьютерная микросхема.
А ДНК-чип –
Технологии ДНК-чипов
Микрочип – это, как вы знаете, компьютерная микросхема.
А ДНК-чип –
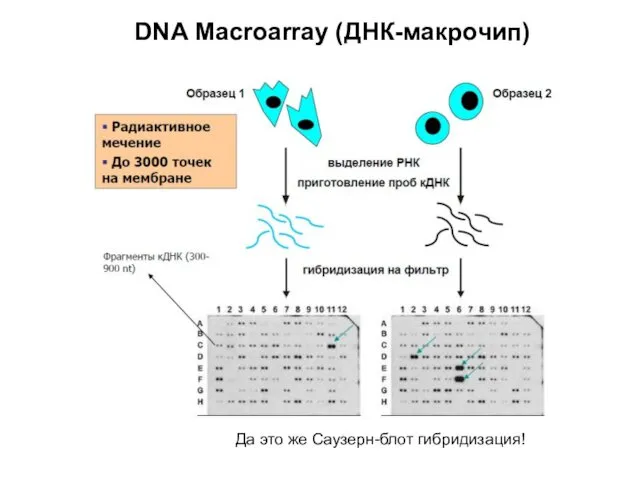
DNA Macroarray (ДНК-макрочип)
Да это же Саузерн-блот гибридизация!
DNA Macroarray (ДНК-макрочип)
Да это же Саузерн-блот гибридизация!
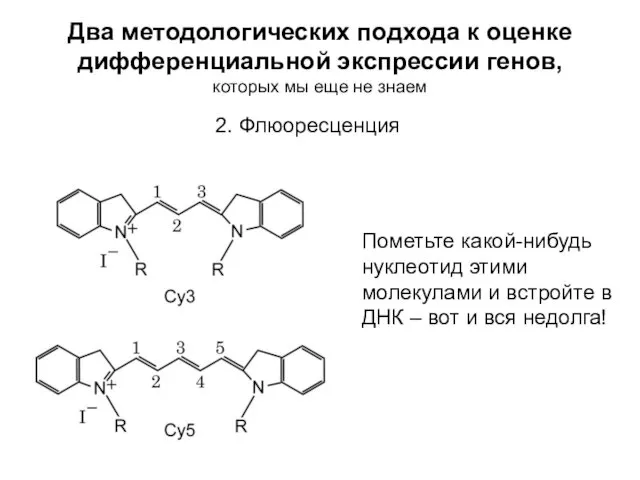
2. Флюоресценция
Пометьте какой-нибудь нуклеотид этими молекулами и встройте в ДНК –
2. Флюоресценция
Пометьте какой-нибудь нуклеотид этими молекулами и встройте в ДНК –
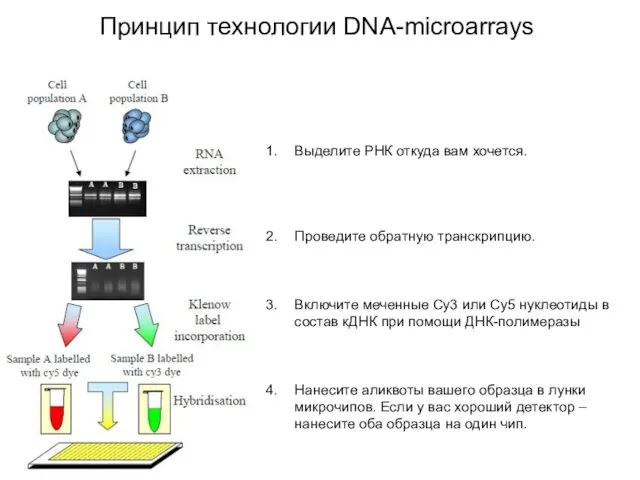
Принцип технологии DNA-microarrays
Выделите РНК откуда вам хочется.
Проведите обратную транскрипцию.
Включите меченные Cy3
Принцип технологии DNA-microarrays
Выделите РНК откуда вам хочется.
Проведите обратную транскрипцию.
Включите меченные Cy3
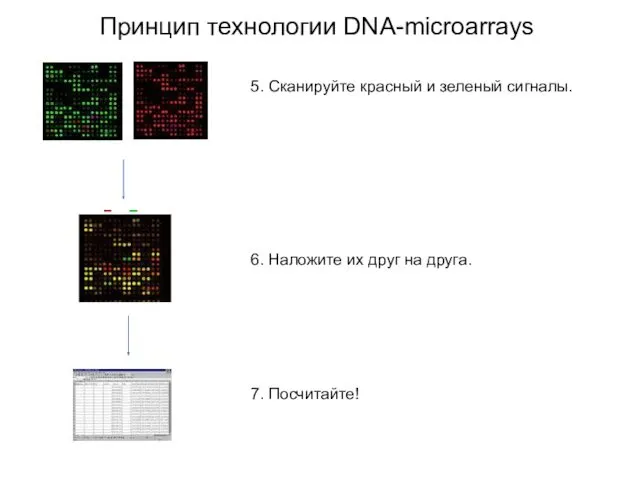
Принцип технологии DNA-microarrays
5. Сканируйте красный и зеленый сигналы.
6. Наложите их друг
Принцип технологии DNA-microarrays
5. Сканируйте красный и зеленый сигналы.
6. Наложите их друг

Анализ экспрессии дрожжевых генов в зависимости от типа питания (аэробное /
Анализ экспрессии дрожжевых генов в зависимости от типа питания (аэробное /

Cy3
Cy5
Cy5
Cy3
log2
Анализ результатов использования ДНК-чипов
Наложите друг на друга картинки с Cy3 и
Cy3
Cy5
Cy5
Cy3
log2
Анализ результатов использования ДНК-чипов
Наложите друг на друга картинки с Cy3 и
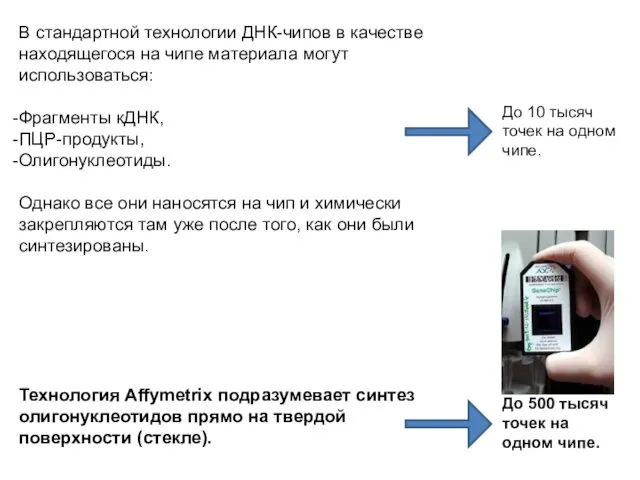
В стандартной технологии ДНК-чипов в качестве находящегося на чипе материала могут
В стандартной технологии ДНК-чипов в качестве находящегося на чипе материала могут
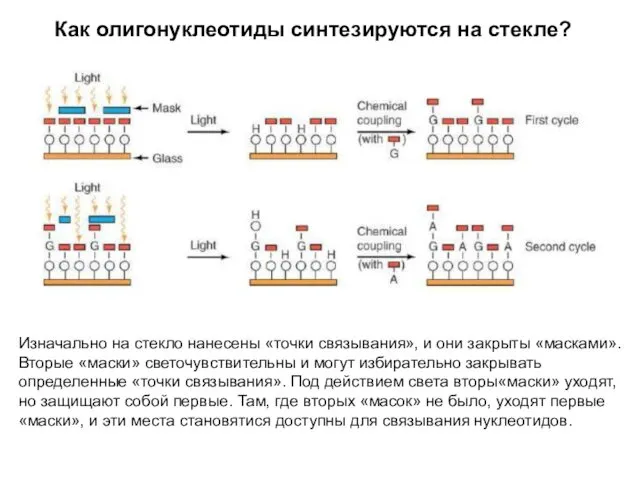
Как олигонуклеотиды синтезируются на стекле?
Изначально на стекло нанесены «точки связывания», и
Как олигонуклеотиды синтезируются на стекле?
Изначально на стекло нанесены «точки связывания», и
 Сестринский процесс при хроническом энтероколите
Сестринский процесс при хроническом энтероколите Методы синтеза и анализа цифровых фильтров. Введение
Методы синтеза и анализа цифровых фильтров. Введение Systemy transportowe – transport globalny
Systemy transportowe – transport globalny Колонизация Сибири русским народом
Колонизация Сибири русским народом Дедуктивные выводы из сложных суждений. Логика
Дедуктивные выводы из сложных суждений. Логика Михаил Васильевич Ломоносов
Михаил Васильевич Ломоносов Вербальная модель в сервисе
Вербальная модель в сервисе Глаз, как оптическая система
Глаз, как оптическая система 2015.03.20.Open house.Odesa.Application
2015.03.20.Open house.Odesa.Application Паровой двигатель
Паровой двигатель Организация объединенных наций (ООН)
Организация объединенных наций (ООН) Пластиковый Апокалипсис
Пластиковый Апокалипсис Развитие коммуникативных игр
Развитие коммуникативных игр Мини Мастер конструктор керамический Печь для итальянской пиццы, как настоящий, 1 : 10
Мини Мастер конструктор керамический Печь для итальянской пиццы, как настоящий, 1 : 10 Литосфера. Основные особенности литосферы. Влияние деятельности человека
Литосфера. Основные особенности литосферы. Влияние деятельности человека Дикорастущие плодовые и орехоплодные растения разных типов леса, их запасы и заготовка
Дикорастущие плодовые и орехоплодные растения разных типов леса, их запасы и заготовка Стипендия. Виды стипендий
Стипендия. Виды стипендий Классификация извещателей пожарных (ИП)
Классификация извещателей пожарных (ИП) Манометры и гидравлическое оборудование
Манометры и гидравлическое оборудование Презентация Стихийные бедствия 2 часть
Презентация Стихийные бедствия 2 часть Числа 3 и 8 (1 класс)
Числа 3 и 8 (1 класс) Увольнение с военной службы
Увольнение с военной службы Интерактивный тренажер Процент от числа
Интерактивный тренажер Процент от числа Материалы и изделия из древесины. Тема 2
Материалы и изделия из древесины. Тема 2 Кишечный шов
Кишечный шов Мотивация учения- основное условие успешного обучения Выступление на педсовете
Мотивация учения- основное условие успешного обучения Выступление на педсовете Салтыков – Щедрин История одного города
Салтыков – Щедрин История одного города Строительство высокоскоростной магистрали ВСМ-2 Москва-Казань
Строительство высокоскоростной магистрали ВСМ-2 Москва-Казань