- Нейрогуморальная регуляция полового процесса

Содержание
- 2. Проявление признаков полового цикла: течки, выделение яйцеклетки из яичника.
- 3. Поддержание беременности и другие изменения в организме самки во время воспроизводительного цикла у коров регулируются гормонами.
- 4. Примечательно, что зачастую ткань может являться одновременно эндокринной железой и тканью – получателем (яичник). Из эндокринных
- 5. Действие гормонов То, что гормон вырабатывается тканью, вовсе не означает, что он произведет свое действие где-либо
- 6. Поэтому, например, что бы ткань реагировала на эстроген, ее клетки должны обладать клетками эстрогена. После того
- 7. Органы, вырабатывающие гормоны и влияющие на процесс воспроизводства. Четыре эндокринные железы, вырабатывающие гормоны, которые регулируют 21-дневный
- 8. Гипоталамус находится в основании мозга. Он содержит нервные окончания и очень чувствителен к сигналам, передаваемым гормонами.
- 9. Гипофиз расположен под гипоталамусом, располагается в основании черепа на дне турецкого седла. В настоящее время установлено,
- 10. Основные половые гормоны, выделяемые передней долей гипофиза, называются гонадотропинами. Они стимулируют органы, производящие гаметы (яичники) у
- 11. Превращении овулированного фолликула в желтое тело и секреции им прогестерона, в развитии фолликулов и секреции эстрогенов,
- 12. Основное действие ФСГ заключается в стимуляции развития множественных фолликулов, приводя к пролиферации клеток гранулезы В свете
- 13. Фолликул в яичнике – Эстроген Эстроген – это гормон, выделяемый фолликулами по мере их созревания в
- 14. Таким образом, эстроген координирует одновременный допуск коровой к себе быка и выход яйцеклетки из яичника. Такой
- 15. Желтое тело-Прогестерон После завершения овуляции ткань, которая мгновение назад была частью фолликула, начинает подвергаться резкому изменению
- 16. Желтое тело-Прогестерон Подготавливает матку к беременности, Предотвращает созревание новых фолликулов и, следовательно, повторение полового цикла (течки),
- 17. Желтое тело персистентное (задержавшееся)
- 18. Таким образом, при наступлении беременности желтое тело остается активным на протяжении всего периода беременности. Если беременность
- 19. Матка-простагландины Спустя по меньшей мере 16-18 дней после начала нового цикла матка коровы может определить наличие
- 20. В результате подавляющий эффект прогестерона устраняется, фолликулы получают возможность продолжать развитие до полного созревания и через
- 21. Гормональная регуляция полового цикла (течки) Эстроген доминирует на предтечковой и течковой фазах цикла, тогда как прогестерон
- 22. В течение предтечкового периода желтое тело рассасывается в результате воздействия простагландина, выделяемого маткой нестельной коровы. Содержание
- 23. Сводка основных гормонов, их источников, органов-получателей и вызываемых эффектов приведена в таблице
- 24. Развитие фолликулов на яичнике Хотя одновременно могут развиваться несколько больших фолликулов, только один (два или три
- 25. Концентрация эстрогена в пикограммах на миллилитр крови (пг/мл) возрастает и достигает пика, равного приблизительно 7-8 пг/мл,
- 26. Формирование желтого тела после овуляции Отделение фолликула от яичника, или овуляция, происходит через 24-32 часа после
- 27. Половой цикл Если организм коровы определяет, что она беременна, то желтое тело продолжает функционировать по сигналу,
- 28. Половой цикл у сельскохозяйственных животных включает две основные фазы: лютеиновую и фолликулярную, которые, достаточно четко гормонально
- 29. Известные способы синхронизации половой охоты и овуляции у домашних животных основаны на двух подходах. Первый в
- 30. Этот подход базируется на торможении функции яичников, последующем прекращении этого ингибирующего влияния и обеспечении роста и
- 31. Второй подход основан на вызывании преждевременного удаления желтого тела и последующего роста и развития фолликулов также
- 32. Цикличная активность яичников у коров, особенно у высокопродуктивных молочных пород и у подсосных мясных коров задерживается
- 33. Циклическая активность яичников у коров в послеотельный период может быть ускорена гонадотропными гормонами, но эта обработка
- 34. Неблагоприятное влияние анэструса на эффективность оплодотворения может быть преодолено путем применения прогестагенов, отдельно или в комплексе
- 36. Скачать презентацию

Проявление признаков полового цикла: течки, выделение яйцеклетки из яичника.
Проявление признаков полового цикла: течки, выделение яйцеклетки из яичника.

Поддержание беременности и другие изменения в организме самки во время воспроизводительного
Поддержание беременности и другие изменения в организме самки во время воспроизводительного

Примечательно, что зачастую ткань может являться одновременно эндокринной железой и
Примечательно, что зачастую ткань может являться одновременно эндокринной железой и
Действие гормонов
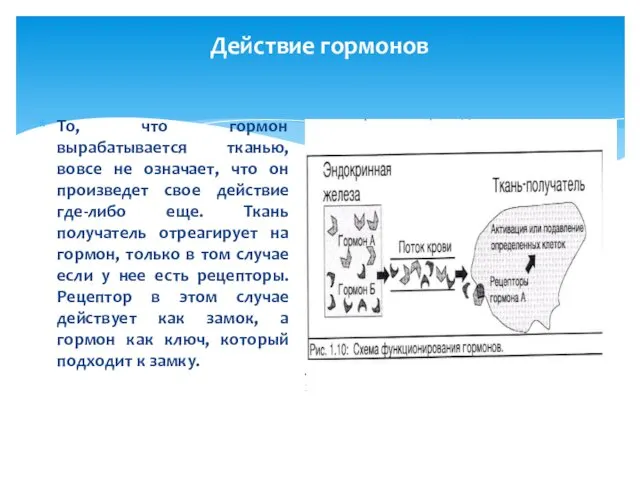
То, что гормон вырабатывается тканью, вовсе не означает, что он
Действие гормонов
То, что гормон вырабатывается тканью, вовсе не означает, что он

Поэтому, например, что бы ткань реагировала на эстроген, ее клетки должны
Поэтому, например, что бы ткань реагировала на эстроген, ее клетки должны

Органы, вырабатывающие гормоны и влияющие на процесс воспроизводства.
Четыре эндокринные железы, вырабатывающие
Органы, вырабатывающие гормоны и влияющие на процесс воспроизводства.
Четыре эндокринные железы, вырабатывающие

Гипоталамус находится в основании мозга. Он содержит нервные окончания и очень
Гипоталамус находится в основании мозга. Он содержит нервные окончания и очень

Гипофиз расположен под гипоталамусом, располагается в основании черепа на дне турецкого
Гипофиз расположен под гипоталамусом, располагается в основании черепа на дне турецкого

Основные половые гормоны, выделяемые передней долей гипофиза, называются гонадотропинами. Они стимулируют
Основные половые гормоны, выделяемые передней долей гипофиза, называются гонадотропинами. Они стимулируют

Превращении овулированного фолликула в желтое тело и секреции им прогестерона, в
Превращении овулированного фолликула в желтое тело и секреции им прогестерона, в

Основное действие ФСГ заключается в стимуляции развития множественных фолликулов, приводя к
Основное действие ФСГ заключается в стимуляции развития множественных фолликулов, приводя к

Фолликул в яичнике – Эстроген
Эстроген – это гормон, выделяемый фолликулами по
Фолликул в яичнике – Эстроген
Эстроген – это гормон, выделяемый фолликулами по

Таким образом, эстроген координирует одновременный допуск коровой к себе быка и
Таким образом, эстроген координирует одновременный допуск коровой к себе быка и
Желтое тело-Прогестерон
После завершения овуляции ткань, которая мгновение назад была частью фолликула,
Желтое тело-Прогестерон
После завершения овуляции ткань, которая мгновение назад была частью фолликула,
Желтое тело-Прогестерон
Подготавливает матку к беременности,
Предотвращает созревание новых фолликулов и, следовательно, повторение
Желтое тело-Прогестерон
Подготавливает матку к беременности,
Предотвращает созревание новых фолликулов и, следовательно, повторение
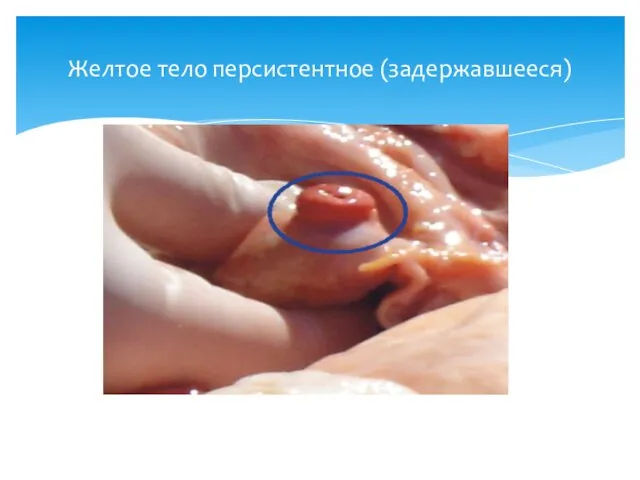
Желтое тело персистентное (задержавшееся)
Желтое тело персистентное (задержавшееся)

Таким образом, при наступлении беременности желтое тело остается активным на протяжении
Таким образом, при наступлении беременности желтое тело остается активным на протяжении
Матка-простагландины
Спустя по меньшей мере 16-18 дней после начала нового цикла матка
Матка-простагландины
Спустя по меньшей мере 16-18 дней после начала нового цикла матка

В результате подавляющий эффект прогестерона устраняется, фолликулы получают возможность продолжать развитие
В результате подавляющий эффект прогестерона устраняется, фолликулы получают возможность продолжать развитие
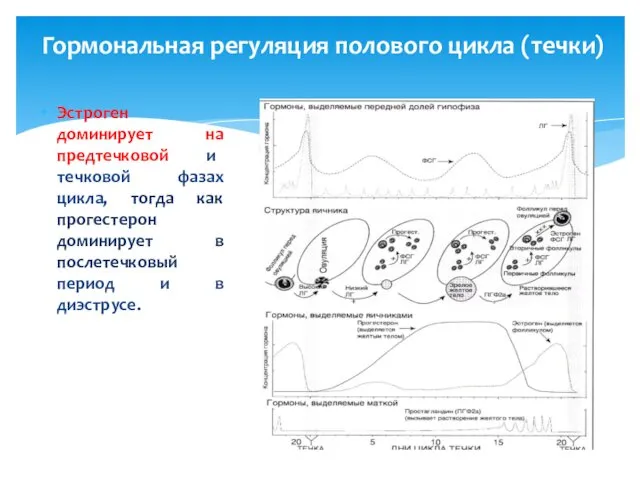
Гормональная регуляция полового цикла (течки)
Эстроген доминирует на предтечковой и течковой фазах
Гормональная регуляция полового цикла (течки)
Эстроген доминирует на предтечковой и течковой фазах

В течение предтечкового периода желтое тело рассасывается в результате воздействия простагландина,
В течение предтечкового периода желтое тело рассасывается в результате воздействия простагландина,
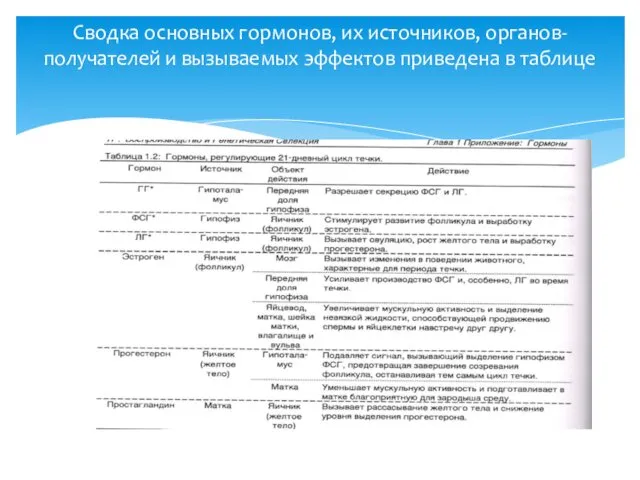
Сводка основных гормонов, их источников, органов-получателей и вызываемых эффектов приведена в
Сводка основных гормонов, их источников, органов-получателей и вызываемых эффектов приведена в

Развитие фолликулов на яичнике
Хотя одновременно могут развиваться несколько больших фолликулов, только
Развитие фолликулов на яичнике
Хотя одновременно могут развиваться несколько больших фолликулов, только

Концентрация эстрогена в пикограммах на миллилитр крови (пг/мл) возрастает и достигает
Концентрация эстрогена в пикограммах на миллилитр крови (пг/мл) возрастает и достигает
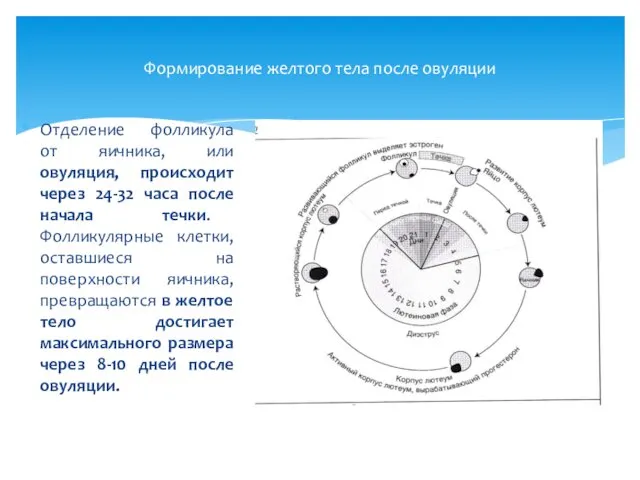
Формирование желтого тела после овуляции
Отделение фолликула от яичника, или овуляция, происходит
Формирование желтого тела после овуляции
Отделение фолликула от яичника, или овуляция, происходит
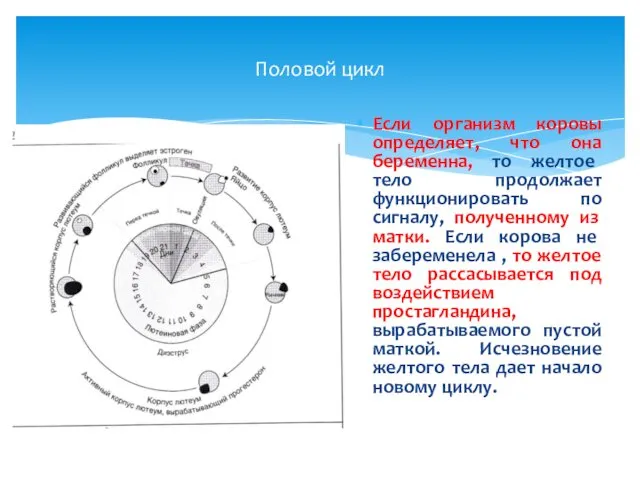
Половой цикл
Если организм коровы определяет, что она беременна, то желтое тело
Половой цикл
Если организм коровы определяет, что она беременна, то желтое тело

Половой цикл у сельскохозяйственных животных включает две основные фазы: лютеиновую и
Половой цикл у сельскохозяйственных животных включает две основные фазы: лютеиновую и

Известные способы синхронизации половой охоты и овуляции у домашних животных основаны
Известные способы синхронизации половой охоты и овуляции у домашних животных основаны

Этот подход базируется на торможении функции яичников, последующем прекращении этого ингибирующего
Этот подход базируется на торможении функции яичников, последующем прекращении этого ингибирующего

Второй подход основан на вызывании преждевременного удаления желтого тела и последующего
Второй подход основан на вызывании преждевременного удаления желтого тела и последующего

Цикличная активность яичников у коров, особенно у высокопродуктивных молочных пород и
Цикличная активность яичников у коров, особенно у высокопродуктивных молочных пород и

Циклическая активность яичников у коров в послеотельный период может быть ускорена
Циклическая активность яичников у коров в послеотельный период может быть ускорена

Неблагоприятное влияние анэструса на эффективность оплодотворения может быть преодолено путем применения
Неблагоприятное влияние анэструса на эффективность оплодотворения может быть преодолено путем применения
 Формирование учебно-познавательных компетенций на уроке математики
Формирование учебно-познавательных компетенций на уроке математики Т хәрефе. 1 сыйныф.Татар төркеме
Т хәрефе. 1 сыйныф.Татар төркеме Лесная промышленность России
Лесная промышленность России Рельеф, геологическое строение и полезные ископаемые России. Обощающий урок. 8 класс.
Рельеф, геологическое строение и полезные ископаемые России. Обощающий урок. 8 класс. Крещение Иисуса Христа
Крещение Иисуса Христа Особенности подготовки публичной речи
Особенности подготовки публичной речи Презентация педагогического проекта Мир эмоций
Презентация педагогического проекта Мир эмоций Влажно-тепловая обработка швейных изделий
Влажно-тепловая обработка швейных изделий Нагнетатели. Мельничные вентиляторы в машине
Нагнетатели. Мельничные вентиляторы в машине Завод гидроизоляции Арена. Производство строительных смесей для гидроизоляции, ремонта и защиты бетонных конструкций
Завод гидроизоляции Арена. Производство строительных смесей для гидроизоляции, ремонта и защиты бетонных конструкций Программа для детей с ОВЗ Дети с ринолалией в ДОУ
Программа для детей с ОВЗ Дети с ринолалией в ДОУ Город Белоусово в Жуковском районе Калужской области России
Город Белоусово в Жуковском районе Калужской области России Я и мои страхи
Я и мои страхи Факультатив Олимпиадное программирование. Олимпиада школьников по математике и криптографии
Факультатив Олимпиадное программирование. Олимпиада школьников по математике и криптографии Методологические исторического развития основы таможенного анализа дела и таможенной политики
Методологические исторического развития основы таможенного анализа дела и таможенной политики Гуманизм: абстрактный или классовый, по роману А.А. Фадеева Разгром
Гуманизм: абстрактный или классовый, по роману А.А. Фадеева Разгром Эксплуатация различных видов серверного программного обеспечения АИС и ее особенности
Эксплуатация различных видов серверного программного обеспечения АИС и ее особенности Десмургия. Десмургия дегеніміз
Десмургия. Десмургия дегеніміз Мировые религии
Мировые религии Права, смежные с авторскими
Права, смежные с авторскими Кислотные дожди
Кислотные дожди Презентация к уроку окружающий мир 3 класс на тему Разнообразие животных
Презентация к уроку окружающий мир 3 класс на тему Разнообразие животных Презентация Лесостепи и степи России
Презентация Лесостепи и степи России Методика выявления Коммуникативных и организаторских способностей (КОС)
Методика выявления Коммуникативных и организаторских способностей (КОС) Моя семья
Моя семья Анализ нарушений по организации социального пространства рабочих мест, бытовых и офисных помещений
Анализ нарушений по организации социального пространства рабочих мест, бытовых и офисных помещений Утренний сбор в начальной школе. Презентация
Утренний сбор в начальной школе. Презентация Вводные слова
Вводные слова