- Обмен веществ. Взаимосвязи липидного, углеводного и азотистого обмена. (Лекция 1)

Содержание
- 2. Стадии обмена Поступление веществ с пищей, переваривание, всасывание Межуточный обмен – переработка веществ и энергии в
- 3. Белки Полисахариды Липиды Большие молекулы Аминокислоты Глюкоза Глицерин, жирные кислоты Молекулы, играющие роль строительных блоков пируват
- 4. Липиды Углеводы Белки Глюкоза Гликоген Глюкозо-6-Ф НАДФH Пируват Лактат НАДH Триозофосфат Ацетил-CoA Ацетоацетат Стерины Жирные кислоты
- 5. Анаболическая фаза обмена веществ начинается с потреблением пищи Катаболическая фаза обмена веществ через 4-6 часов после
- 6. Роль печени в регуляции обмена веществ.
- 9. Печень выполняет важнейшую роль в адаптации обменных процессов к режиму питания, перераспределяя потоки питательных веществ между
- 10. Связи печени и жировой ткани
- 11. аэробный процесс окисления ЖК анаэробный гликолиз кетолиз – утилизация кетоновых тел, образующихся из ЖК
- 12. печень мозг глюкоза аммиак в составе глутамата и глутамина мочевина Связи печени и мозга голодание нейромедиаторные
- 13. печень почка глутамин глюкоза выведение H+ в форме NH4+ ацидоз, диабет Связь печени и почек
- 14. Направленность и интенсивность обменных процессов зависят от: доступности субстратов и энергии, активности ферментов. Регуляция активности ферментов:
- 15. Сродство к лиганду Соединения с исключительно сигнальной функцией (гормоны) Обычные метаболиты Минорные метаболиты Рецепторы Сенсоры Рецепторы
- 16. Аденилат-циклазы цАМФ Фосфо-липазы Cβδ IP3 Ca2+ Эффекты Гормон G-белок Рецептор Вторые посредники Глюкоза Фруктозо-6-Ф Фруктозо-1,6-Ф2 PFK1
- 17. глюкоза
- 18. Как и рецепторные протеинкиназы, сенсоры обычных метаболитов могут действовать на множество мишеней
- 19. Азотистый обмен
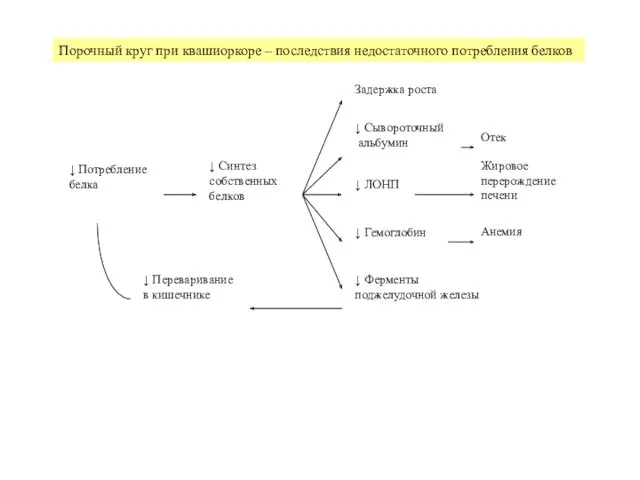
- 20. Азотистый баланс – отношение потребленного азота к экскретируемому,- важный показатель ростовых и анаболических процессов Срок жизни
- 21. Гормоны с анаболической и катаболической направленностью действия
- 23. Переваривание белков Пища Пепсиногены I и II желудка (синтез и секреция) Пепсины цАМФ (секретин, ВИП, катехоламины)
- 24. Na+ H+ H+ олигопептид Na+ K+ Na+,K+-АТФаза H+ Na+/H+-обменник PepT1, H+-пептидный котранспортер олигопептид аминокислоты пептидаза Базолатеральная
- 25. Системы транспорта аминокислот в кишечнике, почках, синапсах, других тканях Другие субстраты: тиреоидные гормоны, биогенные амины
- 26. Щеточная каемка Просвет кишечника Na+/H+ обменник (NHE3) Сопряженный с протонами транспортер аминокислот (PAT1) а.к. H+ Na+
- 27. SN1 Na+ H+ а.к. A ATA1 Na+ H+ а.к. Б Варианты зависимых от Na+ транспортеров аминокислот.
- 28. А: Димерный транспортер аминокислот b0.+ построен из тяжелой цепи rbAT (красный цвет) и легкой цепи b0.+AT
- 29. Межуточный обмен, образование конечных продуктов и их выведение
- 30. Строительный материал для белков и азотсодержащих соединений; источник энергии
- 31. обратимые реакции переаминирования окислительное дезаминирование – удаление избытка аминокислот Баланс между аминокислотами поддерживается за счет взаимопревращений.
- 32. Взаимопревращения глутамата и глутамина цитозоль митохондрии Глюкагон, глюкокортикоиды Глютамин – источник энергии, предшественник для глюконеогенеза, переносчик
- 33. Регуляция обмена аммиака в печени и почках ГЧЭ ГК-Рц ГЧЭ интрона 1 обеспечивает индукцию глюкокортикоидами экспрессии
- 34. Сдвиги в метаболических процессах запускают процесс неопластической трансформации? Oncogene (2015), 1–7
- 35. World Journal of Surgical Oncology (2016) 14:15
- 36. Недостаточность ферментов ( гипераммониемия, цитруллинемия, аргининсукциникэсидурия, аргининемия) - рвота, летаргия, припадки, задержка умственного развития и психомоторной
- 37. Отдаленный энхансер обеспечивает стимуляцию транскрипции гена карбамоилфосфатсинтазы глюкокортикоидами и глюкагоном Энхансер гена аргиназы-1 обеспечивает стимуляцию глюкагоном
- 38. Корковый слой кортикальная собирательная трубочка наружная медуллярная собирательная трубочка внутренняя медуллярная собирательная трубочка Мозговой слой Экскреция
- 39. Регуляция циркадных ритмов и метаболизма с участием ядерного рецептора Rev-erbα
- 40. Figure. Bidirectional relationship between circadian clock, sleep-wake, and fasting-feeding behavior underlies maintenance of energy balance in
- 41. Сигналы света с сетчатки через ретиногипоталамический тракт попадают в супрахиазматическое ядро мозга – центральная синхронизация циркадных
- 42. Гетеродимер CLOCK- BMAL1связывается с энхансером E-box, локализованным в промоторном районе генов PER и CRY, активируя их
- 43. Rev-erbα координирует циркадные ритмы и метаболизм гем-зависимым путем. Связывающий элемент Rev-erbα - RORE, присутствует не только
- 44. Ауторегуляция уровня гема с участием сенсора гема Rev-erbα мРНК PGC-1α Гем Rev-erb Rev-erb Гем Гем мРНК
- 46. Скачать презентацию
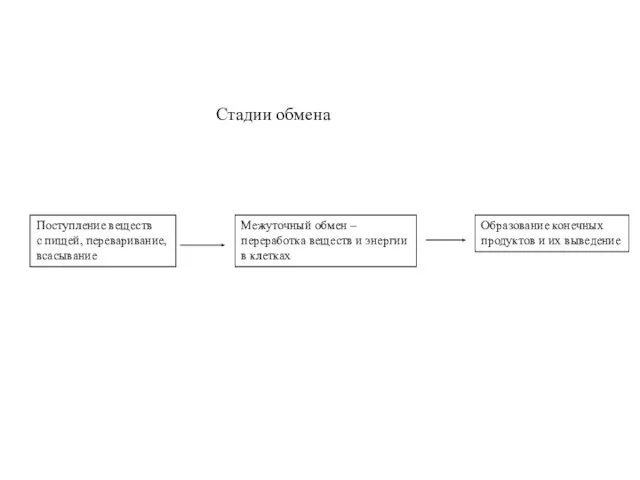
Стадии обмена
Поступление веществ
с пищей, переваривание,
всасывание
Межуточный обмен –
переработка веществ и
Стадии обмена
Поступление веществ
с пищей, переваривание,
всасывание
Межуточный обмен –
переработка веществ и
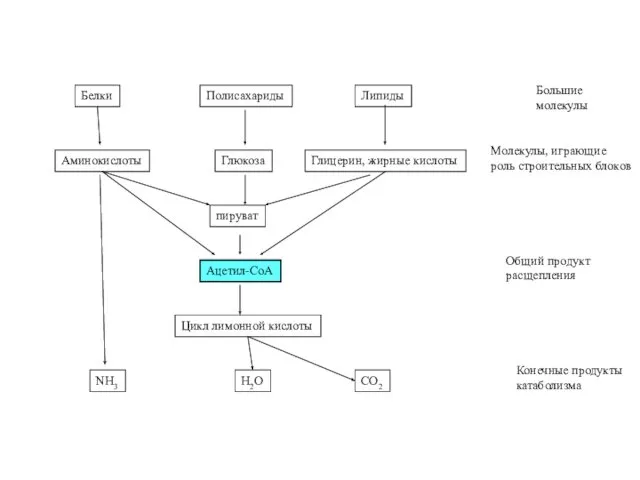
Белки
Полисахариды
Липиды
Большие
молекулы
Аминокислоты
Глюкоза
Глицерин, жирные кислоты
Молекулы, играющие
роль строительных блоков
пируват
Ацетил-СоА
Общий продукт
расщепления
Цикл лимонной кислоты
NH3
H2O
CO2
Конечные
Белки
Полисахариды
Липиды
Большие
молекулы
Аминокислоты
Глюкоза
Глицерин, жирные кислоты
Молекулы, играющие
роль строительных блоков
пируват
Ацетил-СоА
Общий продукт
расщепления
Цикл лимонной кислоты
NH3
H2O
CO2
Конечные
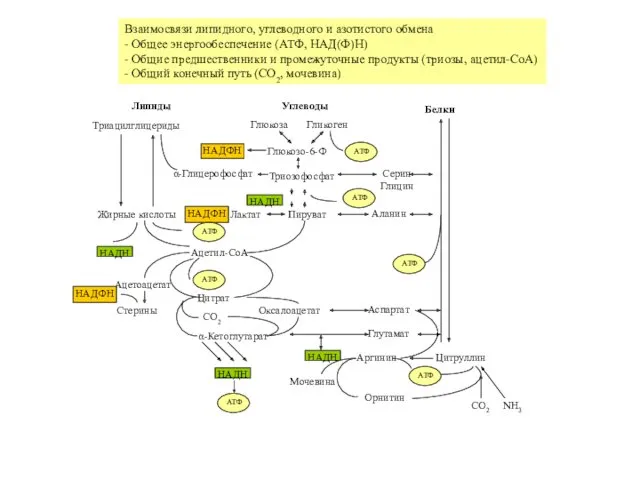
Липиды
Углеводы
Белки
Глюкоза
Гликоген
Глюкозо-6-Ф
НАДФH
Пируват
Лактат
НАДH
Триозофосфат
Ацетил-CoA
Ацетоацетат
Стерины
Жирные кислоты
Оксалоацетат
Цитрат
α-Кетоглутарат
CO2
НАДH
НАДФH
Триацилглицериды
НАДФH
НАДH
α-Глицерофосфат
Аспартат
Глутамат
НАДH
Цитруллин
Аргинин
Орнитин
CO2
NH3
Мочевина
Аланин
Серин
Глицин
Взаимосвязи липидного, углеводного и азотистого обмена
- Общее энергообеспечение (АТФ, НАД(Ф)H)
-
Липиды
Углеводы
Белки
Глюкоза
Гликоген
Глюкозо-6-Ф
НАДФH
Пируват
Лактат
НАДH
Триозофосфат
Ацетил-CoA
Ацетоацетат
Стерины
Жирные кислоты
Оксалоацетат
Цитрат
α-Кетоглутарат
CO2
НАДH
НАДФH
Триацилглицериды
НАДФH
НАДH
α-Глицерофосфат
Аспартат
Глутамат
НАДH
Цитруллин
Аргинин
Орнитин
CO2
NH3
Мочевина
Аланин
Серин
Глицин
Взаимосвязи липидного, углеводного и азотистого обмена
- Общее энергообеспечение (АТФ, НАД(Ф)H)
-
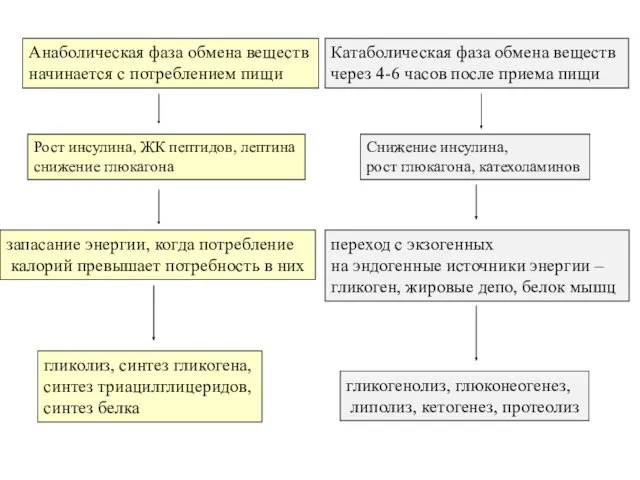
Анаболическая фаза обмена веществ
начинается с потреблением пищи
Катаболическая фаза обмена веществ
через 4-6
Анаболическая фаза обмена веществ
начинается с потреблением пищи
Катаболическая фаза обмена веществ
через 4-6

Роль печени в регуляции обмена веществ.
Роль печени в регуляции обмена веществ.
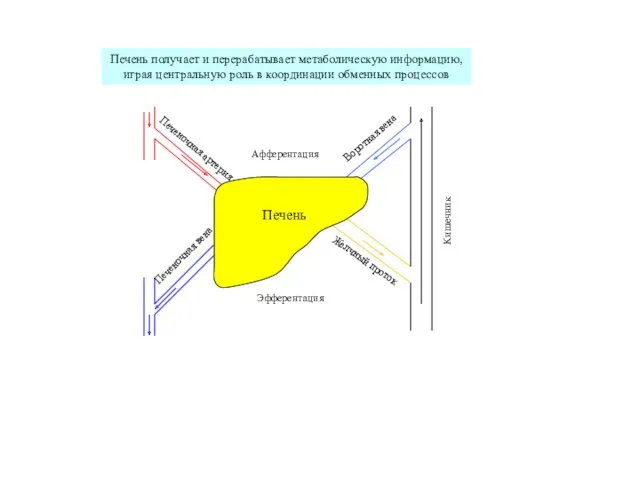
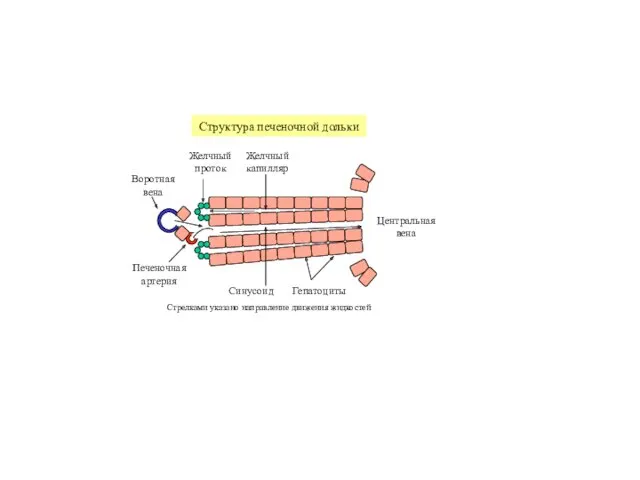

Печень выполняет важнейшую роль в адаптации обменных процессов к режиму питания,
Печень выполняет важнейшую роль в адаптации обменных процессов к режиму питания,
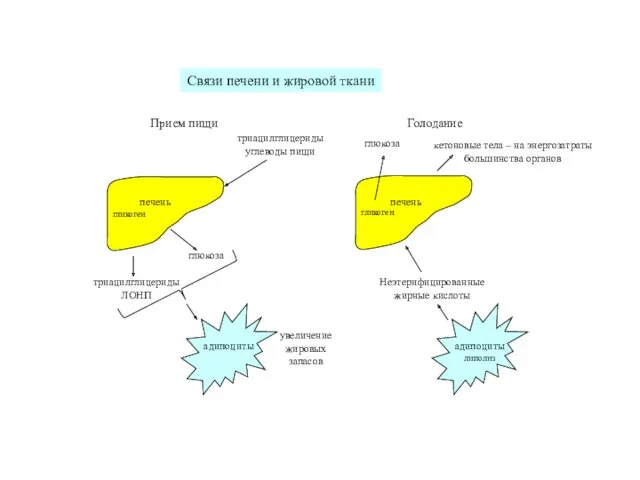
Связи печени и жировой ткани
Связи печени и жировой ткани
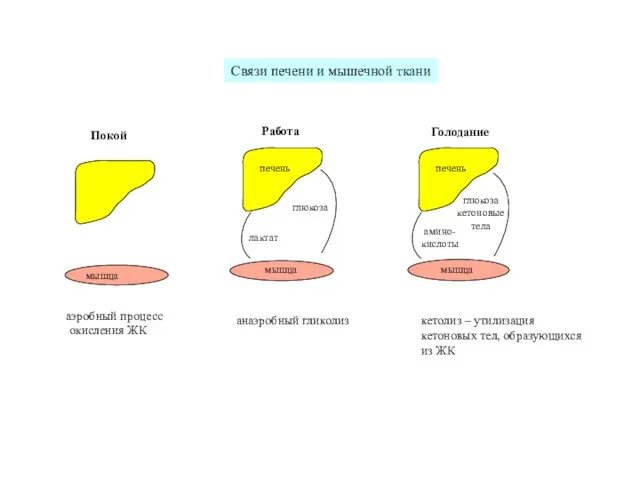
аэробный процесс
окисления ЖК
анаэробный гликолиз
кетолиз – утилизация
кетоновых тел, образующихся
из ЖК
аэробный процесс
окисления ЖК
анаэробный гликолиз
кетолиз – утилизация
кетоновых тел, образующихся
из ЖК
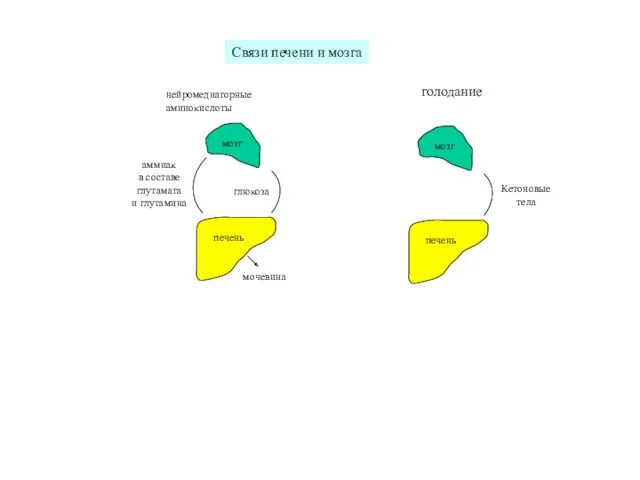
печень
мозг
глюкоза
аммиак
в составе
глутамата
и глутамина
мочевина
Связи печени и мозга
голодание
нейромедиаторные
аминокислоты
печень
мозг
глюкоза
аммиак
в составе
глутамата
и глутамина
мочевина
Связи печени и мозга
голодание
нейромедиаторные
аминокислоты
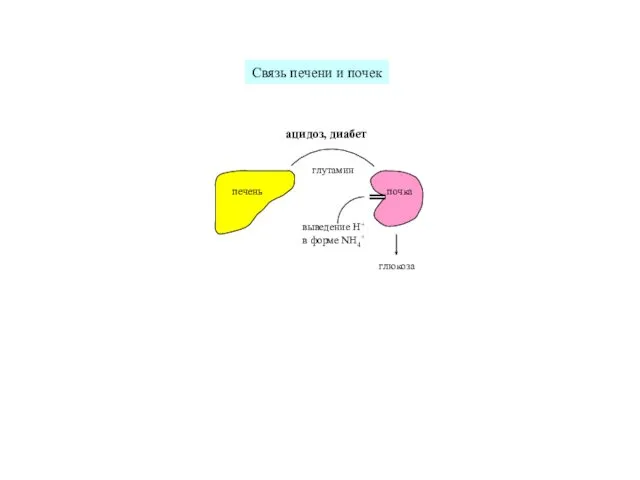
печень
почка
глутамин
глюкоза
выведение H+
в форме NH4+
ацидоз, диабет
Связь печени и почек
печень
почка
глутамин
глюкоза
выведение H+
в форме NH4+
ацидоз, диабет
Связь печени и почек

Направленность и интенсивность обменных процессов зависят от:
доступности субстратов и энергии,
активности ферментов.
Регуляция
Направленность и интенсивность обменных процессов зависят от:
доступности субстратов и энергии,
активности ферментов.
Регуляция
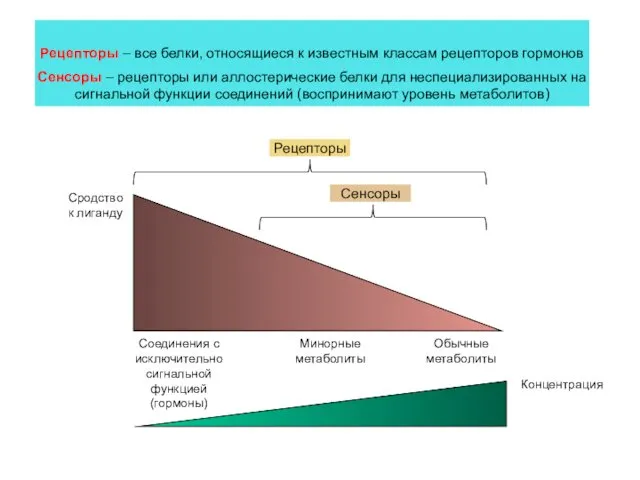
Сродство к лиганду
Соединения с исключительно сигнальной функцией (гормоны)
Обычные метаболиты
Минорные метаболиты
Рецепторы
Сенсоры
Рецепторы –
Сродство к лиганду
Соединения с исключительно сигнальной функцией (гормоны)
Обычные метаболиты
Минорные метаболиты
Рецепторы
Сенсоры
Рецепторы –
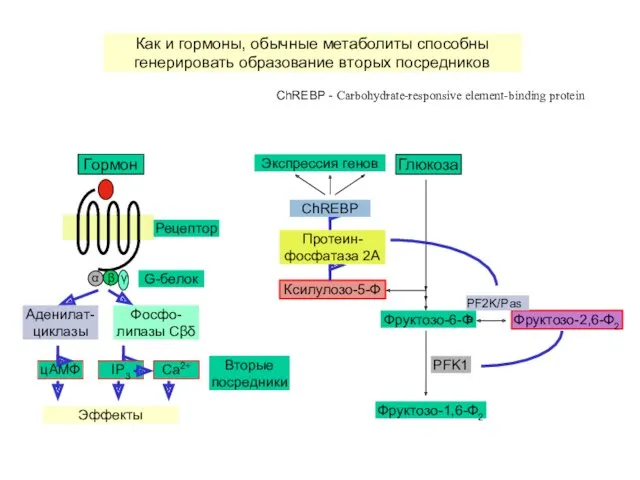
Аденилат-циклазы
цАМФ
Фосфо-липазы Cβδ
IP3
Ca2+
Эффекты
Гормон
G-белок
Рецептор
Вторые посредники
Глюкоза
Фруктозо-6-Ф
Фруктозо-1,6-Ф2
PFK1
Фруктозо-2,6-Ф2
PF2K/Pase
Ксилулозо-5-Ф
Протеин-фосфатаза 2A
ChREBP
Экспрессия генов
Как и гормоны, обычные метаболиты способны генерировать
Аденилат-циклазы
цАМФ
Фосфо-липазы Cβδ
IP3
Ca2+
Эффекты
Гормон
G-белок
Рецептор
Вторые посредники
Глюкоза
Фруктозо-6-Ф
Фруктозо-1,6-Ф2
PFK1
Фруктозо-2,6-Ф2
PF2K/Pase
Ксилулозо-5-Ф
Протеин-фосфатаза 2A
ChREBP
Экспрессия генов
Как и гормоны, обычные метаболиты способны генерировать
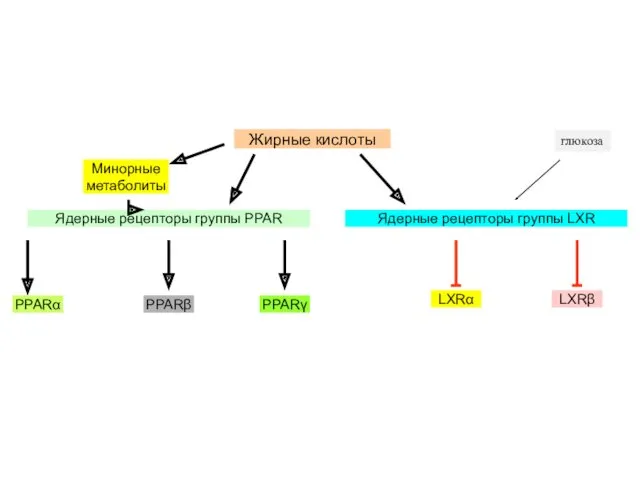
глюкоза
глюкоза
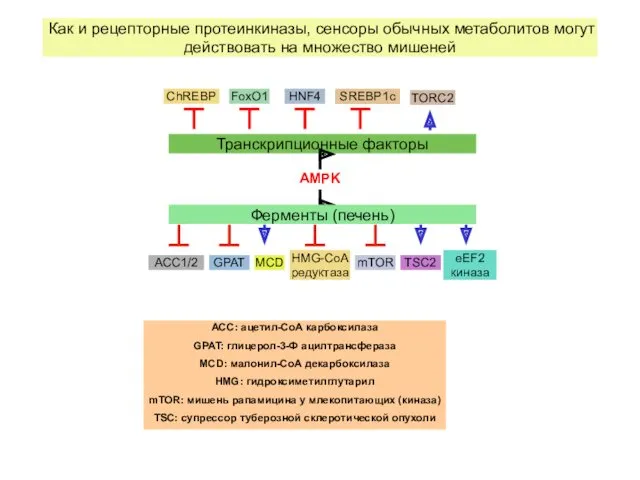
Как и рецепторные протеинкиназы, сенсоры обычных метаболитов могут действовать на
Как и рецепторные протеинкиназы, сенсоры обычных метаболитов могут действовать на

Азотистый обмен
Азотистый обмен

Азотистый баланс – отношение
потребленного азота к экскретируемому,-
важный показатель ростовых и
Азотистый баланс – отношение
потребленного азота к экскретируемому,-
важный показатель ростовых и
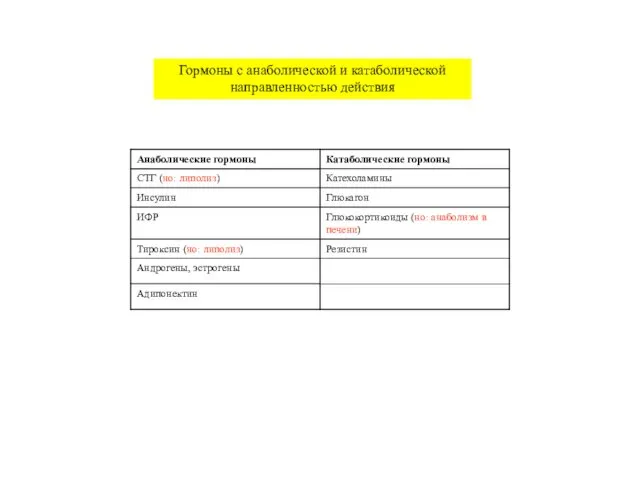
Гормоны с анаболической и катаболической направленностью действия
Гормоны с анаболической и катаболической направленностью действия
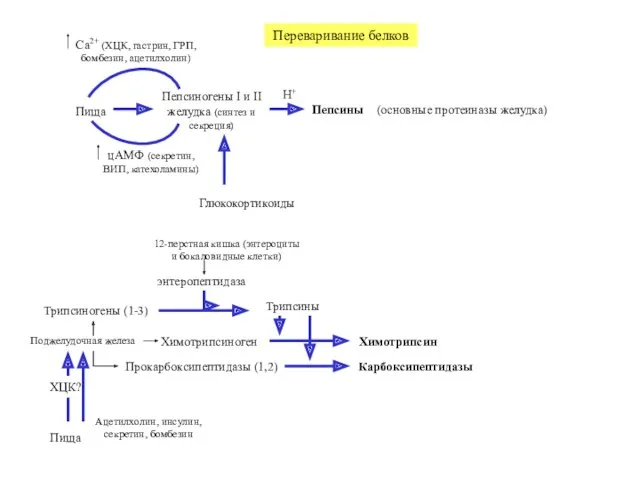
Переваривание белков
Пища
Пепсиногены I и II
желудка (синтез и секреция)
Пепсины
цАМФ (секретин, ВИП,
Переваривание белков
Пища
Пепсиногены I и II
желудка (синтез и секреция)
Пепсины
цАМФ (секретин, ВИП,
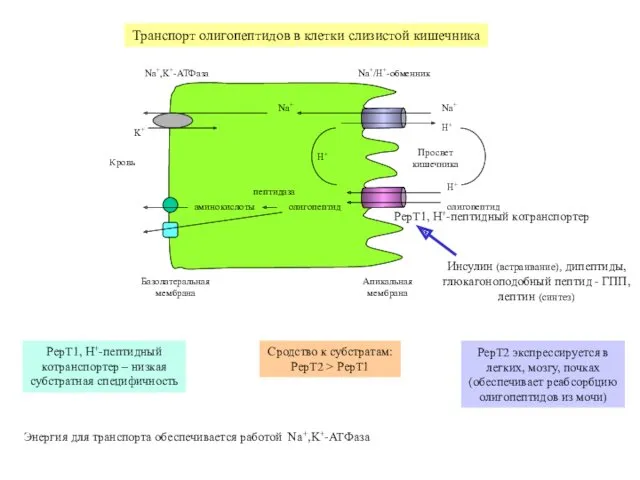
Na+
H+
H+
олигопептид
Na+
K+
Na+,K+-АТФаза
H+
Na+/H+-обменник
PepT1, H+-пептидный котранспортер
олигопептид
аминокислоты
пептидаза
Базолатеральная
мембрана
Апикальная
мембрана
Кровь
Просвет
кишечника
Транспорт олигопептидов в клетки слизистой кишечника
Инсулин (встраивание),
Na+
H+
H+
олигопептид
Na+
K+
Na+,K+-АТФаза
H+
Na+/H+-обменник
PepT1, H+-пептидный котранспортер
олигопептид
аминокислоты
пептидаза
Базолатеральная
мембрана
Апикальная
мембрана
Кровь
Просвет
кишечника
Транспорт олигопептидов в клетки слизистой кишечника
Инсулин (встраивание),

Системы транспорта аминокислот в кишечнике, почках, синапсах, других тканях
Другие субстраты: тиреоидные
Системы транспорта аминокислот в кишечнике, почках, синапсах, других тканях
Другие субстраты: тиреоидные
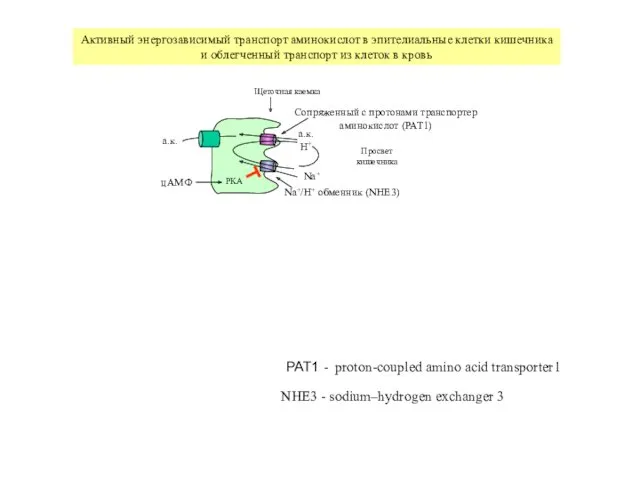
Щеточная каемка
Просвет
кишечника
Na+/H+ обменник (NHE3)
Сопряженный с протонами транспортер
аминокислот (PAT1)
а.к.
H+
Na+
PKA
цАМФ
Активный энергозависимый транспорт
Щеточная каемка
Просвет
кишечника
Na+/H+ обменник (NHE3)
Сопряженный с протонами транспортер
аминокислот (PAT1)
а.к.
H+
Na+
PKA
цАМФ
Активный энергозависимый транспорт
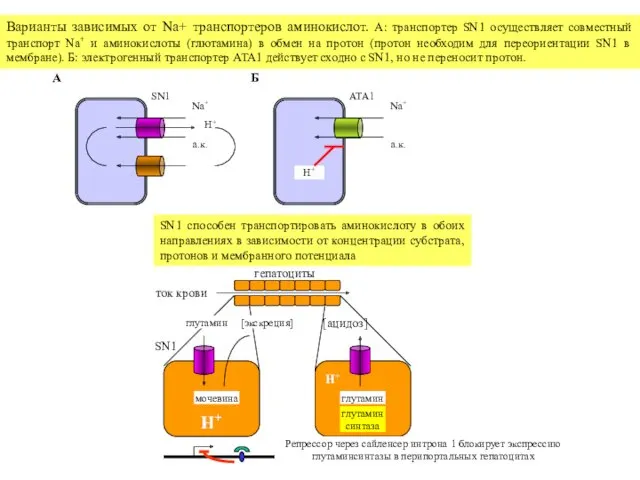
SN1
Na+
H+
а.к.
A
ATA1
Na+
H+
а.к.
Б
Варианты зависимых от Na+ транспортеров аминокислот. А: транспортер SN1 осуществляет совместный
SN1
Na+
H+
а.к.
A
ATA1
Na+
H+
а.к.
Б
Варианты зависимых от Na+ транспортеров аминокислот. А: транспортер SN1 осуществляет совместный
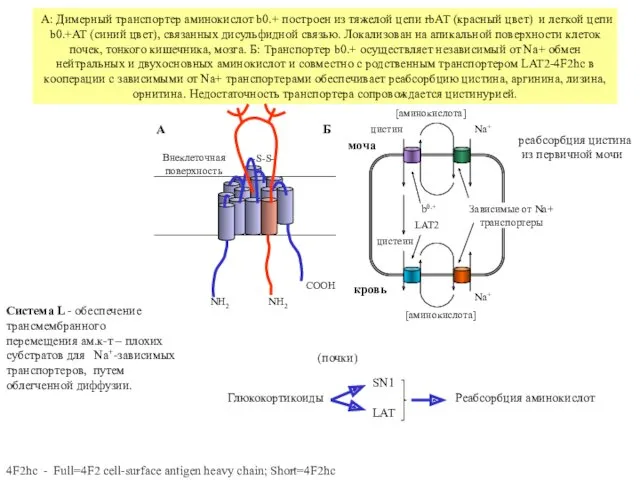
А: Димерный транспортер аминокислот b0.+ построен из тяжелой цепи rbAT (красный
А: Димерный транспортер аминокислот b0.+ построен из тяжелой цепи rbAT (красный

Межуточный обмен,
образование конечных продуктов и их выведение
Межуточный обмен,
образование конечных продуктов и их выведение

Строительный материал для белков
и азотсодержащих соединений;
источник энергии
Строительный материал для белков
и азотсодержащих соединений;
источник энергии
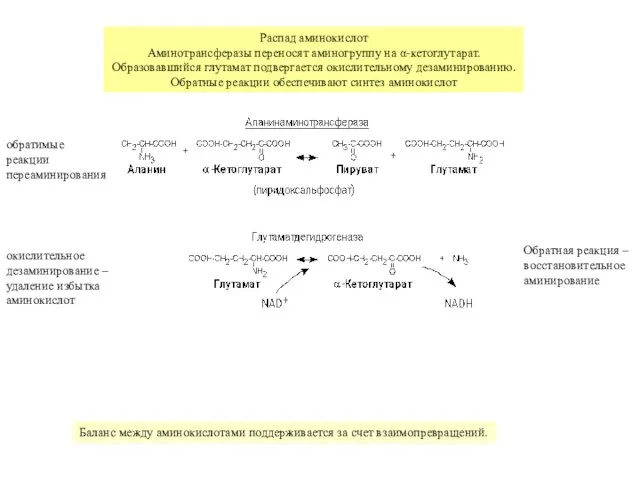
обратимые
реакции
переаминирования
окислительное
дезаминирование –
удаление избытка
аминокислот
Баланс между аминокислотами поддерживается за счет взаимопревращений.
Обратная реакция –
обратимые
реакции
переаминирования
окислительное
дезаминирование –
удаление избытка
аминокислот
Баланс между аминокислотами поддерживается за счет взаимопревращений.
Обратная реакция –
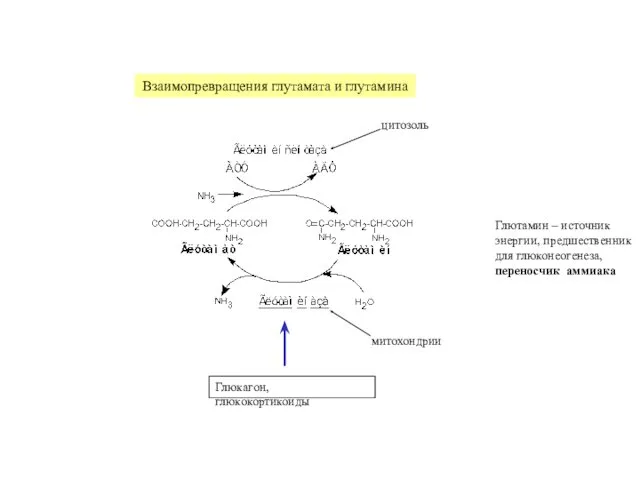
Взаимопревращения глутамата и глутамина
цитозоль
митохондрии
Глюкагон, глюкокортикоиды
Глютамин – источник
энергии, предшественник
для глюконеогенеза,
переносчик аммиака
Взаимопревращения глутамата и глутамина
цитозоль
митохондрии
Глюкагон, глюкокортикоиды
Глютамин – источник
энергии, предшественник
для глюконеогенеза,
переносчик аммиака
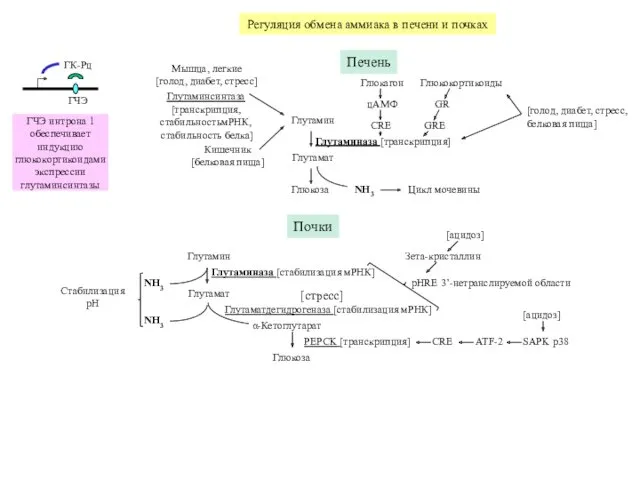
Регуляция обмена аммиака в печени и почках
ГЧЭ
ГК-Рц
ГЧЭ интрона 1 обеспечивает индукцию
Регуляция обмена аммиака в печени и почках
ГЧЭ
ГК-Рц
ГЧЭ интрона 1 обеспечивает индукцию
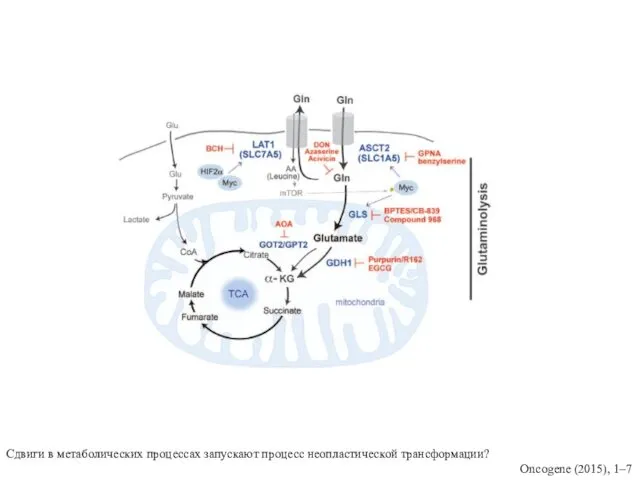
Сдвиги в метаболических процессах запускают процесс неопластической трансформации?
Oncogene (2015), 1–7
Сдвиги в метаболических процессах запускают процесс неопластической трансформации?
Oncogene (2015), 1–7
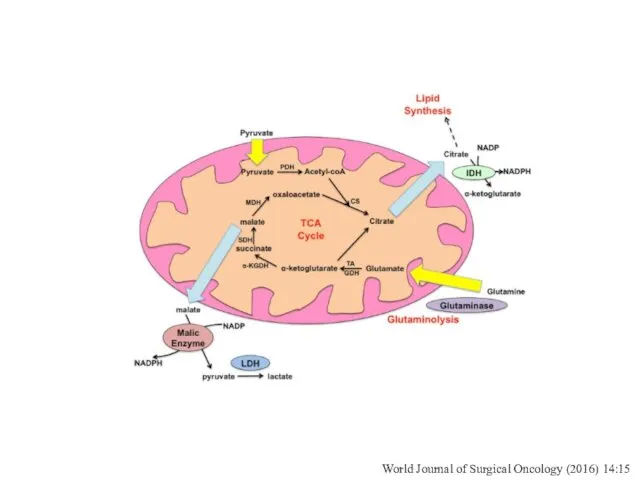
World Journal of Surgical Oncology (2016) 14:15
World Journal of Surgical Oncology (2016) 14:15
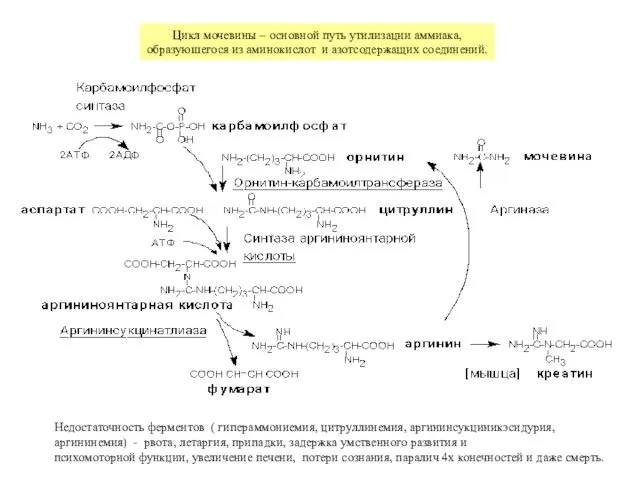
Недостаточность ферментов ( гипераммониемия, цитруллинемия, аргининсукциникэсидурия,
аргининемия) - рвота, летаргия, припадки, задержка
Недостаточность ферментов ( гипераммониемия, цитруллинемия, аргининсукциникэсидурия,
аргининемия) - рвота, летаргия, припадки, задержка
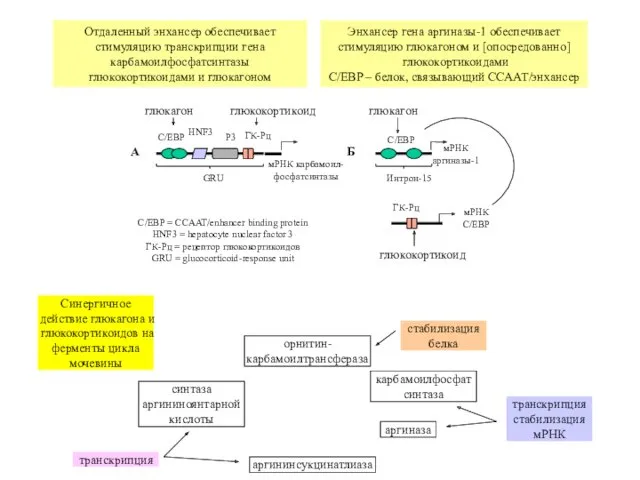
Отдаленный энхансер обеспечивает стимуляцию транскрипции гена карбамоилфосфатсинтазы
глюкокортикоидами и глюкагоном
Энхансер гена
Отдаленный энхансер обеспечивает стимуляцию транскрипции гена карбамоилфосфатсинтазы
глюкокортикоидами и глюкагоном
Энхансер гена
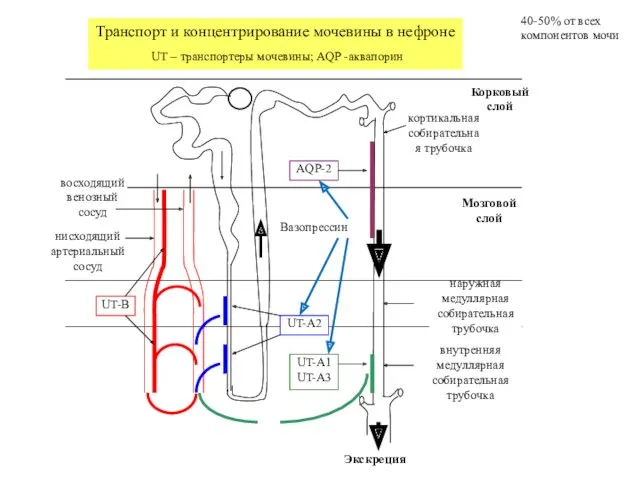
Корковый слой
кортикальная собирательная трубочка
наружная медуллярная собирательная трубочка
внутренняя медуллярная собирательная трубочка
Корковый слой
кортикальная собирательная трубочка
наружная медуллярная собирательная трубочка
внутренняя медуллярная собирательная трубочка

Регуляция циркадных ритмов и метаболизма с участием ядерного
рецептора Rev-erbα
Регуляция циркадных ритмов и метаболизма с участием ядерного
рецептора Rev-erbα
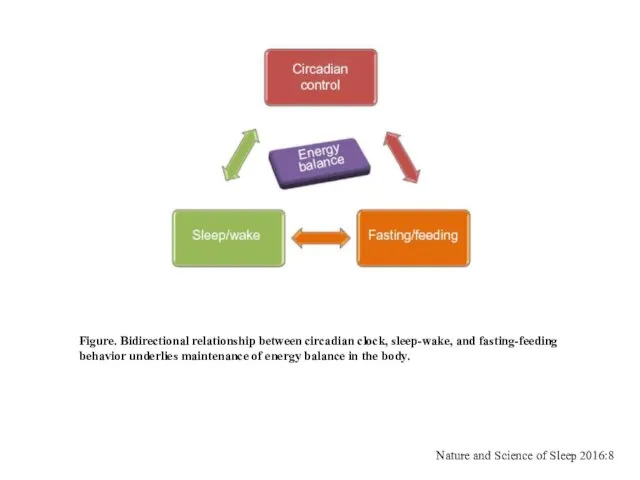
Figure. Bidirectional relationship between circadian clock, sleep-wake, and fasting-feeding
behavior underlies
Figure. Bidirectional relationship between circadian clock, sleep-wake, and fasting-feeding
behavior underlies

Сигналы света с сетчатки через ретиногипоталамический тракт попадают в
супрахиазматическое ядро мозга
Сигналы света с сетчатки через ретиногипоталамический тракт попадают в
супрахиазматическое ядро мозга
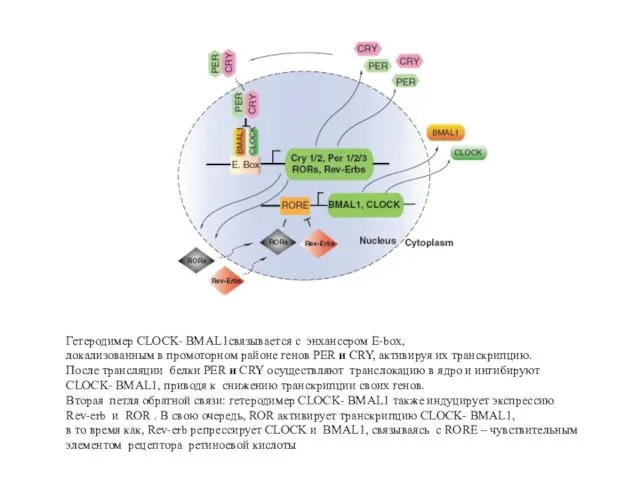
Гетеродимер CLOCK- BMAL1связывается с энхансером E-box,
локализованным в промоторном районе генов PER
Гетеродимер CLOCK- BMAL1связывается с энхансером E-box,
локализованным в промоторном районе генов PER
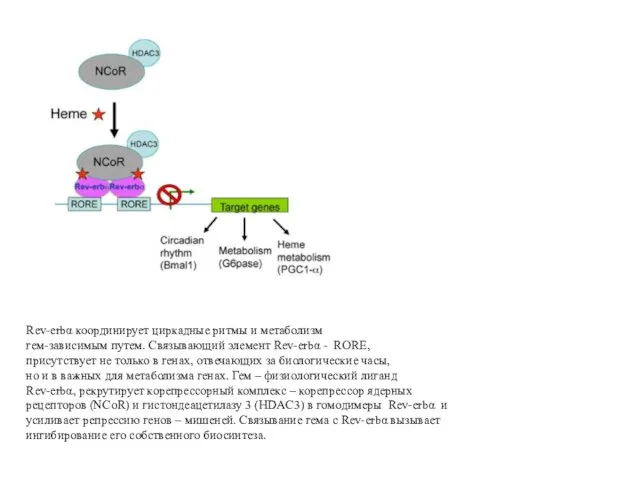
Rev-erbα координирует циркадные ритмы и метаболизм
гем-зависимым путем. Связывающий элемент Rev-erbα -
Rev-erbα координирует циркадные ритмы и метаболизм
гем-зависимым путем. Связывающий элемент Rev-erbα -
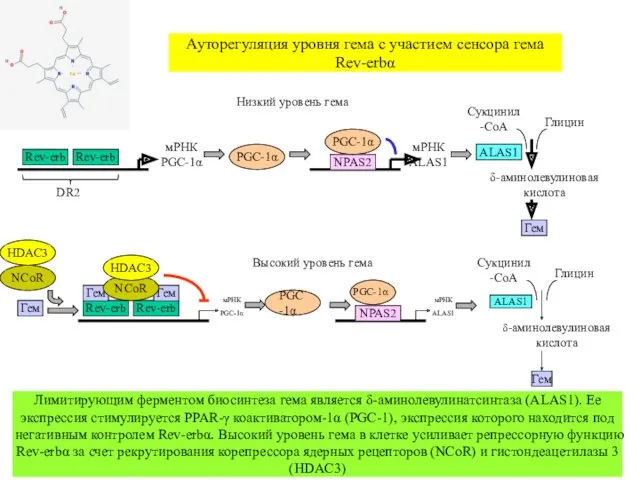
Ауторегуляция уровня гема с участием сенсора гема Rev-erbα
мРНК PGC-1α
Гем
Rev-erb
Rev-erb
Гем
Гем
мРНК PGC-1α
Ауторегуляция уровня гема с участием сенсора гема Rev-erbα
мРНК PGC-1α
Гем
Rev-erb
Rev-erb
Гем
Гем
мРНК PGC-1α
 Основные понятия и показатели надежности
Основные понятия и показатели надежности Конспект открытого интегрированного занятия в подготовительной группе Путешествие в школу. Диск
Конспект открытого интегрированного занятия в подготовительной группе Путешествие в школу. Диск Литература 07.02.24
Литература 07.02.24 Презентация к уроку географии 11 класс Зарубежная Азия.Зачет
Презентация к уроку географии 11 класс Зарубежная Азия.Зачет Татар теле дәресе 4 сыйныф
Татар теле дәресе 4 сыйныф Русская литература XVIII века
Русская литература XVIII века Обобщённая структурная схема радиотехнической системы передачи информации
Обобщённая структурная схема радиотехнической системы передачи информации Семья
Семья План и поисковые маркетинговые исследования: вторичная информация
План и поисковые маркетинговые исследования: вторичная информация Урок географии в 8-9 классах Формирование территории России
Урок географии в 8-9 классах Формирование территории России Изменение политической карты мира. Географические открытия. Конец XV – середина XX вв
Изменение политической карты мира. Географические открытия. Конец XV – середина XX вв Системы оповещения и управления эвакуацией при пожаре (далее СОУЭ)
Системы оповещения и управления эвакуацией при пожаре (далее СОУЭ) Магнітний запис інформації
Магнітний запис інформації Режим дня школьника
Режим дня школьника Шаблоны презентаций по химии
Шаблоны презентаций по химии презентация Найди отличия
презентация Найди отличия Исследовательский проект - лекарственные растения
Исследовательский проект - лекарственные растения Становление и развитие дошкольной педагогики
Становление и развитие дошкольной педагогики Уборочные машины и технологии
Уборочные машины и технологии Мейоз. Гаметогенез
Мейоз. Гаметогенез Иерусалим царя Давида и Соломона. Лекция 2
Иерусалим царя Давида и Соломона. Лекция 2 Гидродинамические исследования скважин
Гидродинамические исследования скважин Славное море - священный Байкал
Славное море - священный Байкал презентация Углерод
презентация Углерод Классный час,посвящённый Дню Победы. Диск Диск Диск
Классный час,посвящённый Дню Победы. Диск Диск Диск История, политическая программа, статистика
История, политическая программа, статистика Презентация Кто есть кто
Презентация Кто есть кто Требования, предъявляемые к строительным конструкциям
Требования, предъявляемые к строительным конструкциям