Последовательность событий в химическом синапсе после активации пресинаптического окончания презентация
- Последовательность событий в химическом синапсе после активации пресинаптического окончания

Содержание
- 2. Стадии экзоцитоза медиатора из синаптических везикул Транспорт (мобилизация, англ., trafficking) вновь синтезированных везикул из перикариона в
- 3. Функциональные субпопуляции (пулы) синаптических везикул Three vesicle pools a) The classic three-pool model. The reserve pool
- 5. Стадии экзоцитоза медиатора из синаптических везикул Специальный процесс контроля (англ., targeting) с участием малых G-белков rab-семейства,
- 6. Стадии экзоцитоза медиатора из синаптических везикул Стыковка (англ., docking) везикулы с местом освобождения медиатора в активной
- 7. Стадии экзоцитоза медиатора из синаптических везикул Слияние (англ., fusion) мембран является определяющей фазой регулируемого экзоцитоза. При
- 8. Альтернативный способ выделения медиатора из синаптических везикул Существует альтернативная форма тесного слияния везикулы с мембраной клетки
- 9. Стадии экзоцитоза медиатора из синаптических везикул Восстановление (англ., retrieval), или так называемое рециклирование (англ., recycling) везикул
- 10. Морфо-функциональная организация активных зон Методом электронно-микроскопичес-кой томографии на мембранах активных зон выявлена электронно-плотная упорядоченная решетка цитоматрикса
- 11. Морфо-функциональная организация активных зон Плазматическая мембрана активных зон включает: ионные каналы, рецепторные белки, специальные клеточные молекулы,
- 12. Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс экзоцитоза
- 13. Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс экзоцитоза
- 14. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза Протонный насос Н+-АТФаза V-типа переносит внутрь везикул
- 15. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза Синаптические SNARE белки и SNARE комплексы (англ.,
- 16. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза SNARE комплексы образованы тремя белками: везикулярный белок
- 17. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза Синаптофизин связывается с холестеролом и вовлечен в
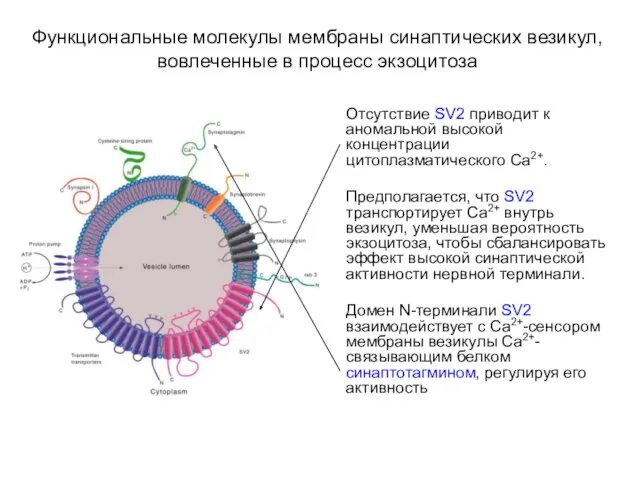
- 18. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза Отсутствие SV2 приводит к аномальной высокой концентрации
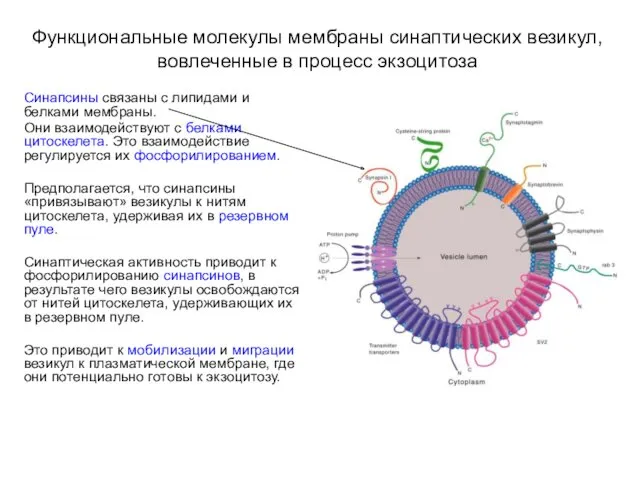
- 19. Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза Синапсины связаны с липидами и белками мембраны.
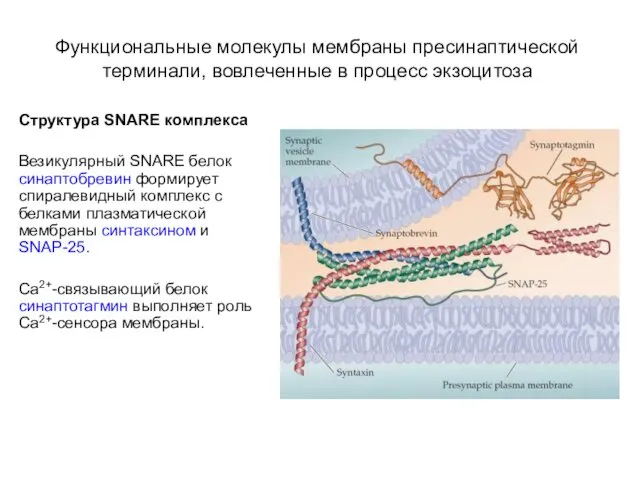
- 20. Функциональные молекулы мембраны пресинаптической терминали, вовлеченные в процесс экзоцитоза Структура SNARE комплекса Везикулярный SNARE белок синаптобревин
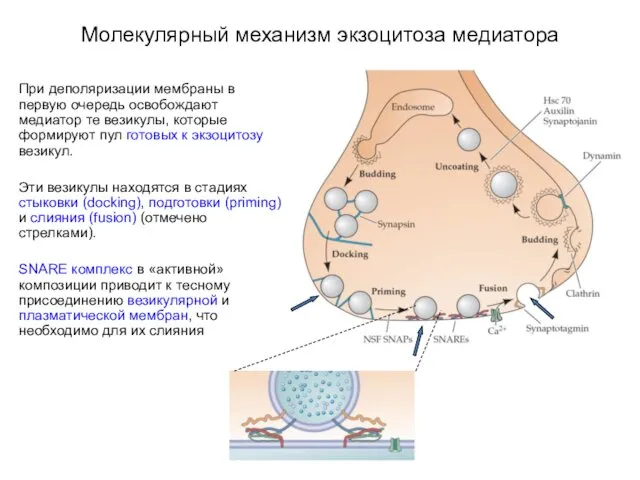
- 21. Молекулярный механизм экзоцитоза медиатора При деполяризации мембраны в первую очередь освобождают медиатор те везикулы, которые формируют
- 22. Молекулярный механизм экзоцитоза медиатора SNARE комплекс в «активной» композиции приводит к тесному присоединению везикулярной и плазматической
- 23. Молекулярный механизм экзоцитоза медиатора Синаптотагмин является ключевым Са2+-сенсором и связывает Са2+, что необходимо для запуска слияния
- 24. Молекулярный механизм экзоцитоза медиатора Клатрин захватывает «пустую» везикулу, «отпочковывая» ее от плазматической мембраны. Молекулы клатрина формируют
- 25. Молекулярный механизм экзоцитоза медиатора (1) Другой белок динамин сдавливает края мембранной полусферы, замыкая ее в «одетый»
- 26. Молекулярный механизм экзоцитоза медиатора (1) Вновь образованные везикулы могут сливаться с внутриклеточными эндосомами по механизму экзоцитоза.
- 27. Synaptic vesicles partially or completely fuse with the presynaptic membrane at the active zone and release
- 28. Альтернативный способ выделения медиатора из синаптических везикул При частичном выделении медиатора способом «поцеловал и убежал» не
- 29. Альтернативный способ выделения медиатора из синаптических везикул Динамика тока, отражающего сложную кинетику высвобождения катехоламина из электронноплотной
- 30. Разные формы экзоцитоза медиатора
- 31. Несинаптическое выделение медиаторов по механизму экзоцитоза Медиаторы могут выделяться из клеток и за пределами синаптической щели.
- 32. Несинаптическое выделение медиаторов по механизму экзоцитоза 2) Вторая модель постулирует, что кольцо из белкового комплекса сначала
- 33. Несинаптическая диффузная передача В отличие от синаптической, несинаптическая диффузная передача в большей степени зависит от скорости
- 35. Скачать презентацию

Стадии экзоцитоза медиатора из синаптических везикул
Транспорт (мобилизация, англ., trafficking) вновь синтезированных
Стадии экзоцитоза медиатора из синаптических везикул
Транспорт (мобилизация, англ., trafficking) вновь синтезированных
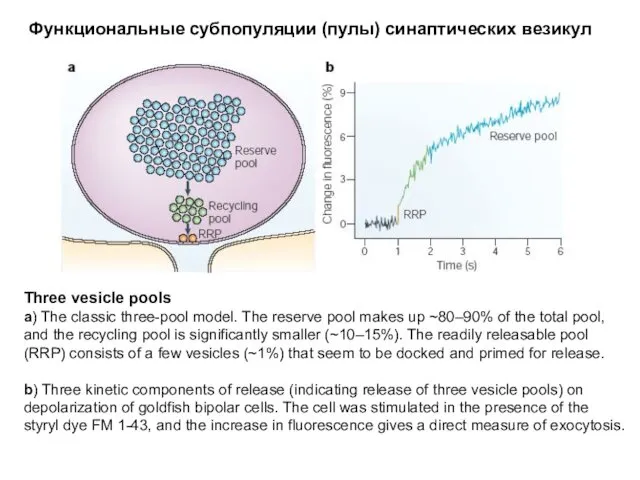
Функциональные субпопуляции (пулы) синаптических везикул
Three vesicle pools
a) The classic three-pool model.
Функциональные субпопуляции (пулы) синаптических везикул
Three vesicle pools
a) The classic three-pool model.


Стадии экзоцитоза медиатора из синаптических везикул
Специальный процесс контроля (англ., targeting) с
Стадии экзоцитоза медиатора из синаптических везикул
Специальный процесс контроля (англ., targeting) с

Стадии экзоцитоза медиатора из синаптических везикул
Стыковка (англ., docking) везикулы с местом
Стадии экзоцитоза медиатора из синаптических везикул
Стыковка (англ., docking) везикулы с местом

Стадии экзоцитоза медиатора из синаптических везикул
Слияние (англ., fusion) мембран является определяющей
Стадии экзоцитоза медиатора из синаптических везикул
Слияние (англ., fusion) мембран является определяющей

Альтернативный способ выделения медиатора из синаптических везикул
Существует альтернативная форма тесного слияния
Альтернативный способ выделения медиатора из синаптических везикул
Существует альтернативная форма тесного слияния

Стадии экзоцитоза медиатора из синаптических везикул
Восстановление (англ., retrieval), или так называемое
Стадии экзоцитоза медиатора из синаптических везикул
Восстановление (англ., retrieval), или так называемое
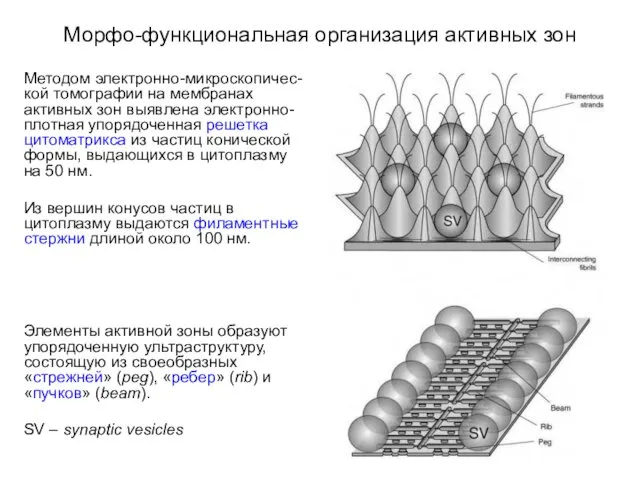
Морфо-функциональная организация активных зон
Методом электронно-микроскопичес-кой томографии на мембранах активных зон выявлена
Морфо-функциональная организация активных зон
Методом электронно-микроскопичес-кой томографии на мембранах активных зон выявлена

Морфо-функциональная организация активных зон
Плазматическая мембрана активных зон включает:
ионные каналы,
рецепторные
Морфо-функциональная организация активных зон
Плазматическая мембрана активных зон включает:
ионные каналы,
рецепторные
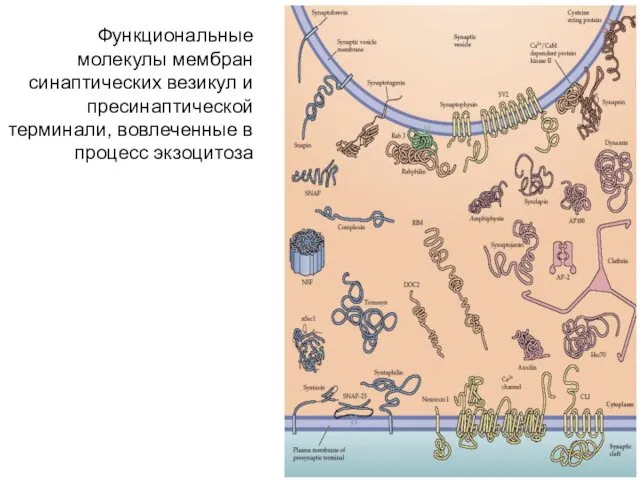
Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс
Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс
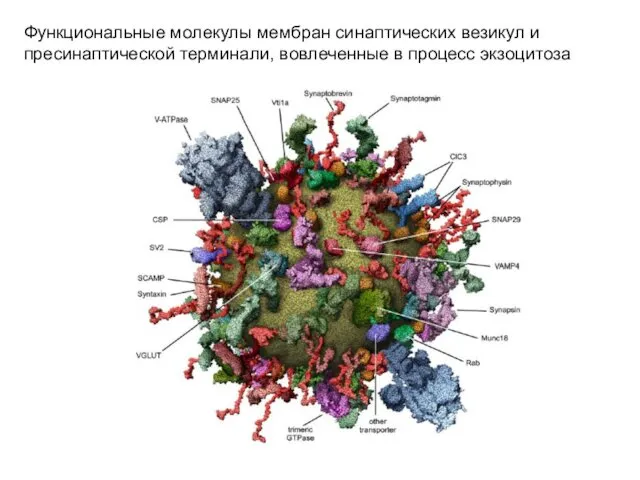
Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс
Функциональные молекулы мембран синаптических везикул и пресинаптической терминали, вовлеченные в процесс
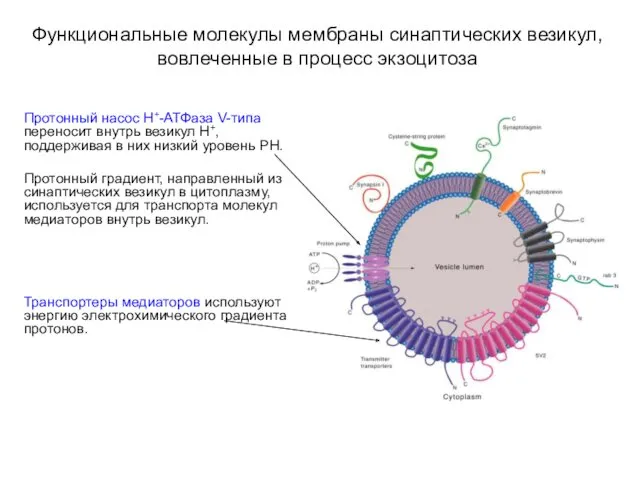
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Протонный насос Н+-АТФаза
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Протонный насос Н+-АТФаза
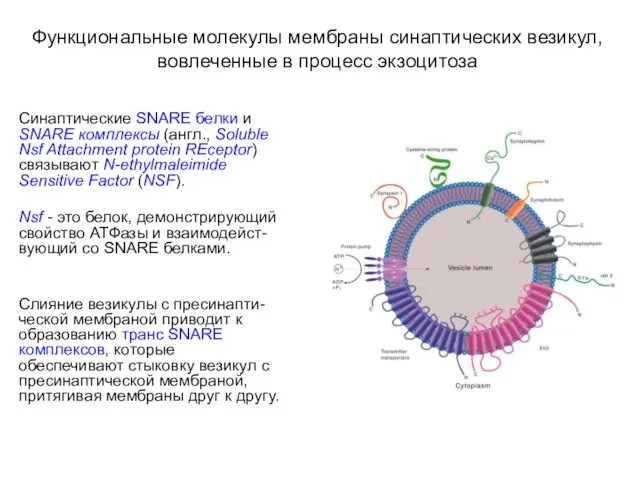
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синаптические SNARE белки
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синаптические SNARE белки
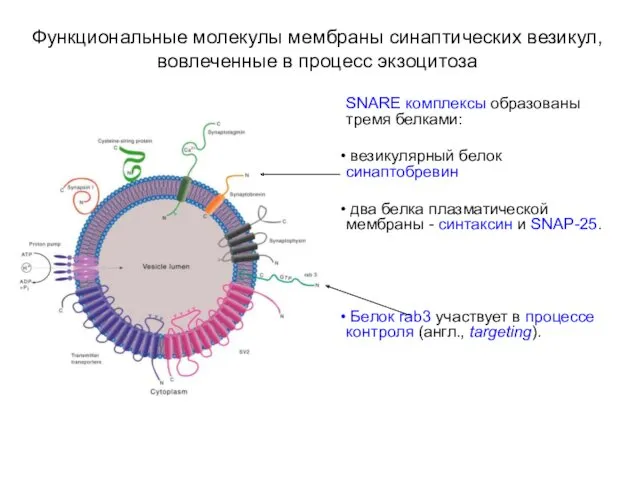
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
SNARE комплексы образованы
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
SNARE комплексы образованы
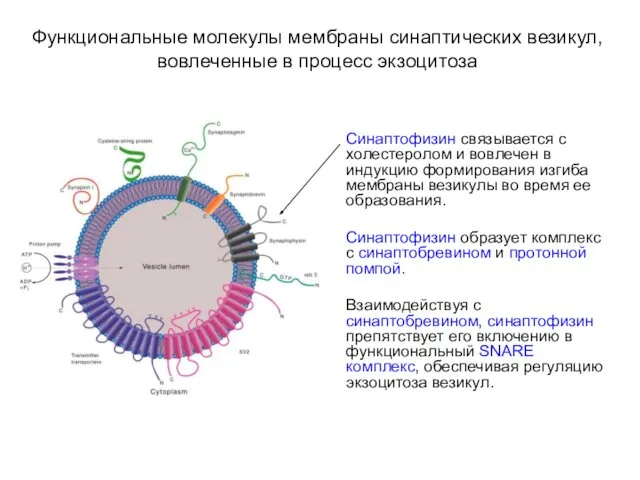
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синаптофизин связывается с
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синаптофизин связывается с
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Отсутствие SV2 приводит
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Отсутствие SV2 приводит
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синапсины связаны с
Функциональные молекулы мембраны синаптических везикул, вовлеченные в процесс экзоцитоза
Синапсины связаны с
Функциональные молекулы мембраны пресинаптической терминали, вовлеченные в процесс экзоцитоза
Структура SNARE комплекса
Функциональные молекулы мембраны пресинаптической терминали, вовлеченные в процесс экзоцитоза
Структура SNARE комплекса
Молекулярный механизм экзоцитоза медиатора
При деполяризации мембраны в первую очередь освобождают медиатор
Молекулярный механизм экзоцитоза медиатора
При деполяризации мембраны в первую очередь освобождают медиатор
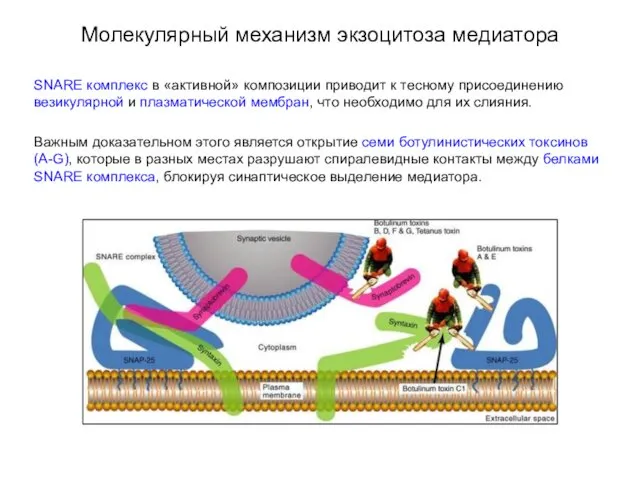
Молекулярный механизм экзоцитоза медиатора
SNARE комплекс в «активной» композиции приводит к тесному
Молекулярный механизм экзоцитоза медиатора
SNARE комплекс в «активной» композиции приводит к тесному
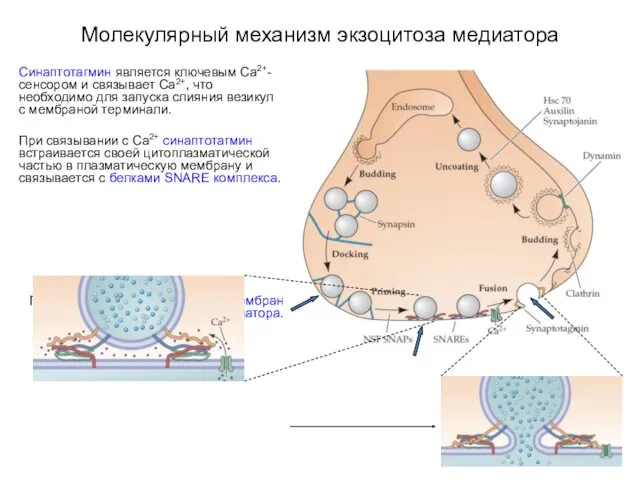
Молекулярный механизм экзоцитоза медиатора
Синаптотагмин является ключевым Са2+-сенсором и связывает Са2+, что
Молекулярный механизм экзоцитоза медиатора
Синаптотагмин является ключевым Са2+-сенсором и связывает Са2+, что
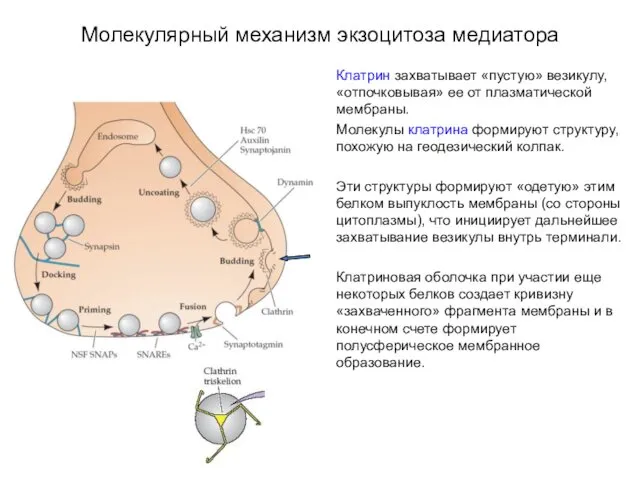
Молекулярный механизм экзоцитоза медиатора
Клатрин захватывает «пустую» везикулу, «отпочковывая» ее от плазматической
Молекулярный механизм экзоцитоза медиатора
Клатрин захватывает «пустую» везикулу, «отпочковывая» ее от плазматической
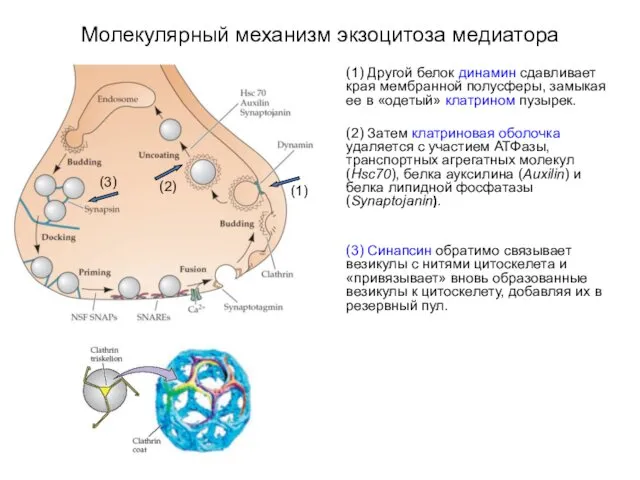
Молекулярный механизм экзоцитоза медиатора
(1) Другой белок динамин сдавливает края мембранной полусферы,
Молекулярный механизм экзоцитоза медиатора
(1) Другой белок динамин сдавливает края мембранной полусферы,
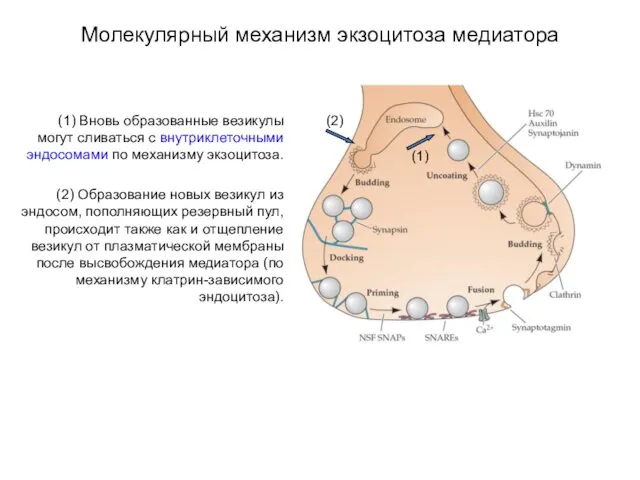
Молекулярный механизм экзоцитоза медиатора
(1) Вновь образованные везикулы могут сливаться с внутриклеточными
Молекулярный механизм экзоцитоза медиатора
(1) Вновь образованные везикулы могут сливаться с внутриклеточными
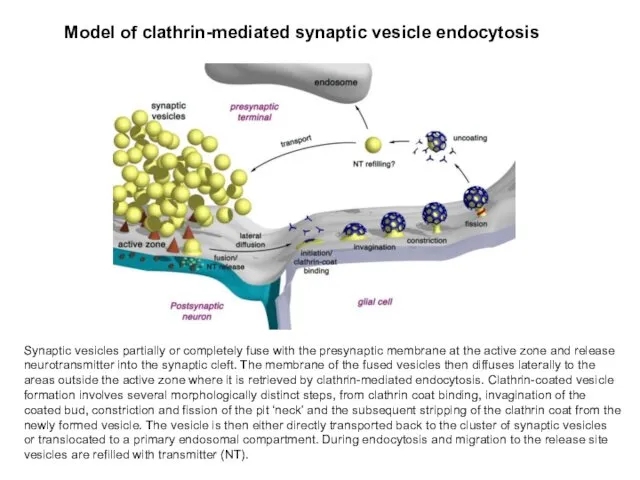
Synaptic vesicles partially or completely fuse with the presynaptic membrane at
Synaptic vesicles partially or completely fuse with the presynaptic membrane at
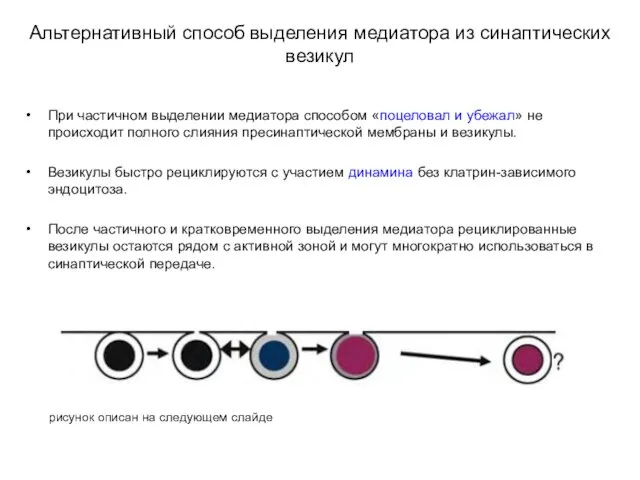
Альтернативный способ выделения медиатора из синаптических везикул
При частичном выделении медиатора способом
Альтернативный способ выделения медиатора из синаптических везикул
При частичном выделении медиатора способом
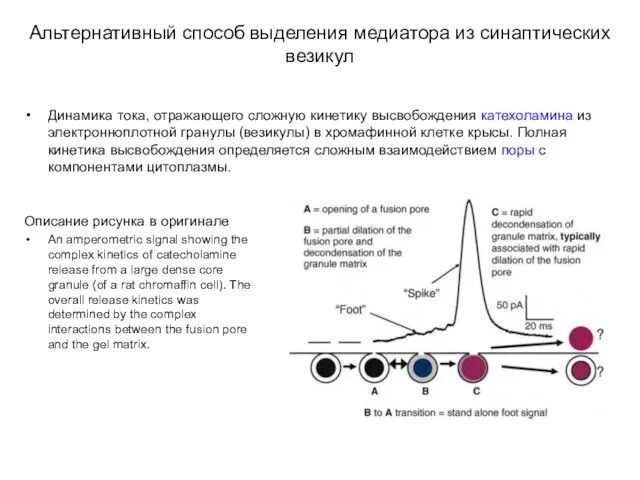
Альтернативный способ выделения медиатора из синаптических везикул
Динамика тока, отражающего сложную кинетику
Альтернативный способ выделения медиатора из синаптических везикул
Динамика тока, отражающего сложную кинетику
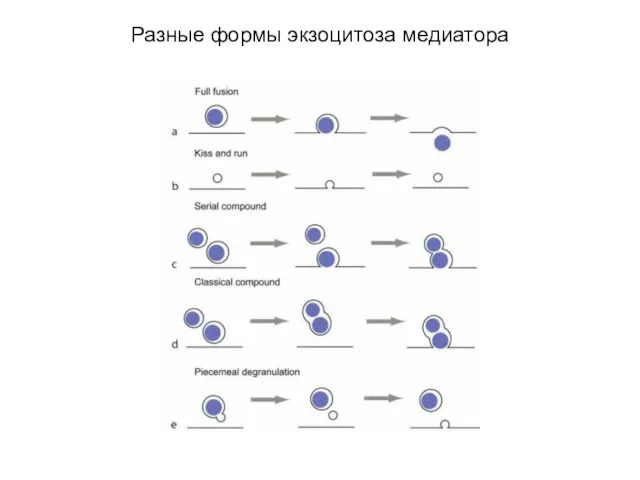
Разные формы экзоцитоза медиатора
Разные формы экзоцитоза медиатора
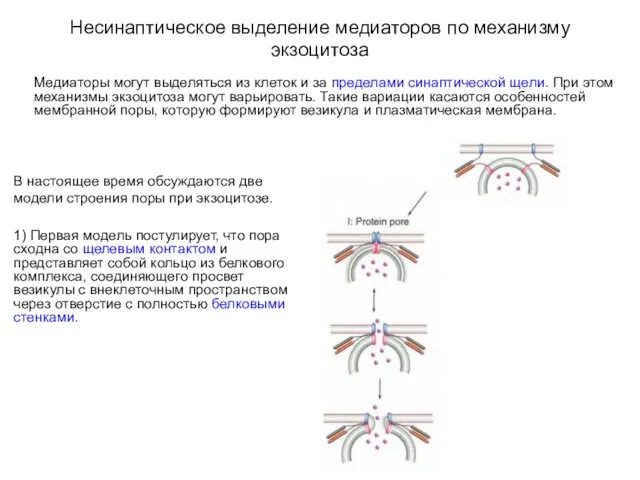
Несинаптическое выделение медиаторов по механизму экзоцитоза
Медиаторы могут выделяться из клеток и
Несинаптическое выделение медиаторов по механизму экзоцитоза
Медиаторы могут выделяться из клеток и

Несинаптическое выделение медиаторов по механизму экзоцитоза
2) Вторая модель постулирует, что кольцо
Несинаптическое выделение медиаторов по механизму экзоцитоза
2) Вторая модель постулирует, что кольцо
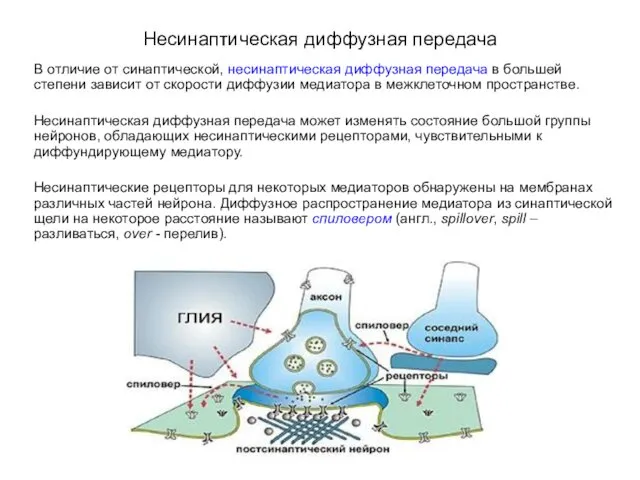
Несинаптическая диффузная передача
В отличие от синаптической, несинаптическая диффузная передача в большей
Несинаптическая диффузная передача
В отличие от синаптической, несинаптическая диффузная передача в большей
 Как хороша ты, родная весна!
Как хороша ты, родная весна! Водный транспорт. Морской и речной транспорт
Водный транспорт. Морской и речной транспорт Этапы вязания крючком
Этапы вязания крючком Компанія ЗМ
Компанія ЗМ Принципы имитационного моделирования. Математические методы моделирования
Принципы имитационного моделирования. Математические методы моделирования Словарные слова (1-2 класс)
Словарные слова (1-2 класс) Визволення України від нацистських загарбників
Визволення України від нацистських загарбників Компанія Federal. Розподільче обладнання
Компанія Federal. Розподільче обладнання Halloween. On the 31st of October British people celebrate Halloween
Halloween. On the 31st of October British people celebrate Halloween Единые сроки каникул в московских школах
Единые сроки каникул в московских школах Глобальные сети и перспективные сетевые технологии. Глобальные сети (Wide Area Networks, WAN)
Глобальные сети и перспективные сетевые технологии. Глобальные сети (Wide Area Networks, WAN) презентация 6 класс Горные породы и минералы
презентация 6 класс Горные породы и минералы Федеральные СМИ
Федеральные СМИ Leongardt_mikrofinansovye_organizatsii
Leongardt_mikrofinansovye_organizatsii Брюгге. Достопримечательности
Брюгге. Достопримечательности Бронхиальная астма
Бронхиальная астма Формирование у детей старшего дошкольного возраста положительной учебной мотивации
Формирование у детей старшего дошкольного возраста положительной учебной мотивации Проверочная работа по теме Мировой океан Диск
Проверочная работа по теме Мировой океан Диск История и методология науки
История и методология науки Презентация Металлические деньги в истории России
Презентация Металлические деньги в истории России Программная обработка данных на компьютере
Программная обработка данных на компьютере Праздник Троицы. Церковное песнопение,
Праздник Троицы. Церковное песнопение, Счетчики. Четырехразрядный суммирующий асинхронный двоичный счетчик
Счетчики. Четырехразрядный суммирующий асинхронный двоичный счетчик Электронный конструктор Знаток
Электронный конструктор Знаток Интегрированный урок по английскому языку и физической культуре. 3 класс
Интегрированный урок по английскому языку и физической культуре. 3 класс Энергетические характеристики агрегатов и ГЭС
Энергетические характеристики агрегатов и ГЭС Защита прав человека в Японии
Защита прав человека в Японии Лепка фигуры человека с каркасом
Лепка фигуры человека с каркасом