Повреждения при оксидативном стрессе, защитные механизмы, регуляторы стрессовой реакции, другие типы стрессов презентация
- Повреждения при оксидативном стрессе, защитные механизмы, регуляторы стрессовой реакции, другие типы стрессов

Содержание
- 2. АФК-сенсоры (рецепторы) в клетках растений: 1. Двухкомпонентные гистидин-киназы; 2. Редокс-чувствительные факторы транскрипции, такие как NPR1 или
- 3. Кальций-связывающие белки (CBPs) – 5 классов: - Кальмодулины - кальмодулин-подобные белки - Ca2+-зависимые протеин-киназы (CDPKs) -
- 4. Гидроперекисное окисление липидов (липиды – наиболее чувствительны к оксидативному стрессы, они способны к лавинообразному окислению и
- 5. Липидный радикал (L•). L• может активировать O2 и формировать липидный пероксильный радикал (LOO•), который способен отнимать
- 6. (3) Фаза терминации Липиды разлагаются во время стадии пропагации до терминальных низкомолекулярных продуктов (т.н. продуктов гидроперекисного
- 7. Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman, 1997; Shacter 2000; Stadtman and
- 8. Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman, 1997; Shacter 2000; Stadtman and
- 9. Основные окислительные повреждения биомолекул при стрессе: Оксидативное повреждение белков А - окисление серусодержащих аминокислот: цистеина, метионина
- 10. Б - Карбонилирование – формирование или встраивание свободной карбонильной группы (C=O) Вторая по встречаемости реакция оксидативного
- 11. Карбонилирование приводит к образованию реактивных кетонов и альдегидов, детектируемых тестом Бради с 2,4-динитрофенилгидразином (в тесте образуются
- 12. Two-dimensional profiles of protein abundance and oxidation in dry mature Arabidopsis seeds. Job C et al.
- 13. В – S-нитрозилирование. Недавно открытое повреждение серусодержащих аминокислот белков под действием нитрозативного стресса. Механизм: NO• реагирует
- 14. Оксидативное «повреждение» углеводов. Моно- и дисахариды – важные АФК-связывающие агенты, проявляющие антиоксидантные свойства. Это показывает, что
- 15. Оксидативное «повреждение» нуклеиновых кислот. Для большинства однолетних культурных растений проблема рака не актуальна. Тем не менее
- 16. Основные антиоксидантные системы растений (см. также предыдущие лекции). Ключевые ферментативные антиоксиданты (обычно отличающиеся высокой специфичностью к
- 17. Основные антиоксидантные системы растений (см. также предыдущие лекции). Ключевые неферментативные антиоксиданты (обычно отличающиеся невысокой специфичностью к
- 18. Вспомогательные системы антиоксидатной защиты растений. Служат для поддержания антиоксидантов в функциональном состоянии. Часто это восстановленное состояние.
- 19. Пероксидазы растений: Три «классических» типа: класс-I, класс-II and класс-III. Пероксиредоксины сейчас часто относят с четвертому классу
- 20. Пероксидазы растений: Недавно найдены гибридные пероксидазы первого класса, совмещающие структуру каталазо-пероксидазы и цитохром-c-пероксидазы. Аскорбат-пероксидаза (APX) –
- 21. Пероксидазы растений: Второй класс пероксидаз существует только у грибов – включает марганец-пероксидазы, лигнин-пероксидазы, и так-называемые версатильные
- 22. Общие защитные механизмы (против практически любого стрессового повреждения): усиление катаболических процессов для высвобождения энергии активация ионных
- 23. Способность культурных растений выдерживать засоление
- 24. Способы защиты растений от избытка солей Галофиты: — растения, способные жить при высоких уровнях засоления (солянки,
- 25. Механизм поступления натрия и борьбы с засолением
- 26. Механизм поступления натрия и борьбы с засолением 1. Вход Na+ 2. Активация сигнала засоления через АФК
- 27. Механизм поступления натрия и борьбы с засолением
- 28. Минимальные температуры роста вегетативных и генеративных органов различных растений, оС
- 30. Скачать презентацию

АФК-сенсоры (рецепторы) в клетках растений:
1. Двухкомпонентные гистидин-киназы;
2. Редокс-чувствительные факторы
АФК-сенсоры (рецепторы) в клетках растений:
1. Двухкомпонентные гистидин-киназы;
2. Редокс-чувствительные факторы

Кальций-связывающие белки (CBPs) – 5 классов:
- Кальмодулины
- кальмодулин-подобные белки
- Ca2+-зависимые
Кальций-связывающие белки (CBPs) – 5 классов:
- Кальмодулины
- кальмодулин-подобные белки
- Ca2+-зависимые

Гидроперекисное окисление липидов
(липиды – наиболее чувствительны к оксидативному стрессы, они способны
Гидроперекисное окисление липидов
(липиды – наиболее чувствительны к оксидативному стрессы, они способны

Липидный радикал (L•). L• может активировать O2 и формировать липидный пероксильный

(3) Фаза терминации
Липиды разлагаются во время стадии пропагации до терминальных низкомолекулярных
(3) Фаза терминации
Липиды разлагаются во время стадии пропагации до терминальных низкомолекулярных
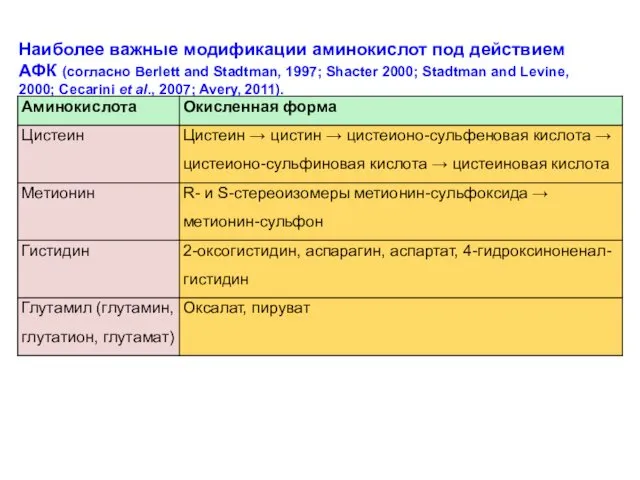
Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman,
Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman,
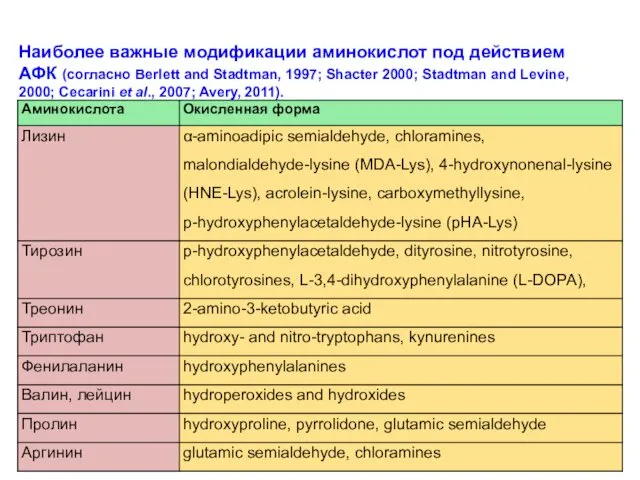
Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman,
Наиболее важные модификации аминокислот под действием АФК (согласно Berlett and Stadtman,

Основные окислительные повреждения биомолекул при стрессе:
Оксидативное повреждение белков
А - окисление
Основные окислительные повреждения биомолекул при стрессе:
Оксидативное повреждение белков
А - окисление

Б - Карбонилирование – формирование или встраивание свободной карбонильной группы (C=O)
Вторая
Б - Карбонилирование – формирование или встраивание свободной карбонильной группы (C=O)
Вторая

Карбонилирование приводит к образованию реактивных кетонов и альдегидов, детектируемых тестом Бради
Карбонилирование приводит к образованию реактивных кетонов и альдегидов, детектируемых тестом Бради
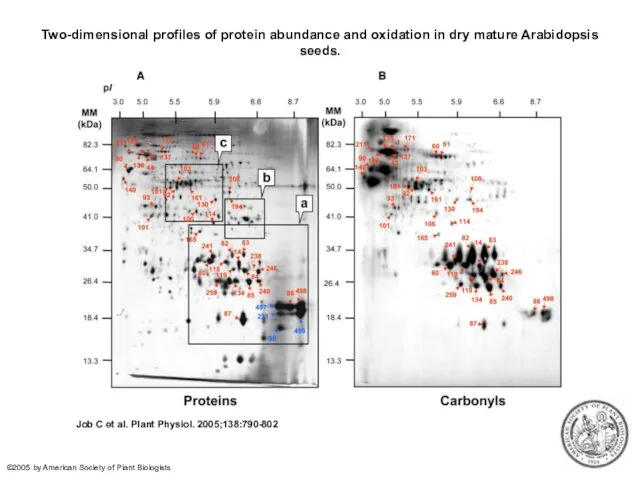
Two-dimensional profiles of protein abundance and oxidation in dry mature Arabidopsis
Two-dimensional profiles of protein abundance and oxidation in dry mature Arabidopsis

В – S-нитрозилирование.
Недавно открытое повреждение серусодержащих аминокислот белков под действием
В – S-нитрозилирование.
Недавно открытое повреждение серусодержащих аминокислот белков под действием

Оксидативное «повреждение» углеводов.
Моно- и дисахариды – важные АФК-связывающие агенты, проявляющие
Оксидативное «повреждение» углеводов.
Моно- и дисахариды – важные АФК-связывающие агенты, проявляющие

Оксидативное «повреждение» нуклеиновых кислот.
Для большинства однолетних культурных растений проблема рака
Оксидативное «повреждение» нуклеиновых кислот.
Для большинства однолетних культурных растений проблема рака

Основные антиоксидантные системы растений (см. также предыдущие лекции).
Ключевые ферментативные антиоксиданты
Основные антиоксидантные системы растений (см. также предыдущие лекции).
Ключевые ферментативные антиоксиданты

Основные антиоксидантные системы растений (см. также предыдущие лекции).
Ключевые неферментативные антиоксиданты
Основные антиоксидантные системы растений (см. также предыдущие лекции).
Ключевые неферментативные антиоксиданты

Вспомогательные системы антиоксидатной защиты растений.
Служат для поддержания антиоксидантов в функциональном
Вспомогательные системы антиоксидатной защиты растений.
Служат для поддержания антиоксидантов в функциональном

Пероксидазы растений:
Три «классических» типа: класс-I, класс-II and класс-III.
Пероксиредоксины сейчас
Пероксидазы растений:
Три «классических» типа: класс-I, класс-II and класс-III.
Пероксиредоксины сейчас

Пероксидазы растений:
Недавно найдены гибридные пероксидазы первого класса, совмещающие структуру каталазо-пероксидазы
Пероксидазы растений:
Недавно найдены гибридные пероксидазы первого класса, совмещающие структуру каталазо-пероксидазы

Пероксидазы растений:
Второй класс пероксидаз существует только у грибов – включает
Пероксидазы растений:
Второй класс пероксидаз существует только у грибов – включает

Общие защитные механизмы (против практически любого стрессового повреждения):
усиление катаболических процессов
Общие защитные механизмы (против практически любого стрессового повреждения):
усиление катаболических процессов
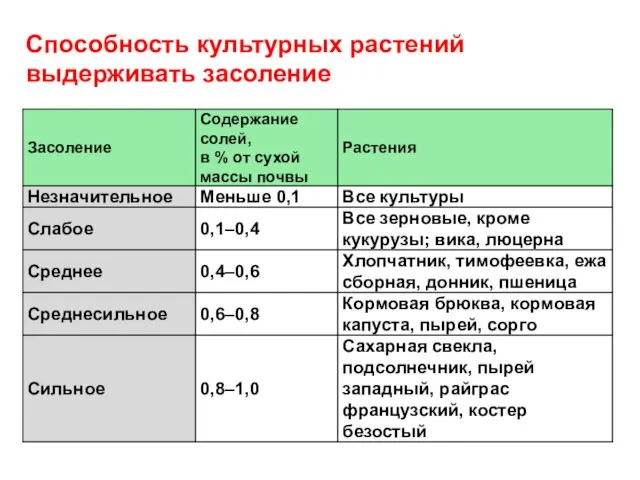
Способность культурных растений
выдерживать засоление
Способность культурных растений
выдерживать засоление
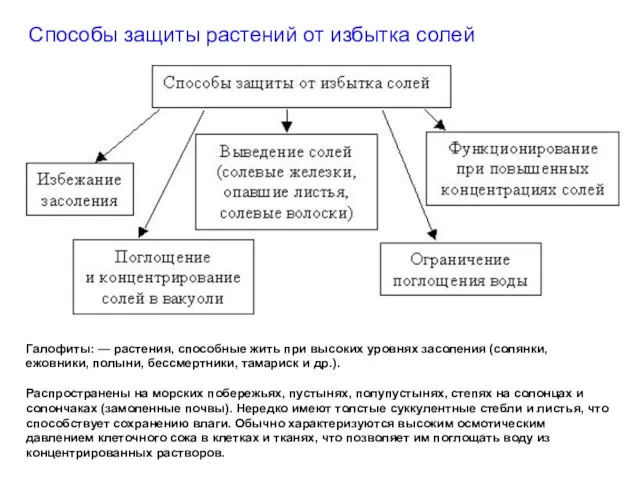
Способы защиты растений от избытка солей
Галофиты: — растения, способные жить при
Способы защиты растений от избытка солей
Галофиты: — растения, способные жить при
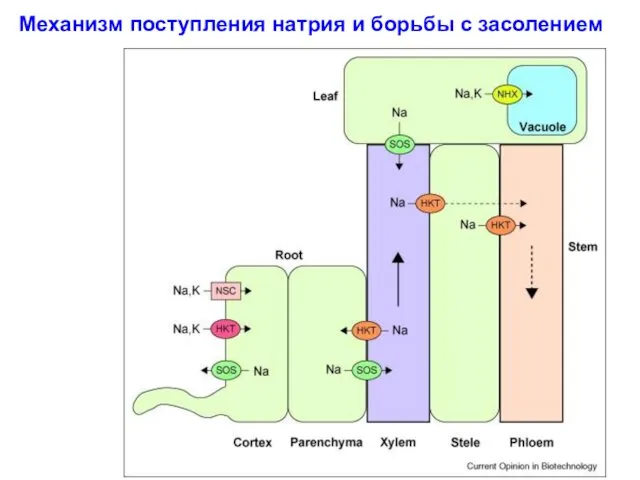
Механизм поступления натрия и борьбы с засолением
Механизм поступления натрия и борьбы с засолением

Механизм поступления натрия и борьбы с засолением
1. Вход Na+
2. Активация сигнала
Механизм поступления натрия и борьбы с засолением
1. Вход Na+
2. Активация сигнала
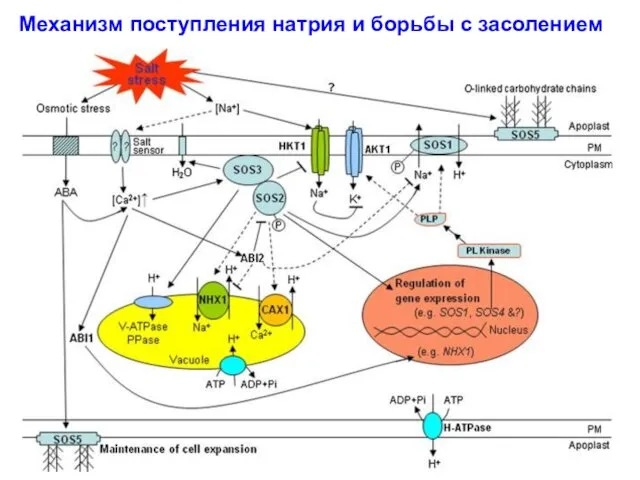
Механизм поступления натрия и борьбы с засолением
Механизм поступления натрия и борьбы с засолением
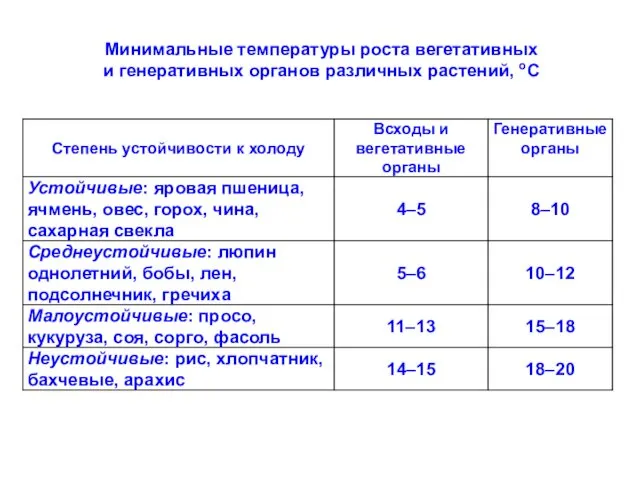
Минимальные температуры роста вегетативных
и генеративных органов различных растений, оС
Минимальные температуры роста вегетативных
и генеративных органов различных растений, оС
 Становлення глобальної економіки
Становлення глобальної економіки Мочекаменная болезнь: этиология и патогенез
Мочекаменная болезнь: этиология и патогенез 20240130_master-klass
20240130_master-klass My home
My home Web-дизайн. Введение в цветоведение
Web-дизайн. Введение в цветоведение Уроки настоящего. Новые технологии для предприятий Краснодарского края
Уроки настоящего. Новые технологии для предприятий Краснодарского края Архитектор Михаил Павлович Малахов (1781-1842)
Архитектор Михаил Павлович Малахов (1781-1842) Физкультминутки в детском саду
Физкультминутки в детском саду Повседневная жизнь горожан на Руси X-XIII века
Повседневная жизнь горожан на Руси X-XIII века Формирование учетной политики организации
Формирование учетной политики организации Аналитическая геометрия
Аналитическая геометрия Самозанятые. Краткое руководство
Самозанятые. Краткое руководство Физические основы механики молекулярная физика. Основы термодинамики
Физические основы механики молекулярная физика. Основы термодинамики Что такое толерантность?
Что такое толерантность? Методы и устройства согласования трактов СВЧ. Лекция 2
Методы и устройства согласования трактов СВЧ. Лекция 2 Животные весной
Животные весной Изображение и обозначение резьбы
Изображение и обозначение резьбы Make an opening statement on the subject of the project work
Make an opening statement on the subject of the project work Имануил Валлерстайн и Теда Скопкол. Историческая ориентация в марксизме
Имануил Валлерстайн и Теда Скопкол. Историческая ориентация в марксизме Презентация по краеведению
Презентация по краеведению Методы увеличения углеводородоотдачи
Методы увеличения углеводородоотдачи Agriculture Dialogue 2015
Agriculture Dialogue 2015 Вводное занятие по геометрии для учащихся 7 класса
Вводное занятие по геометрии для учащихся 7 класса Презентация Берингово море
Презентация Берингово море G`alamat online school
G`alamat online school Стальной низкотемпературный водогрейный котёл. Logano SK655/SK755
Стальной низкотемпературный водогрейный котёл. Logano SK655/SK755 Дресс-код делового человека
Дресс-код делового человека Богословие IV века. Святой Кирилл Иерусалимский (315-387 годы)
Богословие IV века. Святой Кирилл Иерусалимский (315-387 годы)