- Репликация

Содержание
- 2. План лекции: Репликация: определение, типы репликации геномов. Принципы репликации. Основные этапы репликации. Репликация теломерных отделов ДНК.
- 3. Литература: 1) Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология клетки т. 2 «Мир»
- 4. Если истинно утверждение, что сущность жизни состоит в накоплении и передаче опыта от поколения к поколению,
- 5. Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации. Суть репликации ДНК - образование идентичных копий
- 6. Типы репликации геномов. 1. Θ (тэта)-тип репликации кольцевой ДНК у бактерий. Начинается с определенной точки, идет
- 7. 2. σ (сигма)-тип репликации кольцевой ДНК-вирусов, называется «катящимся кольцом». В одной цепи кольцевой хромосомы происходит разрыв
- 9. 3. Полирепликонная репликация линейных молекул ДНК. Репликация в линейных хромосомах начинается в одной или нескольких точках,
- 10. Репликация ДНК идет на основе следующих принципов: Полуконсервативность Синтез ДНК начинается с расхождения цепей, каждая из
- 11. Консервативный способ репликации – когда после удвоения одна молекула состоит из двух старых цепей, другая –
- 12. Комплементарность Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В состав растущей цепи включается
- 13. Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном. В репликоне различают точку начала
- 14. Молекула ДНК, вступающая в репликацию: Точка, в которой происходит репликация называется репликационной вилкой (иногда наз. точкой
- 15. Репликация может идти либо в одном направлении, либо в двух направлениях. Это зависит от количества репликационных
- 16. Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот глазок выглядит одинаково при однонаправленной
- 17. При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную вилку. При двунаправленной репликации он
- 18. Этапы репликации: 1.Инициация идет с участием белков и ферментов, которые должны обеспечить: 1)Раскручивание ДНК 2)Связь инициирующих
- 19. Точки начала репликации богаты парами А-Т. Геликаза (от helix - спираль) расплетает двойную цепь родительской ДНК
- 20. Инициация репликации у эукариот. Белки инициации и происходящие процессы сходны с прокариотами. Отличия: 1.Участие дополнительного белка
- 21. Инициация репликации и клеточный цикл у дрожжей.
- 22. 2. Элонгация. Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых цепей ДНК, новая цепь
- 23. Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь называется запаздывающей. Длина ФО у
- 24. ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз: 1) ДНК-П I 2) ДНК-П II 3) ДНК-П III
- 25. ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её называют ферментом Корнберга. ДНК-П II
- 26. 3. Терминация репликации. У кишечной палочки есть ter-сайты, где происходит терминация репликации. У эукариот терминация репликации
- 28. В 80-х годах 20 века было установлено, что на концах хромосом есть особые структуры – теломеры,
- 29. Во время деления теломеры теряют от 5 до 20 фрагментов и с каждым делением становятся короче,
- 30. Проблема концевой недорепликации. Репликация на отстающей цепи ДНК начинается с синтеза коротких РНК-праймеров или затравок, с
- 31. Затем РНК-затравки удаляются, а образовавшиеся пустоты (бреши, гэпы) заполняются фрагментами ДНК. Причем при синтезе фрагментов ДНК
- 32. Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была открыта, а проблема неполной репликации
- 33. Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3` конец ДНК. Теломераза удлиняет не
- 34. По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной последовательности. В итоге дочерние цепи
- 35. Альтернативный механизм удлинения теломер (ALT – Alternative Lengtheining of Telomeres) без участия теломеразы (встречается у дрозофилы,
- 36. В клетках зародышевого пути теломеразы обладают высокой активностью, поэтому теломеры имеют нормальную длину. В соматических клетках,
- 37. При этом длина теломер стала удлиняться и увеличилась продолжительность жизни клеточных культур. Таким образом, теломеры имеют
- 38. Структура теломер. 1) определенный нуклеотидный состав 2) специфические белки, которые отличаются от обычных гистонов и не
- 39. Функции теломер 1) механическая: фиксация хромосом к ядерному матриксу; сцепление друг с другом концов сестринских хроматид
- 40. Выводы: Репликация – матричный процесс. Во время репликации каждая из двух цепей ДНК служит матрицей для
- 41. ДНК полимеразы :δ α синтез новых цепей ДНК ε ДНК - полимераза β- удаление праймеров 4.
- 42. 5. По завершении репликации образуются две молекулы 2-х спиральной ДНК, каждая из которых содержит одну матричную
- 44. Скачать презентацию

План лекции:
Репликация: определение, типы репликации геномов.
Принципы репликации.
Основные этапы репликации.
Репликация теломерных
План лекции:
Репликация: определение, типы репликации геномов.
Принципы репликации.
Основные этапы репликации.
Репликация теломерных

Литература:
1) Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология
Литература:
1) Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология

Если истинно утверждение, что сущность жизни состоит в накоплении и передаче
Если истинно утверждение, что сущность жизни состоит в накоплении и передаче
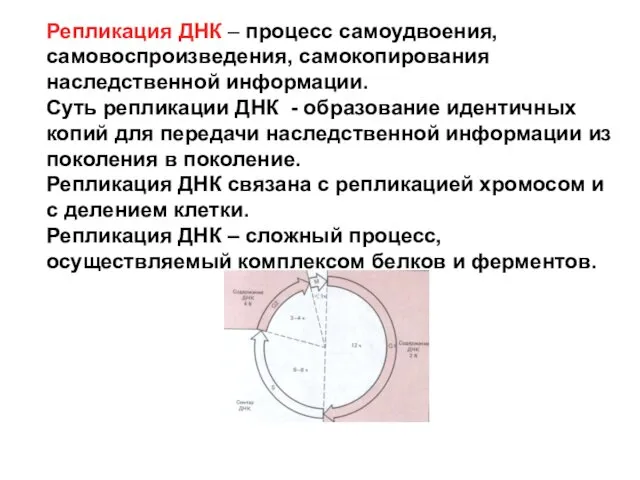
Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации.
Суть репликации ДНК
Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации.
Суть репликации ДНК
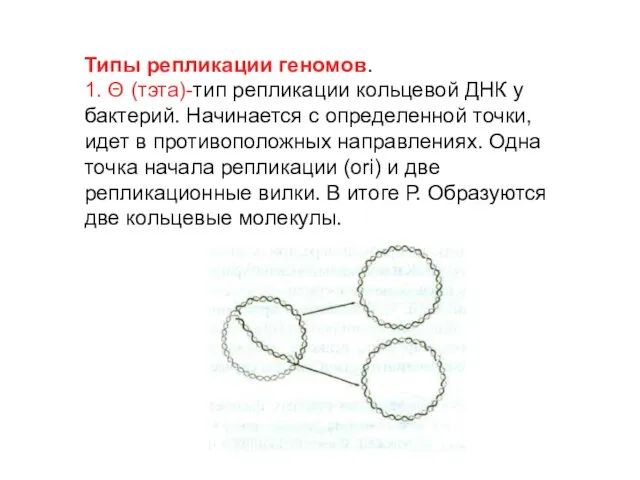
Типы репликации геномов.
1. Θ (тэта)-тип репликации кольцевой ДНК у бактерий. Начинается
Типы репликации геномов.
1. Θ (тэта)-тип репликации кольцевой ДНК у бактерий. Начинается

2. σ (сигма)-тип репликации кольцевой ДНК-вирусов, называется «катящимся кольцом». В одной
2. σ (сигма)-тип репликации кольцевой ДНК-вирусов, называется «катящимся кольцом». В одной

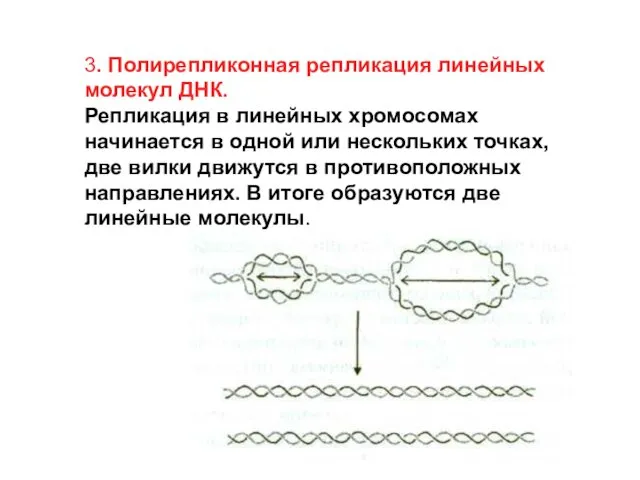
3. Полирепликонная репликация линейных молекул ДНК.
Репликация в линейных хромосомах начинается в
3. Полирепликонная репликация линейных молекул ДНК.
Репликация в линейных хромосомах начинается в
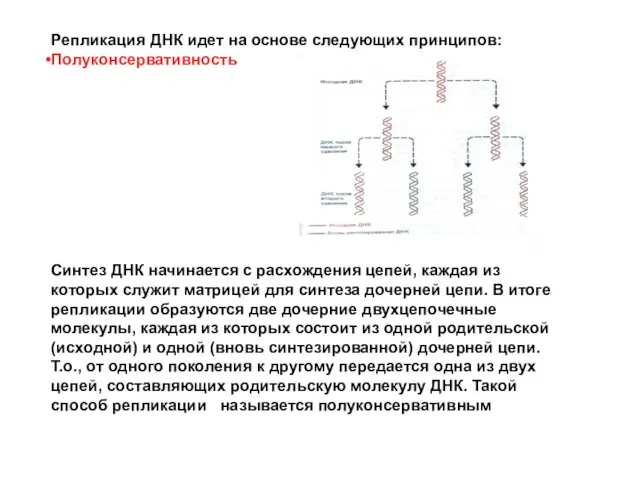
Репликация ДНК идет на основе следующих принципов:
Полуконсервативность
Синтез ДНК начинается с расхождения
Репликация ДНК идет на основе следующих принципов:
Полуконсервативность
Синтез ДНК начинается с расхождения

Консервативный способ репликации – когда после удвоения одна молекула состоит из
Консервативный способ репликации – когда после удвоения одна молекула состоит из

Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В
Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В

Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном.
Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном.

Молекула ДНК, вступающая в репликацию:
Точка, в которой происходит репликация называется репликационной
Молекула ДНК, вступающая в репликацию:
Точка, в которой происходит репликация называется репликационной

Репликация может идти либо в одном направлении, либо в двух направлениях.
Репликация может идти либо в одном направлении, либо в двух направлениях.
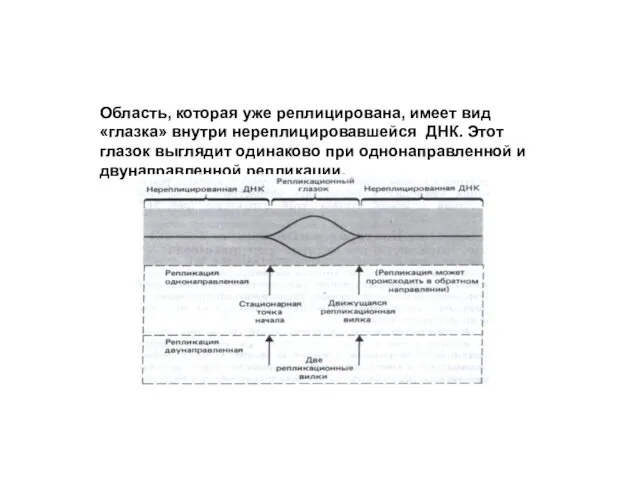
Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот
Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот

При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную
При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную

Этапы репликации:
1.Инициация идет с участием белков и ферментов, которые должны обеспечить:
1)Раскручивание
Этапы репликации:
1.Инициация идет с участием белков и ферментов, которые должны обеспечить:
1)Раскручивание

Точки начала репликации богаты парами А-Т.
Геликаза (от helix - спираль)
Точки начала репликации богаты парами А-Т.
Геликаза (от helix - спираль)

Инициация репликации у эукариот.
Белки инициации и происходящие процессы сходны с прокариотами.
Инициация репликации у эукариот.
Белки инициации и происходящие процессы сходны с прокариотами.
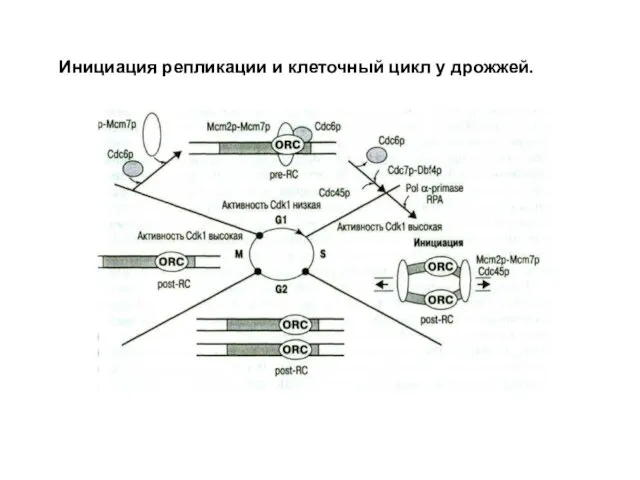
Инициация репликации и клеточный цикл у дрожжей.
Инициация репликации и клеточный цикл у дрожжей.

2. Элонгация.
Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых
2. Элонгация.
Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых

Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь
Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь

ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз:
1) ДНК-П I
2) ДНК-П II
3)
ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз:
1) ДНК-П I
2) ДНК-П II
3)

ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её
ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её

3. Терминация репликации.
У кишечной палочки есть ter-сайты, где происходит терминация репликации.
У
3. Терминация репликации.
У кишечной палочки есть ter-сайты, где происходит терминация репликации.
У

В 80-х годах 20 века было установлено, что на концах хромосом
В 80-х годах 20 века было установлено, что на концах хромосом

Во время деления теломеры теряют от 5 до 20 фрагментов и
Во время деления теломеры теряют от 5 до 20 фрагментов и

Проблема концевой недорепликации.
Репликация на отстающей цепи ДНК начинается с синтеза
Проблема концевой недорепликации.
Репликация на отстающей цепи ДНК начинается с синтеза

Затем РНК-затравки удаляются, а образовавшиеся пустоты (бреши, гэпы) заполняются фрагментами ДНК.
Затем РНК-затравки удаляются, а образовавшиеся пустоты (бреши, гэпы) заполняются фрагментами ДНК.

Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была
Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была

Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3`
Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3`

По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной
По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной

Альтернативный механизм удлинения теломер (ALT – Alternative Lengtheining of Telomeres) без
Альтернативный механизм удлинения теломер (ALT – Alternative Lengtheining of Telomeres) без

В клетках зародышевого пути теломеразы обладают высокой активностью, поэтому теломеры имеют
В клетках зародышевого пути теломеразы обладают высокой активностью, поэтому теломеры имеют

При этом длина теломер стала удлиняться и увеличилась продолжительность жизни клеточных
При этом длина теломер стала удлиняться и увеличилась продолжительность жизни клеточных

Структура теломер.
1) определенный нуклеотидный состав
2) специфические белки, которые отличаются от
Структура теломер.
1) определенный нуклеотидный состав
2) специфические белки, которые отличаются от

Функции теломер
1) механическая: фиксация хромосом к ядерному матриксу; сцепление друг с
Функции теломер
1) механическая: фиксация хромосом к ядерному матриксу; сцепление друг с

Выводы:
Репликация – матричный процесс. Во время репликации каждая из двух цепей
Выводы:
Репликация – матричный процесс. Во время репликации каждая из двух цепей

ДНК полимеразы :δ
α синтез новых цепей ДНК
ε
ДНК - полимераза
α синтез новых цепей ДНК
ε
ДНК - полимераза

5. По завершении репликации образуются две молекулы 2-х спиральной ДНК, каждая
5. По завершении репликации образуются две молекулы 2-х спиральной ДНК, каждая
 Стартап за неделю
Стартап за неделю Презентация Любовь - основа жизни, ОРКСЭ, урок № 20, к учебнику Шемшуриной А. (Дрофа), 4 класс
Презентация Любовь - основа жизни, ОРКСЭ, урок № 20, к учебнику Шемшуриной А. (Дрофа), 4 класс Презентация театрального кружка в детском саду ТЕАТР И ДЕТИ
Презентация театрального кружка в детском саду ТЕАТР И ДЕТИ Grade can we speak to rik morell
Grade can we speak to rik morell Правила поведения детей на железнодорожном транспорте
Правила поведения детей на железнодорожном транспорте Система коррекционной работы с детьми раннего возраста
Система коррекционной работы с детьми раннего возраста Электр станциялары
Электр станциялары Разъединители. Коммутационная аппаратура
Разъединители. Коммутационная аппаратура Начало освоения Новороссии и Крыма
Начало освоения Новороссии и Крыма Теория отраслевых рынков (разделы курса)
Теория отраслевых рынков (разделы курса) Бессмертный полк
Бессмертный полк Цветущая ветка вишни
Цветущая ветка вишни Деепричастие. Деепричастие оборот
Деепричастие. Деепричастие оборот Типы и виды очистных снарядов
Типы и виды очистных снарядов Реализм. Новые жанры
Реализм. Новые жанры педагогический проект
педагогический проект 70 лет Победы
70 лет Победы 3 ФГОС общие ХАРАКТЕРИСТИКИ 3 ФГОС общие ХАРАКТЕРИСТИКИ
3 ФГОС общие ХАРАКТЕРИСТИКИ 3 ФГОС общие ХАРАКТЕРИСТИКИ h1495273913
h1495273913 Высказывания: Истинные, Ложные
Высказывания: Истинные, Ложные Морская лоция. Несение ходовой навигационной вахты на вспомогательном уровне ПДМНВ ІІ/4
Морская лоция. Несение ходовой навигационной вахты на вспомогательном уровне ПДМНВ ІІ/4 Спортивное питание
Спортивное питание Муниципальное бюджетное учреждение Спортивная школа №2 г. Ижевск
Муниципальное бюджетное учреждение Спортивная школа №2 г. Ижевск Радиоактивные изотопы в биологии и медицине
Радиоактивные изотопы в биологии и медицине Проект: Борьба Хуреш- игра сильных!
Проект: Борьба Хуреш- игра сильных! Germany
Germany Алкоголь и его влияние на здоровье человека
Алкоголь и его влияние на здоровье человека Как рисовать красивую кошку (1 класс)
Как рисовать красивую кошку (1 класс)