- Репликация ДНК. Мутации

Содержание
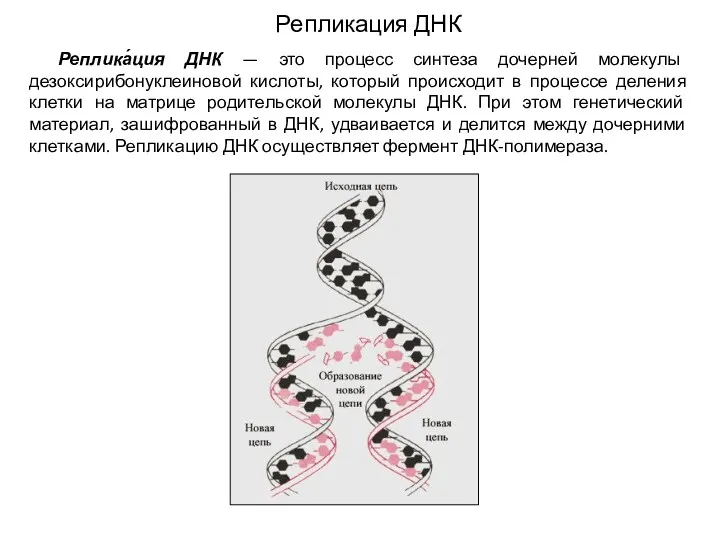
- 2. Репликация ДНК Реплика́ция ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, который происходит в процессе
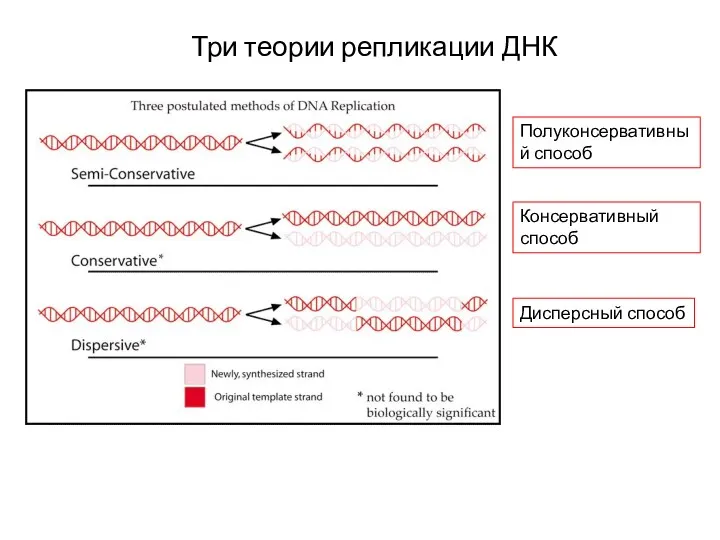
- 3. Три теории репликации ДНК Полуконсервативный способ Консервативный способ Дисперсный способ
- 4. Схема эксперимента Мезельсона и Сталя, доказывающего полуконсервативный способ репликации ДНК.
- 5. Цепи молекулы ДНК расходятся и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь.
- 6. Кольцевая вирусная ДНК Кольцевая бактериальная ДНК Эукариотическая ДНК У вирусов и прокариот репликация ДНК начинается с
- 7. В репликация участвует комплекс специфических ферментов. Хеликаза, топоизомераза и ДНК-связывающие белки, которые расплетают ДНК, удерживают матрицу
- 8. Изучение ферментативного синтеза показало, одновременно копируются обе цепи ДНК. Полимеризация одной цепи происходит в направление от
- 9. Основные этапы репликации ДНК
- 10. Раскручивание двойной цепи ДНК происходит с участием трёх разных белков: А) белки (SSB) , дестабилизирующие спираль
- 11. У эукариот известно пять ДНК-полимераз (α, β, γ, δ, ε), из которых главную роль в репликации
- 12. Репликационное поведение хромосом основывается на трёх фундаментальных свойствах: 1) непосредственно репликация; 2) сегрегация хромосом при репликации
- 13. Мутации Мутации – это изменение генов и хромосом фенотипически проявляющиеся в изменении свойств и признаков организма.
- 14. Организмы, содержащие в своём геноме одну или несколько мутаций называются мутантными. Они могут отличаться от исходных
- 15. Генеративные мутации происходят в генитальных и половых клетках. Если мутация происходит в генитальных клетках, то мутантный
- 17. На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных
- 18. Полиплоидия – хромосомная мутация в виде увеличения числа полных гаплоидных наборов хромосом. Наблюдается в основном у
- 19. Гаплоидия - противоположное полиплоидии явление, заключающееся в кратном уменьшении числа хромосом у потомства в сравнении с
- 20. Гетероплоидия. Изменение числа хромосом, некратное гаплоидному набору. Так, в диплоидном наборе может быть всего на 1
- 21. В зависимости от происхождения различают спонтанные и индуцированные мутации. Спонтанными называют мутации, которые возникают у организмов
- 22. Индуцированными являются те мутации, которые возникают после обработки клеток (организмов) мутагенными факторами – мутагенами. Мутагены -
- 23. Физические мутагены Среди физических мутагенов наибольшим эффектом обладает ионизирующие излучения и УФ – излучение. Ионизирующие излучения
- 24. Химические мутагены Некоторые химические мутагены нарушаю нормальный процесс мейоза, что приводи к нерасхождению хромосом, а также
- 25. Биологические мутагены Вирусы. Вызывают хромосомные абберации в культивируемых клетках. Такой способностью обладает вирус гриппа, Эпштена-Барра, кори,
- 26. Репарация повреждений ДНК Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы
- 27. Р. Сетлоу, К. Руперт (США) и др. установили, что фотореактивация - фотохимический процесс, протекающий с участием
- 29. Скачать презентацию
Репликация ДНК
Реплика́ция ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты,
Репликация ДНК
Реплика́ция ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты,
Три теории репликации ДНК
Полуконсервативный способ
Консервативный способ
Дисперсный способ
Три теории репликации ДНК
Полуконсервативный способ
Консервативный способ
Дисперсный способ
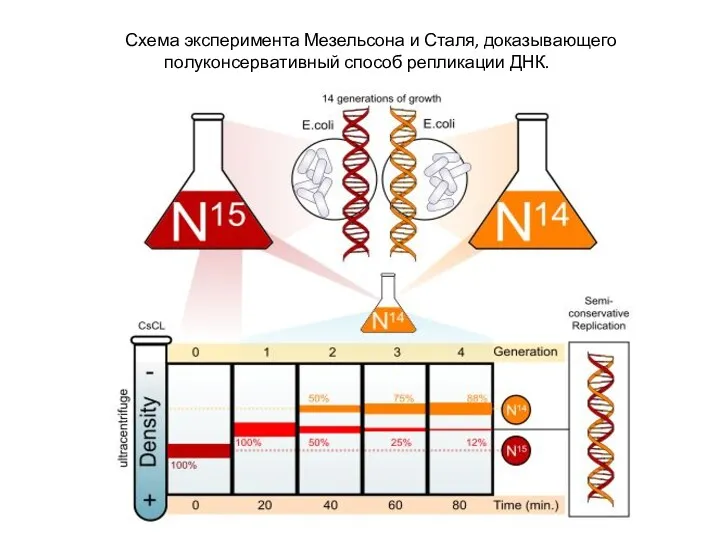
Схема эксперимента Мезельсона и Сталя, доказывающего полуконсервативный способ репликации ДНК.
Схема эксперимента Мезельсона и Сталя, доказывающего полуконсервативный способ репликации ДНК.
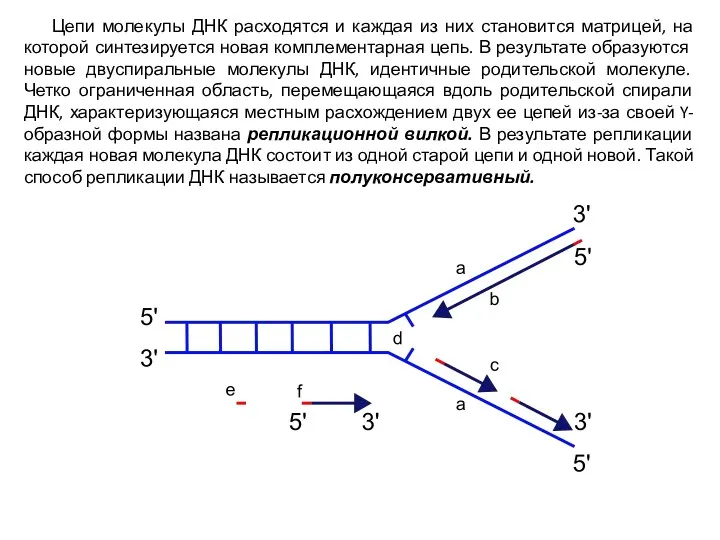
Цепи молекулы ДНК расходятся и каждая из них становится матрицей, на
Цепи молекулы ДНК расходятся и каждая из них становится матрицей, на
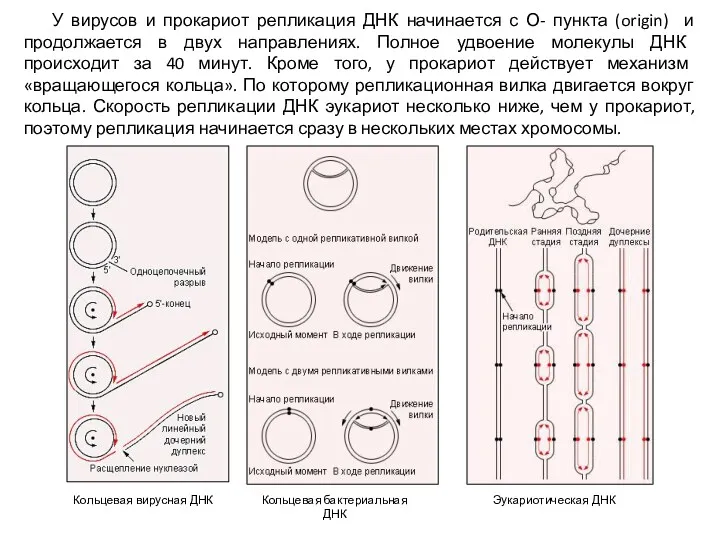
Кольцевая вирусная ДНК
Кольцевая бактериальная ДНК
Эукариотическая ДНК
У вирусов и прокариот репликация ДНК
Кольцевая вирусная ДНК
Кольцевая бактериальная ДНК
Эукариотическая ДНК
У вирусов и прокариот репликация ДНК
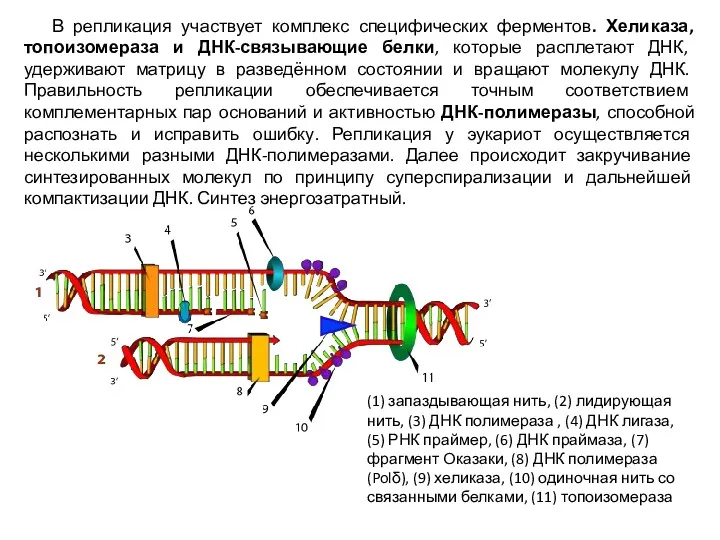
В репликация участвует комплекс специфических ферментов. Хеликаза, топоизомераза и ДНК-связывающие белки,
В репликация участвует комплекс специфических ферментов. Хеликаза, топоизомераза и ДНК-связывающие белки,
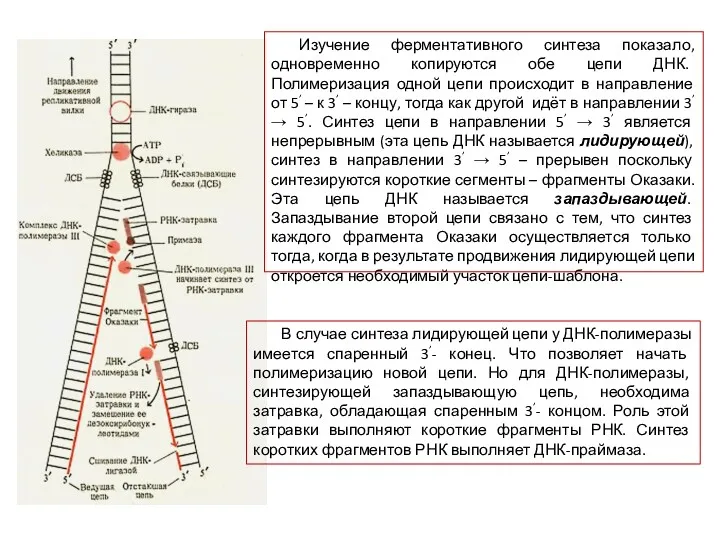
Изучение ферментативного синтеза показало, одновременно копируются обе цепи ДНК. Полимеризация одной
Изучение ферментативного синтеза показало, одновременно копируются обе цепи ДНК. Полимеризация одной

Основные этапы репликации ДНК
Основные этапы репликации ДНК
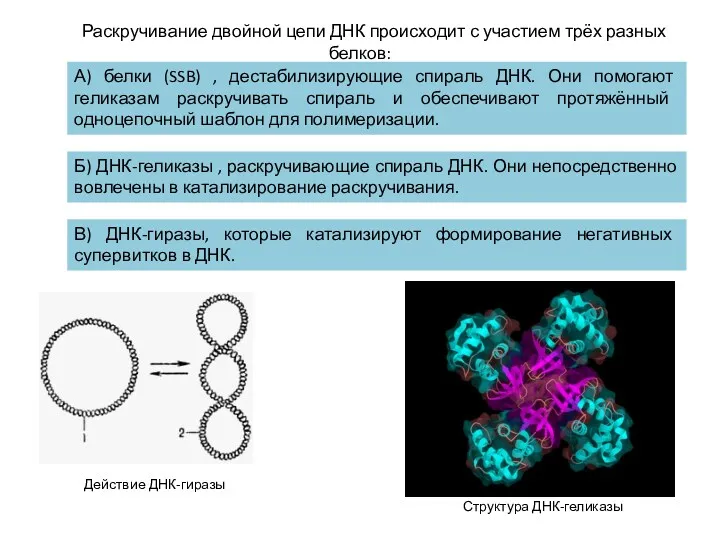
Раскручивание двойной цепи ДНК происходит с участием трёх разных белков:
А) белки
Раскручивание двойной цепи ДНК происходит с участием трёх разных белков:
А) белки
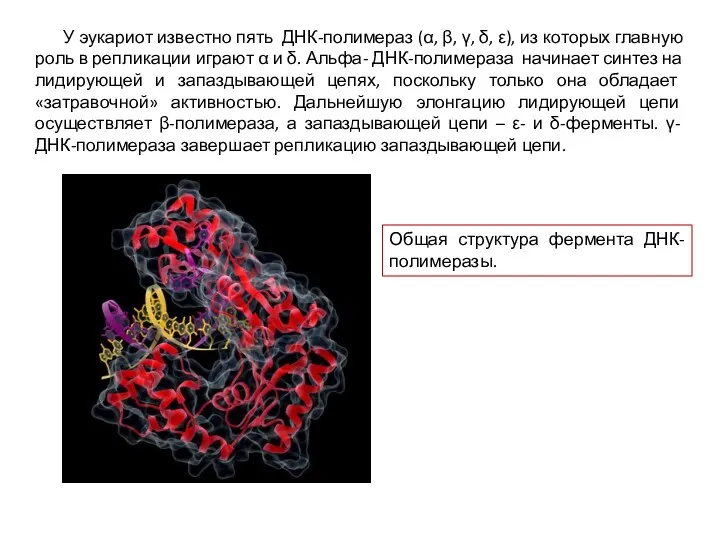
У эукариот известно пять ДНК-полимераз (α, β, γ, δ, ε), из
У эукариот известно пять ДНК-полимераз (α, β, γ, δ, ε), из
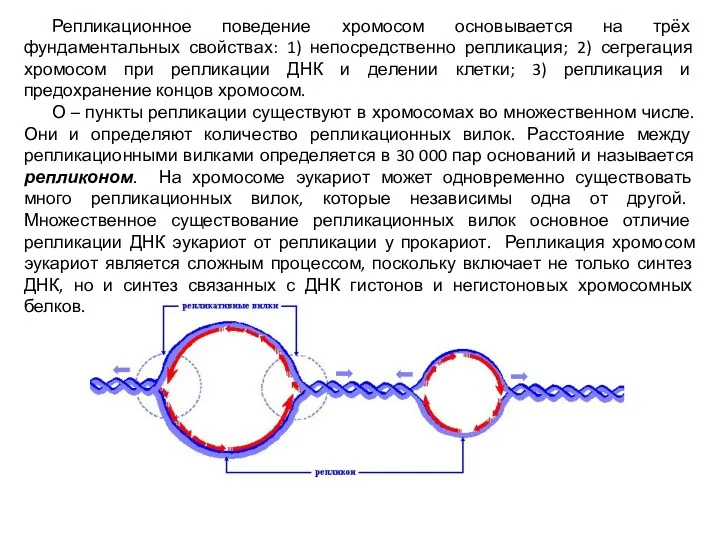
Репликационное поведение хромосом основывается на трёх фундаментальных свойствах: 1) непосредственно репликация;
Репликационное поведение хромосом основывается на трёх фундаментальных свойствах: 1) непосредственно репликация;

Мутации
Мутации – это изменение генов и хромосом фенотипически проявляющиеся в изменении
Мутации
Мутации – это изменение генов и хромосом фенотипически проявляющиеся в изменении

Организмы, содержащие в своём геноме одну или несколько мутаций называются мутантными.
Организмы, содержащие в своём геноме одну или несколько мутаций называются мутантными.
Генеративные мутации происходят в генитальных и половых клетках. Если мутация происходит
Генеративные мутации происходят в генитальных и половых клетках. Если мутация происходит
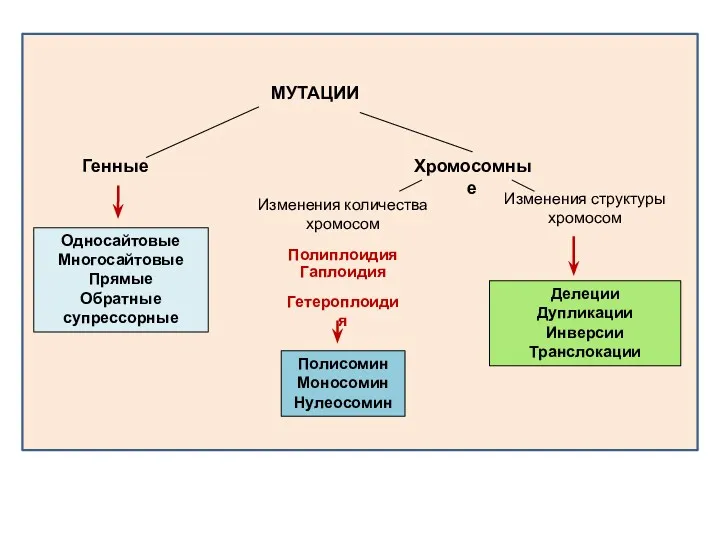
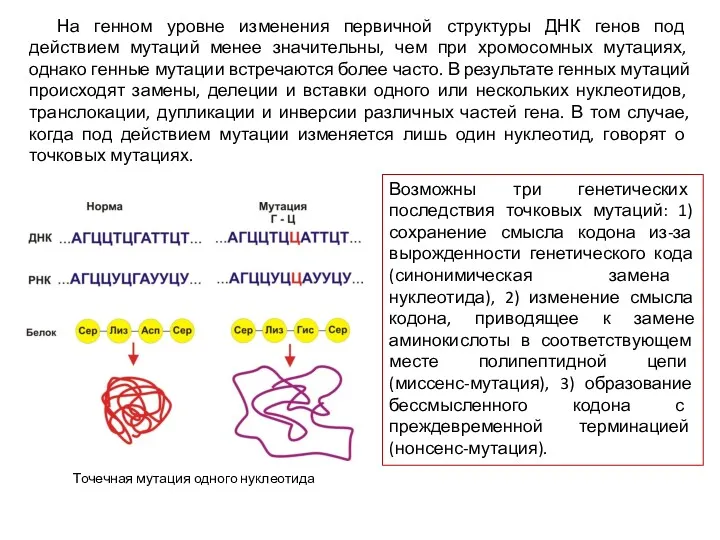
На генном уровне изменения первичной структуры ДНК генов под действием мутаций
На генном уровне изменения первичной структуры ДНК генов под действием мутаций
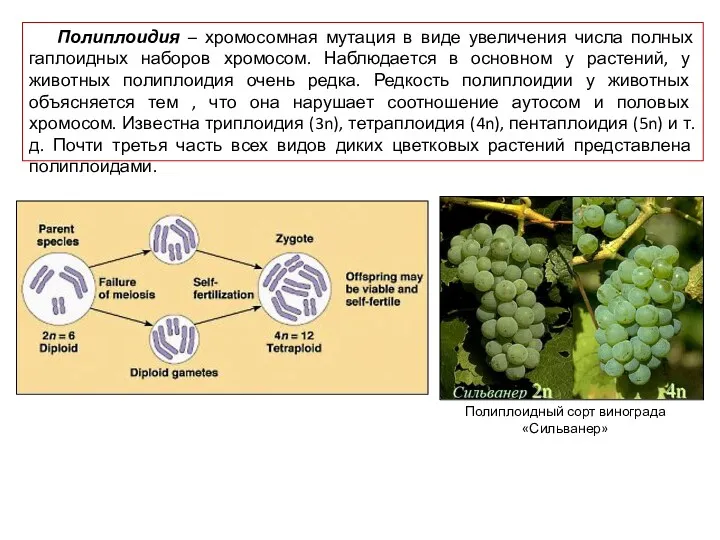
Полиплоидия – хромосомная мутация в виде увеличения числа полных гаплоидных наборов
Полиплоидия – хромосомная мутация в виде увеличения числа полных гаплоидных наборов

Гаплоидия - противоположное полиплоидии явление, заключающееся в кратном уменьшении числа хромосом
Гаплоидия - противоположное полиплоидии явление, заключающееся в кратном уменьшении числа хромосом

Гетероплоидия. Изменение числа хромосом, некратное гаплоидному набору. Так, в диплоидном наборе
Гетероплоидия. Изменение числа хромосом, некратное гаплоидному набору. Так, в диплоидном наборе

В зависимости от происхождения различают спонтанные и индуцированные мутации.
Спонтанными называют мутации,
В зависимости от происхождения различают спонтанные и индуцированные мутации.
Спонтанными называют мутации,
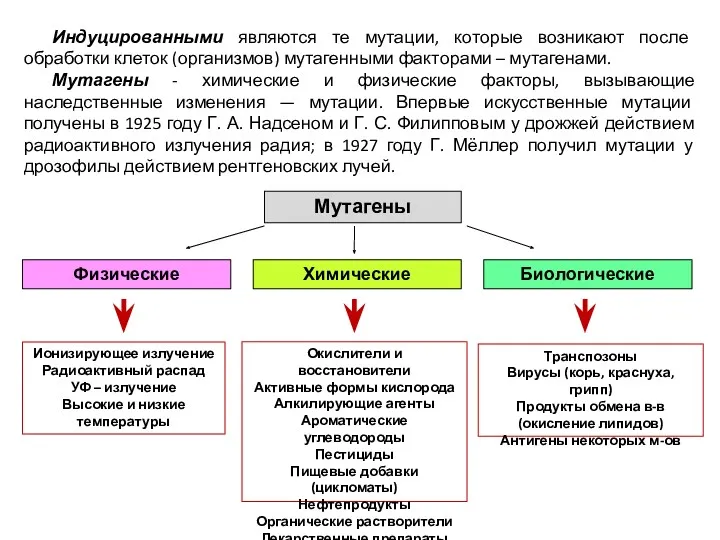
Индуцированными являются те мутации, которые возникают после обработки клеток (организмов) мутагенными
Индуцированными являются те мутации, которые возникают после обработки клеток (организмов) мутагенными
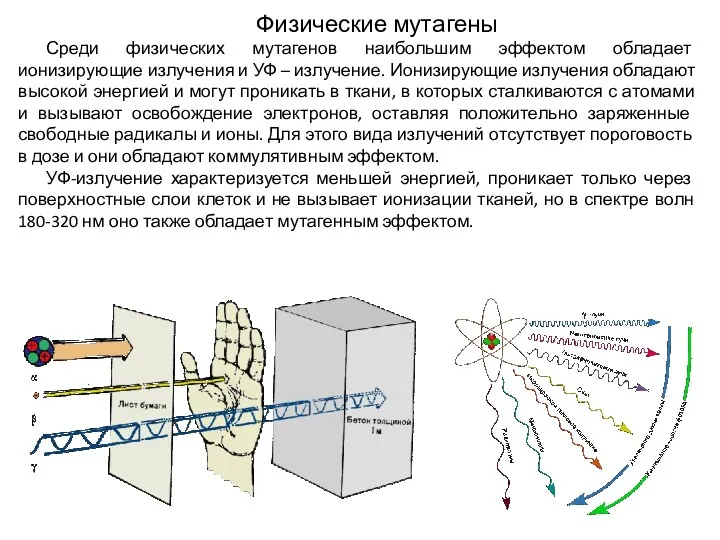
Физические мутагены
Среди физических мутагенов наибольшим эффектом обладает ионизирующие излучения и УФ
Физические мутагены
Среди физических мутагенов наибольшим эффектом обладает ионизирующие излучения и УФ

Химические мутагены
Некоторые химические мутагены нарушаю нормальный процесс мейоза, что приводи к
Химические мутагены
Некоторые химические мутагены нарушаю нормальный процесс мейоза, что приводи к

Биологические мутагены
Вирусы. Вызывают хромосомные абберации в культивируемых клетках. Такой способностью обладает
Биологические мутагены
Вирусы. Вызывают хромосомные абберации в культивируемых клетках. Такой способностью обладает
Репарация повреждений ДНК
Репарация — особая функция клеток, заключающаяся в способности исправлять
Репарация повреждений ДНК
Репарация — особая функция клеток, заключающаяся в способности исправлять
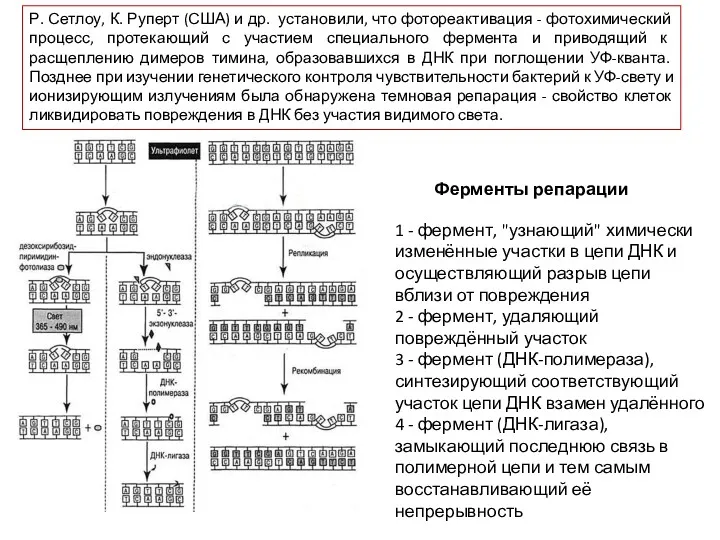
Р. Сетлоу, К. Руперт (США) и др. установили, что фотореактивация -
Р. Сетлоу, К. Руперт (США) и др. установили, что фотореактивация -
 Вид. Структура и критерии вида
Вид. Структура и критерии вида 20200117_svoystva_tkaney.ppt
20200117_svoystva_tkaney.ppt Внешняя политика Александра I
Внешняя политика Александра I Развитие школьника как языковой личности в процессе обучения и воспитания
Развитие школьника как языковой личности в процессе обучения и воспитания Террористический акт
Террористический акт Учет взносов и отчислений на обязательное социальное медицинское страхование (ОСМС)
Учет взносов и отчислений на обязательное социальное медицинское страхование (ОСМС) Животные – переносчики заболеваний
Животные – переносчики заболеваний 40-55 жас аралығындағы ішек өткізбеушілігі диагнозымен ауыратын науқастарда перитониттің алдын алу үшін энтеростомияға қарағанда
40-55 жас аралығындағы ішек өткізбеушілігі диагнозымен ауыратын науқастарда перитониттің алдын алу үшін энтеростомияға қарағанда Построение сложных запросов. Инсерт
Построение сложных запросов. Инсерт мастер-класс по теме ФГОС через призму своего опыта: формируем метапредметные и личностные результаты
мастер-класс по теме ФГОС через призму своего опыта: формируем метапредметные и личностные результаты Презентация История одной песни Смуглянка
Презентация История одной песни Смуглянка Адаптация – результат эволюции
Адаптация – результат эволюции Четырехэтапная мозговая атака
Четырехэтапная мозговая атака Сетевой протокол
Сетевой протокол Механический привод
Механический привод Текстильные изделия в интерьере
Текстильные изделия в интерьере Организация развивающей предметно-пространственной среды логопедического кабинета МБДОУ в условиях реализации ФГОС.
Организация развивающей предметно-пространственной среды логопедического кабинета МБДОУ в условиях реализации ФГОС. Античный миф
Античный миф Развитие духовно-нравственных основ личности ребенка
Развитие духовно-нравственных основ личности ребенка Нормативное обоснование введения в образовательный процесс электронных форм учебников
Нормативное обоснование введения в образовательный процесс электронных форм учебников Монголо-татарское нашествие на Русь (1237-1240)
Монголо-татарское нашествие на Русь (1237-1240) Сборка компьютера с умом
Сборка компьютера с умом Скульптура. Жанры скульптуры
Скульптура. Жанры скульптуры 공공시설 이용
공공시설 이용 Умножение десятичных дробей на натуральное число.
Умножение десятичных дробей на натуральное число. Новый сценарий Вашей жизни. Семинар на волшебном острове Тенерифе с 21 февраля по 1 марта
Новый сценарий Вашей жизни. Семинар на волшебном острове Тенерифе с 21 февраля по 1 марта Освобождение Заполярья от фашистских захватчиков
Освобождение Заполярья от фашистских захватчиков Генеральный план
Генеральный план