- Транскрипция. Центральная догма молекулярной биологии

Содержание
- 2. Центральная догма молекулярной биологии
- 3. Пространственная структура РНК Одноцепочечная; Шпильки – спирализованные участки (водородные связи); Не соблюдается правило Чаргаффа; Виды РНК:
- 4. тРНК Структура тРНК: 1 – шпильки 2 - петли молекулы-адапторы: переводят информацию мРНК в последовательность аминокислот
- 5. структурный компонент рибосом; 80% от общего количества РНК в клетке; 4 типа у эукариот: 5S, 5,8S,
- 6. Транскрипция - (лат. transcriptio — переписывание) процесс синтеза РНК с использованием ДНК в качестве матрицы. Направление
- 7. Транскрипция: общая характеристика Работа РНК-полимеразы, скорость ~15-20 нукл./сек. Субстрат – рибонуклеозидтрифосфаты.
- 8. Транскрипция: общая характеристика Инициация – Элонгация - Терминация
- 9. Транскрипция: общая характеристика Транскрипция генов, наблюдаемая в электронный микроскоп
- 10. Транскрипция у прокариот Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции и мРНК деградации могут
- 11. Транскрипция у прокариот Промотор – точка старта РНК-полимеразы; При инициации σ субъединица освобождается
- 12. Транскрипция у прокариот Скорость РНК-полимеразы составляет примерно 40 нуклеотидов в секунду; В течении транскрипции ДНК перед
- 13. Регуляция Транскрипции у прокариот Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны
- 14. Регуляция Транскрипции у прокариот
- 15. Транскрипция у эукариот Транскрипция у эукариот происходит в ядре; Синтез молекул РНК начинается с промоторов, и
- 16. Транскрипция у эукариот 1. Инициация Активация промотора происходит с помощью большого белка - ТАТА-фактора, связывающегося с
- 17. Транскрипция у эукариот 1. Инициация Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают раскручивание примерно одного
- 18. Транскрипция у эукариот 2. Элонгация На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18
- 19. Транскрипция у эукариот 3. Терминация Завершается синтез РНК в строго определенных участках матрицы - сайты терминации
- 20. Факторы Транскрипции у эукариот Регуляция базальной экспрессии генов; Регуляция онтогенеза; Ответ на изменение окружающей среды; Контроль
- 21. Процессинг мрнк у эукариот: кэпирование Модифицированный 5'-конец обеспечивает инициацию трансляции, удлиняет время жизни мРНК, защищая её
- 22. Процессинг мрнк у эукариот: полиаденилирование Специальным ферментом полиА-полимеразой формируется полиА-последовательность (полиА-"хвост"), состоящая из 100-200 остатков аденозина.
- 23. Процессинг мрнк у эукариот: сплайсинг Последовательности интронов "вырезаются" из первичного транскрипта, концы экзонов соединяются. Гены эукариотов
- 25. Скачать презентацию
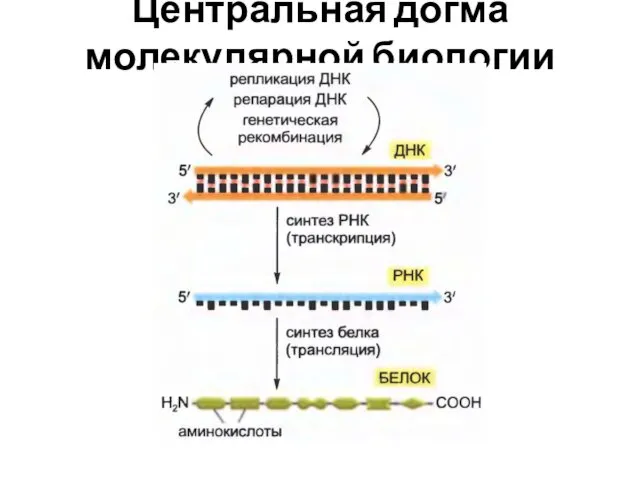
Центральная догма молекулярной биологии
Центральная догма молекулярной биологии

Пространственная структура РНК
Одноцепочечная;
Шпильки – спирализованные участки (водородные связи);
Не соблюдается правило Чаргаффа;
Виды
Пространственная структура РНК
Одноцепочечная;
Шпильки – спирализованные участки (водородные связи);
Не соблюдается правило Чаргаффа;
Виды
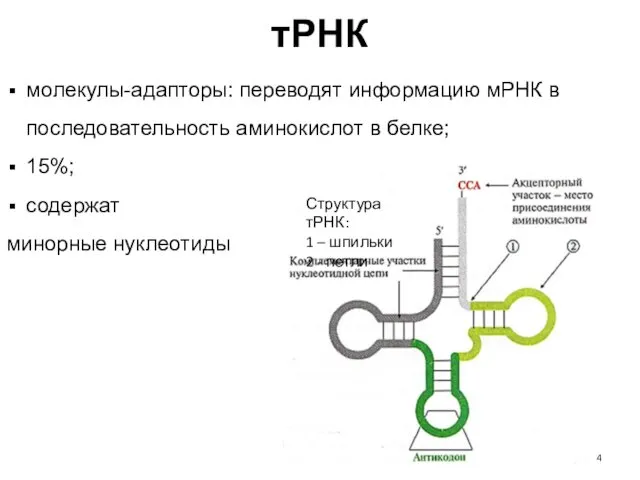
тРНК
Структура тРНК:
1 – шпильки
2 - петли
молекулы-адапторы: переводят информацию мРНК в последовательность
тРНК
Структура тРНК:
1 – шпильки
2 - петли
молекулы-адапторы: переводят информацию мРНК в последовательность
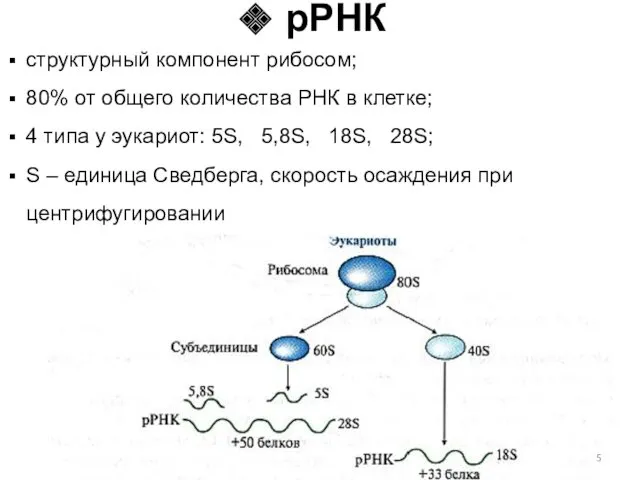
структурный компонент рибосом;
80% от общего количества РНК в клетке;
4 типа у
структурный компонент рибосом;
80% от общего количества РНК в клетке;
4 типа у
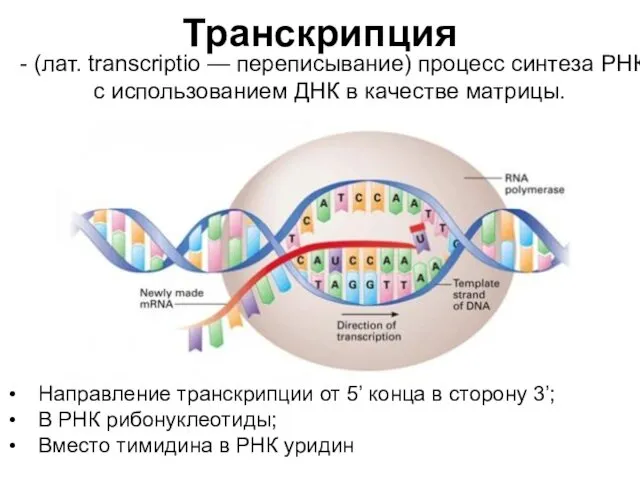
Транскрипция
- (лат. transcriptio — переписывание) процесс синтеза РНК с использованием ДНК
Транскрипция
- (лат. transcriptio — переписывание) процесс синтеза РНК с использованием ДНК
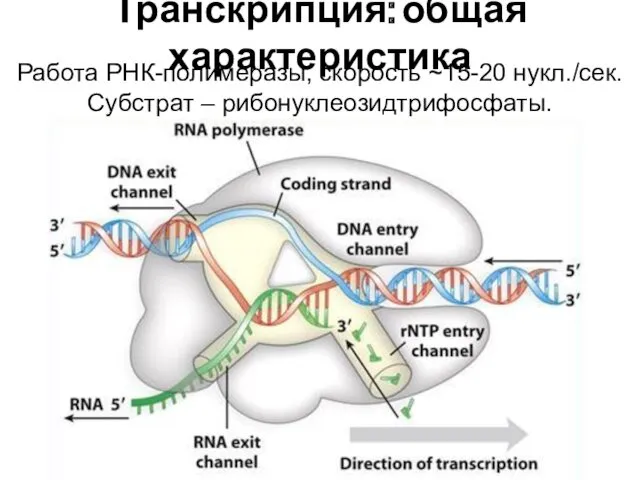
Транскрипция: общая характеристика
Работа РНК-полимеразы, скорость ~15-20 нукл./сек. Субстрат – рибонуклеозидтрифосфаты.
Транскрипция: общая характеристика
Работа РНК-полимеразы, скорость ~15-20 нукл./сек. Субстрат – рибонуклеозидтрифосфаты.
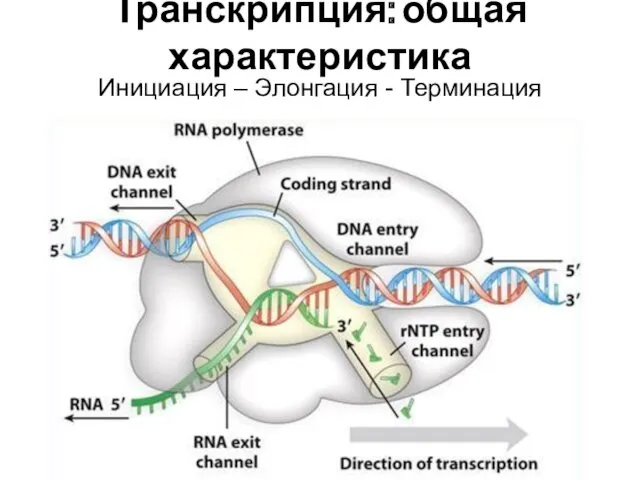
Транскрипция: общая характеристика
Инициация – Элонгация - Терминация
Транскрипция: общая характеристика
Инициация – Элонгация - Терминация
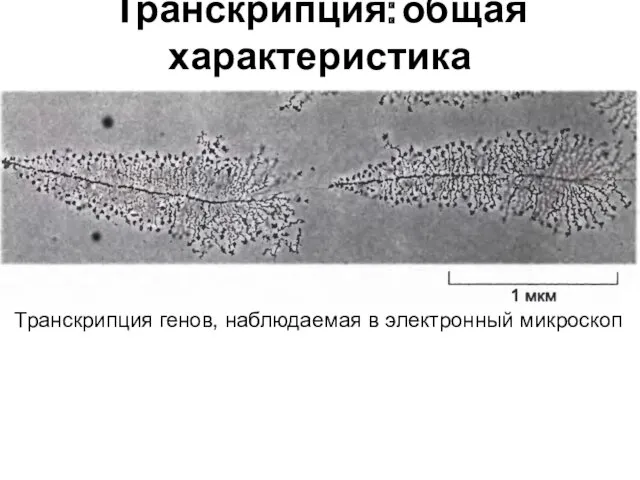
Транскрипция: общая характеристика
Транскрипция генов, наблюдаемая в электронный микроскоп
Транскрипция: общая характеристика
Транскрипция генов, наблюдаемая в электронный микроскоп
Транскрипция у прокариот
Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции
Транскрипция у прокариот
Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции
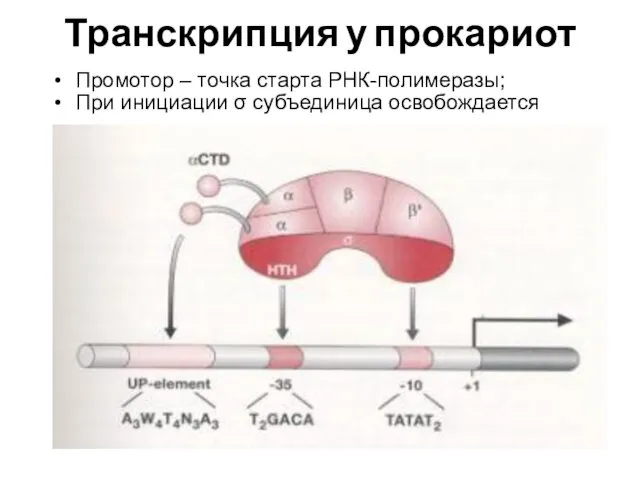
Транскрипция у прокариот
Промотор – точка старта РНК-полимеразы;
При инициации σ субъединица освобождается
Транскрипция у прокариот
Промотор – точка старта РНК-полимеразы;
При инициации σ субъединица освобождается
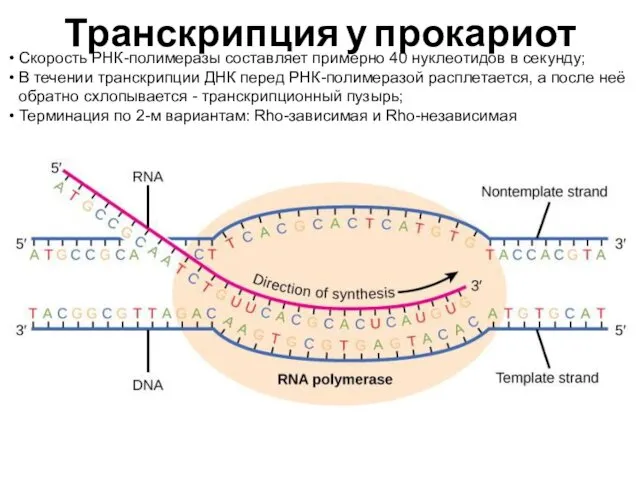
Транскрипция у прокариот
Скорость РНК-полимеразы составляет примерно 40 нуклеотидов в секунду;
В течении
Транскрипция у прокариот
Скорость РНК-полимеразы составляет примерно 40 нуклеотидов в секунду;
В течении
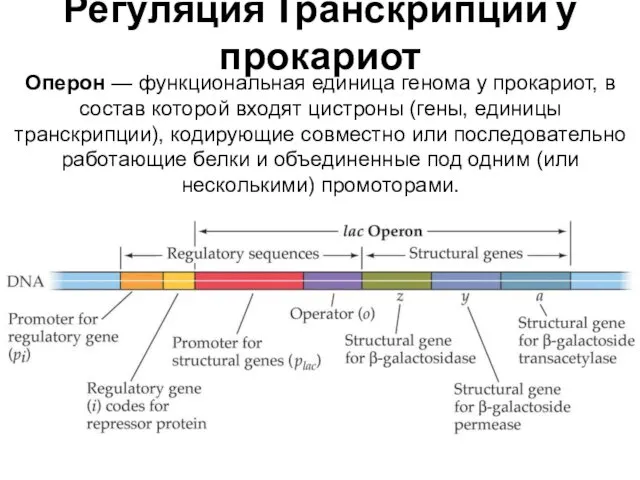
Регуляция Транскрипции у прокариот
Оперон — функциональная единица генома у прокариот, в
Регуляция Транскрипции у прокариот
Оперон — функциональная единица генома у прокариот, в
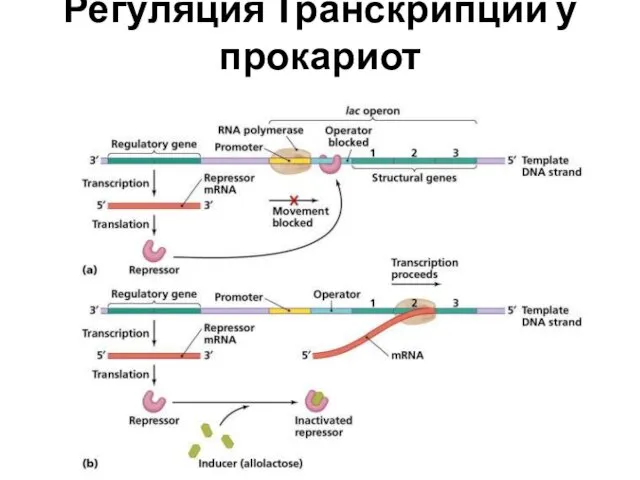
Регуляция Транскрипции у прокариот
Регуляция Транскрипции у прокариот

Транскрипция у эукариот
Транскрипция у эукариот происходит в ядре;
Синтез молекул РНК начинается
Транскрипция у эукариот
Транскрипция у эукариот происходит в ядре;
Синтез молекул РНК начинается
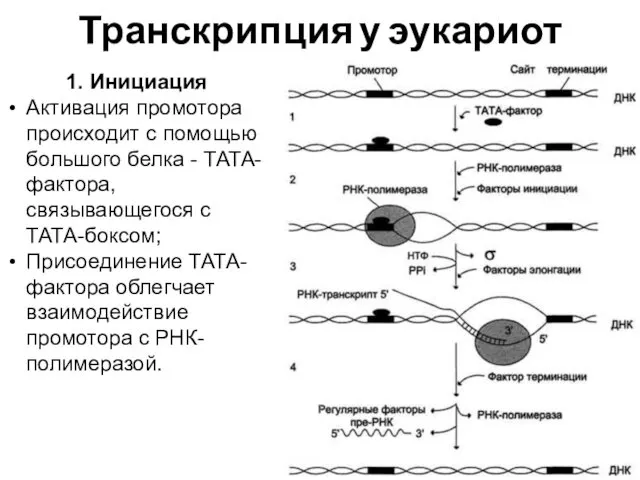
Транскрипция у эукариот
1. Инициация
Активация промотора происходит с помощью большого белка -
Транскрипция у эукариот
1. Инициация
Активация промотора происходит с помощью большого белка -
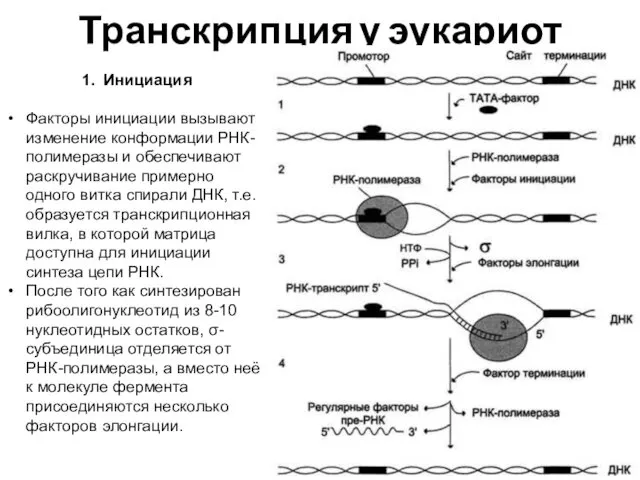
Транскрипция у эукариот
1. Инициация
Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают
Транскрипция у эукариот
1. Инициация
Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают
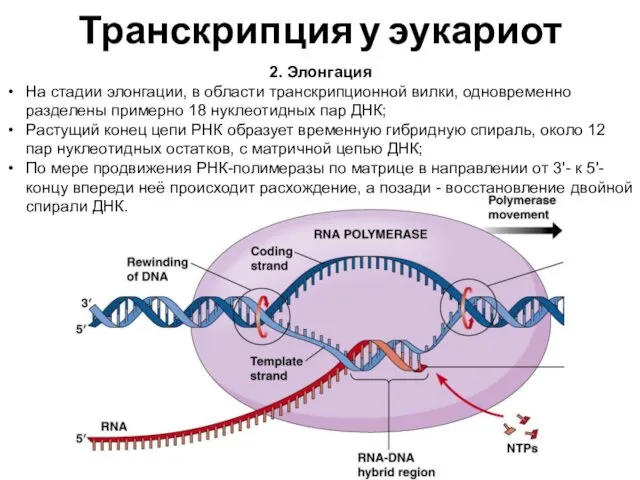
Транскрипция у эукариот
2. Элонгация
На стадии элонгации, в области транскрипционной вилки, одновременно
Транскрипция у эукариот
2. Элонгация
На стадии элонгации, в области транскрипционной вилки, одновременно
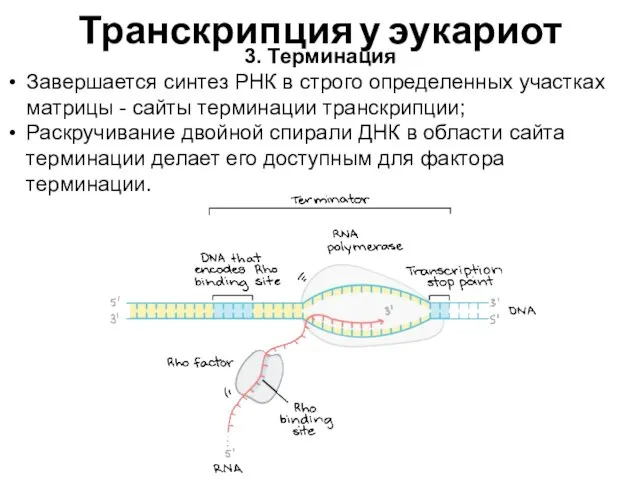
Транскрипция у эукариот
3. Терминация
Завершается синтез РНК в строго определенных участках матрицы
Транскрипция у эукариот
3. Терминация
Завершается синтез РНК в строго определенных участках матрицы

Факторы Транскрипции у эукариот
Регуляция базальной экспрессии генов;
Регуляция онтогенеза;
Ответ на изменение окружающей
Факторы Транскрипции у эукариот
Регуляция базальной экспрессии генов;
Регуляция онтогенеза;
Ответ на изменение окружающей
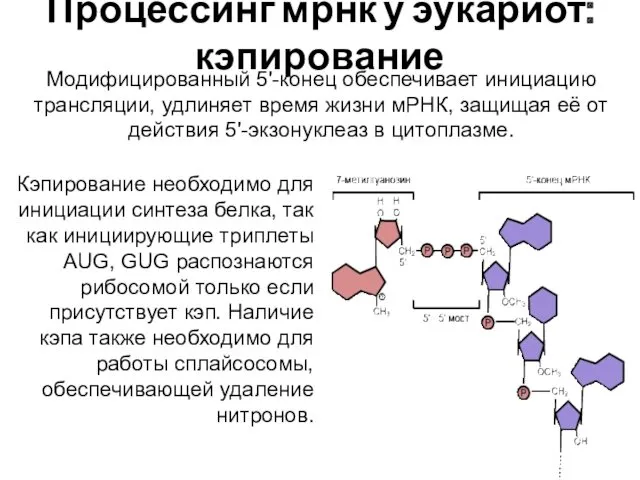
Процессинг мрнк у эукариот: кэпирование
Модифицированный 5'-конец обеспечивает инициацию трансляции, удлиняет время
Процессинг мрнк у эукариот: кэпирование
Модифицированный 5'-конец обеспечивает инициацию трансляции, удлиняет время

Процессинг мрнк у эукариот: полиаденилирование
Специальным ферментом полиА-полимеразой формируется полиА-последовательность (полиА-"хвост"), состоящая
Процессинг мрнк у эукариот: полиаденилирование
Специальным ферментом полиА-полимеразой формируется полиА-последовательность (полиА-"хвост"), состоящая
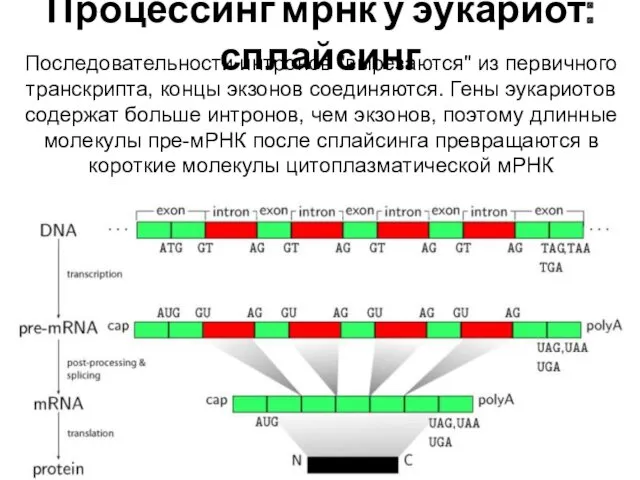
Процессинг мрнк у эукариот: сплайсинг
Последовательности интронов "вырезаются" из первичного транскрипта, концы
Процессинг мрнк у эукариот: сплайсинг
Последовательности интронов "вырезаются" из первичного транскрипта, концы
 Винсент Ван Гог
Винсент Ван Гог Дисциплина на улицах, дорогах и в транспорте - залог безопасности
Дисциплина на улицах, дорогах и в транспорте - залог безопасности Good health is above wealth
Good health is above wealth Отчёт-презентация на тему:Социально-личностное развитие дошкольников
Отчёт-презентация на тему:Социально-личностное развитие дошкольников Требования к фитнес-зонам
Требования к фитнес-зонам О работе Молодежного парламента при Совете депутатов муниципального образования Киясовский район
О работе Молодежного парламента при Совете депутатов муниципального образования Киясовский район Роль методического объединения классных руководителей в современной школе
Роль методического объединения классных руководителей в современной школе Производство банкнотной Бумаги
Производство банкнотной Бумаги Планировка участка по ремонту двигателей
Планировка участка по ремонту двигателей Испытание машин
Испытание машин Практическое занятие №1 по ТЭС и АЭС
Практическое занятие №1 по ТЭС и АЭС Защита программы
Защита программы Коралловая свадьба
Коралловая свадьба Ваша личная бизнес-модель профессионального развития
Ваша личная бизнес-модель профессионального развития Урок окружающего мира Как из зерна получилась булка
Урок окружающего мира Как из зерна получилась булка Видоизменения корней растений
Видоизменения корней растений Самопрезентация Самая лучшая ПРОФЕССИЯ!
Самопрезентация Самая лучшая ПРОФЕССИЯ! Отрочество – особая пора жизни
Отрочество – особая пора жизни Антитела. 6 курс
Антитела. 6 курс Отчет по производственной практике. Архитектура
Отчет по производственной практике. Архитектура Дигибридное скрещивание. Третий закон Г. Менделя
Дигибридное скрещивание. Третий закон Г. Менделя Онлайн собрание профоргов
Онлайн собрание профоргов Интеллектуальный марафон - 15 (2 класс)
Интеллектуальный марафон - 15 (2 класс) Влияние музыки на речевое развитие детей
Влияние музыки на речевое развитие детей Астана қаласының сәулет өнері
Астана қаласының сәулет өнері офтальмотренаж
офтальмотренаж Иоганн Себастьян Бах
Иоганн Себастьян Бах ОАО Брестский ликероводочный завод Белалко
ОАО Брестский ликероводочный завод Белалко