- Аэробное окисление углеводов

Содержание
- 2. Аэробное окисление углеводов
- 3. Содержание 1. «Аэробное окисление глюкозы» /дихотомический путь/ -окислительное декарбоксилирование пировиноградной кислоты; - цикл трикарбоновых кислот 2.
- 4. 1.Аэробное окисление глюкозы Валовое уравнение: C6H12O6 + 6O2=>6CO2 + 6H2O + W Полный выход энергии (W)
- 5. Глицерофосфатный челночный механизм. NADH2 NAD+ Цитозоль Цитоплазматическая глицерол 3-фосфат - ДГ Диоксиацетон фосфат Глицерол - 3
- 6. Внутренняя мембрана митохондрии Цитозоль Матрикс Mt малат малат оксалоацетат оксалоацетат NAD NAD NADH2 NADH2 глутамат глутамат
- 7. При гликолизе пировиноградная кислота восстанавливается и превращается в молочную кислоту – конечный продукт анаэробного обмена, а
- 8. Окислительное декарбоксилирование пирувата (Mt) пируват (ПВК) ТДФ
- 9. Амид липоевой кислоты или Окисленная форма Восстановленная форма
- 10. Тиаминдифосфат (ТДФ) или Е1-ТПФ Коэнзим А (HS-KoA) Пантотеновая кислота (вит. В3) 3’-фосфоаденозин- 5’-дифосфат
- 11. Стадии окислительного декарбоксилирования пирувата: Первая стадия катализируется пируватдегидрогеназой (Е1), коферментом служит ТДФ. В результате данной реакции
- 12. В третьей стадии ацетил-липоат (связанный с ферментым комплексом) взаимо- действует с коэнзимом А. Реакция катализируется ферментом
- 13. На четвертой стадии происходит окисление восстановленной липоевой кислоты до ее дисульфидной формы. Реакция катализируется ферментом дигидро-
- 14. Пируватдегидрогеназный комплекс СО2 Е3-ФАД NAD NADH2 Е1 – пируват-ДГ Е2 – дигидролипоил-ТФ Е3 – дигидролипоил-ДГ
- 15. Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальше окислению с образованием в конечном счете СО2 и
- 16. 3 этап. Цикл трикарбоновых кислот (цикл Кребса) Итак, образовавшийся в результате окислительного декарбоксилирования пирувата ацетил-КоА вступает
- 17. Цикл лимонной кислоты (цикл Кребса). Цис-аконитат Изоцитрат Сукцинил - КоА Сукцинат Цитрат Оксалоацетат Малат Фумарат 12
- 18. В первой реакции, катализируемой ферментом цитрат-синтазой, происходит конденсация ацетил-КоА с оксалоацетатом. В результате образуется лимонная кислота
- 19. Во второй реакции цикла образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты. Последняя, при- соединяя
- 20. В третьей реакции, которая , во-видимому, является лимитирующей реакцией цикла Кребса, происходит дегидрирование изолимонной кислоты в
- 21. В четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты до сукцинил-КоА. Механизм этой реакции сходен с реакцией
- 22. В пятой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту. Одновременно происходит
- 23. В шестой реакции янтарная кислота дегидрируется в фумаровую кислоту. Данная реакция катализируется сукцинатдегидрогеназой, в молекуле которой
- 24. В седьмой реакции образовавшаяся фумаровая кислота гидратируется под влиянием фермента фумаразы. Продуктом данной реакции является яблочная
- 25. В восьмой реакции цикла трикарбоновых кислот под влиянием митохондри- альной НАД-зависимой малатлегидрогеназы происходит окисление L-яблочной кислоты
- 26. образуется 3 молекулы АТФ (в процессе сопряженного окислительного фосфо- рилирования), а всего, следовательно, 9 молекул АТФ.
- 28. 2840 кДж до 50% ее аккумулируется в митохондриях в форме, которая может быть использована для выполнения
- 29. Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфат-дегидрогеназы и кофермента НАДФ+. 6-фосфоглюконо-лак- тон соединение нестабильное,
- 30. В следующей окислительной реакции, катализируемой 6-фосфоглюконат- дегидрогеназой (декарбоксилизирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная
- 31. Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза – ксилулозо-5-фосфат. Кроме того, рибу- лозо-5-фосфат
- 32. Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов:
- 33. Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седугептулозо-7-фосфата на глицеральдегид-3- фосфат:
- 34. Транскетолазная реакция в пентозном цикле встречается дважды, второй раз при образовании фруктозо-6-фосфата и триозофосфата в результате
- 36. Как видно, 6 молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, Образуют 6 молекул рибулозо-5-фосфата и 6 молекул
- 39. Скачать презентацию

Аэробное окисление углеводов
Аэробное окисление углеводов

Содержание
1. «Аэробное окисление глюкозы» /дихотомический путь/
-окислительное декарбоксилирование пировиноградной кислоты;
- цикл трикарбоновых
Содержание
1. «Аэробное окисление глюкозы» /дихотомический путь/
-окислительное декарбоксилирование пировиноградной кислоты;
- цикл трикарбоновых

1.Аэробное окисление глюкозы
Валовое уравнение: C6H12O6 + 6O2=>6CO2 + 6H2O + W
Полный
1.Аэробное окисление глюкозы
Валовое уравнение: C6H12O6 + 6O2=>6CO2 + 6H2O + W
Полный
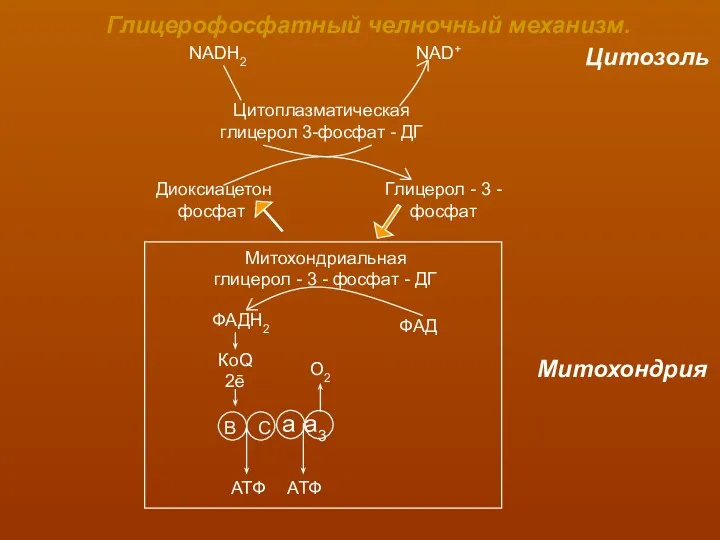
Глицерофосфатный челночный механизм.
NADH2
NAD+
Цитозоль
Цитоплазматическая
глицерол 3-фосфат - ДГ
Диоксиацетон
фосфат
Глицерол - 3 -
фосфат
Митохондриальная
глицерол
Глицерофосфатный челночный механизм.
NADH2
NAD+
Цитозоль
Цитоплазматическая
глицерол 3-фосфат - ДГ
Диоксиацетон
фосфат
Глицерол - 3 -
фосфат
Митохондриальная
глицерол
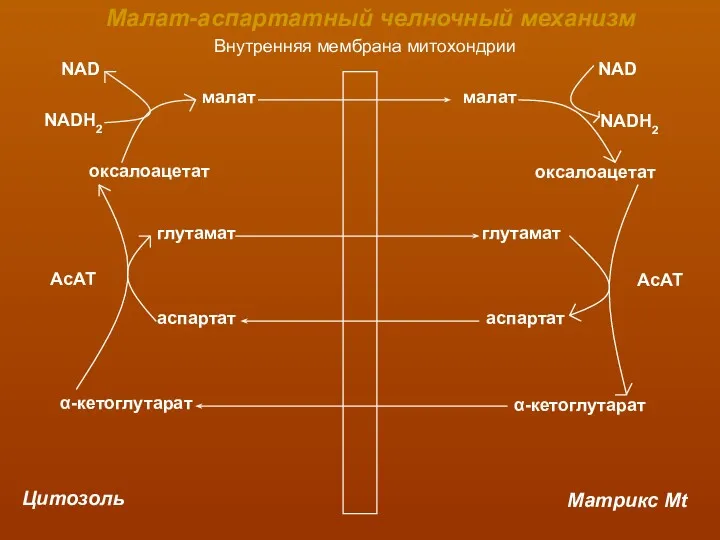
Внутренняя мембрана митохондрии
Цитозоль
Матрикс Mt
малат
малат
оксалоацетат
оксалоацетат
NAD
NAD
NADH2
NADH2
глутамат
глутамат
аспартат
аспартат
α-кетоглутарат
α-кетоглутарат
АсАТ
АсАТ
Малат-аспартатный челночный механизм
Внутренняя мембрана митохондрии
Цитозоль
Матрикс Mt
малат
малат
оксалоацетат
оксалоацетат
NAD
NAD
NADH2
NADH2
глутамат
глутамат
аспартат
аспартат
α-кетоглутарат
α-кетоглутарат
АсАТ
АсАТ
Малат-аспартатный челночный механизм
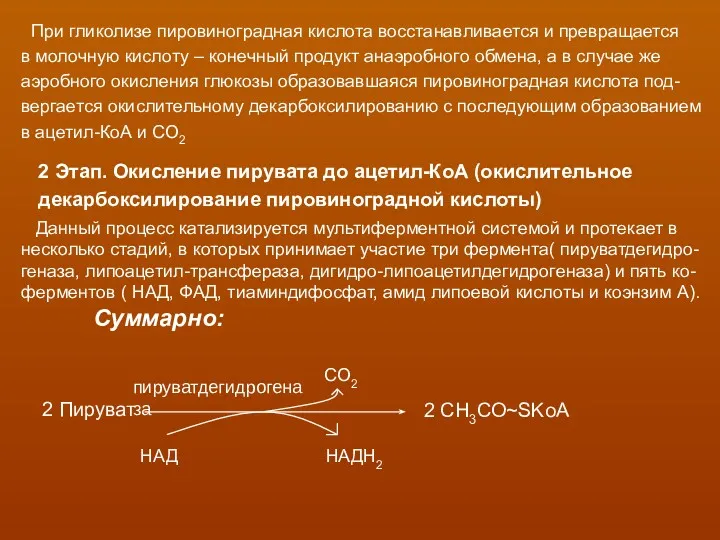
При гликолизе пировиноградная кислота восстанавливается и превращается
в молочную кислоту
При гликолизе пировиноградная кислота восстанавливается и превращается
в молочную кислоту
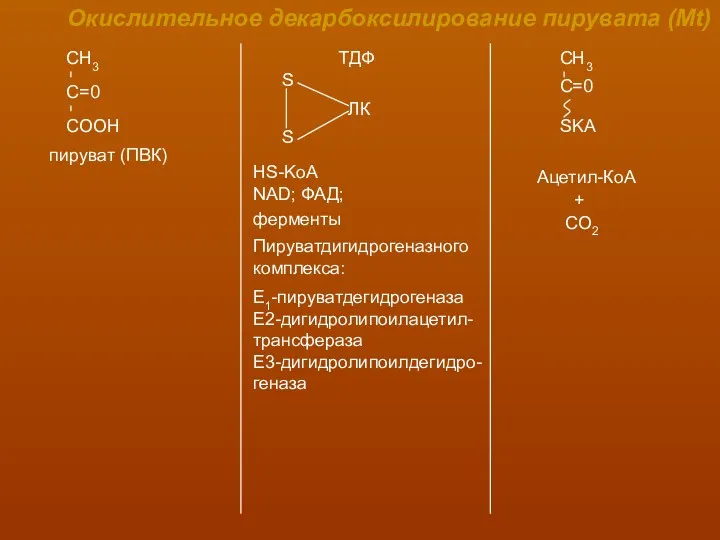
Окислительное декарбоксилирование пирувата (Mt)
пируват (ПВК)
ТДФ
Окислительное декарбоксилирование пирувата (Mt)
пируват (ПВК)
ТДФ
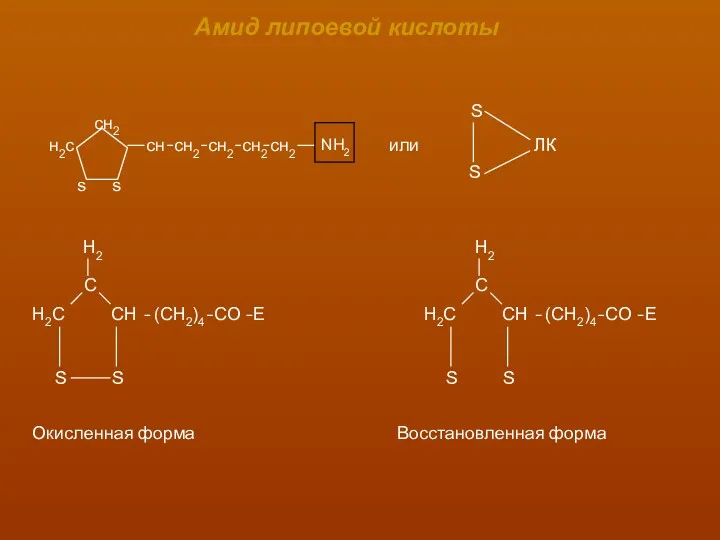
Амид липоевой кислоты
или
Окисленная форма
Восстановленная форма
Амид липоевой кислоты
или
Окисленная форма
Восстановленная форма
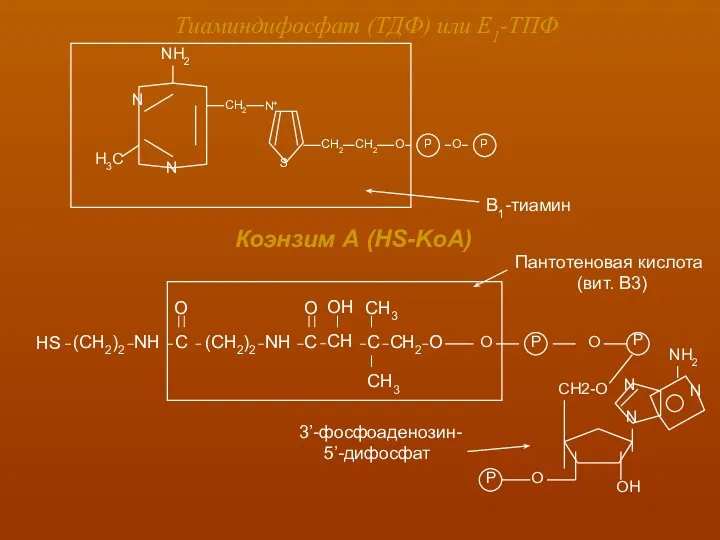
Тиаминдифосфат (ТДФ) или Е1-ТПФ
Коэнзим А (HS-KoA)
Пантотеновая кислота
(вит. В3)
3’-фосфоаденозин-
5’-дифосфат
Тиаминдифосфат (ТДФ) или Е1-ТПФ
Коэнзим А (HS-KoA)
Пантотеновая кислота
(вит. В3)
3’-фосфоаденозин-
5’-дифосфат

Стадии окислительного декарбоксилирования
пирувата:
Первая стадия катализируется пируватдегидрогеназой (Е1), коферментом служит
Стадии окислительного декарбоксилирования
пирувата:
Первая стадия катализируется пируватдегидрогеназой (Е1), коферментом служит
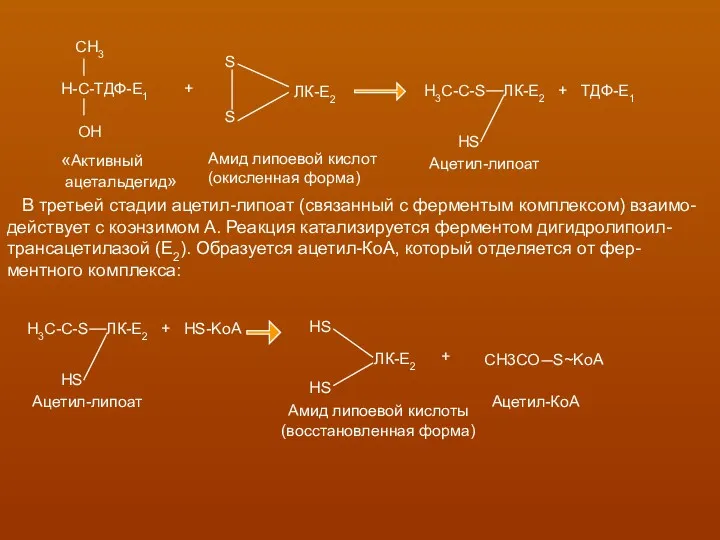
В третьей стадии ацетил-липоат (связанный с ферментым комплексом) взаимо-
действует с
В третьей стадии ацетил-липоат (связанный с ферментым комплексом) взаимо-
действует с

На четвертой стадии происходит окисление восстановленной липоевой кислоты
до ее дисульфидной
На четвертой стадии происходит окисление восстановленной липоевой кислоты
до ее дисульфидной
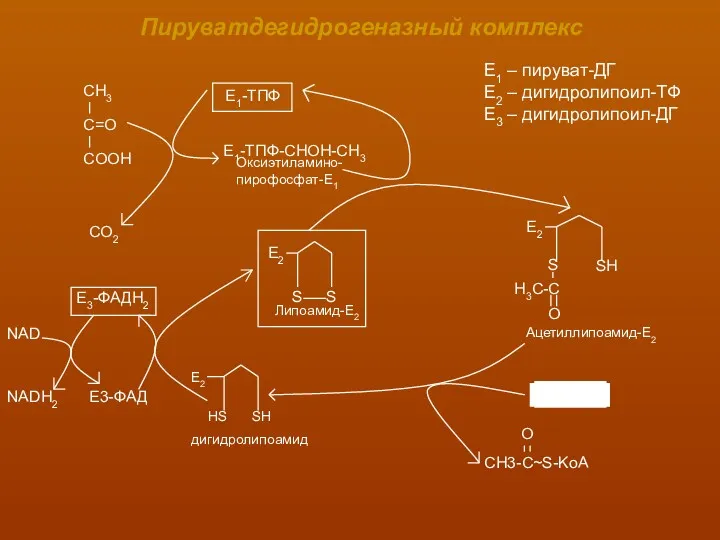
Пируватдегидрогеназный комплекс
СО2
Е3-ФАД
NAD
NADH2
Е1 – пируват-ДГ
Е2 – дигидролипоил-ТФ
Е3 – дигидролипоил-ДГ
Пируватдегидрогеназный комплекс
СО2
Е3-ФАД
NAD
NADH2
Е1 – пируват-ДГ
Е2 – дигидролипоил-ТФ
Е3 – дигидролипоил-ДГ
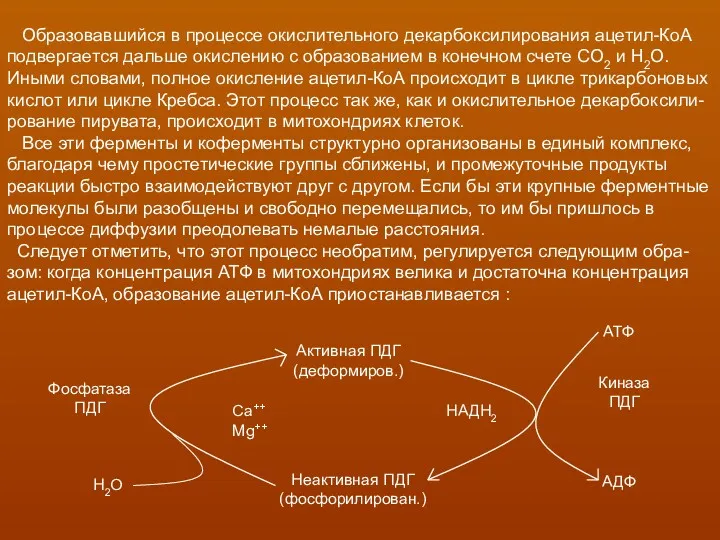
Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА
подвергается дальше окислению с образованием
Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА
подвергается дальше окислению с образованием

3 этап. Цикл трикарбоновых кислот (цикл Кребса)
Итак, образовавшийся в результате
3 этап. Цикл трикарбоновых кислот (цикл Кребса)
Итак, образовавшийся в результате
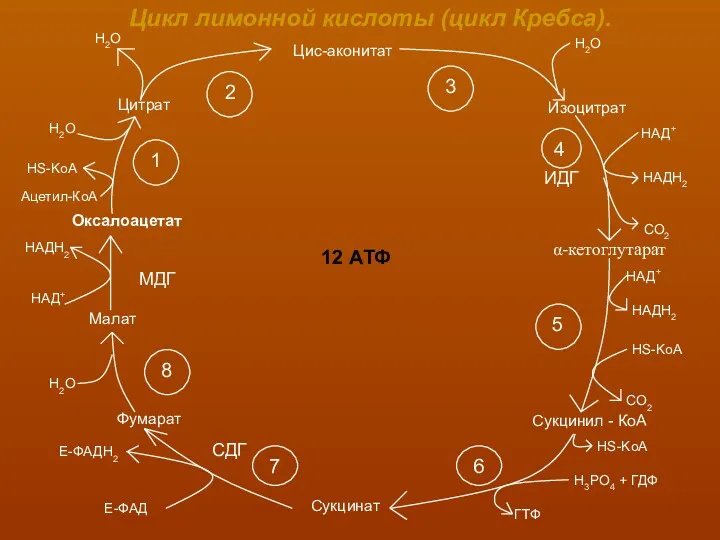
Цикл лимонной кислоты (цикл Кребса).
Цис-аконитат
Изоцитрат
Сукцинил - КоА
Сукцинат
Цитрат
Оксалоацетат
Малат
Фумарат
12 АТФ
МДГ
СДГ
ИДГ
α-кетоглутарат
Цикл лимонной кислоты (цикл Кребса).
Цис-аконитат
Изоцитрат
Сукцинил - КоА
Сукцинат
Цитрат
Оксалоацетат
Малат
Фумарат
12 АТФ
МДГ
СДГ
ИДГ
α-кетоглутарат

В первой реакции, катализируемой ферментом цитрат-синтазой, происходит
конденсация ацетил-КоА с
В первой реакции, катализируемой ферментом цитрат-синтазой, происходит
конденсация ацетил-КоА с
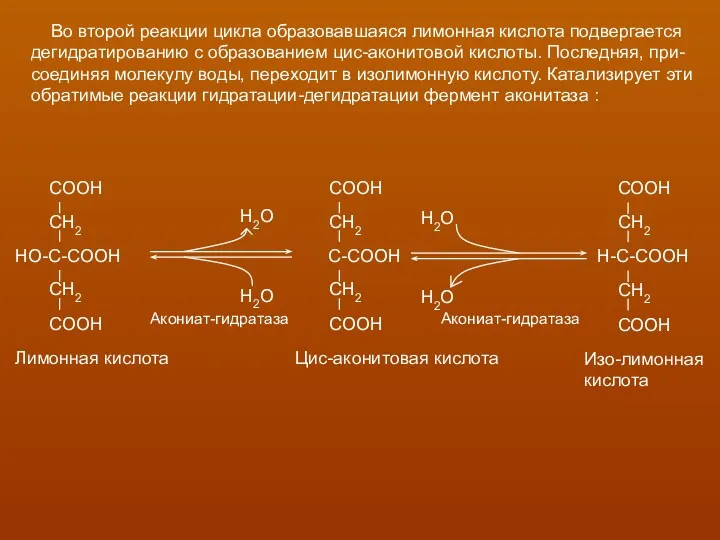
Во второй реакции цикла образовавшаяся лимонная кислота подвергается
дегидратированию с образованием
Во второй реакции цикла образовавшаяся лимонная кислота подвергается
дегидратированию с образованием

В третьей реакции, которая , во-видимому, является лимитирующей реакцией
цикла Кребса,
В третьей реакции, которая , во-видимому, является лимитирующей реакцией
цикла Кребса,
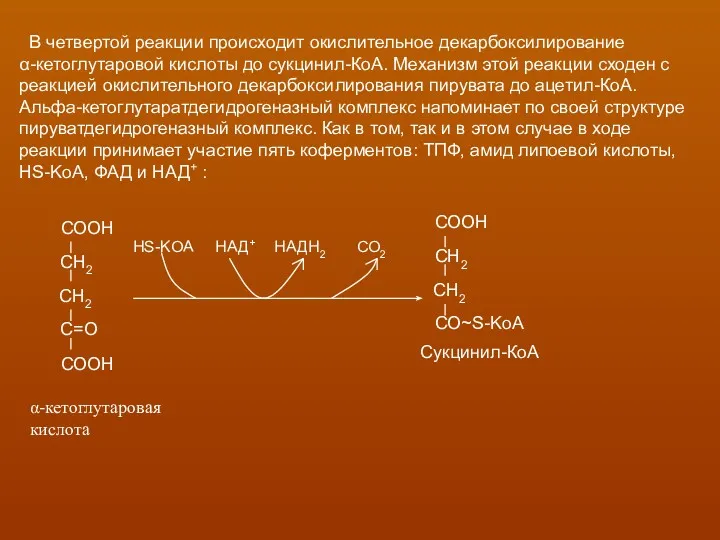
В четвертой реакции происходит окислительное декарбоксилирование
α-кетоглутаровой кислоты до сукцинил-КоА.
В четвертой реакции происходит окислительное декарбоксилирование
α-кетоглутаровой кислоты до сукцинил-КоА.

В пятой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата
превращается
В пятой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата
превращается
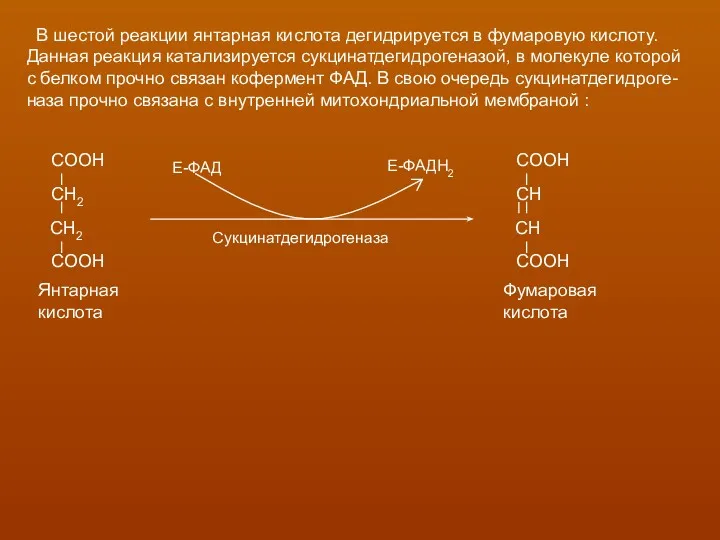
В шестой реакции янтарная кислота дегидрируется в фумаровую кислоту.
Данная реакция
В шестой реакции янтарная кислота дегидрируется в фумаровую кислоту.
Данная реакция
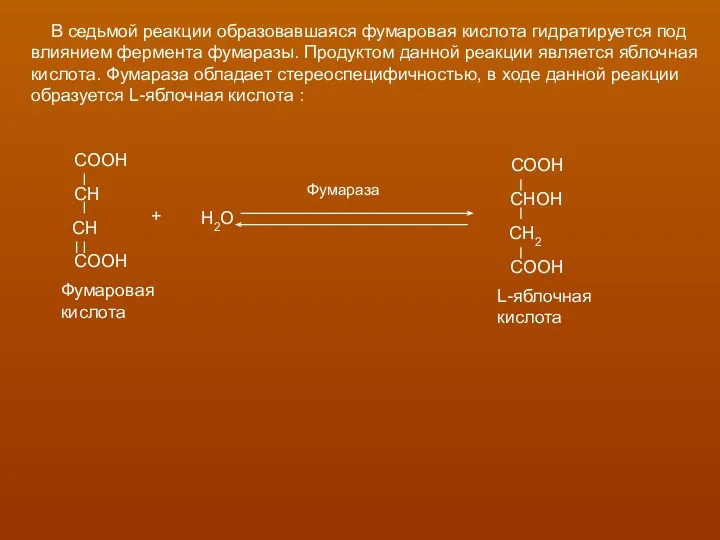
В седьмой реакции образовавшаяся фумаровая кислота гидратируется под
влиянием фермента
В седьмой реакции образовавшаяся фумаровая кислота гидратируется под
влиянием фермента

В восьмой реакции цикла трикарбоновых кислот под влиянием митохондри-
альной НАД-зависимой
В восьмой реакции цикла трикарбоновых кислот под влиянием митохондри-
альной НАД-зависимой

образуется 3 молекулы АТФ (в процессе сопряженного окислительного фосфо-
рилирования), а всего,
образуется 3 молекулы АТФ (в процессе сопряженного окислительного фосфо-
рилирования), а всего,
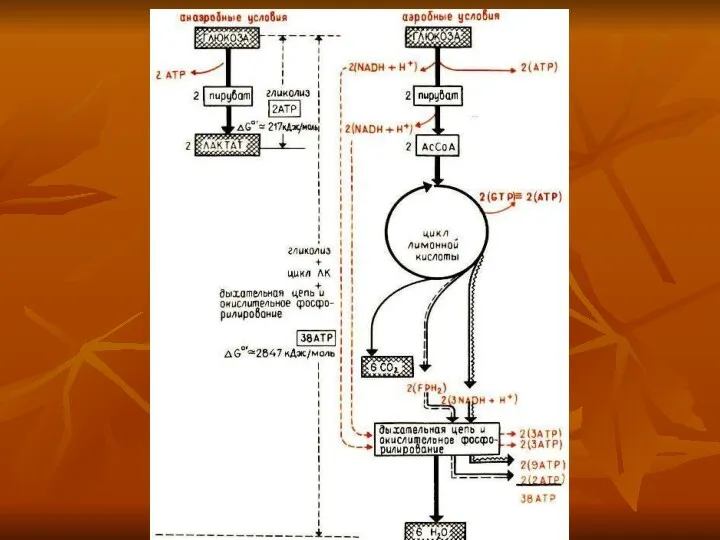

2840 кДж до 50% ее аккумулируется в митохондриях в форме, которая
2840 кДж до 50% ее аккумулируется в митохондриях в форме, которая
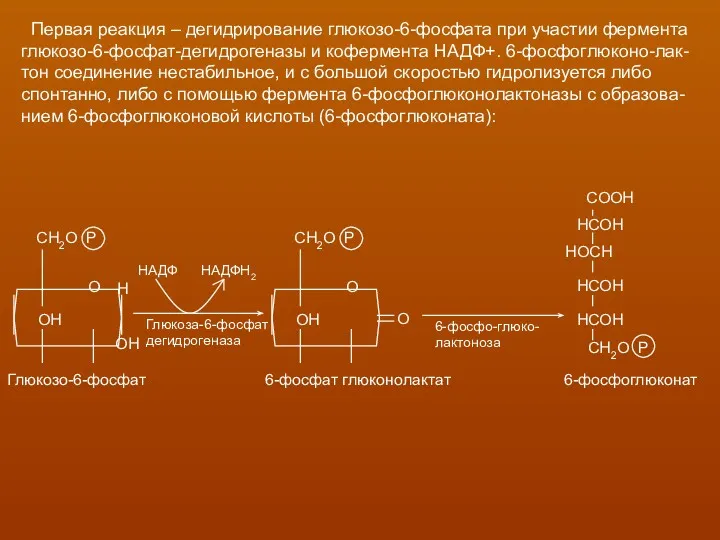
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента
глюкозо-6-фосфат-дегидрогеназы и
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента
глюкозо-6-фосфат-дегидрогеназы и
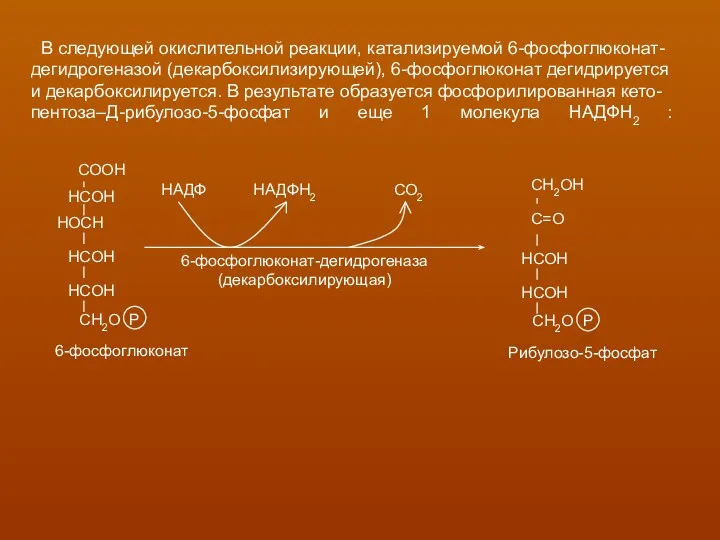
В следующей окислительной реакции, катализируемой 6-фосфоглюконат-
дегидрогеназой (декарбоксилизирующей), 6-фосфоглюконат дегидрируется
и
В следующей окислительной реакции, катализируемой 6-фосфоглюконат-
дегидрогеназой (декарбоксилизирующей), 6-фосфоглюконат дегидрируется
и
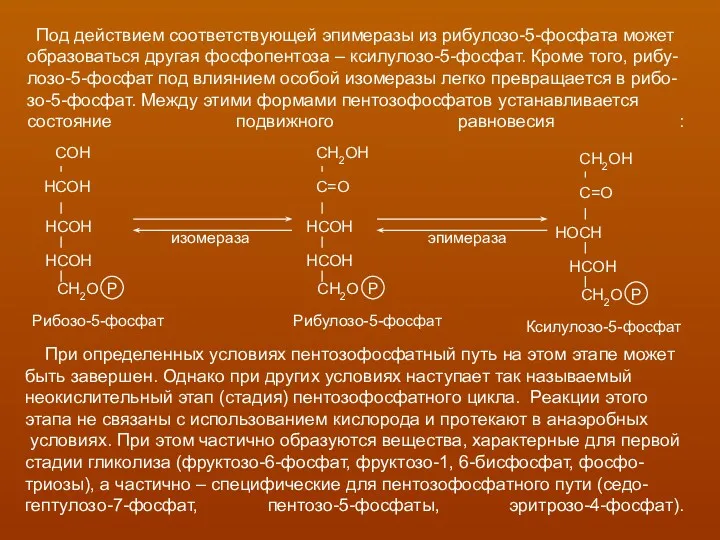
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может
образоваться другая фосфопентоза
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может
образоваться другая фосфопентоза
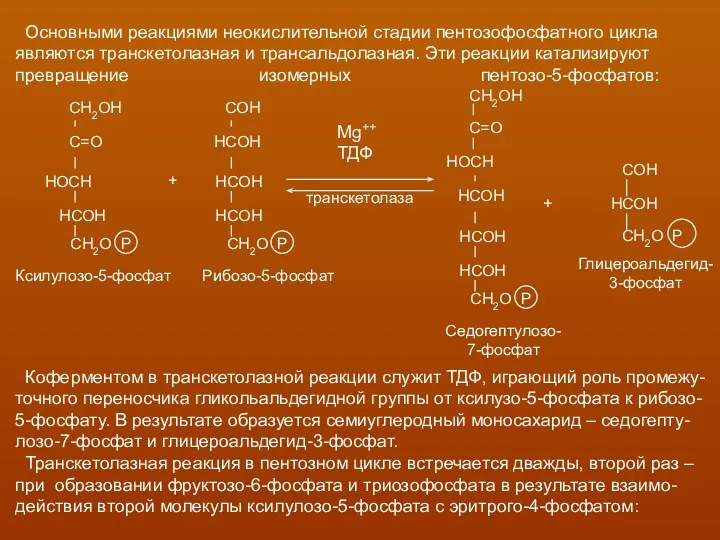
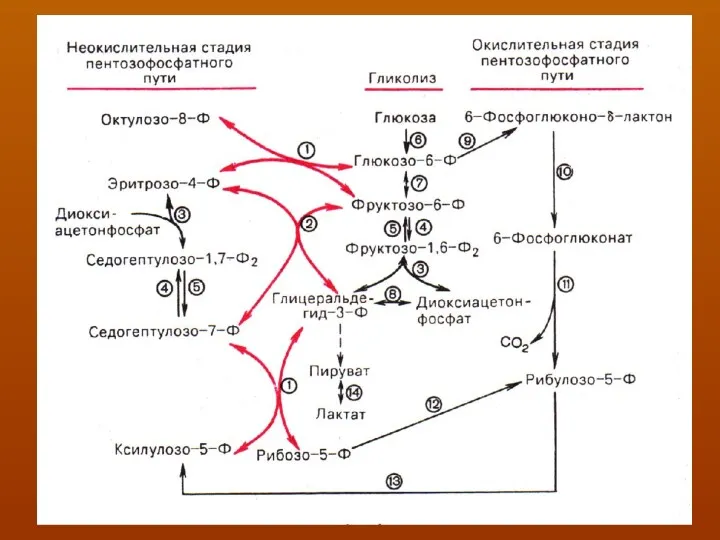
Основными реакциями неокислительной стадии пентозофосфатного цикла
являются транскетолазная и трансальдолазная. Эти
Основными реакциями неокислительной стадии пентозофосфатного цикла
являются транскетолазная и трансальдолазная. Эти
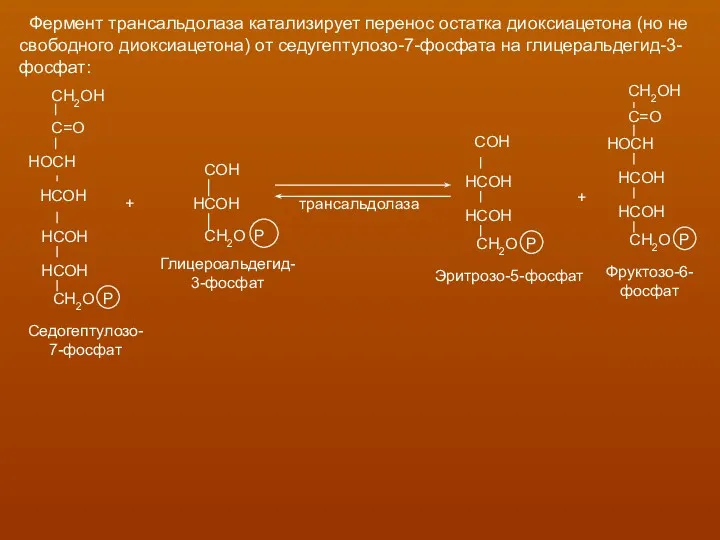
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не
свободного диоксиацетона)
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не
свободного диоксиацетона)
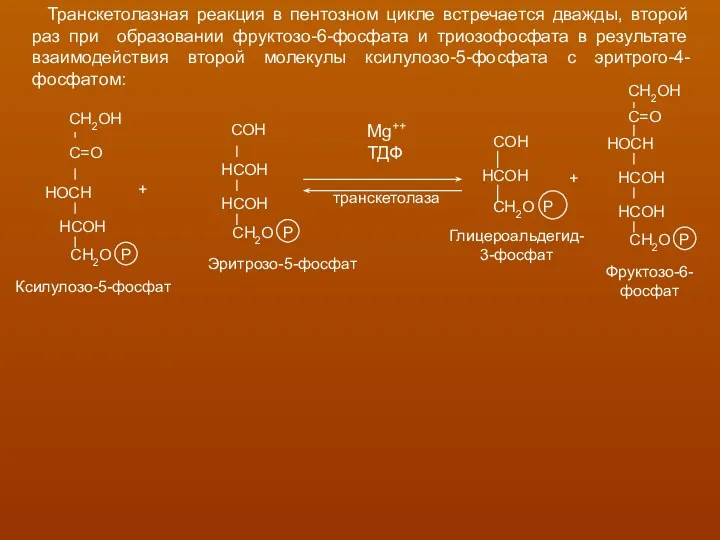
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз при
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз при
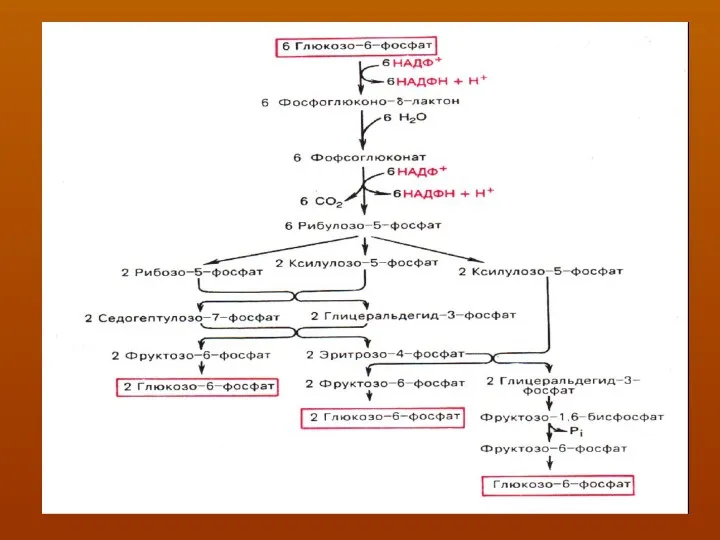

Как видно, 6 молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл,
Образуют
Как видно, 6 молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл,
Образуют
 Стебель. Побег
Стебель. Побег Structure, function and hygiene of teeth
Structure, function and hygiene of teeth Транспорт питательных веществ
Транспорт питательных веществ Общая характеристика нарушений голоса. История изучения
Общая характеристика нарушений голоса. История изучения Мини-викторина Естественные науки
Мини-викторина Естественные науки Жер бетінде тіршіліктің пайда болуы
Жер бетінде тіршіліктің пайда болуы Клеточная оболочка
Клеточная оболочка Витамины жирорастворимые и водорастворимые
Витамины жирорастворимые и водорастворимые Биосинтез белка
Биосинтез белка Клетка Беца коры больших полушарий
Клетка Беца коры больших полушарий Вегетативные органы растений. Корень. Лекция 5
Вегетативные органы растений. Корень. Лекция 5 Деление клеток. Митоз
Деление клеток. Митоз Организменный уровень. Тесты
Организменный уровень. Тесты Круги кровообращения
Круги кровообращения Кожные покровы хордовых
Кожные покровы хордовых Клеточная теория. Особенности строения клетки
Клеточная теория. Особенности строения клетки Сохранение природы средней полосы России от вторжения борщевика Сосновского
Сохранение природы средней полосы России от вторжения борщевика Сосновского Применение гербицидов. Лекция-6
Применение гербицидов. Лекция-6 Обитатели водоёмов
Обитатели водоёмов Генетические законы
Генетические законы викторина по биологии
викторина по биологии Динозавры жили на планете
Динозавры жили на планете Технология подготовки территория под создание цветника. Правила выращивания георгин различных сортов
Технология подготовки территория под создание цветника. Правила выращивания георгин различных сортов Класс млекопитающие. Общая характеристика класса
Класс млекопитающие. Общая характеристика класса Организация цитоплазмы
Организация цитоплазмы Половое и бесполое размножение организмов
Половое и бесполое размножение организмов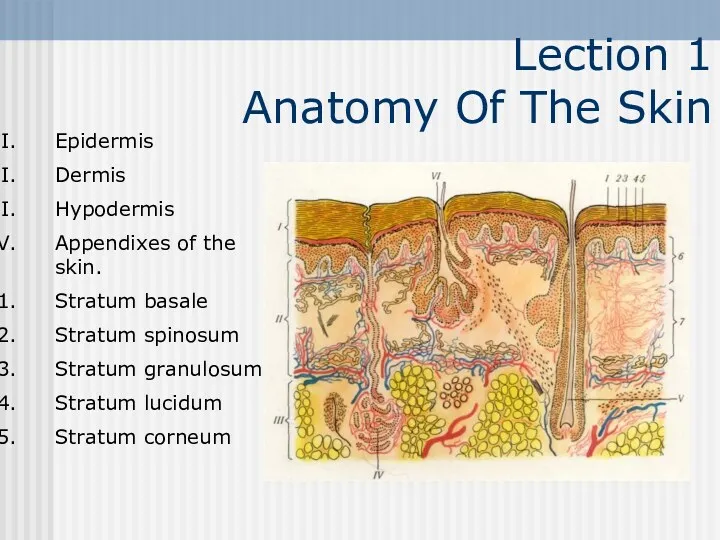 Anatomy Of The Skin. Lecture 1
Anatomy Of The Skin. Lecture 1 Отряд Хоботные
Отряд Хоботные