- Белки II. Уровни организации белков

Содержание
- 2. Первичная структура Вторичная структура Амино- кислоты α-спираль Третичная структура Четвертичная структура Глобула белка Несколько глобул белков
- 3. Простые белки – протеины. Состоят только из аминокислот а) альбумины б) глобулины в) гистоны г) протамины
- 4. Наиболее распространенные Альбумины Молекулярная масса – 15-70 тыс. ИЭТ – 4,7 (кислые) Мало аминокислоты глицина, но
- 5. Глобулины. Масса – 100 тыс. и выше ИЭТ - 5,5 – 7,3 Глицина в 3 раза
- 6. Гистоны. Белки ядра. Масса – 12 – 24 тыс. ИЭТ – 8 – 9 (основные) В
- 7. Белки растительного происхождения Глютелины. Белки зерен. ИЭТ - 6 -8 Пролина – 10 -15% Глютаминовой кислоты
- 8. Протеиноиды (склеропротеины). Белки опорных тканей (кости, хрящи, сухожилия, шерсть, копыта). Растворимость – нерастворимы. Много серосодержащих аминокислот.
- 9. Клетки Поперечный разрез волоса Макрофибрила Микрофибрила Протофибрила α-спираль Структура волоса
- 10. Исчерченность Головки тропоколлагеновых молекул Схема объединения 3-х х-цепей тропоколлагеновых молекул Структура коллагеновых волокон Исчерченность Головки тропоколлагеновых
- 11. Строение коллагеновой фибриллы
- 12. Один из типов поперечных связей между параллельными цепями коллагена
- 13. Молекулы тропоэластина, из которых формируется сеть связанных между собой полипептидных цепей эластина
- 15. Восстановление Окисление Искривление Схема завивки волос
- 16. Миозин и актин - два нитевидных белка сократительной системы
- 17. Сложные протеины. нуклеопротеины хромопротеины фосфоропротеины гликопротеины липопротеины металлопротеины
- 18. Нуклеопротеины. ↓ состоят из ↓ Протеины Нуклеиновые кислоты гистоны не гистоновые белки ДНК протамины РНК
- 19. Хромопротеины. Гемоглобин Миоглобин Цитохромы Гемоцианин Родопсин Каталаза Пероксидаза, другие
- 20. Хромопротеины. ↓ ↓ Протеин Небелковое вещество, обуславливающее окраску гемоглобин миоглобин цитохромы каталаза флавопротеины ретинопротеины
- 21. Миоглобин. Содержится в красных мышцах. Масса 17 тыс., 153 аминокислотных остатка + ГЕМ 75% образуют 8
- 22. Третичная структура миоглобина кашалота, установленная методом рентгеноструктурного анализа
- 23. Строение гемоглобина
- 24. Гемоглобин. Масса – 67 - 70 тыс. 96 % белка и 4 % ГЕМа от молекулярной
- 25. Трехмерная структура окси- и дезоксигемоглобина, установленная методом рентгеноструктурного анализа
- 26. Строение гема, входящего в состав миоглобина и гемоглобина
- 28. Транспорт газов. Гемоглобин связывает 4 молекулы кислорода. Оксигенирование гемоглобина сопровождается значительными конформационными изменениями: α/β пара поворачиваются
- 29. Соединения Нb Hb(Fe2+)О2 - оксигемоглобин; Hb (Fe2+) СО2 - карбогемоглобин; Hb (Fe2+) СО - карбоксигемоглобин; Hb
- 30. Схематическое изображение изменений в четвертичной структуре гемоглобина, обусловленных перемещением пары субъединиц α1β1 относительно неподвижной пары α2β2
- 31. Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
- 32. Перенос Н+ и СО2 с кровью. Эффект Бора
- 33. Фосфоропротеины. Белок + остаток фосфорной кислоты. Казеиноген Ововиттелин желтка Фосвитин Пепсин и др. белки-ферменты Ихтуллин икры
- 34. Гликопротеины. Представители: Фибриноген, муцины, мукоиды, церулоплазмин, иммуноглобулины. Белковая часть составляет 80 - 90% от м.м. молекул.
- 35. Белок + гетерополисахарид, присоединен ковалентно. Нейтральные – глобулины, фибриноген. Образуют нестойкие агрегаты. Кислые – муцин, мукоид.
- 36. Образование гликопротеинов
- 37. Протеогликаны Протеогликаны (мукополисахариды, гликозаминогликаны) – высокомолекулярные углеводно-белковые соединения. Образуют основную массу межклеточного матрикса соединительной ткани. Составляют
- 38. Протеогликан
- 39. Строение экстрацеллюлярного матрикса
- 40. Металлопротеины (металлосвязывающие белки). Белок + металл Способ удержания и защиты от осадка. Теряют токсичность. Обеспечивают ферментативную
- 41. Липопротеины плазмы крови
- 42. Общая характеристика липопротеинов Липиды в водной среде (а значит, и в крови) нерастворимы, поэтому для транспорта
- 43. Апопротеины Апопротеины выполняют несколько функций: • формируют структуру липопротеинов; • взаимодействуют с рецепторами на поверхности клеток
- 44. Липопротеины В организме синтезируются следующие типы липопротеинов: 1. хиломикроны (ХМ), 2. липопротеины очень низкой плотности (ЛПОНП),
- 45. Свойства липопротеинов ЛП хорошо растворимы в крови, не опалесцируют, так как имеют небольшой размер и отрицательный
- 46. Гиперхиломикронемия, гипертриглицеронемия После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно, гиперхиломикронемия, которая может продолжаться
- 47. Липопротеины — транспортные формы липидов Примечания: ФЛ — фосфолипиды; ХС — холестерол; ЭХС — эфиры холестерола;
- 48. Сравнение пространственной структуры эластазы и химотрипсина ЭЛАСТАЗА ХИМОТРИПСИН
- 49. Образования димера из одинаковых белковых субъединиц Субъединица Участок связывания Димер
- 50. Ленточная модель димера, образованного из двух идентичных субъединиц
- 51. Одинаковые субъединицы формируют спираль или кольцо Кольцо Спираль Участок связывания Участок связывания Субъединица Субъединица
- 52. Формирование спирали актина из субъединиц Спираль актина
- 53. Формирование пространственных структур из субъединиц белка Субъединица Спиральная трубка Гиксагонально Упакованный слой
- 55. Скачать презентацию
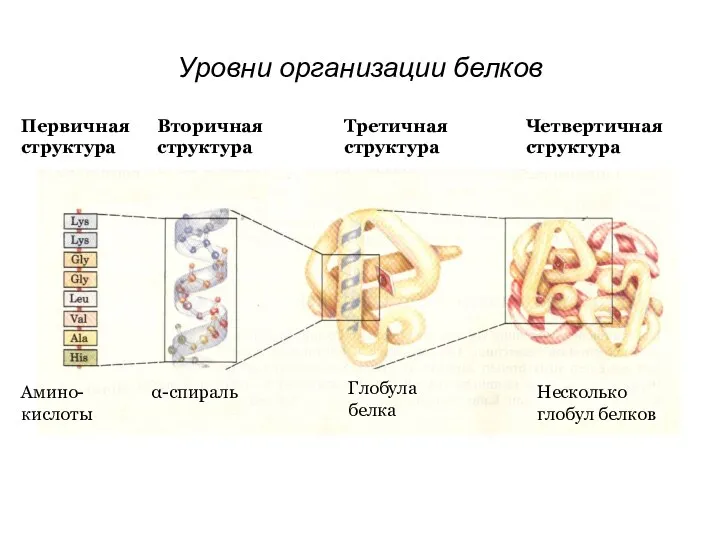
Первичная структура
Вторичная структура
Амино-
кислоты
α-спираль
Третичная структура
Четвертичная структура
Глобула белка
Несколько глобул белков
Уровни организации белков
Первичная структура
Вторичная структура
Амино-
кислоты
α-спираль
Третичная структура
Четвертичная структура
Глобула белка
Несколько глобул белков
Уровни организации белков

Простые белки – протеины.
Состоят только из аминокислот
а) альбумины
б) глобулины
в) гистоны
г) протамины
д)
Простые белки – протеины.
Состоят только из аминокислот
а) альбумины
б) глобулины
в) гистоны
г) протамины
д)

Наиболее распространенные
Альбумины
Молекулярная масса – 15-70 тыс.
ИЭТ – 4,7 (кислые)
Мало аминокислоты глицина,
Наиболее распространенные
Альбумины
Молекулярная масса – 15-70 тыс.
ИЭТ – 4,7 (кислые)
Мало аминокислоты глицина,

Глобулины.
Масса – 100 тыс. и выше
ИЭТ - 5,5 – 7,3
Глицина в
Глобулины.
Масса – 100 тыс. и выше
ИЭТ - 5,5 – 7,3
Глицина в

Гистоны.
Белки ядра.
Масса – 12 – 24 тыс.
ИЭТ – 8 –
Гистоны.
Белки ядра.
Масса – 12 – 24 тыс.
ИЭТ – 8 –

Белки растительного происхождения
Глютелины.
Белки зерен.
ИЭТ - 6 -8
Пролина – 10 -15%
Глютаминовой кислоты
Белки растительного происхождения
Глютелины.
Белки зерен.
ИЭТ - 6 -8
Пролина – 10 -15%
Глютаминовой кислоты

Протеиноиды (склеропротеины).
Белки опорных тканей (кости, хрящи, сухожилия, шерсть, копыта).
Растворимость – нерастворимы.
Много
Протеиноиды (склеропротеины).
Белки опорных тканей (кости, хрящи, сухожилия, шерсть, копыта).
Растворимость – нерастворимы.
Много
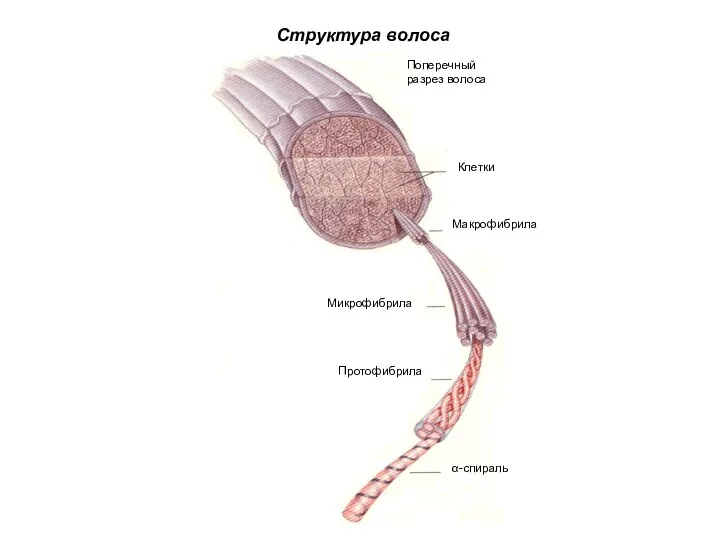
Клетки
Поперечный разрез волоса
Макрофибрила
Микрофибрила
Протофибрила
α-спираль
Структура волоса
Клетки
Поперечный разрез волоса
Макрофибрила
Микрофибрила
Протофибрила
α-спираль
Структура волоса

Исчерченность
Головки тропоколлагеновых молекул
Схема объединения 3-х х-цепей тропоколлагеновых молекул
Структура коллагеновых волокон
Исчерченность
Головки тропоколлагеновых
Исчерченность
Головки тропоколлагеновых молекул
Схема объединения 3-х х-цепей тропоколлагеновых молекул
Структура коллагеновых волокон
Исчерченность
Головки тропоколлагеновых
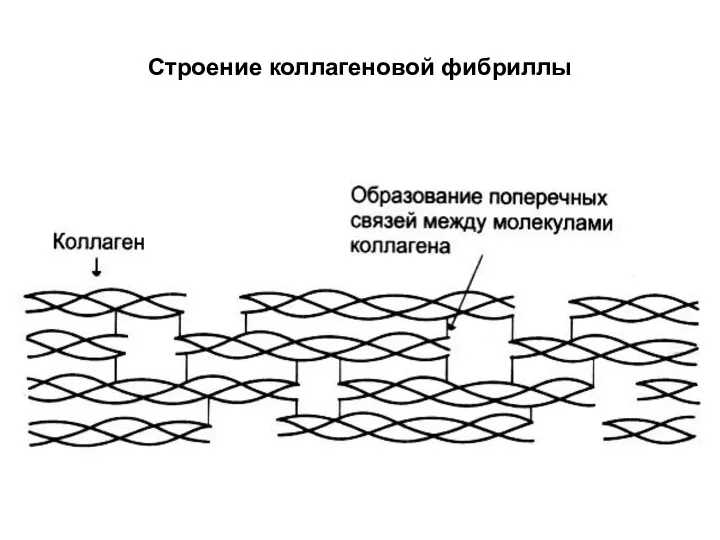
Строение коллагеновой фибриллы
Строение коллагеновой фибриллы
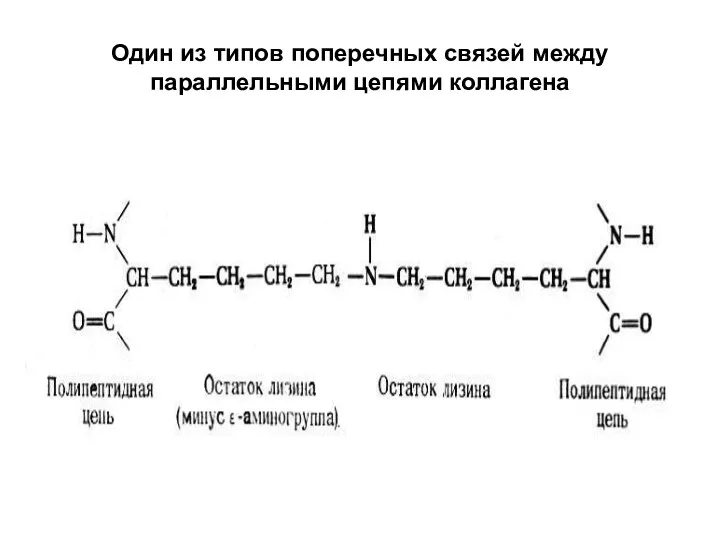
Один из типов поперечных связей между параллельными цепями коллагена
Один из типов поперечных связей между параллельными цепями коллагена
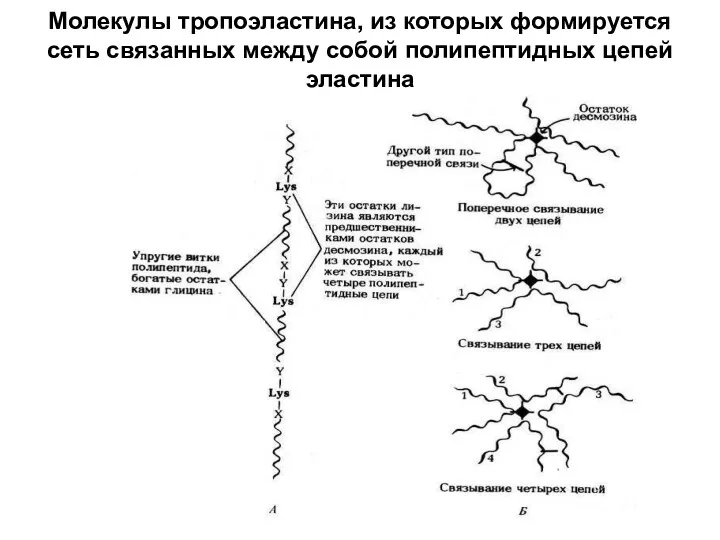
Молекулы тропоэластина, из которых формируется сеть связанных между собой полипептидных цепей
Молекулы тропоэластина, из которых формируется сеть связанных между собой полипептидных цепей

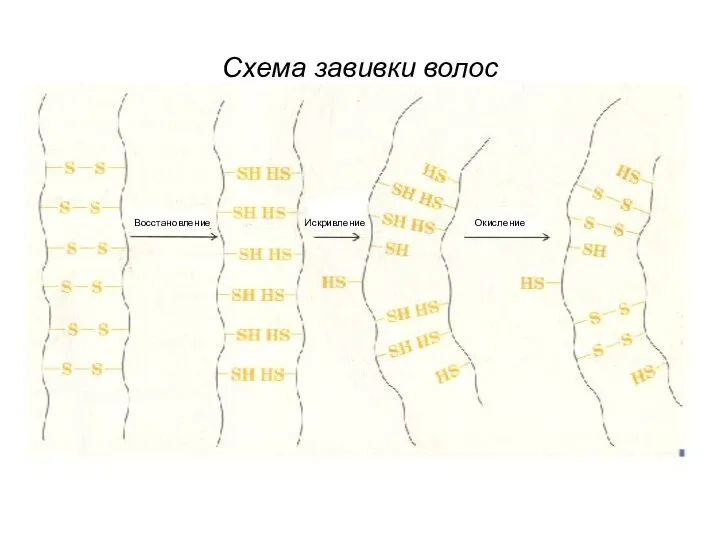
Восстановление
Окисление
Искривление
Схема завивки волос
Восстановление
Окисление
Искривление
Схема завивки волос
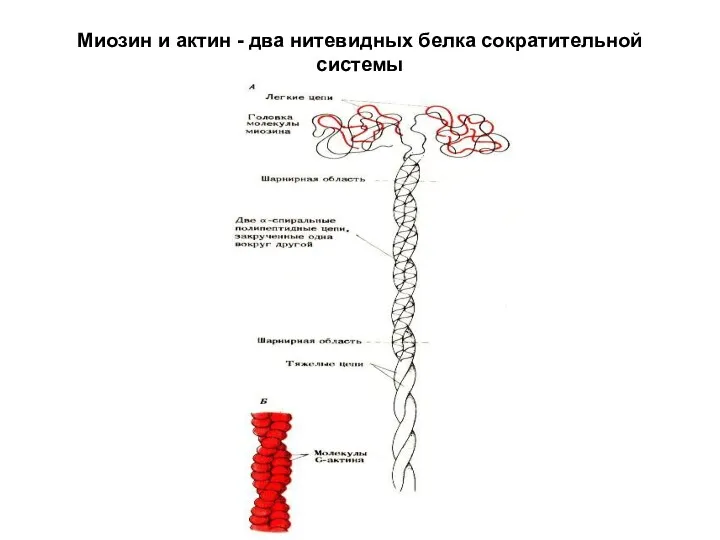
Миозин и актин - два нитевидных белка сократительной системы
Миозин и актин - два нитевидных белка сократительной системы

Сложные протеины.
нуклеопротеины
хромопротеины
фосфоропротеины
гликопротеины
липопротеины
металлопротеины
Сложные протеины.
нуклеопротеины
хромопротеины
фосфоропротеины
гликопротеины
липопротеины
металлопротеины
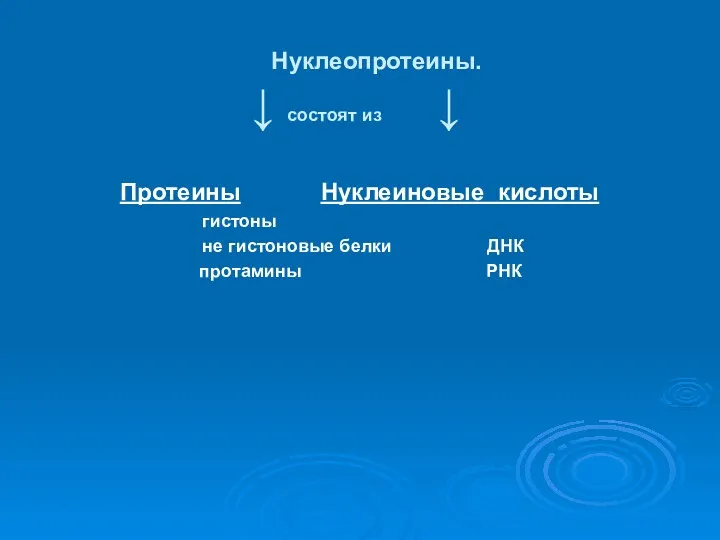
Нуклеопротеины.
↓ состоят из ↓
Протеины Нуклеиновые кислоты
гистоны
Нуклеопротеины.
↓ состоят из ↓
Протеины Нуклеиновые кислоты
гистоны

Хромопротеины.
Гемоглобин
Миоглобин
Цитохромы
Гемоцианин
Родопсин
Каталаза
Пероксидаза, другие
Хромопротеины.
Гемоглобин
Миоглобин
Цитохромы
Гемоцианин
Родопсин
Каталаза
Пероксидаза, другие

Хромопротеины.
↓ ↓
Протеин Небелковое вещество, обуславливающее окраску
гемоглобин
миоглобин цитохромы
каталаза
Хромопротеины.
↓ ↓
Протеин Небелковое вещество, обуславливающее окраску
гемоглобин
миоглобин цитохромы
каталаза

Миоглобин.
Содержится в красных мышцах.
Масса 17 тыс., 153 аминокислотных остатка + ГЕМ
75%
Миоглобин.
Содержится в красных мышцах.
Масса 17 тыс., 153 аминокислотных остатка + ГЕМ
75%
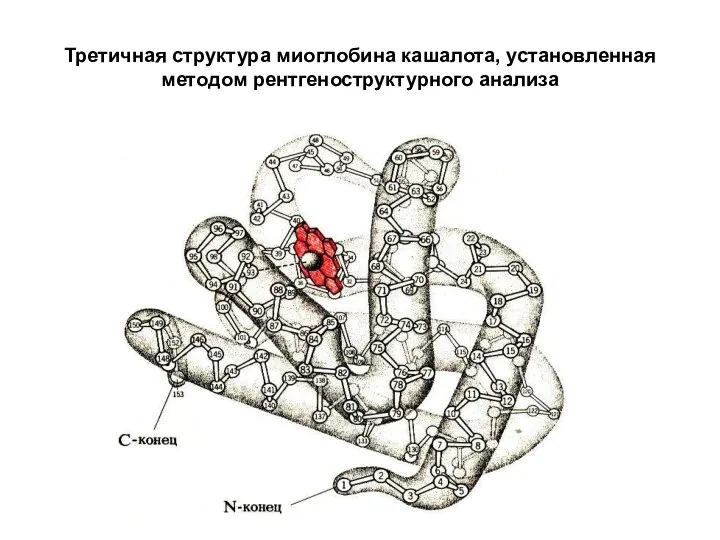
Третичная структура миоглобина кашалота, установленная методом рентгеноструктурного анализа
Третичная структура миоглобина кашалота, установленная методом рентгеноструктурного анализа
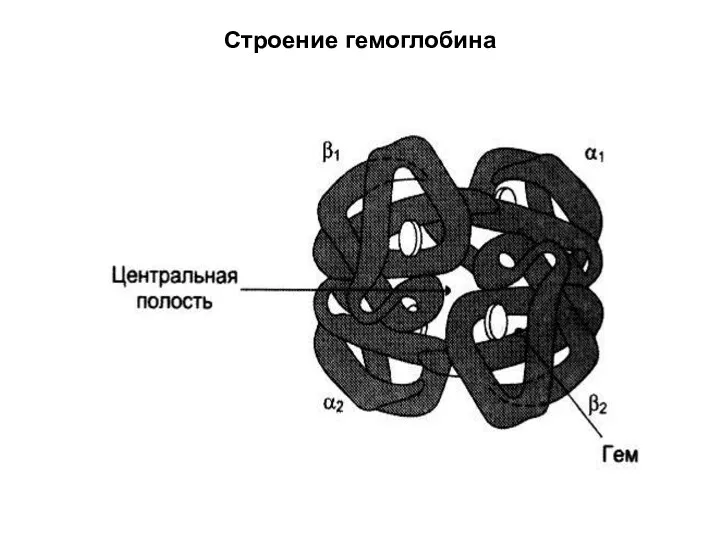
Строение гемоглобина
Строение гемоглобина

Гемоглобин.
Масса – 67 - 70 тыс.
96 % белка и 4 %
Гемоглобин.
Масса – 67 - 70 тыс.
96 % белка и 4 %
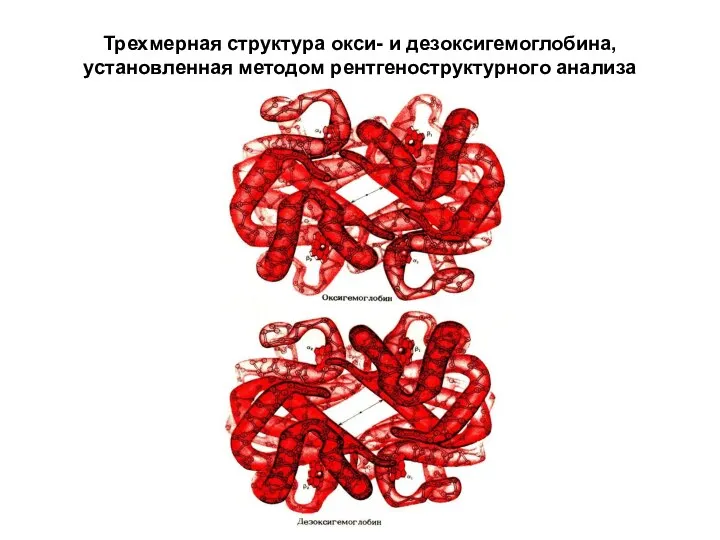
Трехмерная структура окси- и дезоксигемоглобина, установленная методом рентгеноструктурного анализа
Трехмерная структура окси- и дезоксигемоглобина, установленная методом рентгеноструктурного анализа
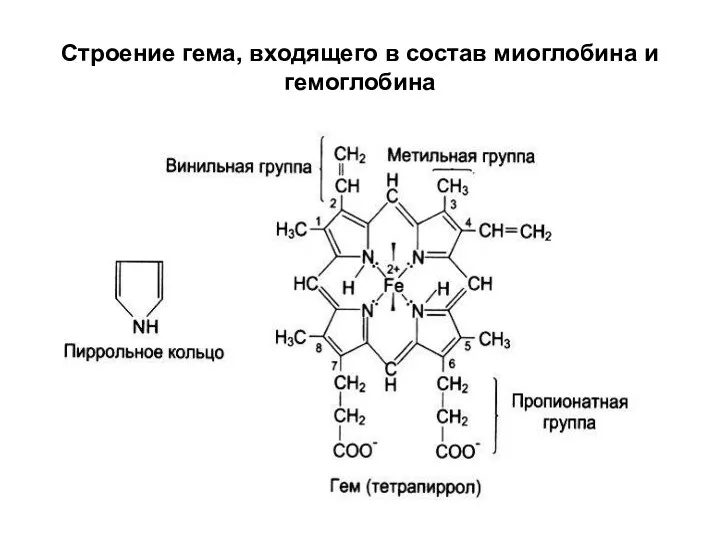
Строение гема, входящего в состав миоглобина и гемоглобина
Строение гема, входящего в состав миоглобина и гемоглобина
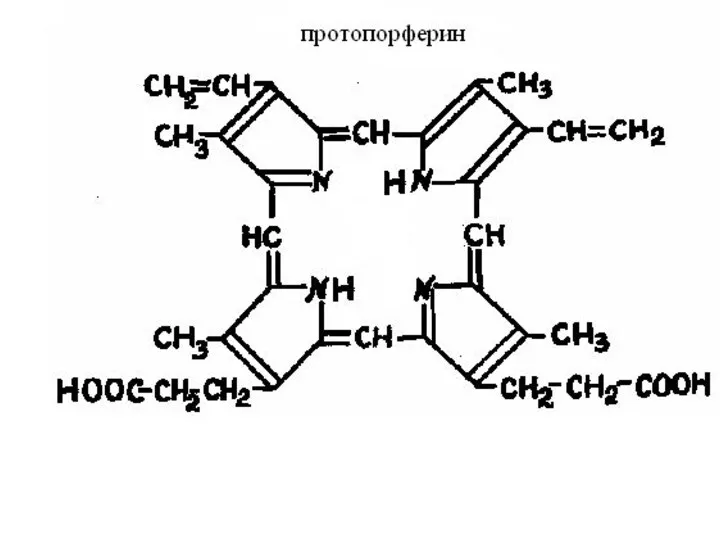

Транспорт газов.
Гемоглобин связывает 4 молекулы кислорода.
Оксигенирование гемоглобина сопровождается значительными конформационными изменениями:
Транспорт газов.
Гемоглобин связывает 4 молекулы кислорода.
Оксигенирование гемоглобина сопровождается значительными конформационными изменениями:

Соединения Нb
Hb(Fe2+)О2 - оксигемоглобин;
Hb (Fe2+) СО2 - карбогемоглобин;
Hb (Fe2+) СО -
Соединения Нb
Hb(Fe2+)О2 - оксигемоглобин;
Hb (Fe2+) СО2 - карбогемоглобин;
Hb (Fe2+) СО -
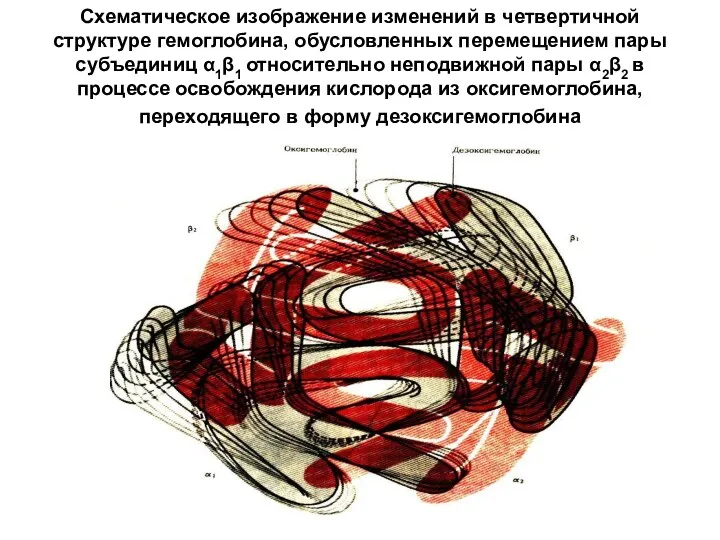
Схематическое изображение изменений в четвертичной структуре гемоглобина, обусловленных перемещением пары субъединиц
Схематическое изображение изменений в четвертичной структуре гемоглобина, обусловленных перемещением пары субъединиц
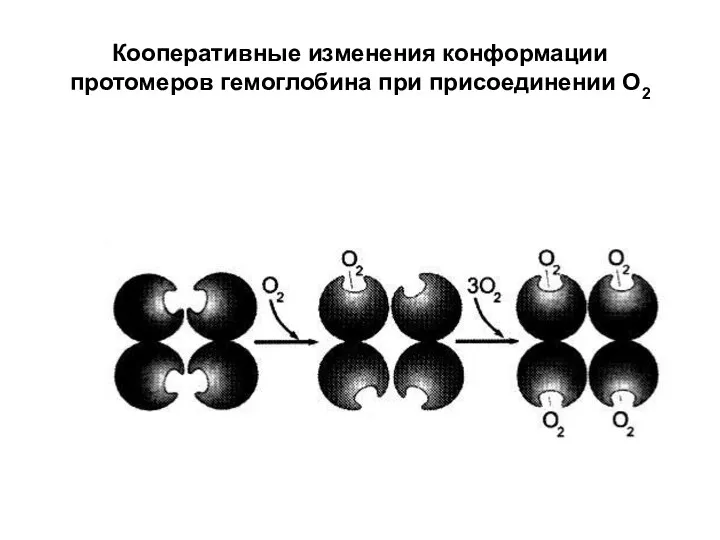
Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
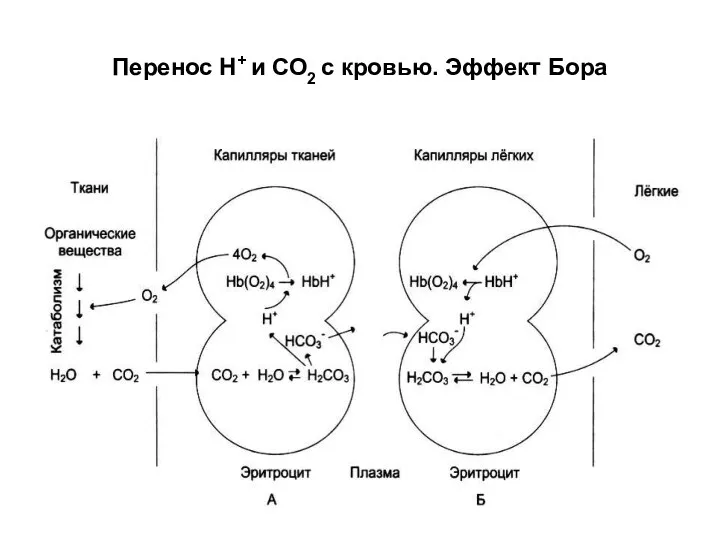
Перенос Н+ и СО2 с кровью. Эффект Бора
Перенос Н+ и СО2 с кровью. Эффект Бора

Фосфоропротеины.
Белок + остаток фосфорной кислоты.
Казеиноген
Ововиттелин желтка
Фосвитин
Пепсин и др. белки-ферменты
Ихтуллин икры
Фосфоропротеины.
Белок + остаток фосфорной кислоты.
Казеиноген
Ововиттелин желтка
Фосвитин
Пепсин и др. белки-ферменты
Ихтуллин икры

Гликопротеины.
Представители: Фибриноген, муцины, мукоиды, церулоплазмин, иммуноглобулины.
Белковая часть составляет 80 - 90%
Гликопротеины.
Представители: Фибриноген, муцины, мукоиды, церулоплазмин, иммуноглобулины.
Белковая часть составляет 80 - 90%

Белок + гетерополисахарид, присоединен ковалентно.
Нейтральные – глобулины, фибриноген. Образуют нестойкие агрегаты.
Кислые
Белок + гетерополисахарид, присоединен ковалентно.
Нейтральные – глобулины, фибриноген. Образуют нестойкие агрегаты.
Кислые

Образование гликопротеинов
Образование гликопротеинов

Протеогликаны
Протеогликаны (мукополисахариды, гликозаминогликаны) – высокомолекулярные углеводно-белковые соединения. Образуют основную массу межклеточного
Протеогликаны
Протеогликаны (мукополисахариды, гликозаминогликаны) – высокомолекулярные углеводно-белковые соединения. Образуют основную массу межклеточного
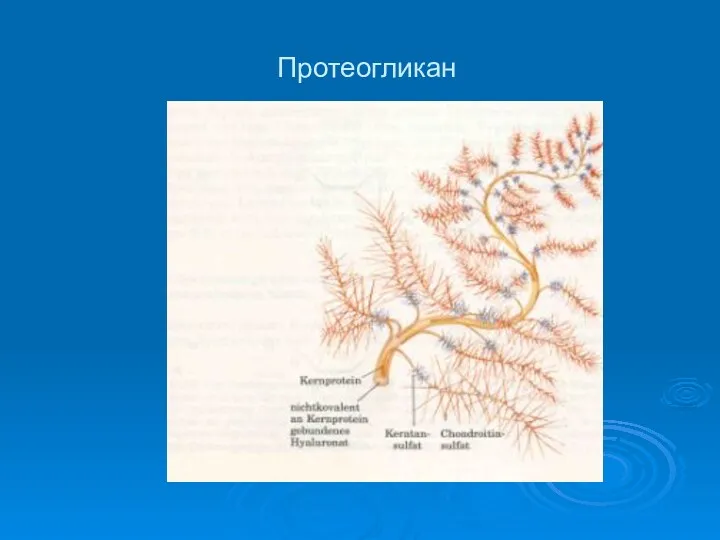
Протеогликан
Протеогликан
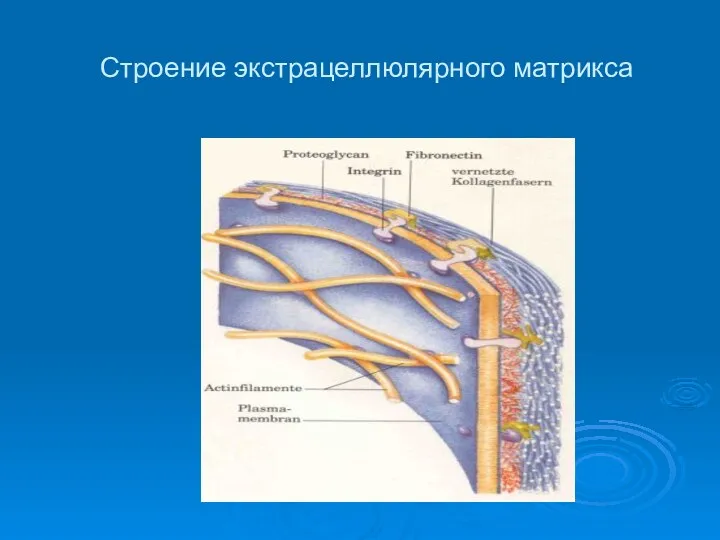
Строение экстрацеллюлярного матрикса
Строение экстрацеллюлярного матрикса

Металлопротеины (металлосвязывающие белки).
Белок + металл
Способ удержания и защиты от осадка.
Теряют токсичность.
Металлопротеины (металлосвязывающие белки).
Белок + металл
Способ удержания и защиты от осадка.
Теряют токсичность.
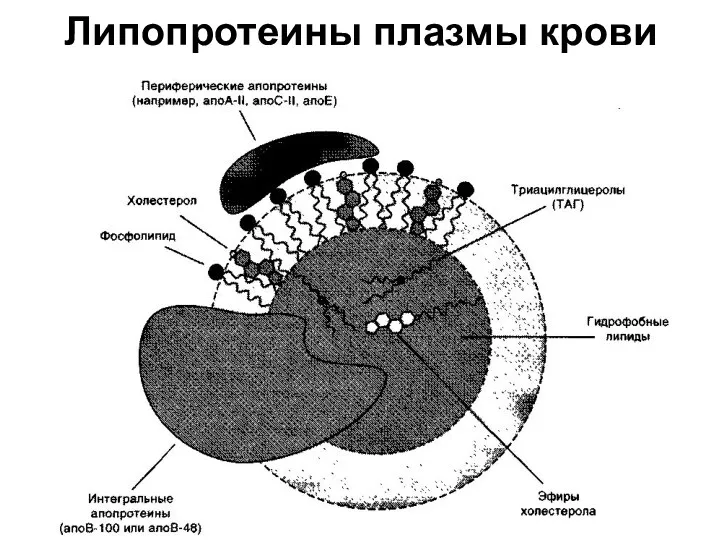
Липопротеины плазмы крови
Липопротеины плазмы крови

Общая характеристика липопротеинов
Липиды в водной среде (а значит, и в крови)
Общая характеристика липопротеинов
Липиды в водной среде (а значит, и в крови)

Апопротеины
Апопротеины выполняют несколько функций:
• формируют структуру липопротеинов;
• взаимодействуют с рецепторами на
Апопротеины
Апопротеины выполняют несколько функций:
• формируют структуру липопротеинов;
• взаимодействуют с рецепторами на

Липопротеины
В организме синтезируются следующие типы липопротеинов: 1. хиломикроны (ХМ), 2. липопротеины
Липопротеины
В организме синтезируются следующие типы липопротеинов: 1. хиломикроны (ХМ), 2. липопротеины

Свойства липопротеинов
ЛП хорошо растворимы в крови, не опалесцируют, так как имеют
Свойства липопротеинов
ЛП хорошо растворимы в крови, не опалесцируют, так как имеют

Гиперхиломикронемия, гипертриглицеронемия
После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно,
Гиперхиломикронемия, гипертриглицеронемия
После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно,
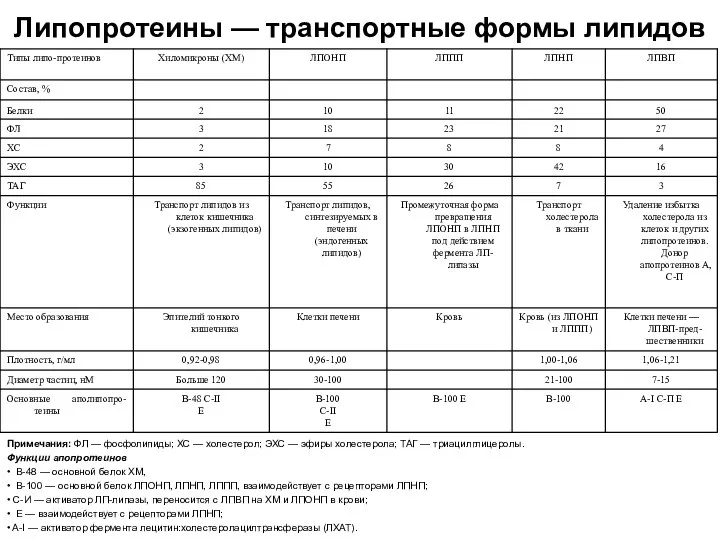
Липопротеины — транспортные формы липидов
Примечания: ФЛ — фосфолипиды; ХС — холестерол;
Липопротеины — транспортные формы липидов
Примечания: ФЛ — фосфолипиды; ХС — холестерол;
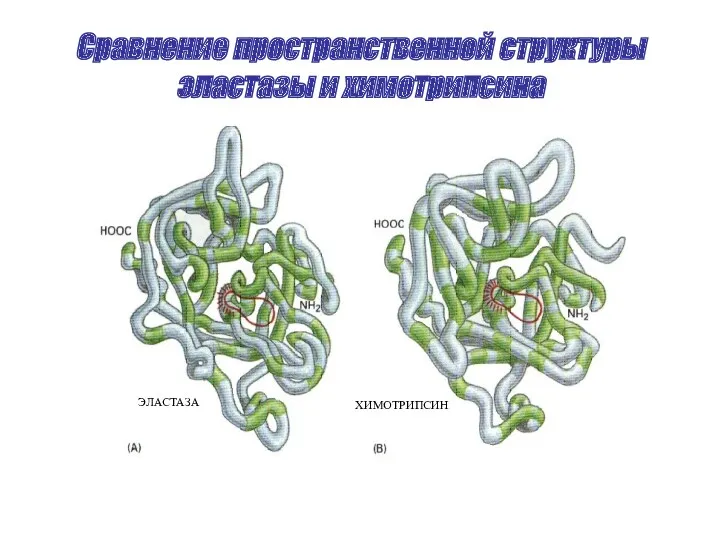
Сравнение пространственной структуры эластазы и химотрипсина
ЭЛАСТАЗА
ХИМОТРИПСИН
Сравнение пространственной структуры эластазы и химотрипсина
ЭЛАСТАЗА
ХИМОТРИПСИН
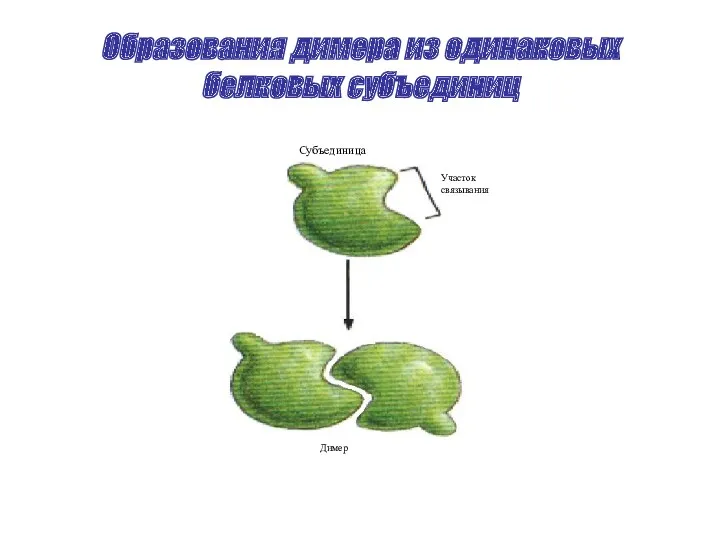
Образования димера из одинаковых белковых субъединиц
Субъединица
Участок
связывания
Димер
Образования димера из одинаковых белковых субъединиц
Субъединица
Участок
связывания
Димер
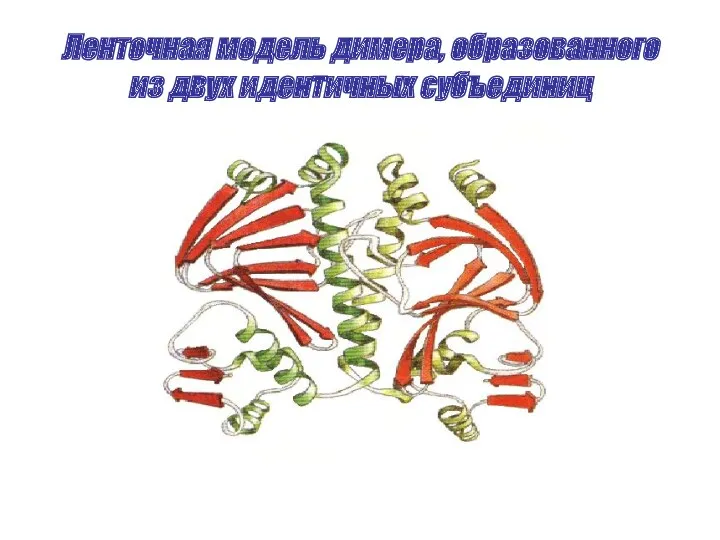
Ленточная модель димера, образованного из двух идентичных субъединиц
Ленточная модель димера, образованного из двух идентичных субъединиц
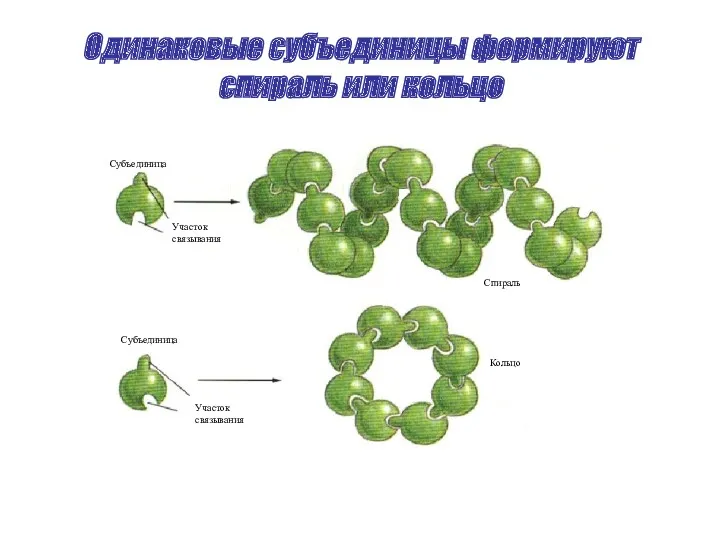
Одинаковые субъединицы формируют спираль или кольцо
Кольцо
Спираль
Участок
связывания
Участок
связывания
Субъединица
Субъединица
Одинаковые субъединицы формируют спираль или кольцо
Кольцо
Спираль
Участок
связывания
Участок
связывания
Субъединица
Субъединица

Формирование спирали актина из субъединиц
Спираль актина
Формирование спирали актина из субъединиц
Спираль актина
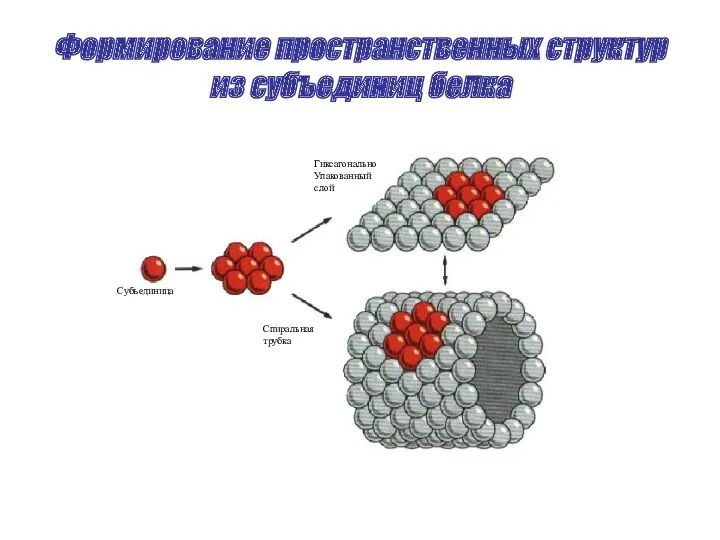
Формирование пространственных структур из субъединиц белка
Субъединица
Спиральная
трубка
Гиксагонально
Упакованный
слой
Формирование пространственных структур из субъединиц белка
Субъединица
Спиральная
трубка
Гиксагонально
Упакованный
слой
 Семейство Паслёновые и Бобовые, биология, 6 класс
Семейство Паслёновые и Бобовые, биология, 6 класс Основы селекции. Работы Н.И. Вавилова
Основы селекции. Работы Н.И. Вавилова Ветеринарные препараты, инструменты и кормовые добавки
Ветеринарные препараты, инструменты и кормовые добавки Макроэволюция: направления и пути эволюции
Макроэволюция: направления и пути эволюции Сезонные изменения в жизни организмов
Сезонные изменения в жизни организмов Биотехнология туралы түсінік
Биотехнология туралы түсінік Підтип саркодові
Підтип саркодові Морфология, физиология и классификация грибов, простейших и вирусов
Морфология, физиология и классификация грибов, простейших и вирусов Рысь
Рысь Хвойные деревья Крыма
Хвойные деревья Крыма Автомобиль и экология
Автомобиль и экология Органические и неорганические вещества, входящие в состав клетки. Нуклеиновые кислоты и их роль в клетке
Органические и неорганические вещества, входящие в состав клетки. Нуклеиновые кислоты и их роль в клетке Солнечная активность и её влияние на человека
Солнечная активность и её влияние на человека Презентация по теме Витамины
Презентация по теме Витамины Собаки
Собаки Приспособления организмов к жизни в природе
Приспособления организмов к жизни в природе Основы сравнительной эмбриологии
Основы сравнительной эмбриологии Кожа – наружный покровный орган
Кожа – наружный покровный орган Игра Счастливый случай для 5-6 классов
Игра Счастливый случай для 5-6 классов Половое размножение. Образование половых клеток
Половое размножение. Образование половых клеток Класс паукообразные. Образ жизни и внешнее строение паука – крестовика
Класс паукообразные. Образ жизни и внешнее строение паука – крестовика Циклы развития растений. Подготовка к выполнению заданий ЕГЭ С5
Циклы развития растений. Подготовка к выполнению заданий ЕГЭ С5 Класс Насекомые. Развитие насекомых
Класс Насекомые. Развитие насекомых эволюция кровеносной системы
эволюция кровеносной системы Імунітет. Види імунітету
Імунітет. Види імунітету Гидротермические условия для выращивания растений
Гидротермические условия для выращивания растений Прецептрон. Практична реалізація
Прецептрон. Практична реалізація Тип Хордовые (chordata)
Тип Хордовые (chordata)