- Биоэнергетика. Биологическое окисление

Содержание
- 2. План Цепь переноса электронов и окислительное фосфорилирование; Окислительно-восстановительный потенциал; Переносчики электронов дыхательной цепи Сопряжение окисления и
- 4. Цикл превращений АТФ АТФ универсальный переносчик энергии для клеток; аутотрофы трансформируют энергию солнечного света в энергию
- 6. Роль окислительно-восстановительных процессов в метаболизме Основной путь получения энергии гетеротрофами - окисление органических веществ путем дегидрирования.
- 7. Коферменты никотиновой кислоты и никотинамида Существуют два кофермента никотиновой кислоты и никотинамида: НАД+ и НАДФ+; НАД+
- 10. Сравнение степеней окисления атомов углерода в биомолекулах
- 11. Окислительно-восстановительные реакции Окислительно-восстановительные реакции включают транспорт электронов, каждая из реакций состоит из двух сопряженных полуреакций: А(окисленный)+В(восстановленный)=В(окисленный)+А(восстановленный)
- 12. Окислительно-восстановительный потенциал. Стандартный окислительно-восстановительный потенциал (редокс потенциал ) для полуреакций – электрический потенциал (E0') в вольтах,
- 13. Стандартные условия: - рН 7; температура 250С; концентрация растворенных веществ 1моль; атмосферное давление 1 атм.
- 15. Аппарат для измерения стандартного окислительно-восстановительного потенциала
- 17. Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2. Δ Go' = -nF ΔEo'
- 18. Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2. ΔE НАДН/ О2 = (О,82-(-
- 19. Строение дыхательной цепи
- 20. Строение дыхательной цепи Четыре белковых комплекса расположены во внутренней митохондриальной мембране Жирорастворимый кофермент кофермент Q и
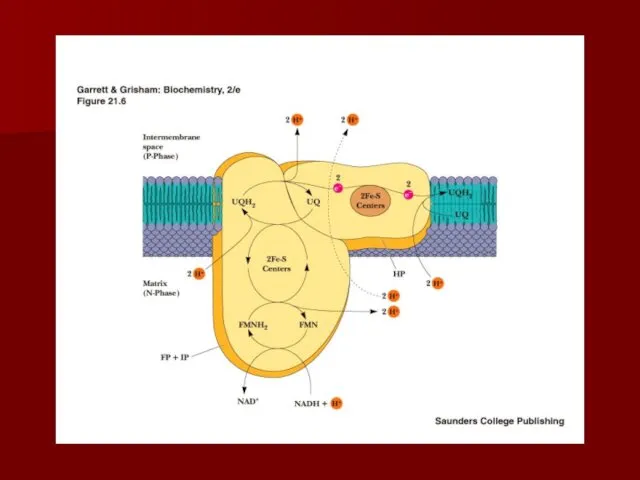
- 25. Комплекс I НАДН-КоQ редуктаза Переносит электроны от НАДН к коферменту Q Состоит из более 30 субъединиц
- 27. Рибофлавин (витамин В2) Молекула содержит спирт рибитол и флавин или изоаллоксазиновое кольцо Активные формы витамина –
- 28. Флавиновые коферменты ФМН и ФАД – одно и двухэлектронные переносчики Название флавин происходит от латинского flavius,
- 29. ФМН
- 30. Комплекс II (сукцинат-кофермент Q редуктаза) сукцинатдегидрогеназа – фермент цикла Кребса; сукцинатдегидрогеназа – ФАД-зависима; Содержит четыре субъединицы,
- 31. ФАД
- 32. Типы железо-серных центров
- 34. Убихинон
- 36. Комплекс III коэнзим Q-цитохром с редуктаза коэнзим Q передаёт электроны на -цитохром с и выкачивает протоны
- 41. Комплекс IV цитохром с оксидаза Электроны с цитохрома с используются для четырехэлектронного восстановления молекулы кислорода, что
- 47. Сопряжение транспорта электронов и окислительного фосфорилирования Много исследователей пытались найти «высокоэнергетический посредник»; Питер Митчелл предложил необычную
- 49. Q цикл Происходит передача двух электронов с восстановленной формы коэнзима Q: один на цитохром с, другой
- 52. Гипотеза Митчелла
- 53. Основные положения теории Митчелла. Перенос электронов по дыхательной цепи от НАД.Н+Н+ к кислороду сопровождается выкачиванием протонов
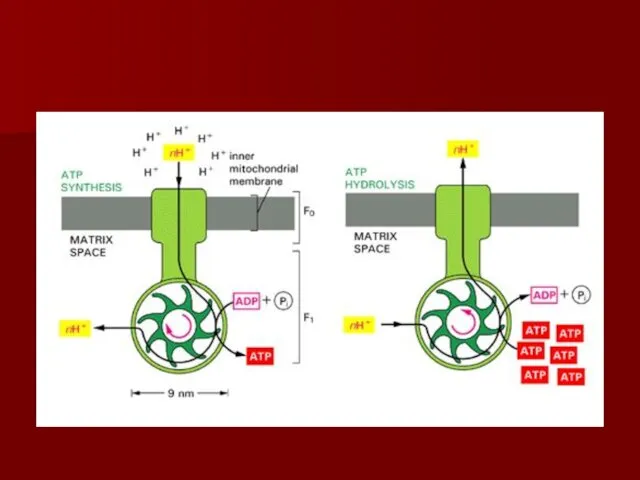
- 54. Протонный градиент и АТФ синтаза
- 57. АТФ синтаза
- 58. Конформационные изменения АТФ синтазы
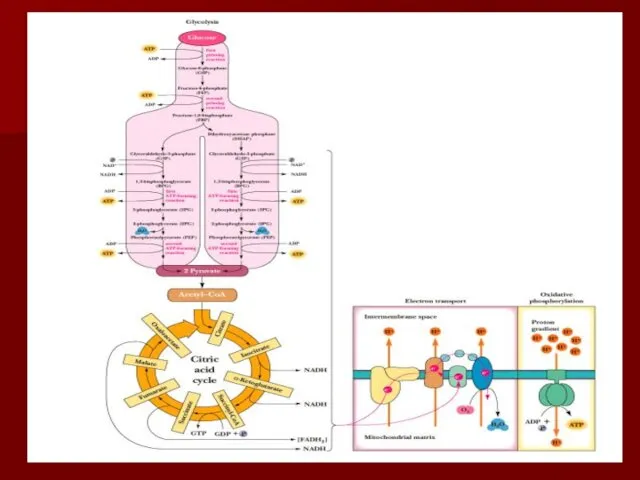
- 61. Резюме Дыхательная цепь- путь переноса протонов и электронов от органических веществ на кислород воздуха Это система
- 63. Ингибиторы окислительного фосфорилирования Ротенон (рыбий яд) ингибирует комплекс I; Цианиды, азиды и угарный газ ингибируют комплекс
- 66. Разобщители Разобщение потока электронов и окислительного фосфорилирования происходит при нарушении протонного градиента; Разобщители являются гидрофобными молекулами,
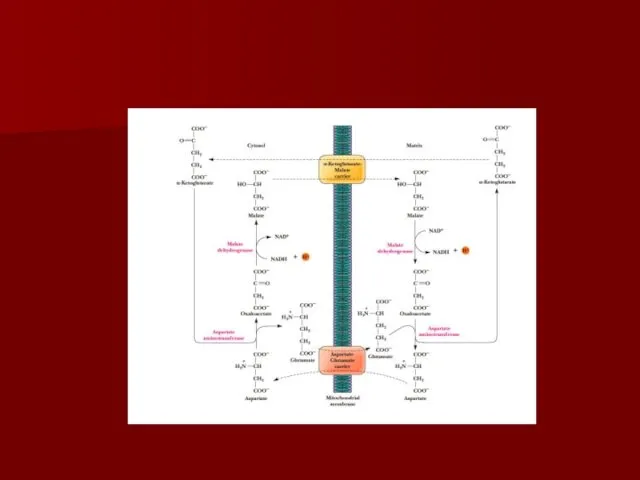
- 70. АТФ-АДФ транслоказа В большинстве клеток синтез основного количества АТФ происходит в митохондриях, а основные потребители АТФ
- 73. Коэффициент P/O Означает сколько молекул АТФ синтезируется при передаче пары электронов от субстрата на кислород; Транспорт
- 74. Активные формы кислорода. Кислород является потенциально токсическим веществом.Токсичность может возрастать в результате неполного восстановления молекулы кислорода
- 75. Супероксид ион Супероксид ион может образовываться в митохондриях при реакции кислорода с восстановленными ФАД или убихиноном.
- 76. Механизм действия супероксиддисмутазы
- 77. Пероксид водорода Н2О2 так же токсичен и разрушается пероксидазой в следующей реакции: 2Н2О2→2Н2О+О2 а. Наиболее распространенная
- 79. Биологическое окисление
- 82. Скачать презентацию

План
Цепь переноса электронов и окислительное фосфорилирование;
Окислительно-восстановительный потенциал;
Переносчики электронов дыхательной цепи
Сопряжение окисления
План
Цепь переноса электронов и окислительное фосфорилирование;
Окислительно-восстановительный потенциал;
Переносчики электронов дыхательной цепи
Сопряжение окисления
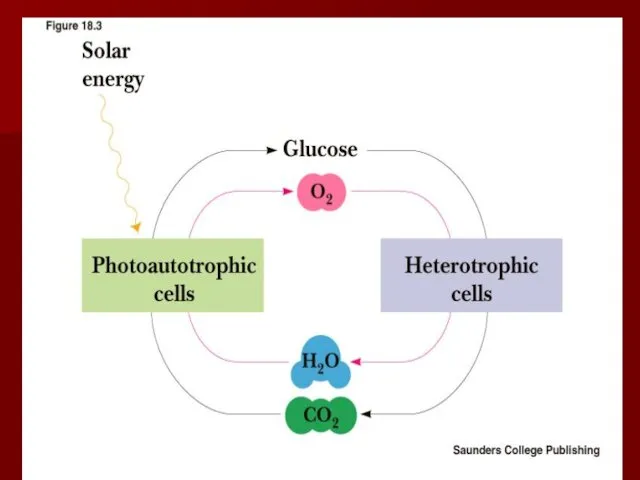

Цикл превращений АТФ
АТФ универсальный переносчик энергии для клеток;
аутотрофы трансформируют энергию солнечного
Цикл превращений АТФ
АТФ универсальный переносчик энергии для клеток;
аутотрофы трансформируют энергию солнечного
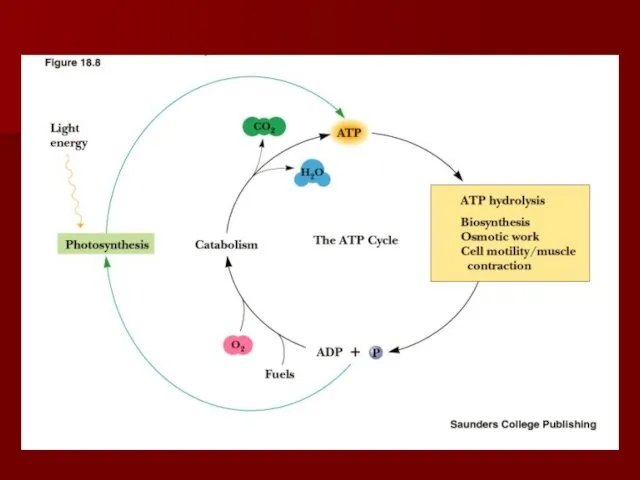

Роль окислительно-восстановительных процессов в метаболизме
Основной путь получения энергии гетеротрофами - окисление
Роль окислительно-восстановительных процессов в метаболизме
Основной путь получения энергии гетеротрофами - окисление

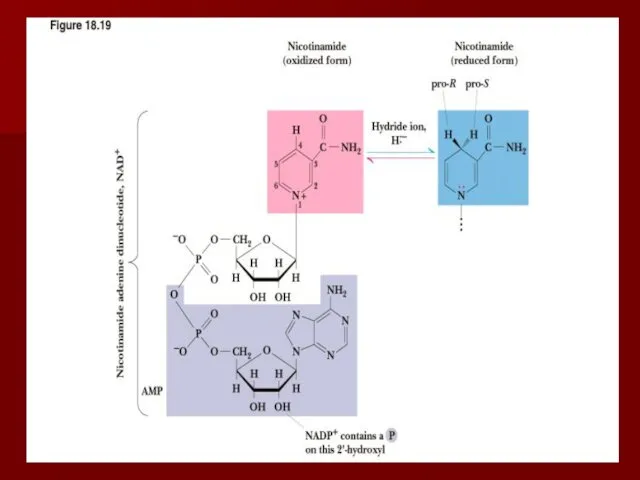
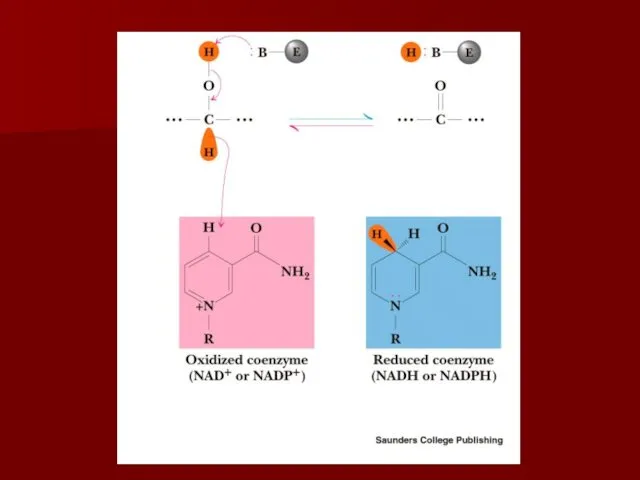
Коферменты никотиновой кислоты и никотинамида
Существуют два кофермента никотиновой кислоты и никотинамида:
Коферменты никотиновой кислоты и никотинамида
Существуют два кофермента никотиновой кислоты и никотинамида:

Сравнение степеней окисления
атомов углерода в биомолекулах
Сравнение степеней окисления
атомов углерода в биомолекулах

Окислительно-восстановительные
реакции
Окислительно-восстановительные реакции включают транспорт электронов, каждая из реакций состоит
Окислительно-восстановительные
реакции
Окислительно-восстановительные реакции включают транспорт электронов, каждая из реакций состоит

Окислительно-восстановительный
потенциал.
Стандартный окислительно-восстановительный
потенциал (редокс потенциал ) для полуреакций –
Окислительно-восстановительный
потенциал.
Стандартный окислительно-восстановительный потенциал (редокс потенциал ) для полуреакций –

Стандартные условия:
- рН 7;
температура 250С;
концентрация растворенных веществ 1моль;
атмосферное давление 1
Стандартные условия:
- рН 7;
температура 250С;
концентрация растворенных веществ 1моль;
атмосферное давление 1
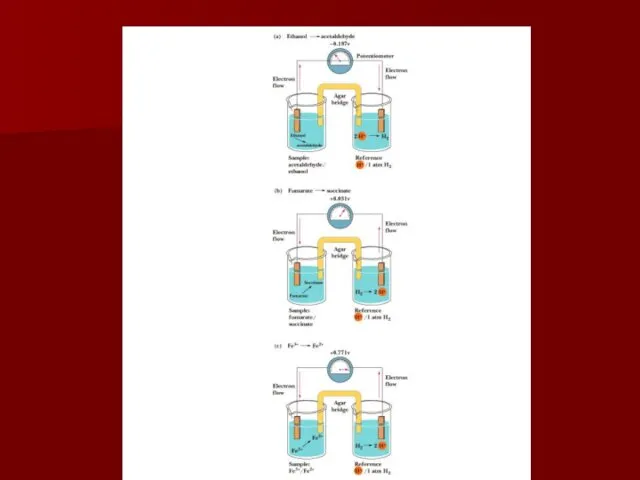
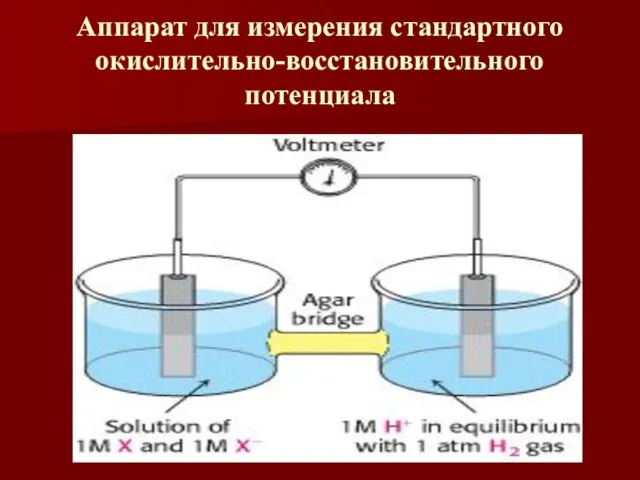
Аппарат для измерения стандартного окислительно-восстановительного потенциала
Аппарат для измерения стандартного окислительно-восстановительного потенциала


Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2.
Δ
Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2.
Δ

Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2.
ΔE
Уравнение расчета энергии переноса 2е от НАДН+ Н+ к ½ О2.
ΔE
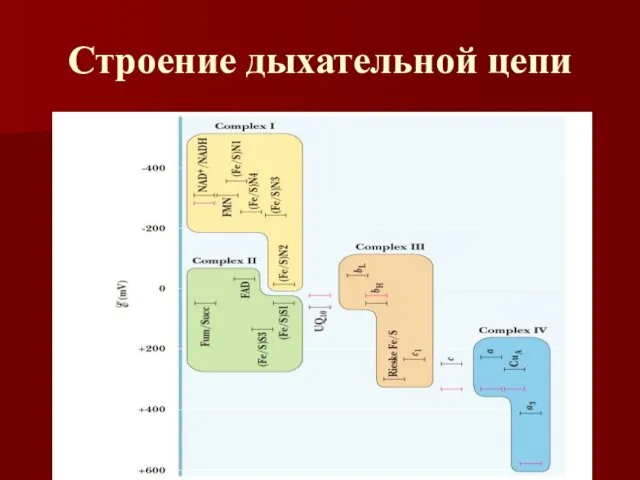
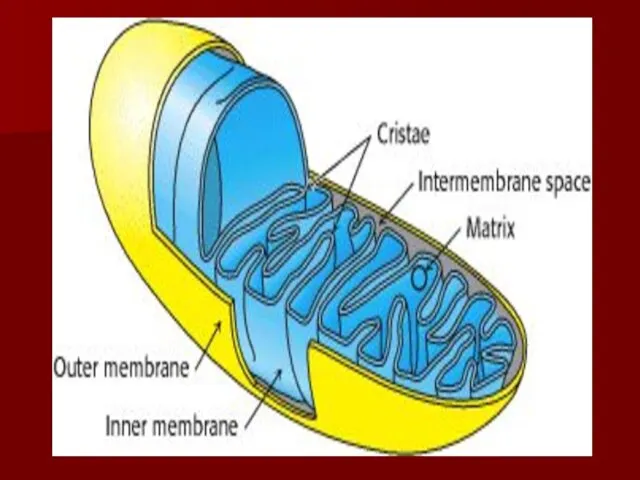
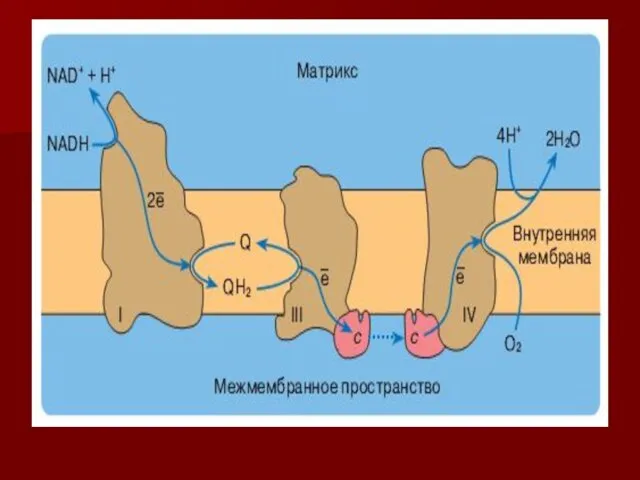
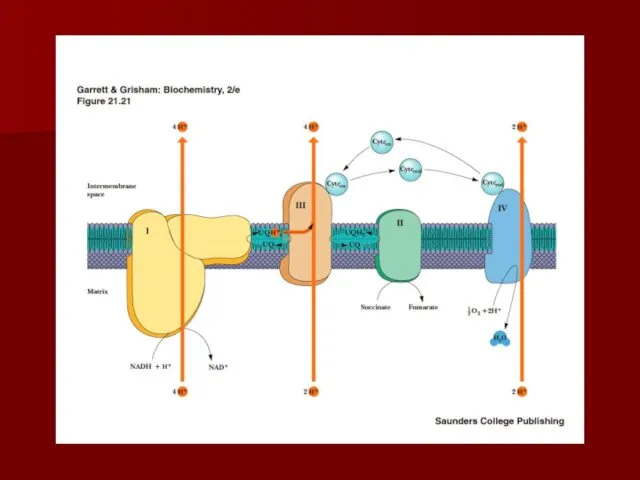
Строение дыхательной цепи
Строение дыхательной цепи

Строение дыхательной цепи
Четыре белковых комплекса расположены во внутренней митохондриальной мембране
Жирорастворимый кофермент
Строение дыхательной цепи
Четыре белковых комплекса расположены во внутренней митохондриальной мембране
Жирорастворимый кофермент
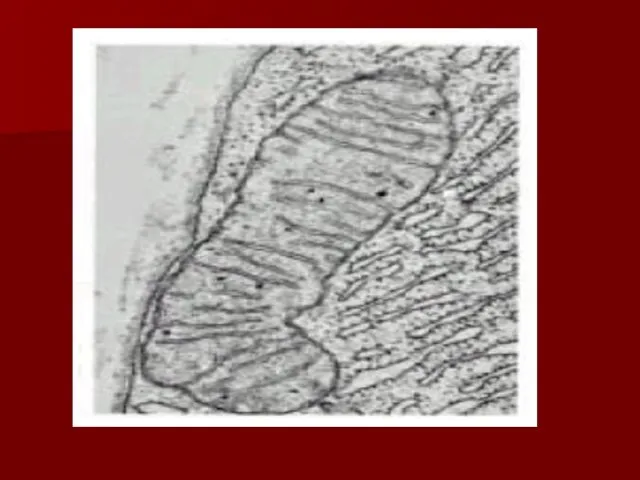
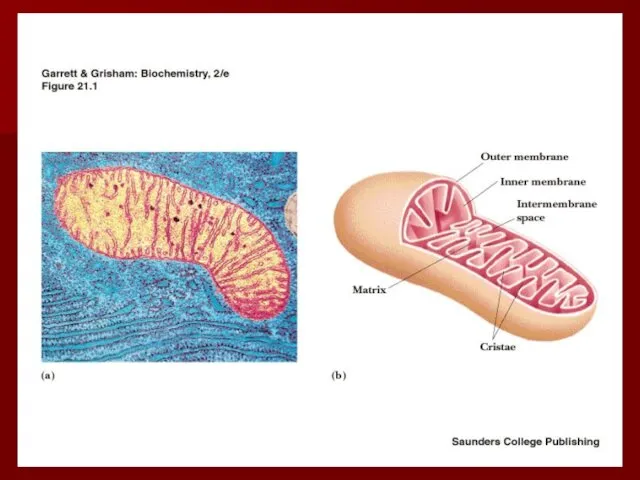
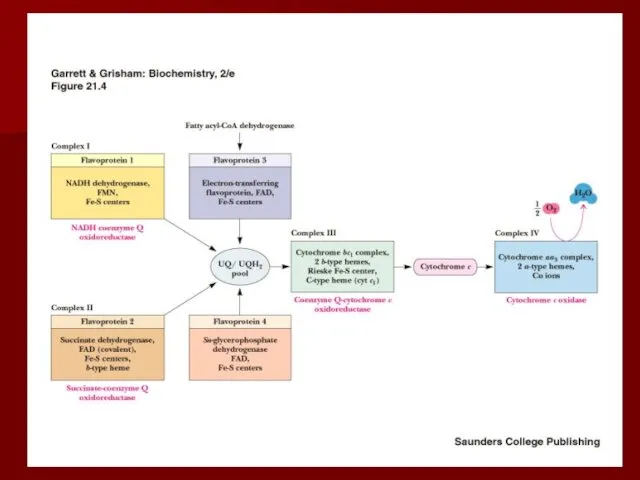

Комплекс I
НАДН-КоQ редуктаза
Переносит электроны от НАДН к коферменту Q
Состоит из
Комплекс I
НАДН-КоQ редуктаза
Переносит электроны от НАДН к коферменту Q
Состоит из

Рибофлавин
(витамин В2)
Молекула содержит спирт рибитол и флавин или изоаллоксазиновое кольцо
Активные
Рибофлавин
(витамин В2)
Молекула содержит спирт рибитол и флавин или изоаллоксазиновое кольцо
Активные

Флавиновые коферменты
ФМН и ФАД – одно и двухэлектронные переносчики
Название флавин происходит
Флавиновые коферменты
ФМН и ФАД – одно и двухэлектронные переносчики
Название флавин происходит
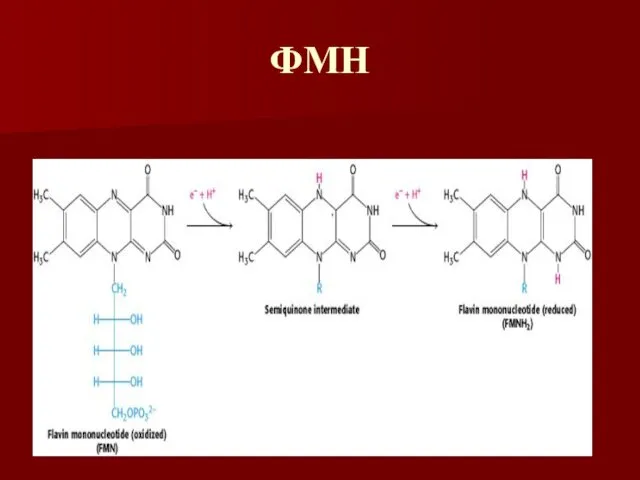
ФМН
ФМН

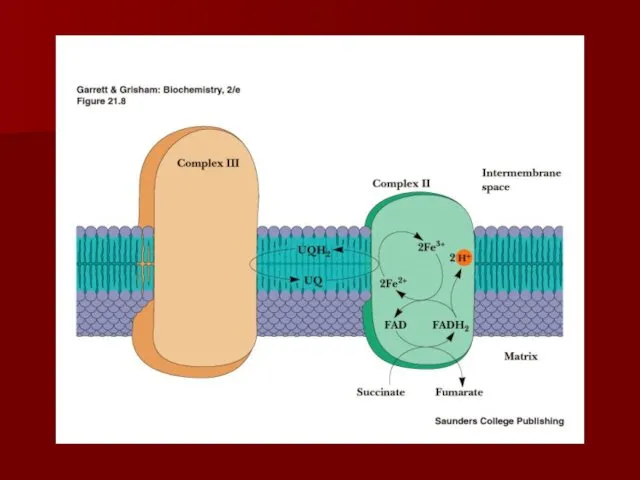
Комплекс II
(сукцинат-кофермент Q редуктаза)
сукцинатдегидрогеназа – фермент цикла Кребса;
сукцинатдегидрогеназа – ФАД-зависима;
Содержит
Комплекс II
(сукцинат-кофермент Q редуктаза)
сукцинатдегидрогеназа – фермент цикла Кребса;
сукцинатдегидрогеназа – ФАД-зависима;
Содержит
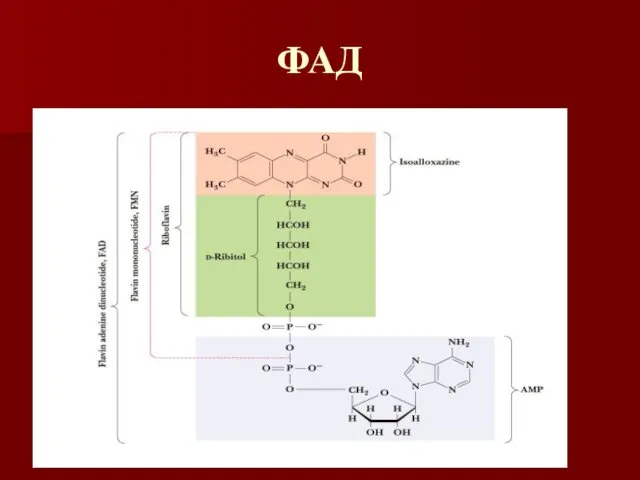
ФАД
ФАД
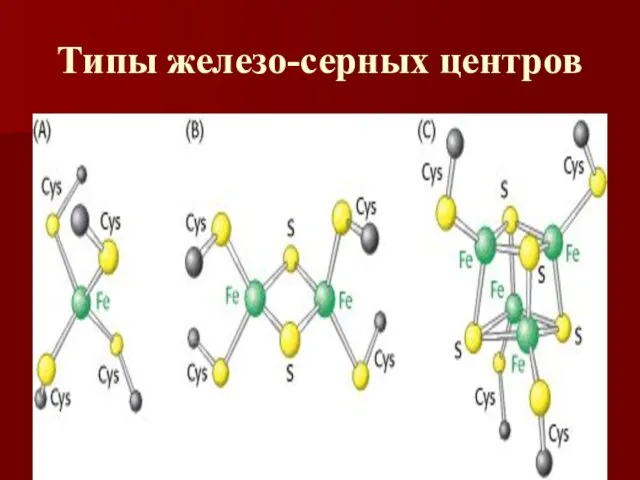
Типы железо-серных центров
Типы железо-серных центров
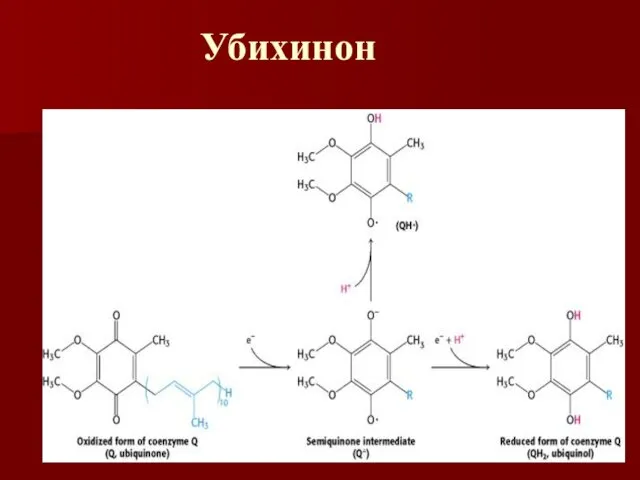
Убихинон
Убихинон
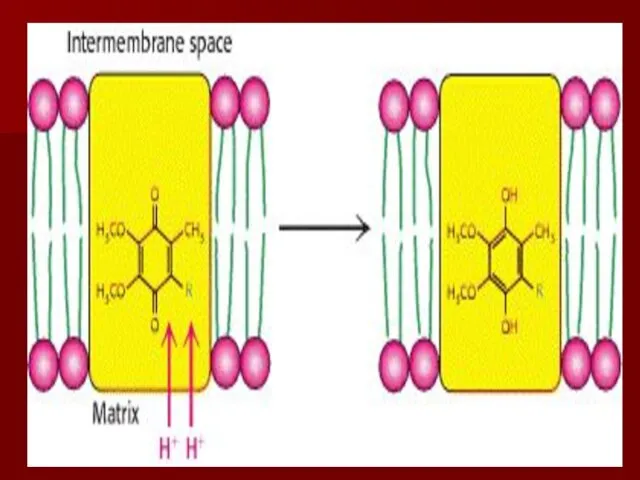

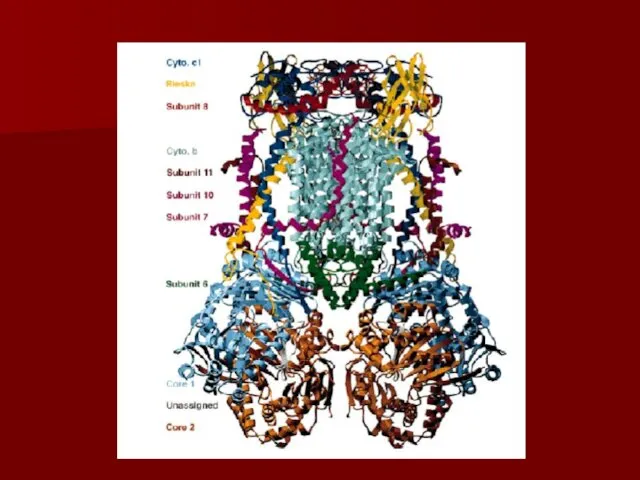
Комплекс III
коэнзим Q-цитохром с редуктаза
коэнзим Q передаёт электроны на -цитохром
Комплекс III
коэнзим Q-цитохром с редуктаза
коэнзим Q передаёт электроны на -цитохром
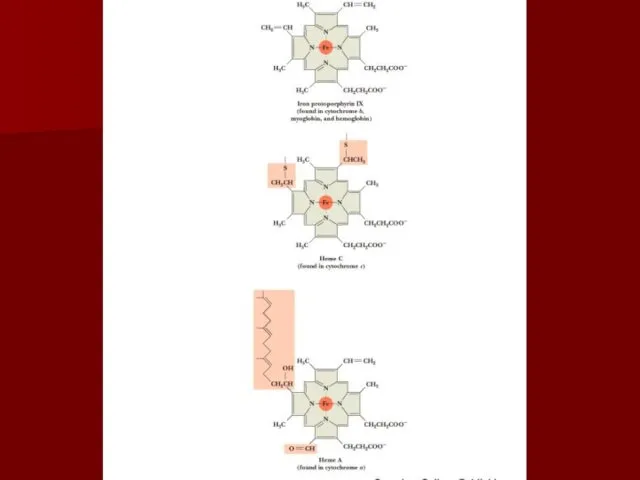

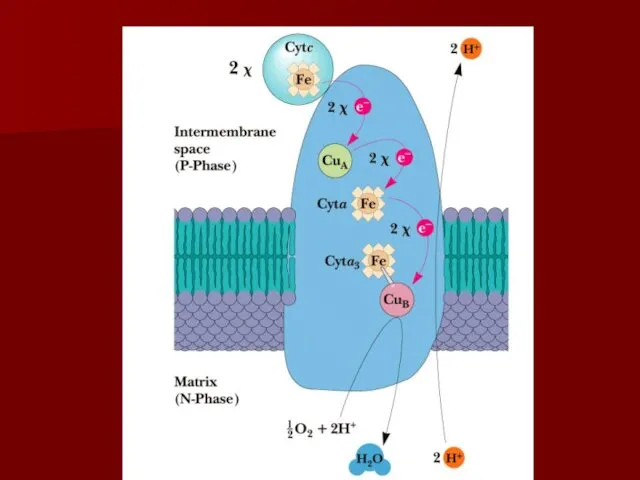
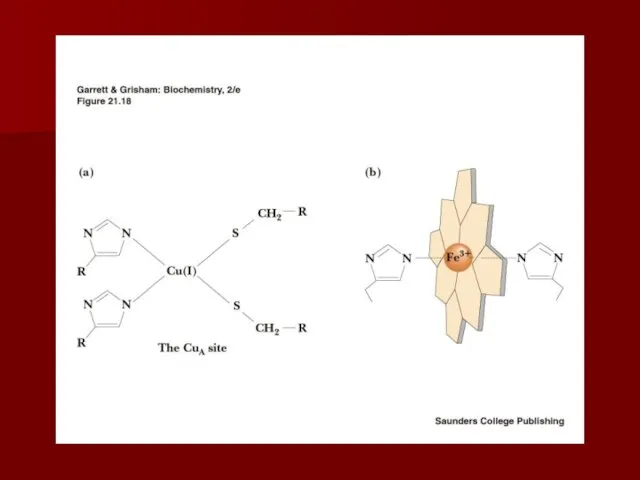
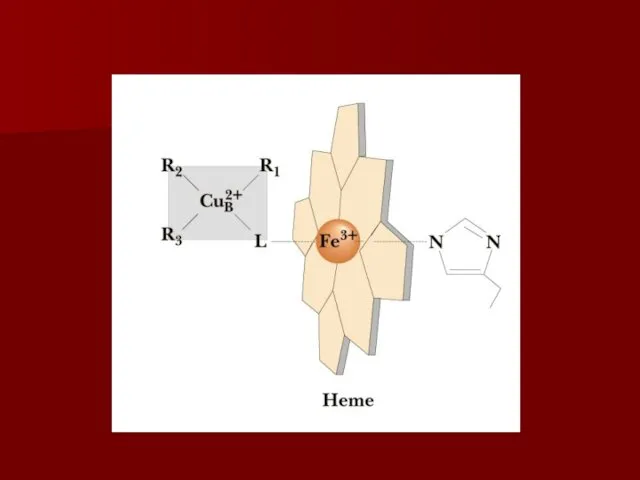
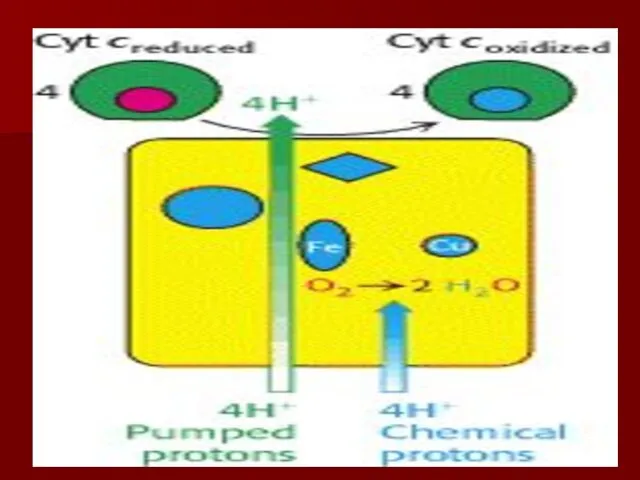
Комплекс IV
цитохром с оксидаза
Электроны с цитохрома с используются для четырехэлектронного восстановления
Комплекс IV
цитохром с оксидаза
Электроны с цитохрома с используются для четырехэлектронного восстановления
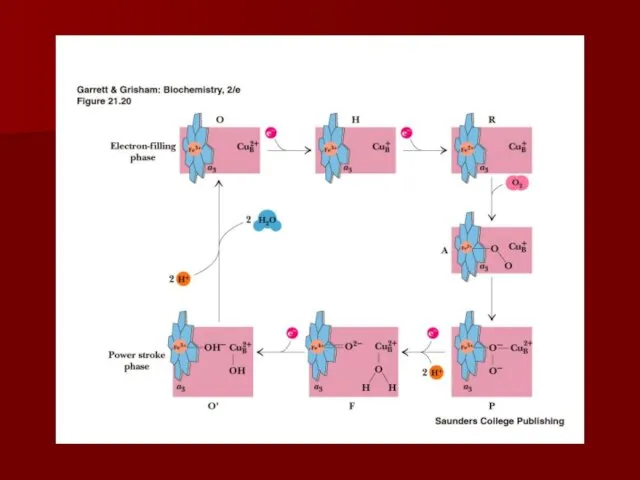

Сопряжение транспорта электронов и окислительного фосфорилирования
Много исследователей пытались найти «высокоэнергетический посредник»;
Сопряжение транспорта электронов и окислительного фосфорилирования
Много исследователей пытались найти «высокоэнергетический посредник»;

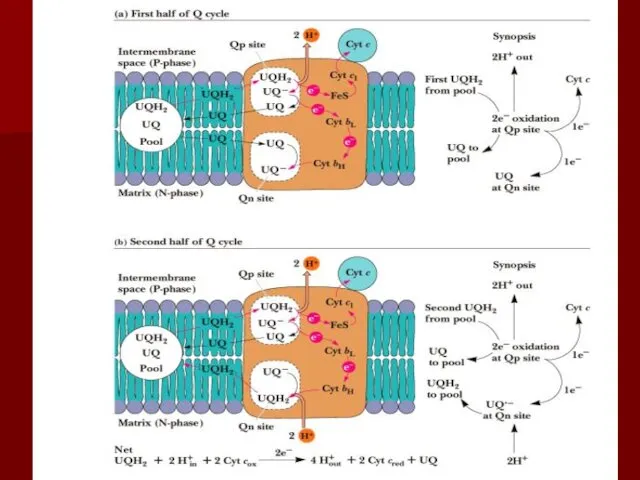
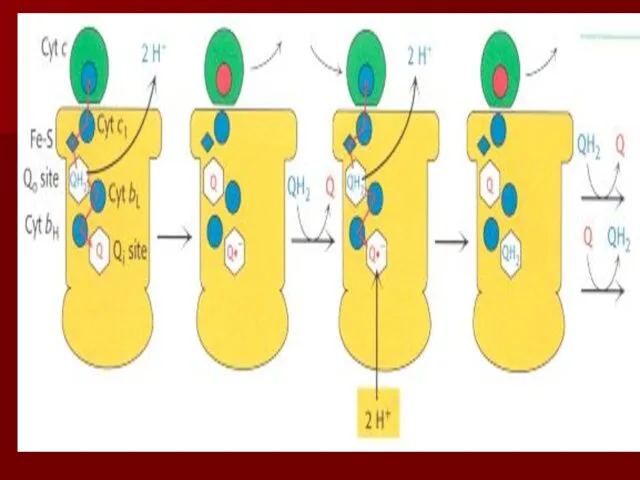
Q цикл
Происходит передача двух электронов с восстановленной формы коэнзима Q: один
Q цикл
Происходит передача двух электронов с восстановленной формы коэнзима Q: один
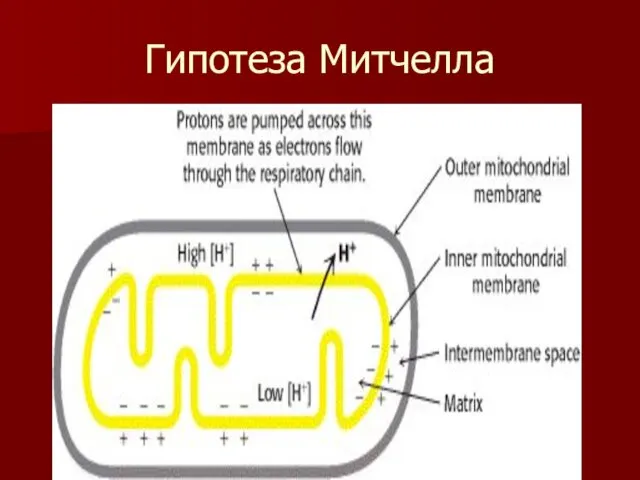
Гипотеза Митчелла
Гипотеза Митчелла

Основные положения теории Митчелла.
Перенос электронов по дыхательной цепи от НАД.Н+Н+ к
Основные положения теории Митчелла.
Перенос электронов по дыхательной цепи от НАД.Н+Н+ к

Протонный градиент
и АТФ синтаза
Протонный градиент
и АТФ синтаза
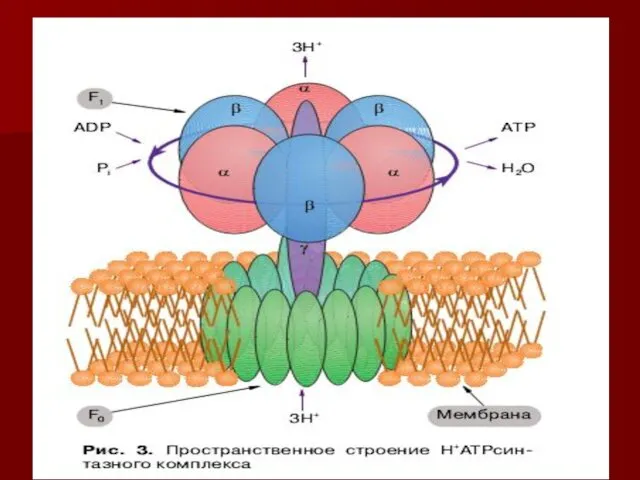
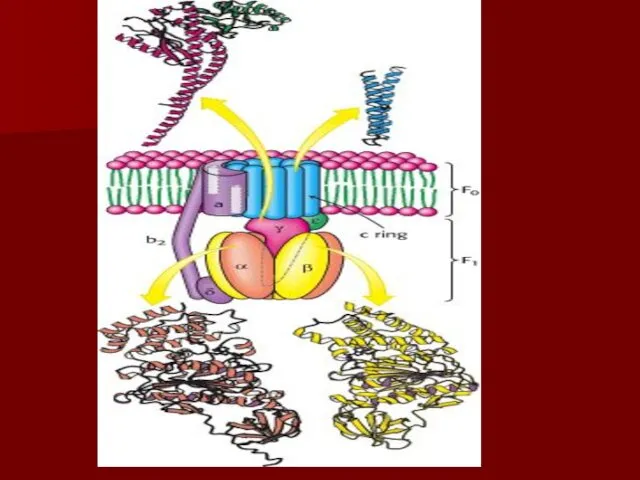
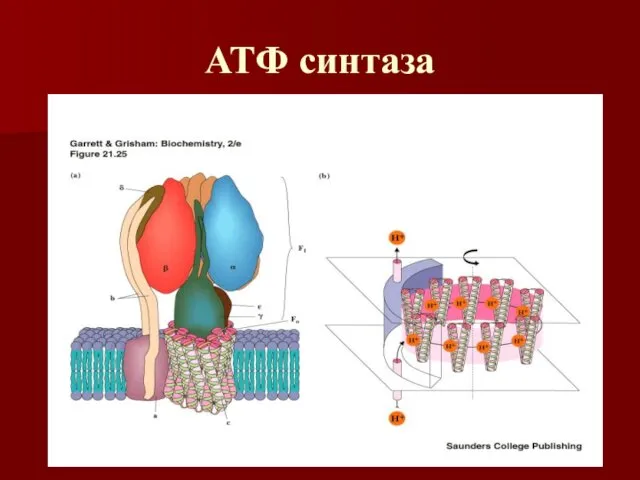
АТФ синтаза
АТФ синтаза
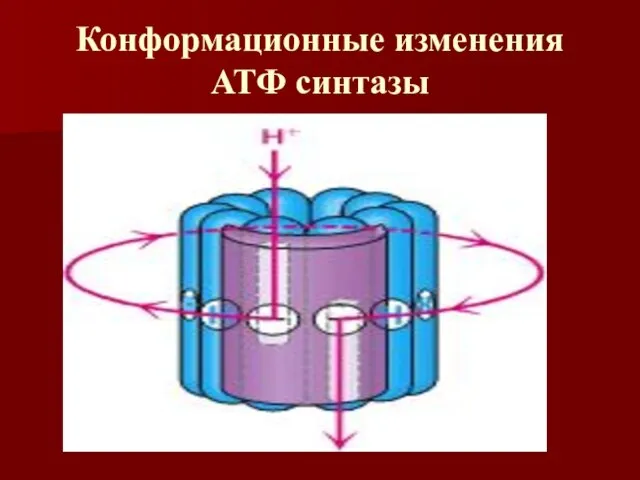
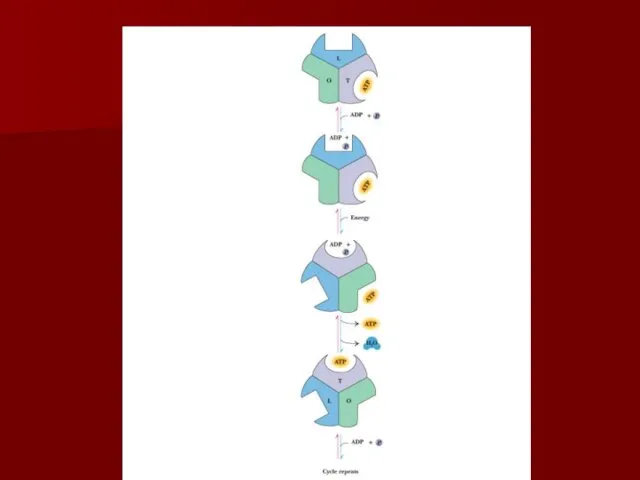
Конформационные изменения АТФ синтазы
Конформационные изменения АТФ синтазы
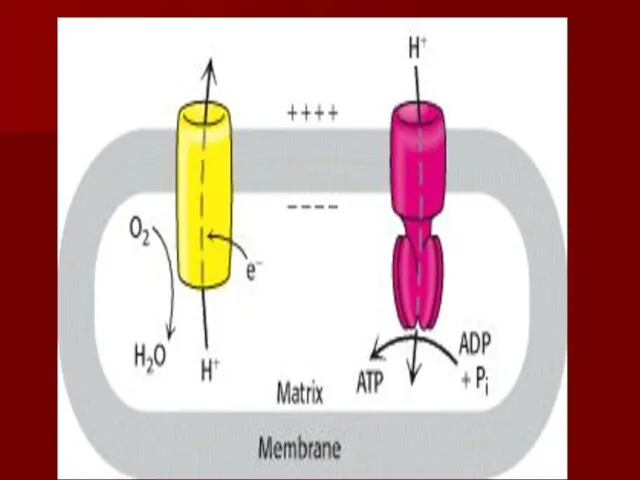


Резюме
Дыхательная цепь- путь переноса протонов и электронов от органических веществ на
Резюме
Дыхательная цепь- путь переноса протонов и электронов от органических веществ на

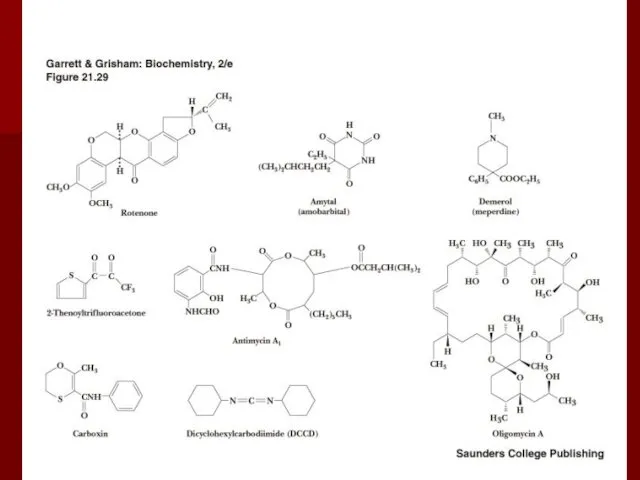
Ингибиторы окислительного фосфорилирования
Ротенон (рыбий яд) ингибирует комплекс I;
Цианиды, азиды и
Ингибиторы окислительного фосфорилирования
Ротенон (рыбий яд) ингибирует комплекс I;
Цианиды, азиды и
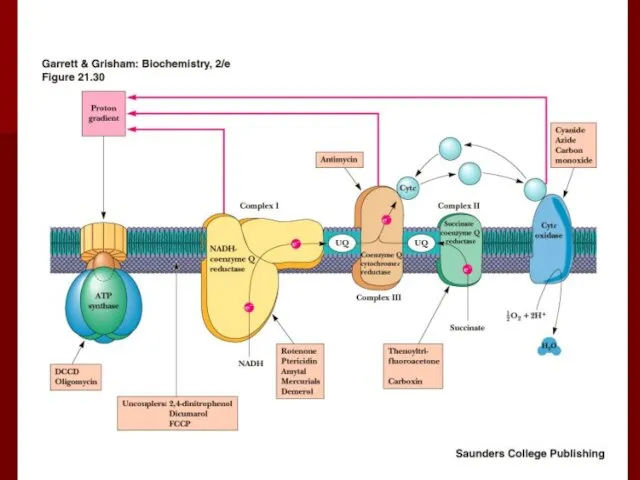

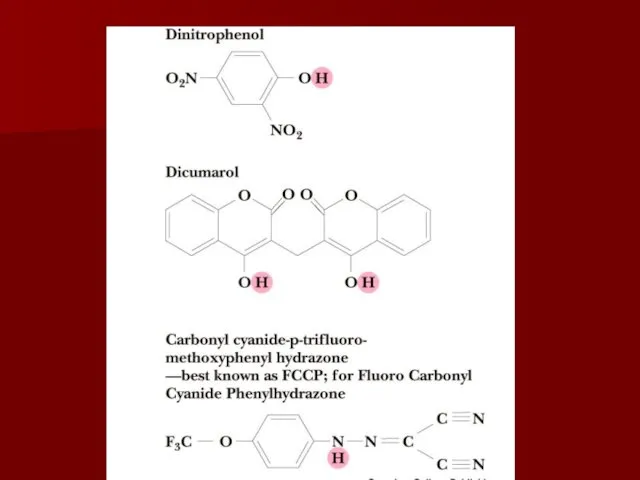
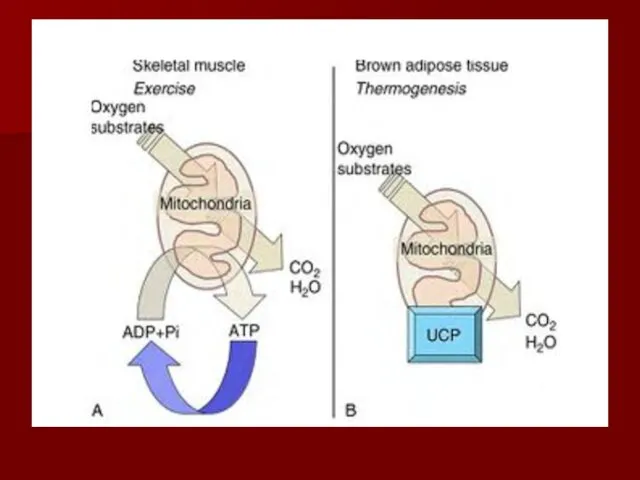
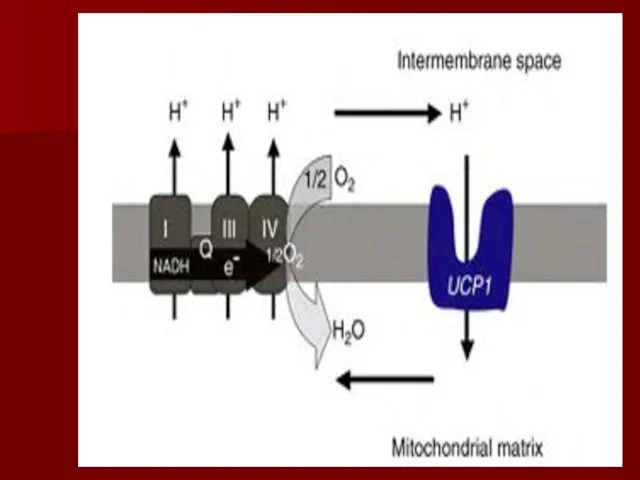
Разобщители
Разобщение потока электронов и окислительного фосфорилирования происходит при нарушении протонного градиента;
Разобщители
Разобщители
Разобщение потока электронов и окислительного фосфорилирования происходит при нарушении протонного градиента;
Разобщители

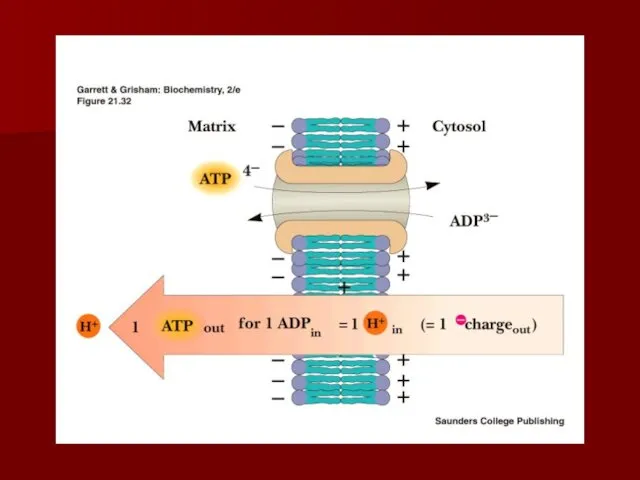
АТФ-АДФ транслоказа
В большинстве клеток синтез основного количества АТФ происходит в митохондриях,
АТФ-АДФ транслоказа
В большинстве клеток синтез основного количества АТФ происходит в митохондриях,

Коэффициент P/O
Означает сколько молекул АТФ синтезируется при передаче пары электронов от
Коэффициент P/O
Означает сколько молекул АТФ синтезируется при передаче пары электронов от

Активные формы кислорода.
Кислород является потенциально токсическим веществом.Токсичность может возрастать в
Активные формы кислорода.
Кислород является потенциально токсическим веществом.Токсичность может возрастать в

Супероксид ион
Супероксид ион может образовываться в митохондриях при реакции кислорода с
Супероксид ион
Супероксид ион может образовываться в митохондриях при реакции кислорода с
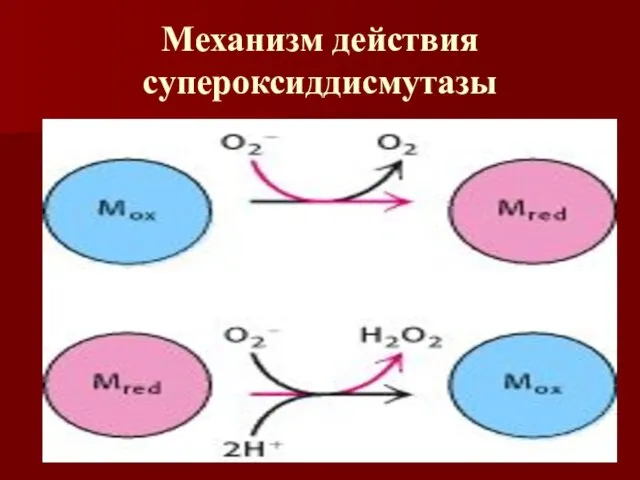
Механизм действия супероксиддисмутазы
Механизм действия супероксиддисмутазы

Пероксид водорода
Н2О2 так же токсичен и разрушается пероксидазой в следующей
Пероксид водорода
Н2О2 так же токсичен и разрушается пероксидазой в следующей
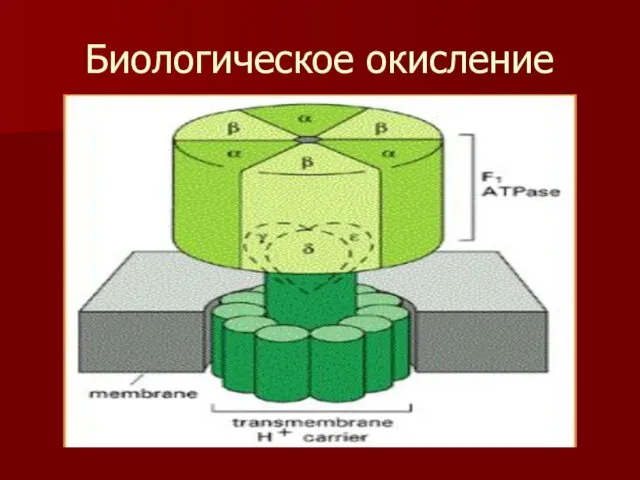
Биологическое окисление
Биологическое окисление
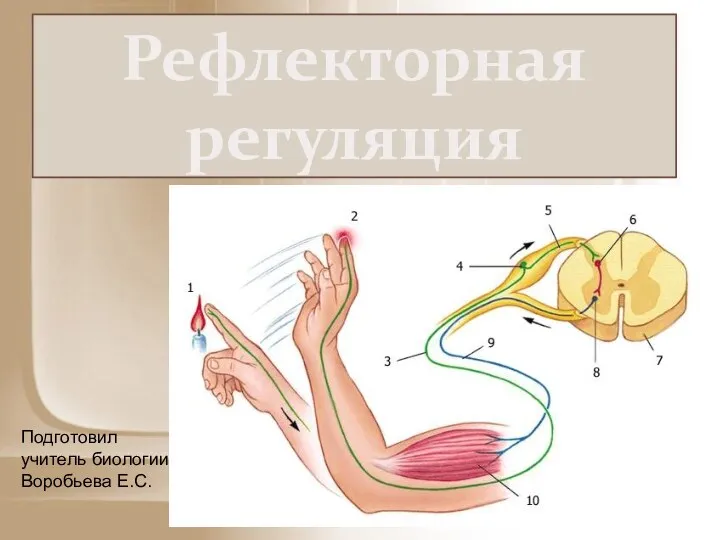 Рефлекторная регуляция
Рефлекторная регуляция Ленинградский зоопарк
Ленинградский зоопарк Изменчивость: наследственная и ненаследственная. Мутации
Изменчивость: наследственная и ненаследственная. Мутации Орієнтування тварин
Орієнтування тварин Витамины. Какие манипуляции с продуктами позволяют сохранить в них витамины
Витамины. Какие манипуляции с продуктами позволяют сохранить в них витамины Репликация. Синтез ДНК по матрице ДНК
Репликация. Синтез ДНК по матрице ДНК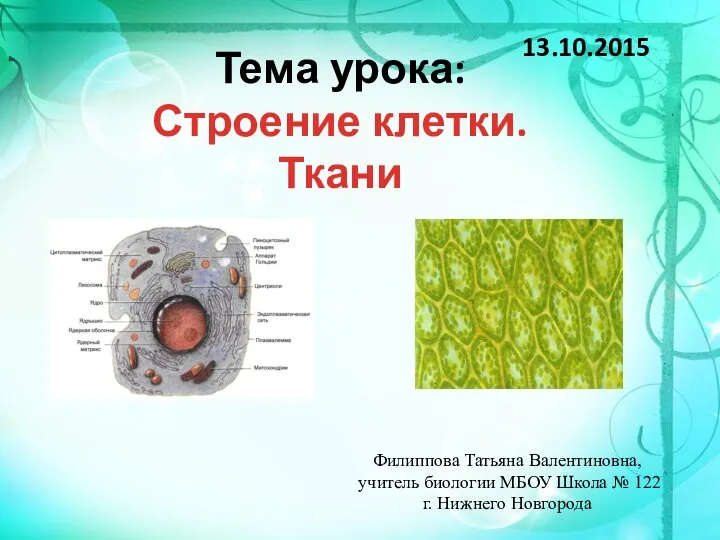 Строение клетки ткани
Строение клетки ткани Живое вещество биосферы, его функции
Живое вещество биосферы, его функции Репликация. Прокариоты. Репликация фагов
Репликация. Прокариоты. Репликация фагов Zoológico de Madrid
Zoológico de Madrid Собачий клещ (Ixodes ricinus)
Собачий клещ (Ixodes ricinus) Ландшафтні квіткові композиції для оздоблення прибудинкових територій
Ландшафтні квіткові композиції для оздоблення прибудинкових територій ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Тест Рыбы 7 класс.
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Тест Рыбы 7 класс. Презентация по биологии на тему _Плауны,хвощи,папоротники_(5 класс) (1)
Презентация по биологии на тему _Плауны,хвощи,папоротники_(5 класс) (1) Основы систематики микроорганизмов
Основы систематики микроорганизмов Хрящевые рыбы
Хрящевые рыбы Основы сравнительной эмбриологии
Основы сравнительной эмбриологии Клетка и методы цитологии
Клетка и методы цитологии Орган слуха и равновесия
Орган слуха и равновесия Функциональная анатомия вегетативной нервной системы. Симпатическая часть ВНС (лекция № 23)
Функциональная анатомия вегетативной нервной системы. Симпатическая часть ВНС (лекция № 23) Комнатные растения в интерьере помещения
Комнатные растения в интерьере помещения Хронобиология и биоритмы человека
Хронобиология и биоритмы человека Окапи
Окапи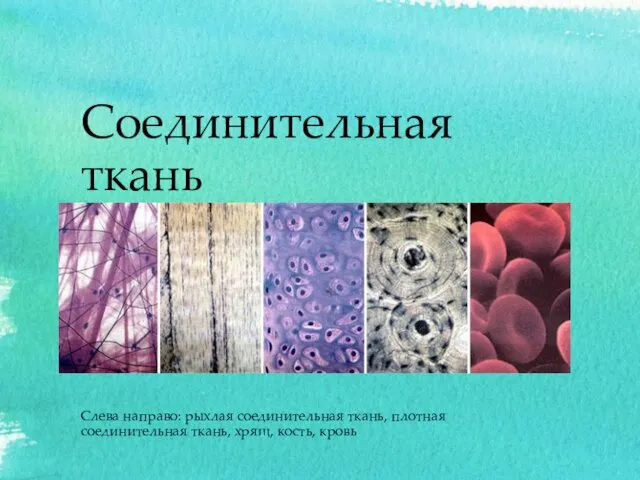 Соединительная ткань
Соединительная ткань Овчарки со всего мира. Фотографии
Овчарки со всего мира. Фотографии Общие признаки для всех живых организмов
Общие признаки для всех живых организмов Изоляция
Изоляция Соцветия. Строение соцветия
Соцветия. Строение соцветия