Биохимия мышечной деятельности. Общая характеристика механизма энергообеспечения. Лекция № 6 презентация
- Биохимия мышечной деятельности. Общая характеристика механизма энергообеспечения. Лекция № 6

Содержание
- 2. Биохимия мышечной деятельности. Общая характеристика механизма энергообеспечения. Подключение энергетических систем при различных физических нагрузках и их
- 3. Мышечная ткань Вся мускулатура состоит из 439 отдельных мышц, которые в совокупности составляют у женщин 30-40%,
- 4. N.B!!! Число мышечных волокон уменьшается с возрастом в результате увеличивающейся неактивности примерно на 30%, при этом
- 5. Мышечные клетки характеризуются наличием сокращающихся белковых структур – миофибрилл. Каждую мышечную ткань сопровождает соединительная ткань, которая
- 6. Продольные срезы поперечно-полосатой, гладкой и сердечной мышцы
- 7. ППМ иннервируется, прежде всего, соматической нервной системой, ГМ – только вегетативной. Сокращения ППМ проходят быстро и
- 8. Каждая мышечная ткань состоит из вытянутых в длину мышечных клеток (волокон) с одним или несколькими клеточными
- 10. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые построены из повторяющихся в продольном
- 11. Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула
- 12. Характеристика мышечных волокон
- 14. Химический состав мышечной ткани
- 15. Основные белки мышц 1. Саркоплазматические – 35%. Белки-ферменты митохондрий, катализируют процессы окислительного фосфорилирования; ферменты гликолиза, азотистого
- 16. Основные биохимические процессы Сокращение запускается нервным импульсом. В синапсе (1) выделяется ацетилхолин (2). АХ вызывает возбуждение
- 17. В таком состоянии миозиновые головки уже способны взаимодействовать с актином, но центр их взаимодействия блокирован тропонином.
- 18. Теория скольжения нитей мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга.
- 19. 3. Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина
- 20. Расслабление скелетной мышцы Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического
- 21. Функция АТФ в процессе сокращения и расслабления В покоящейся мышце – препятствует соединению актиновых нитей с
- 22. Общая характеристика механизмов энергообразования Ресинтез АТФ может осуществляться в реакциях, протекающих без участия кислорода (анаэробные механизмы)
- 23. Анаэробные механизмы Креатинфосфокиназный (фосфогенный или алактатный) механизм, обеспечивающий ресинтез АТФ за счет перефосфорилирования между креатинфосфатом и
- 24. Аэробный механизм ресинтеза АТФ включает в основном реакции окислительного фосфорилирования, протекаемые в митохондриях. Энергетическими субстратами аэробного
- 25. Критерии оценки механизмов энергообеспечения мышечной деятельности
- 26. Креатинфосфокиназный механизм ресинтеза АТФ Алактатный анаэробный механизм ресинтеза АТФ включает использование имеющейся в мышцах АТФ и
- 27. Максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5-0,7-й секунде интенсивной работы, что свидетельствует о большой скорости
- 28. Максимальная мощность креатинфосфокиназной реакции составляет 3,8 кДж•кг-1•мин-1, что значительно выше мощности анаэробного гликолиза (в 1,5-2 раза)
- 29. Запасы АТФ и Крф в скелетных мышцах человека
- 30. Изменение концентрации АТФ, КрФ и лактата в скелетных мышцах в процессе работы
- 31. Содержание КрФ в скелетных мышцах увеличивается в процессе адаптации организма к скоростным и силовым физическим нагрузкам
- 32. Гликолитический механизм ресинтеза АТФ Как только в процессе анаэробной мышечной работы КФК-ный механизм перестает обеспечивать необходимую
- 33. Схема гликолитического механизма ресинтеза АТФ в скелетных мышцах 1 – АТФ-аза миозина. 2 – КрФ (саркоплазма)
- 34. Максимальная мощность гликолиза у хорошо тренированных спортсменов может составлять 3,1 кДж•кг-1•мин-1, а у нетренированных людей –
- 35. Образование молочной кислоты в процессе гликолиза в зависимости от продолжительности упражнений
- 36. Молочная кислота может диффундировать через клеточные мембраны по градиенту концентрации и поступать из работающих мышц в
- 37. Определение ПЛ по кривым зависимости концентрации лактата, рН крови, уровня легочной вентиляции и «избыточного выделения» СО2
- 38. В последнее время при оценке степени адаптационных процессов метаболизма используют более простой метод измерения концентрации лактата
- 39. Изменение концентрации лактата в крови после заплыва на 200 м с заданной скоростью у пловцов в
- 40. Миокиназный механизм Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Она заключается
- 41. Аэробный механизм ресинтеза АТФ Аэробный механизм ресинтеза АТФ в обычных условиях обеспечивает около 90 % общего
- 42. Схема аэробного механизма ресинтеза АТФ, включающего процессы анаэробного превращения (I) и аэробное окисление – тканевое дыхание
- 43. Максимальная мощность аэробного механизма наименьшая и составляет 1,2 кДж•кг-1•мин-1 и в равной степени зависит от скорости
- 44. Максимальная мощность аэробного процесса достигается на 2-3-й минуте неинтенсивной работы у спортсменов и на 4-5-й минуте
- 45. При окислении высших жирных кислот, например пальмитиновой, образуется еще больше энергии: Эффективность энергообразования этого механизма также
- 46. Подключение энергетических систем при различных физических нагрузках и их адаптация в процессе тренировки В условиях относительного
- 47. Последовательность и вклад механизмов анаэробного и аэробного энергообразования в энергетику различных упражнений
- 48. Работа различной мощности и продолжительности обеспечивается различными механизмами энергообразования. С увеличением продолжительности бега ↓ доля анаэробных
- 49. В спортивной практике физические упражнения, в которых вклад анаэробных алактатного и гликолитического процессов составляет более 60%
- 50. Адаптационные изменения мощности (VO2) и емкости отдельных механизмов энергообеспечения мышечной работы в процессе специфической тренировки.
- 51. Мощность аэробного механизма, которая зависит от МПК и активности окислительных ферментов, также увеличивается в процессе адаптации
- 52. В процессе скоростной тренировки существенно изменяются анаэробные механизмы энергообеспечения. Отмечается ↑ их мощности и емкости. Это
- 54. Скачать презентацию

Биохимия мышечной деятельности. Общая характеристика механизма энергообеспечения. Подключение энергетических систем при
Биохимия мышечной деятельности. Общая характеристика механизма энергообеспечения. Подключение энергетических систем при
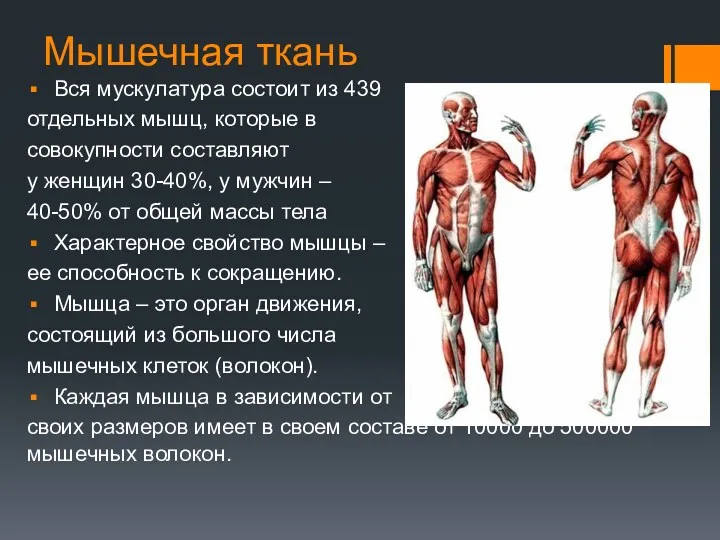
Мышечная ткань
Вся мускулатура состоит из 439
отдельных мышц, которые в
совокупности составляют
Мышечная ткань
Вся мускулатура состоит из 439
отдельных мышц, которые в
совокупности составляют

N.B!!!
Число мышечных волокон уменьшается с возрастом в результате увеличивающейся неактивности примерно
N.B!!!
Число мышечных волокон уменьшается с возрастом в результате увеличивающейся неактивности примерно
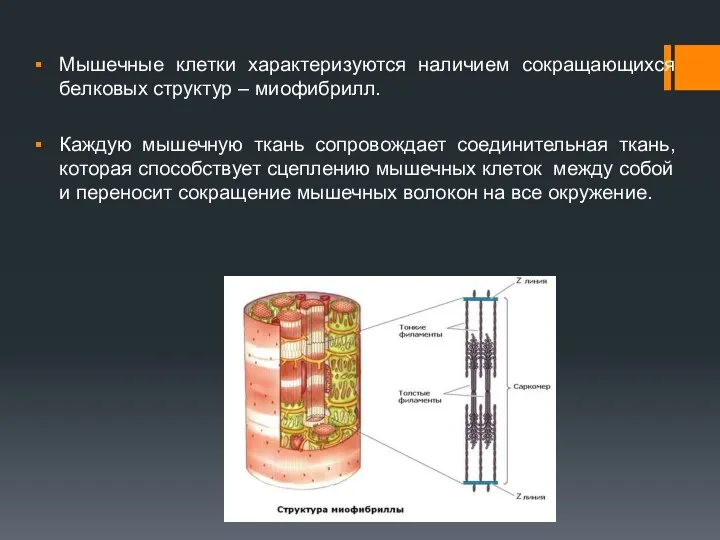
Мышечные клетки характеризуются наличием сокращающихся белковых структур – миофибрилл.
Каждую мышечную ткань
Мышечные клетки характеризуются наличием сокращающихся белковых структур – миофибрилл.
Каждую мышечную ткань
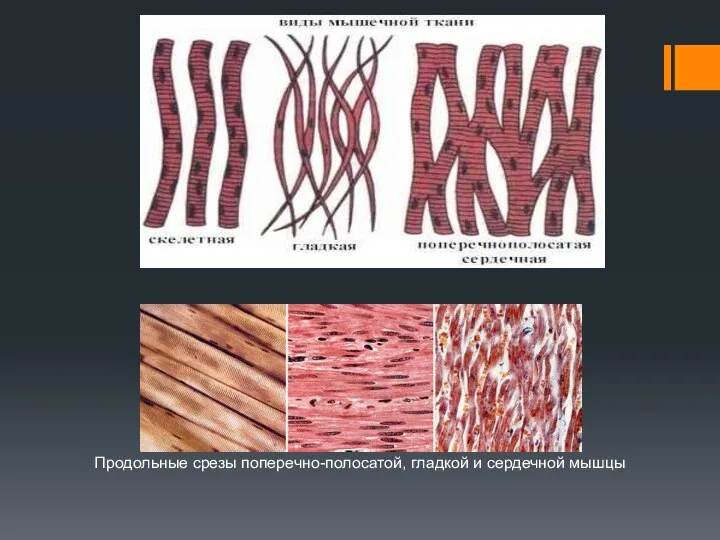
Продольные срезы поперечно-полосатой, гладкой и сердечной мышцы
Продольные срезы поперечно-полосатой, гладкой и сердечной мышцы

ППМ иннервируется, прежде всего, соматической нервной системой, ГМ – только вегетативной.
Сокращения
ППМ иннервируется, прежде всего, соматической нервной системой, ГМ – только вегетативной.
Сокращения
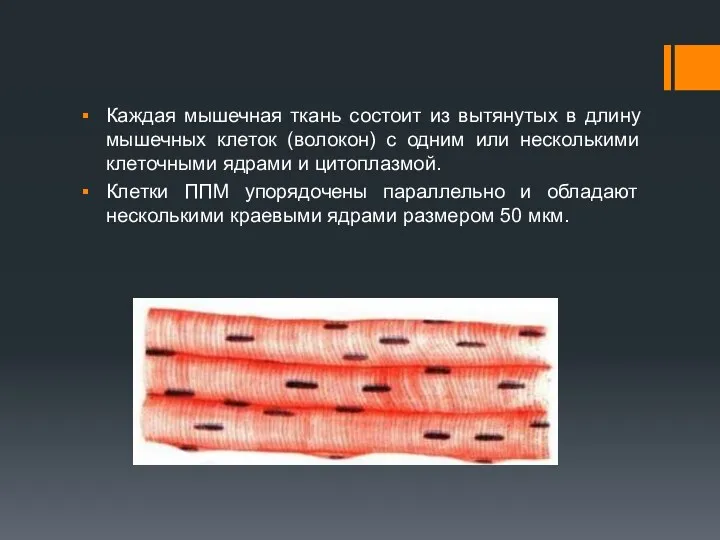
Каждая мышечная ткань состоит из вытянутых в длину мышечных клеток (волокон)
Каждая мышечная ткань состоит из вытянутых в длину мышечных клеток (волокон)
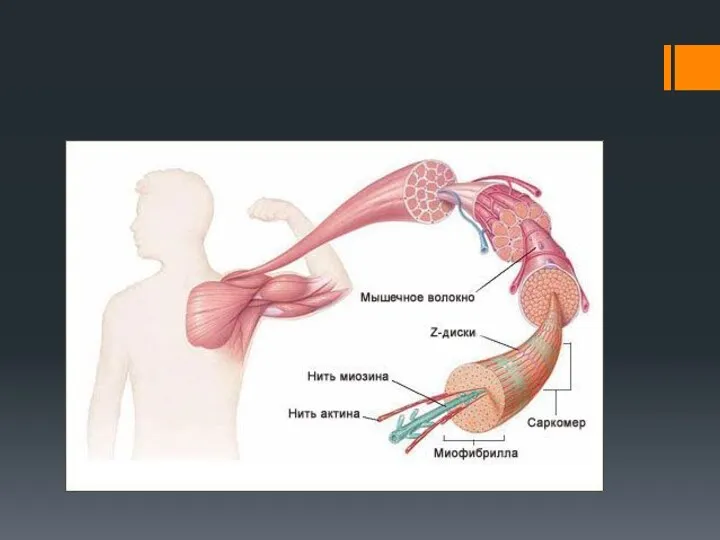
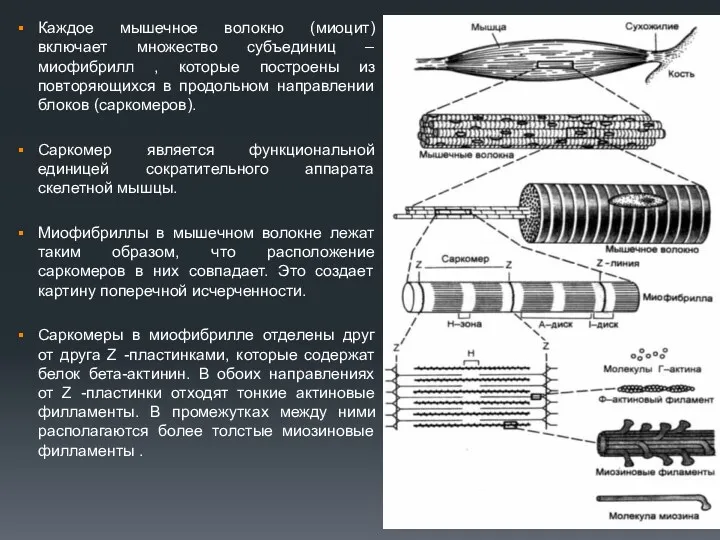
Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые
Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые
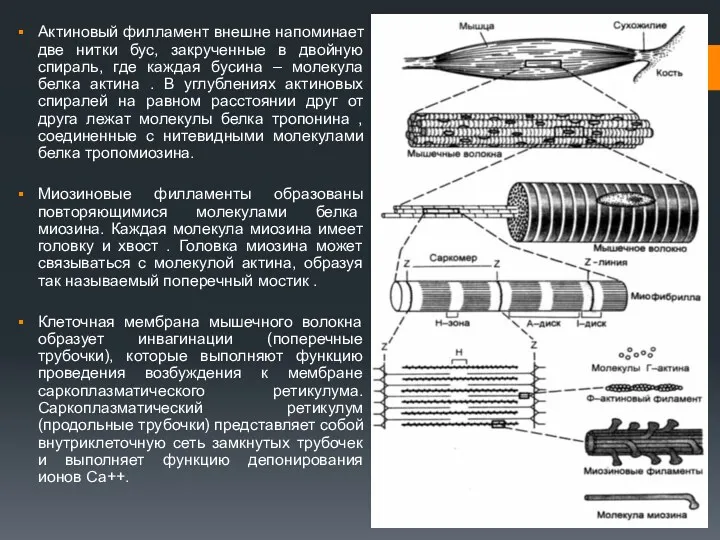
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль,
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль,
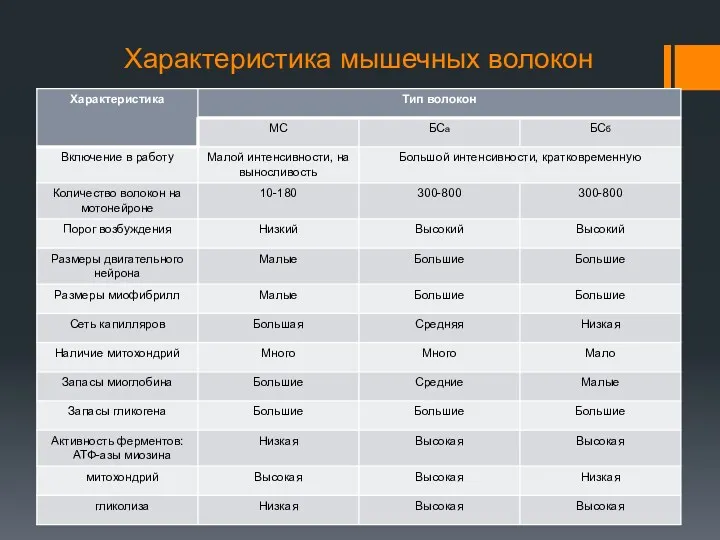
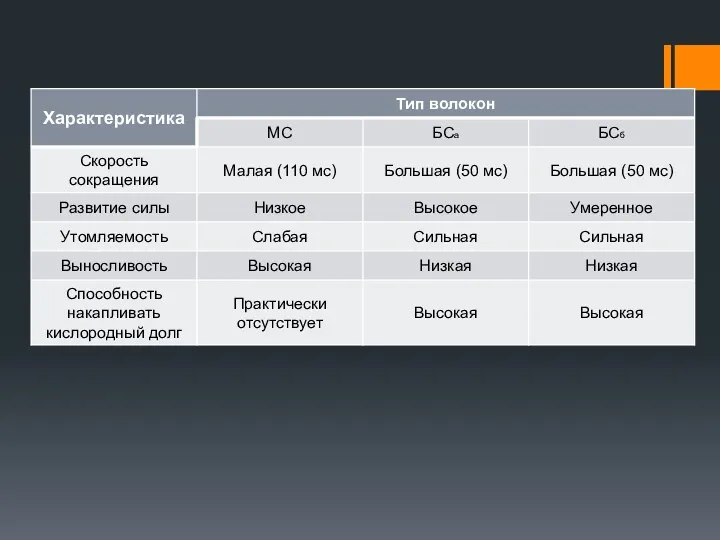
Характеристика мышечных волокон
Характеристика мышечных волокон
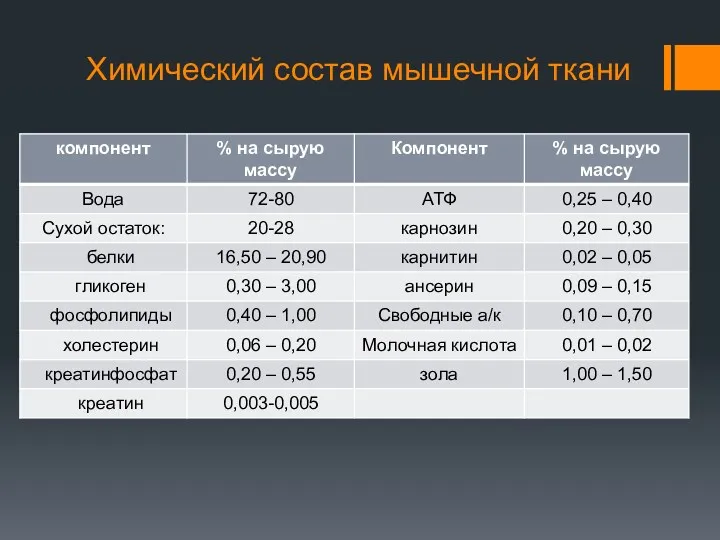
Химический состав мышечной ткани
Химический состав мышечной ткани

Основные белки мышц
1. Саркоплазматические – 35%.
Белки-ферменты митохондрий, катализируют процессы окислительного фосфорилирования;
Основные белки мышц
1. Саркоплазматические – 35%.
Белки-ферменты митохондрий, катализируют процессы окислительного фосфорилирования;
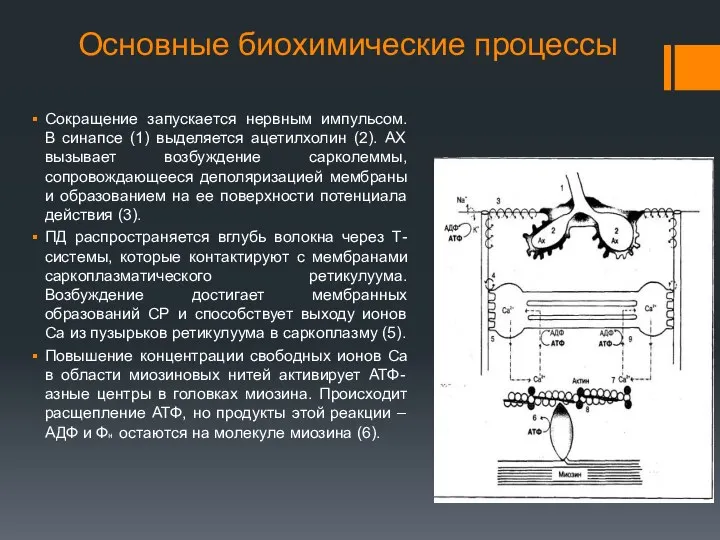
Основные биохимические процессы
Сокращение запускается нервным импульсом. В синапсе (1) выделяется
Основные биохимические процессы
Сокращение запускается нервным импульсом. В синапсе (1) выделяется
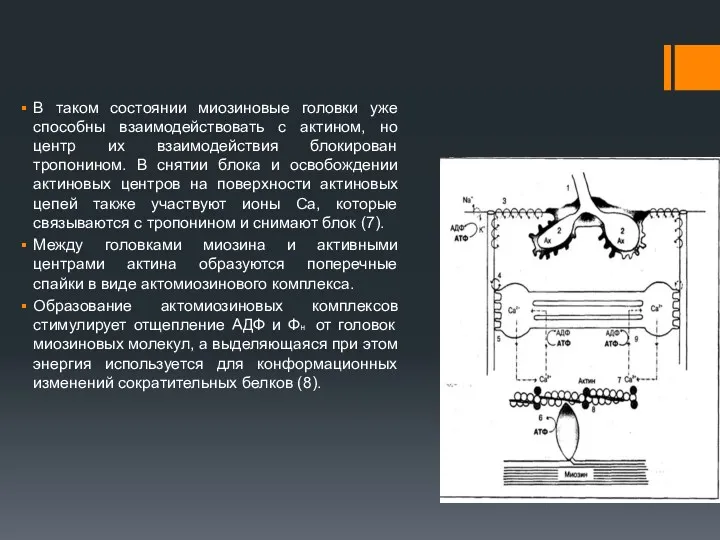
В таком состоянии миозиновые головки уже способны взаимодействовать с актином, но
В таком состоянии миозиновые головки уже способны взаимодействовать с актином, но
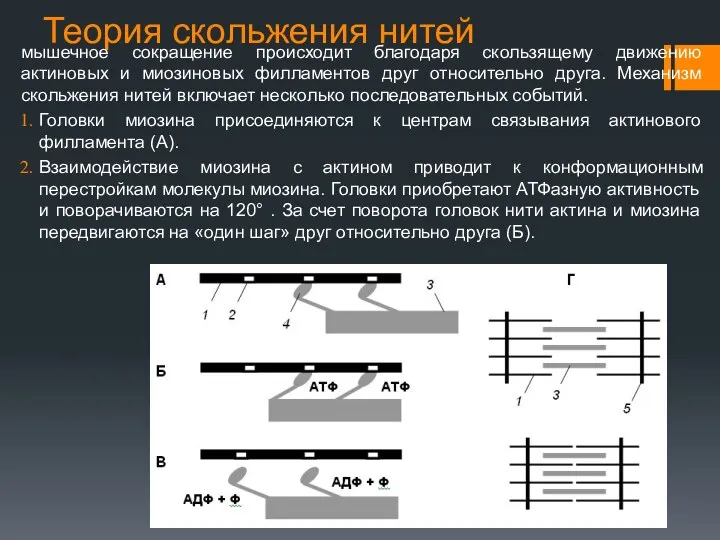
Теория скольжения нитей
мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых
Теория скольжения нитей
мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых

3. Рассоединение актина и миозина и восстановление конформации головки происходит в
3. Рассоединение актина и миозина и восстановление конформации головки происходит в

Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого

Функция АТФ в процессе сокращения и расслабления
В покоящейся мышце – препятствует
Функция АТФ в процессе сокращения и расслабления
В покоящейся мышце – препятствует
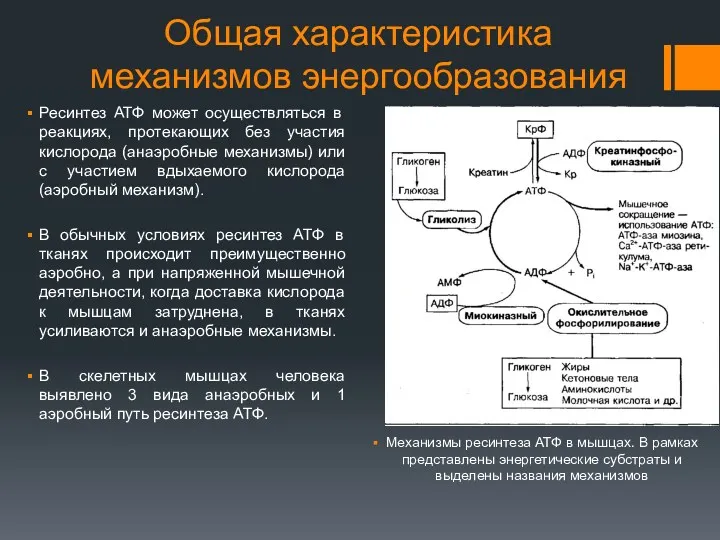
Общая характеристика механизмов энергообразования
Ресинтез АТФ может осуществляться в реакциях, протекающих без
Общая характеристика механизмов энергообразования
Ресинтез АТФ может осуществляться в реакциях, протекающих без

Анаэробные механизмы
Креатинфосфокиназный (фосфогенный или алактатный) механизм, обеспечивающий ресинтез АТФ за счет
Анаэробные механизмы
Креатинфосфокиназный (фосфогенный или алактатный) механизм, обеспечивающий ресинтез АТФ за счет

Аэробный механизм ресинтеза АТФ включает в основном реакции окислительного фосфорилирования, протекаемые
Аэробный механизм ресинтеза АТФ включает в основном реакции окислительного фосфорилирования, протекаемые
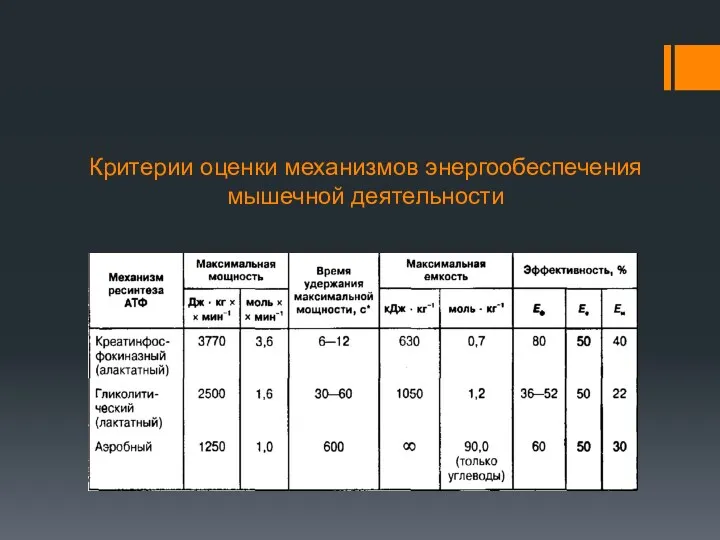
Критерии оценки механизмов энергообеспечения мышечной деятельности
Критерии оценки механизмов энергообеспечения мышечной деятельности

Креатинфосфокиназный механизм ресинтеза АТФ
Алактатный анаэробный механизм ресинтеза АТФ включает использование имеющейся
Креатинфосфокиназный механизм ресинтеза АТФ
Алактатный анаэробный механизм ресинтеза АТФ включает использование имеющейся

Максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5-0,7-й секунде интенсивной работы,
Максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5-0,7-й секунде интенсивной работы,

Максимальная мощность креатинфосфокиназной реакции составляет 3,8 кДж•кг-1•мин-1, что значительно выше мощности
Максимальная мощность креатинфосфокиназной реакции составляет 3,8 кДж•кг-1•мин-1, что значительно выше мощности

Запасы АТФ и Крф в скелетных мышцах человека
Запасы АТФ и Крф в скелетных мышцах человека
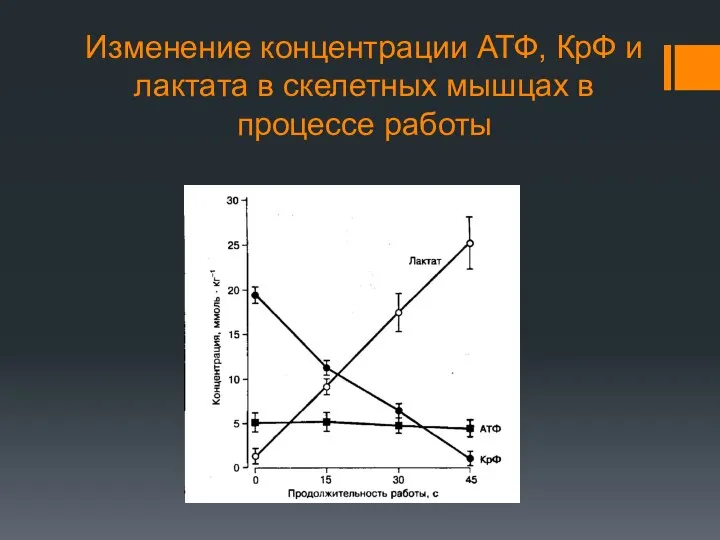
Изменение концентрации АТФ, КрФ и лактата в скелетных мышцах в процессе
Изменение концентрации АТФ, КрФ и лактата в скелетных мышцах в процессе

Содержание КрФ в скелетных мышцах увеличивается в процессе адаптации организма к
Содержание КрФ в скелетных мышцах увеличивается в процессе адаптации организма к

Гликолитический механизм ресинтеза АТФ
Как только в процессе анаэробной мышечной работы КФК-ный
Гликолитический механизм ресинтеза АТФ
Как только в процессе анаэробной мышечной работы КФК-ный
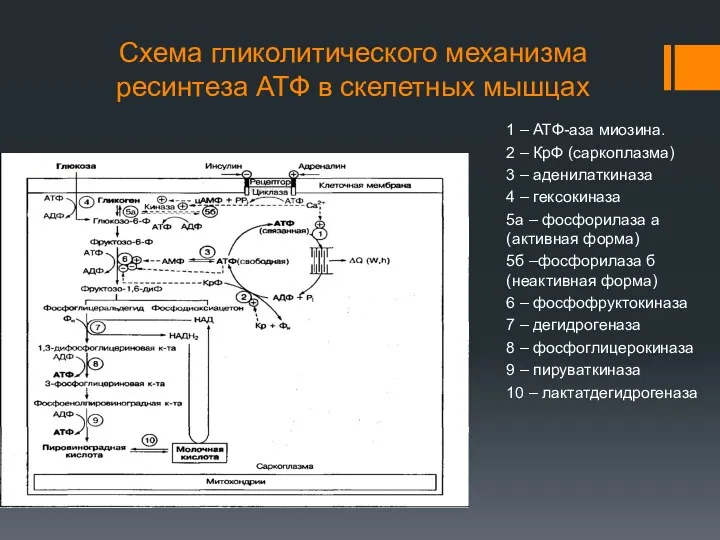
Схема гликолитического механизма ресинтеза АТФ в скелетных мышцах
1 – АТФ-аза миозина.
2
Схема гликолитического механизма ресинтеза АТФ в скелетных мышцах
1 – АТФ-аза миозина.
2

Максимальная мощность гликолиза у хорошо тренированных спортсменов может составлять 3,1 кДж•кг-1•мин-1,
Максимальная мощность гликолиза у хорошо тренированных спортсменов может составлять 3,1 кДж•кг-1•мин-1,
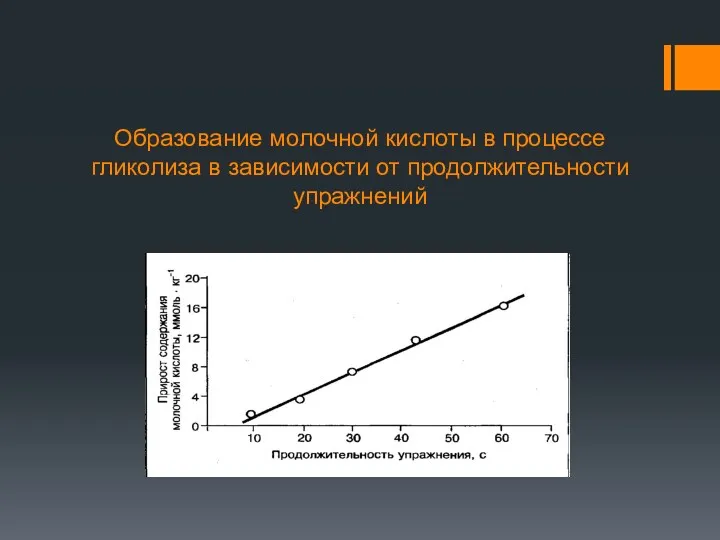
Образование молочной кислоты в процессе гликолиза в зависимости от продолжительности упражнений
Образование молочной кислоты в процессе гликолиза в зависимости от продолжительности упражнений

Молочная кислота может диффундировать через клеточные мембраны по градиенту концентрации и
Молочная кислота может диффундировать через клеточные мембраны по градиенту концентрации и
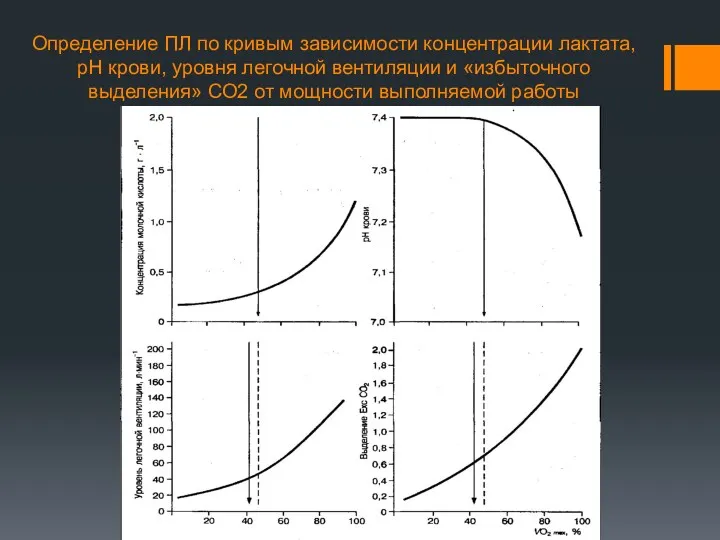
Определение ПЛ по кривым зависимости концентрации лактата, рН крови, уровня легочной
Определение ПЛ по кривым зависимости концентрации лактата, рН крови, уровня легочной
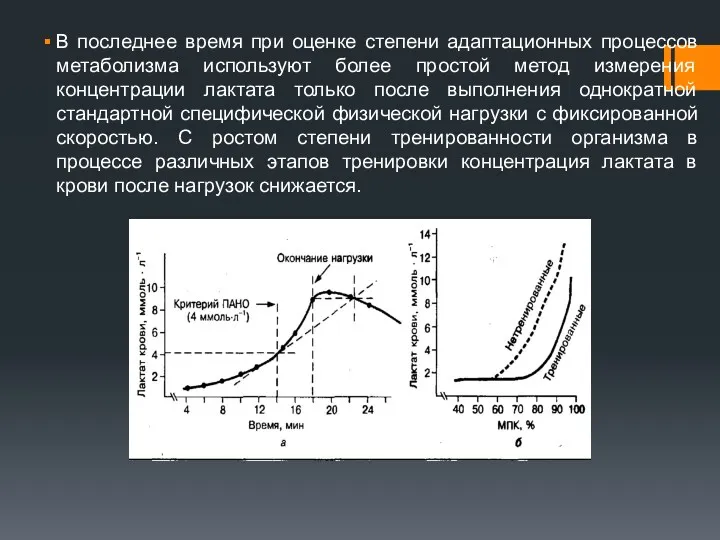
В последнее время при оценке степени адаптационных процессов метаболизма используют более
В последнее время при оценке степени адаптационных процессов метаболизма используют более
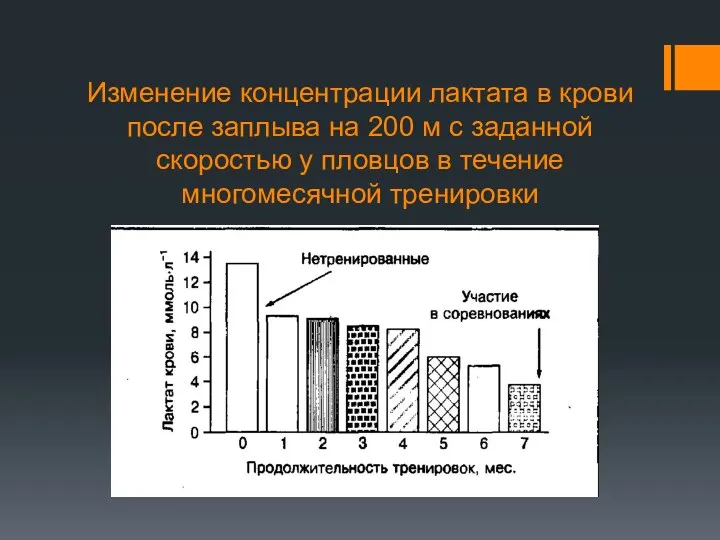
Изменение концентрации лактата в крови после заплыва на 200 м с
Изменение концентрации лактата в крови после заплыва на 200 м с

Миокиназный механизм
Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ
Миокиназный механизм
Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ

Аэробный механизм ресинтеза АТФ
Аэробный механизм ресинтеза АТФ в обычных условиях обеспечивает
Аэробный механизм ресинтеза АТФ
Аэробный механизм ресинтеза АТФ в обычных условиях обеспечивает
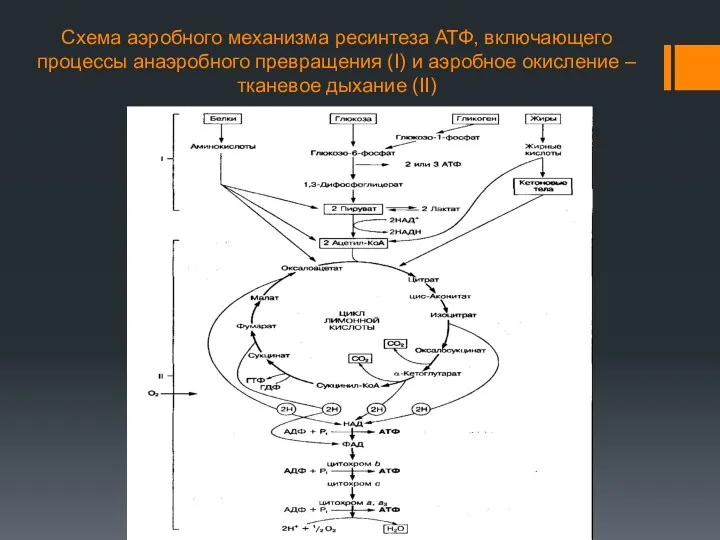
Схема аэробного механизма ресинтеза АТФ, включающего процессы анаэробного превращения (I) и
Схема аэробного механизма ресинтеза АТФ, включающего процессы анаэробного превращения (I) и

Максимальная мощность аэробного механизма наименьшая и составляет 1,2 кДж•кг-1•мин-1 и в

Максимальная мощность аэробного процесса достигается на 2-3-й минуте неинтенсивной работы у
Максимальная мощность аэробного процесса достигается на 2-3-й минуте неинтенсивной работы у

При окислении высших жирных кислот, например пальмитиновой, образуется еще больше энергии:
Эффективность
При окислении высших жирных кислот, например пальмитиновой, образуется еще больше энергии:
Эффективность

Подключение энергетических систем при различных физических нагрузках и их адаптация в
Подключение энергетических систем при различных физических нагрузках и их адаптация в
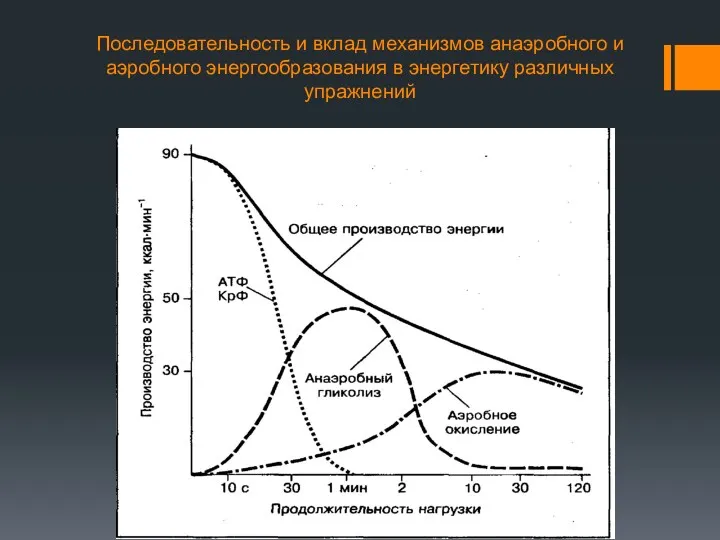
Последовательность и вклад механизмов анаэробного и аэробного энергообразования в энергетику различных
Последовательность и вклад механизмов анаэробного и аэробного энергообразования в энергетику различных
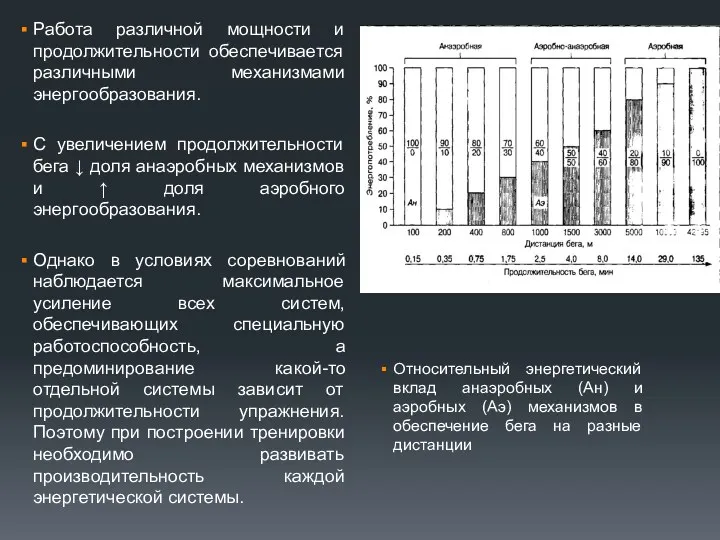
Работа различной мощности и продолжительности обеспечивается различными механизмами энергообразования.
С увеличением
Работа различной мощности и продолжительности обеспечивается различными механизмами энергообразования.
С увеличением

В спортивной практике физические упражнения, в которых вклад анаэробных алактатного и
В спортивной практике физические упражнения, в которых вклад анаэробных алактатного и
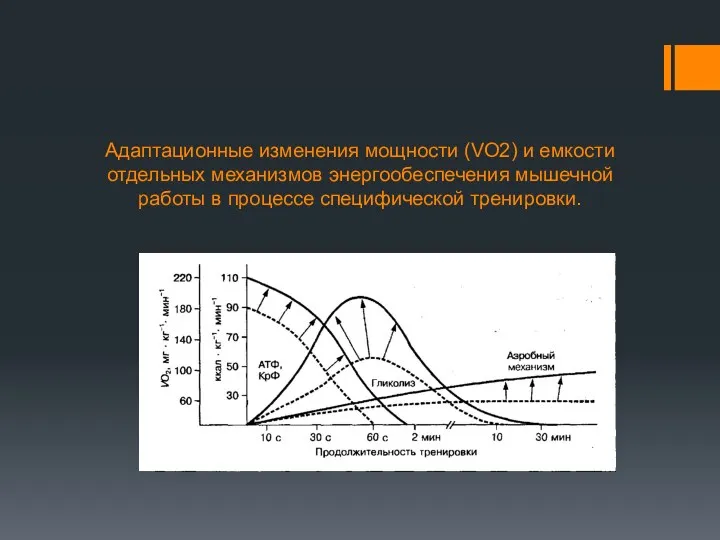
Адаптационные изменения мощности (VO2) и емкости отдельных механизмов энергообеспечения мышечной работы
Адаптационные изменения мощности (VO2) и емкости отдельных механизмов энергообеспечения мышечной работы

Мощность аэробного механизма, которая зависит от МПК и активности окислительных ферментов,
Мощность аэробного механизма, которая зависит от МПК и активности окислительных ферментов,

В процессе скоростной тренировки существенно изменяются анаэробные механизмы энергообеспечения. Отмечается ↑
В процессе скоростной тренировки существенно изменяются анаэробные механизмы энергообеспечения. Отмечается ↑
 Основные этапы развития животного мира на Земле
Основные этапы развития животного мира на Земле Картоптың алтын түстес жұмырқұрты
Картоптың алтын түстес жұмырқұрты Биология клетки в культуре. Клеточная и генная инженерия 2
Биология клетки в культуре. Клеточная и генная инженерия 2 Будова тварин. Клітини, тканини
Будова тварин. Клітини, тканини Зимующие птицы (для дошкольников)
Зимующие птицы (для дошкольников) Жасуша және оның құрылысы
Жасуша және оның құрылысы Parts of Human Skeleton
Parts of Human Skeleton Тип Членистоногие
Тип Членистоногие Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Продолговатый мозг и мост
Продолговатый мозг и мост Микроскопические грибы
Микроскопические грибы Біохімія гормонів
Біохімія гормонів Метаболизм сложных белков. (Тема 2)
Метаболизм сложных белков. (Тема 2) Вивчення захворювань і пошкоджень яблук та овочів
Вивчення захворювань і пошкоджень яблук та овочів Перенос веществ в организме беспозвоночных и позвоночных животных
Перенос веществ в организме беспозвоночных и позвоночных животных газообмен в легких
газообмен в легких Тепловые свойства почв
Тепловые свойства почв Гистология – учение о тканях. Классификация тканей
Гистология – учение о тканях. Классификация тканей Торможение условных рефлексов
Торможение условных рефлексов Межвидовая конкуренция
Межвидовая конкуренция Методическая разработка урока биологии по теме Обмен веществ 6 класс
Методическая разработка урока биологии по теме Обмен веществ 6 класс Жизнь организмов в морях и океанах
Жизнь организмов в морях и океанах Фенологические наблюдения за деревом
Фенологические наблюдения за деревом Птицы леса
Птицы леса История развития эволюционных идей
История развития эволюционных идей Су табиғаттың ерекше туындысы
Су табиғаттың ерекше туындысы Послепосадочный уход за деревьями и кустарниками
Послепосадочный уход за деревьями и кустарниками МОХОВИДНЫЕ 7 класс
МОХОВИДНЫЕ 7 класс