- Биохимия нервной, мышечной и соединительной ткани

Содержание
- 2. План лекции Нервная ткань: определение понятия, классификация по функциям и клеточному составу. Нейрон – как основная
- 3. Функции нервной системы: воспринимает информацию из внешней и внутренней среды; перерабатывает полученную информацию; хранит информацию; генерирует
- 4. Классификация нервной системы Анатомически Физиологически ЦНС ПНС Соматическая Автономная Симпатическая Парасимпатическая головной и спинной мозг периферические
- 5. Функциональной тканью нервной системы является нервная. Нервная ткань – это высокоспециализированная ткань, обладающая возбудимостью и проводимостью,
- 6. Нейрон
- 7. Нейроглия (от греческого glia – клей) это клетки нервной системы, которые не проводят нервные импульсы. Глиальные
- 8. Макроглия 1. Астроцитарная глия Астроциты обеспечивает микроокружение нейронов, выполняет опорную и трофическую функции в сером и
- 9. образует выстилку желудочков головного мозга входит в состав гематоликворного барьера. На снимке - просвет одного из
- 10. встречается в сером и белом веществе; обеспечивает барьерную функцию, участвует в формировании миелиновых оболочек нервных волокон,
- 11. Микроглия – специализированные макрофаги ЦНС. Способны к амёбоидным движениям и фагоцитозу Активизируются при воспалительных и дегенеративных
- 12. Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии (астроциты, олигодендроциты) в ЦНС называется
- 13. Химический состав нервной ткани
- 14. Белки нервной ткани сложные Простые :Нейроальбумины – основные растворимые белки (80%) Нейроглобулины - 5%. Катионные белки
- 15. Сложные белки Гликопротеины –нейрорецепция Протеолипиды – структурная ф -я Нейроспецифические белки Нейроспецифическая енолаза (Белок 14-3-2) -
- 16. Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в астроцитах) и интенсивно нарабатывается в
- 17. Опиоидные нейропептиды
- 18. нейропептиды
- 19. Нейротрофические факторы:
- 20. Ферменты Нейроспецифическая енолаза ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии), АСТ, альдолаза, креатинкиназа (ВВ),
- 21. Аминокислоты Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ Аминокислот в мозге в среднем 34
- 22. Липиды нервной ткани фосфоглицериды в сером веществе составляют более 60% от всех липидов, а в белом
- 23. Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез ганглиозидов связан с дифференциацией нейронов.
- 24. Углеводы нервной ткани много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей индивидуальность и специфичность. мало
- 25. Нуклеотиды нервной ткани Клетки Пуркинье мозжечка содержат избыточное количество ДНК. Необычно короткие нуклеосомные единицы, наличие редких
- 26. Макроэргические соединения нервной ткани Содержание креатина и креатинфосфата более, чем в 2 раза превышает количество адениновых
- 27. Минеральные вещества нервной ткани Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+ распределены в головном мозге
- 28. Строение нервного волокна. Миелиновая оболочка 1 2 3 1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои) 5-липиды;
- 29. Образован: клетками эндотелия капилляров плотной базальной мембраной, не имеющей пор. Астроглией, выстилающие наружную поверхность эндотелия. Р-ГП
- 30. Особенности метаболизма нервной ткани Для мозга характерна высокая интенсивность энергетического обмена с преобладанием аэробных процессов. Головной
- 31. Основной энергетический субстрат для нервной ткани - глюкоза За 1 минуту 100 г ткани мозга потребляют
- 32. Обмен белков и аминокислот нервной ткани Нервная ткань характеризуется высоким обменом аминокислот и белков. Максимальная скорость
- 33. Обмен азота нервной ткани Источником аммиака в головном мозге служит непрямое дезаминирование аминокислот с участием глутаматдегидрогеназы,
- 34. Липидный обмен нервной ткани Особенность: липиды не используются в качестве энергетического материала, а в основном идут
- 35. Механизмы передачи нервного импульса
- 36. Механизмы передачи нервного импульса по нервному волокну 1. Потенциал покоя (-70 мВ) Na-канал К-канал 2. Потенциал
- 37. Са Na K Синтез медиатора Загрузка в везикулу Слияние везикул Специфические рецепторы Открытие Na канала Проведение
- 38. Аминокислотные медиаторы ингибиторные нейтральные (ГАМК, глицин, β-аланин и таурин). возбуждающие кислые (глутамат и аспартат) Глицин открывает
- 39. открывает хлорные каналы вызывает гиперполяризацию тормозит возбудимость постсинаптической мембраны ГАМК
- 40. Синтез холина Печень Нейроны
- 41. Синтез и распад медиатора на примере ацетилхолина (CH3)3N-CH2-CH2-OH холинацетилтрансфераза (CH3)3N-CH2-CH2-O-CO-CH3 H2O ацетилхолинэстераза (CH3)3N-CH2-CH2OH Нейрон ПЕЧЕНЬ CH3-CO-S-KoA
- 42. Рецепторы Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри клетки, которые, взаимодействуя с
- 43. Биохимические синдромы в психиатрии Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина) Нарушения окислительно-восстановительных процессов (гипоксия, накопление молочной к-ты,
- 44. Патобиохимия нервной системы 1) Миастения - уменьшение числа холинорецепторов 3) болезнь Паркинсона - дегенерация дофамин содержащих
- 45. Нейроспецифические белки- МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ белок S100 Нейрон специфическая енолаза (NSE) Нейроны Астроциты Олигодендроциты и миелин
- 46. Нейронспецифическая енолаза (NSE) Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент, катализирующий превращение 2-фосфоглицерата в
- 47. Белок S100 S-100 - кальций-связывающий протеин. Локализация мембраны, цитоплазма. участвует в фосфорилировании белка, обеспечивает работу цитоскелета
- 48. антитела к S100β являются специфичными маркерами повреждения астроцитарной глии. Ген S100 находиться в длинном плече 21-й
- 49. Основной белок миелина (MBP) MBP - щелочной белок, с высоким содержанием (25%) основных аминокислот (аргинина, лизина,
- 50. NSE, S100 и MBP – маркеры повреждения мозговой ткани Нейроспецифические белки в нормальных условиях обнаруживаются в
- 51. NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также других опухолей нейроэктодермального или нейроэндокринного
- 52. концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени, чем у женщин NSE, S100
- 53. Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции в ряде периферических органов (
- 54. Катаболизм дофамина: Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»). В везикулу с помощью протон-зависимой
- 55. ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА Различают семь отдельных подсистем (первые три являются основными): 1. Нигростриатная 2. Мезокортикальная
- 56. Пути Дофамина: Патология: Патология Патология
- 57. Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы, ДОФА-декарбоксилазы. Патология Дофамина. Болезнь
- 58. Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы, ДОФА-декарбоксилазы. Патология Дофамина. Болезнь
- 59. План лекции Функции мышц Классификация мышечных волокон Химический состав мышц Особенности обмена веществ и энергии в
- 60. Мышечное волокно Функциональной единицей мышечной ткани является мышечное волокно Мышечное волокно поперечнополосатой мышцы- это многоядерная клетка.
- 61. Двигательная единица – это совокупность образований – нейрон и все мышечные волокна (обычно 10-1000), которые этот
- 62. ХИМИЧЕСКИЙ СОСТАВ МЫШЕЧНОЙ ТКАНИ Вода............................................72—80 Сухой остаток …........................20—28 В том числе: Белки...........................................16,5—20,9 Гликоген......................................0,3—3,0 Фосфатиды.................................0,4—1,0 Холестерин.................................0,06—0,2 Креатин
- 63. 1. Сократительные (миофибриллярные) белки миозин 55% актин 25% тропомиозин (во всех мышцах) тропонины Т, I и
- 64. Углеводы мышечной ткани Гликоген: 0,3-3,0% ГАГ моносахариды глюкоза, фруктоза Липиды мышечной ткани Фосфолипиды (в миокарде больше)
- 65. ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ В МЫШЕЧНОЙ ТКАНИ Обмен белков и аминокислот Мышцы характеризуются высоким обменом белков и
- 66. Мышцы также синтезируют и выделяют много аланина и глутамина. В реакций трансаминирования вступают ПВК (гликолиз), α-КГ
- 67. Аминокислоты Белки При интенсивной работе для нейтрализации лактата мышцы выделяют аммиак. В мышцах низкая активность глу-ДГ,
- 68. Липидный обмен преобладает катаболизм липидов. Жирные кислоты, кетоновые тела в аэробных условиях окисляются в мышцах для
- 69. Углеводный обмен преобладает катаболизм углеводов. глюкоза окисляется в аэробных или анаэробных условиях для синтеза АТФ. Из
- 70. Энергетический обмен Энергетический обмен в состоянии покоя очень низкий, при интенсивной физической нагрузке значительно возрастает. В
- 71. Мышцы, которые могут длительно находиться в состоянии сокращения, способны резервировать кислород в миоглобине. За счет миоглобина,
- 72. Основной потребитель АТФ - процесс мышечного сокращения. Запасы АТФ быстро истощаются. Источники АТФ: 1) классический путь
- 73. (Окислительное фосфорилирование) АТФ АДФ-АТФ-транслоказа(ААТ) ВНУТРЕННЯЯ МЕМБРАНА МИТОХОНДРИЙ КРЕАТИН КРЕАТИН ФОСФАТ АТФ АДФ САРКОЛЕММА МИОФИБРИЛЛА КРЕАТИН ФОСФАТ
- 74. Креатинфосфатный челнок предотвращает быстрое истощение запасов АТФ в мышце за счет: запаса макроэргических связей в креатинфосфате
- 75. Характеристика быстрых и медленных скелетных мышц
- 76. Миофибрилла
- 77. МИОФИБРИЛЛА Миофибрилла — это цилиндрическое образование толщиной 1-2 мкм, простирающиеся на всю длину мышечного волокна. Миофибрилла
- 78. 1. Тонкие филаменты Тонкие филаменты у гладких мышц образованы F-актином и тропомиозином, У поперечнополосатых - F-актином,
- 79. Актин G-актин - мономерный (глобулярный) белок с массой 43кДа. F-актин. При физиологической величине рН и в
- 80. Тропомиозин - есть во всех мышцах: белок, состоящий из а и р цепей, располагается в щели
- 81. 2. Толстые филаменты образованы миозином, размер 16нм. располагаются в центре саркомера, между «тонкими» филаментами. На поверхности
- 82. Миозин Асимметричный гексамер с мол. массой 460кДа, состоит из 2 тяжелых (Н) и 4 легких (L)
- 83. 3. Z-диски Саркомер ограничен с двух сторон Z-дисками (α-актинин). К Z-дискам присоединены «тонкие» филаменты Z-ДИСК (α-АКТИНИН)
- 84. Строение саркомера Диск А (анизотропная зона)= темный участок –образован «толстыми» нитями миозина. Зона Н - центральная
- 85. Полоса М - пересекает центральную область диска А, образована толстыми нитями, в которых миозин не имеет
- 86. Вследствие движения уменьшается длина каждого саркомера (укорачиваются Н-зона и I-диски) и всей мышцы в целом. При
- 87. В скелетной мышечной ткани саркомеры миофибрилл располагаются параллельно. За счет этого на срезах скелетные мышцы выглядят
- 88. Миозиновая головка может спонтанно гидролизовать АТФ до АДФ и Фн, которые при этом остаются в составе
- 89. При достижении нужного положения миозиновая головка с АТФ или АДФ и Фн может связываться с F-актином,
- 90. У АДФ и Фн низкое сродство к актин-миозиновому комплексу, поэтому они от него отделяются. При этом
- 91. Новая молекула АТФ присоединяется к актин-миозиновому комплексу. Комплекс актин-миозин-АТФ обладает низким сродством к актину, поэтому миозиновая
- 92. Механизм мышечного сокращения
- 93. Регуляция сокращения и расслабления мышц Любое мышечное сокращение опосредуется Са2+. Кальциевые насосы постоянно перекачивают Са2+ из
- 94. Са-регуляция сокращение При действии ацетилхолина на ацетилхолиновые рецепторы происходит возникновение потенциала действия. Он через Т-систему достигает
- 95. Актиновая регуляция Характерна для поперечнополосатых мышц. Мышечное сокращение ингибирует тропомиозиновая система на 2 стадии сокращения: TпI
- 96. Поступающий в саркоплазму Са2+ присоединяется к тропонину ТnС. Комплекс ТnС•Са2+ реагирует с TnI и ТnТ, влияя
- 97. Расслабление происходит, когда: 1) При перекачке Са2+ в ЭПР его содержание в саркоплазме падает ниже 10-7
- 98. Миозиновая регуляция Характерна для гладких мышц. нет тропониновой системы, легкая цепь (р-цепь) миозина подавляет его АТФ-азную
- 99. Расслабление гладких мышц происходит, когда: 1) содержание ионов Са2+ в саркоплазме падает ниже 10-7 моль/л 2)
- 100. Биохимические показатели крови и мочи, отражающие функциональное состояние мышечной ткани Аминотрансферазы -диагностика патологии печени и миокарда.
- 101. Тропонин Т -маркер инфаркта миокарда в острой и подострой фазе. Миоглобин -маркер деструктивных изменений в мышечной
- 102. Миопатии (греч. mys, myos мышца + pathos страдание, болезнь) - нервно-мышечные заболевания, характеризующиеся развитием первичного дистрофического
- 104. БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ В МЫШЦАХ ПРИ ПАТОЛОГИИ: снижение содержания миофибриллярных белков возрастание концентрации белков стромы и некоторых
- 105. нарушение метаболизма креатина и его усиленное выделение с мочой (креатинурия). креатинурия является результатом нарушения в скелетной
- 106. Изменение активности ферментов в мышцах: уменьшается активность ферментов, локализованных в саркоплазме изменяется активность ферментов, связанных с
- 107. Ишемическая болезнь сердца ИБС - патологическое состояние, характеризующееся абсолютным или относительным нарушением кровоснабжения миокарда. Причины: обменные
- 108. 1) первичная остановка кровообращения; 2) стенокардия; 3) инфаркт миокарда; 4) сердечная недостаточность; 5) аритмии. Классификация ишемической
- 109. Причины инфаркта миокарда Атеросклероз коронарных артерий Эмболы, тромбы коронарной артерии Спазм коронарных артерий и резко повышение
- 110. Биохимические изменения при ИБС гипоксия Снижение: активности ферментов аэробного дыхания, синтеза клеточных структур, транспорта субстратов, обмена
- 111. норма Некроз через 2 часа Некроз через 18 часов Неблагоприятные факторы: ↓АТФ ↑ ПОЛ Ацидоз Дисбаланс
- 112. Диагностика ЭКГ
- 113. При гипоксии миокард поглощает ТГ из липопротеинов, которые не используются, а накапливаются, приводя к ожирению миокарда.
- 114. Терапия
- 115. План лекции Биохимия межклеточного матрикса гетерополисахариды структурные и адгезивные белки Минерализованная соединительная ткань. Ремоделирование костной ткани.
- 116. Межклеточный матрикс Гетерополисахариды и протеогликаны Гликозамингликаны гиалуроновая кислота хондроитинсульфат дерматансульфат гепарансульфат гепарин кератансульфат Протеогликаны (гликозамингликаны +
- 117. Гликозамингликаны, или мукополисахариды (ГАГ) Линейные отрицательно заряженные гетерополисахариды, состоящие из повторяющихся дисахаридных звеньев (см. след. слайд):
- 118. Дисахаридная единица гиалуроновой кислоты (всего несколько тыс. дисахаридных единиц) Дисахаридная единица дерматансульфата Структуру других ГАГ для
- 119. Особенности строения отдельных ГАГ Кератансульфаты вместо гексуроновой кислоты содержат галактозу и кроме дисахаридных единиц –фукозу, маннозу,
- 120. Функции гликозамингликанов и протеогликанов Структурный компонент межклеточного матрикса Формируют тургор тканей (гиалуроновая кислота) «Смазка» в суставах
- 121. Протеогликаны – основное вещество межклеточного матрикса Состав протеогликанов: Гликозамингликаны (кроме гиалуроновой кислоты) – 90-95% Белковая часть
- 122. Протеогликаны: связь гликозамингликана с белком А и Б – дисахаридная единица гликозамингликана
- 123. Синтез протеогликанов 1. Синтез белковой части 2. Образование линкерного трисахарида 3. Рост цепи гликозамингликана Реакции 2
- 124. Синтез хондроитинсульфата
- 125. Катаболизм гликозамингликанов и протеогликанов Полупериод жизни: 3 – 10 дней (кератансульфат 120 дней) Осуществляют лизосомные протеазы,
- 126. Структурные белки соединительной ткани: Коллаген Коллагены – семейство полиморфных фибриллярных белков (19 типов коллагена), имеющих трехспиральную
- 127. Коллаген: особенности строения Пептидная левозакрученная α-спираль коллагена содержит 600 – 3000 АК (в отличие от α-спирали
- 128. Синтез и созревание коллагена 1. Внутриклеточный этап (фибробласты, хондробласты, остеобласты): образование пре-про-α-цепи, содержащей сигнальный пептид, облегчающий
- 130. Гидроксилирование пролина и лизина осуществляют монооксигеназы (гидроксилазы). Кофактор – Fe2+ . Для поддержания восстановленной формы железа
- 131. Гликозилирование ГидЛиз путем присоединения остатков глюкозы и галактозы осуществляют гликозилтрансферазы, использующие в качестве субстратов активные формы
- 133. Дезаминирование лизина и гидроксилизина с образованием реакционноспособного альдегида – аллизина и гидроксиаллизина для формирования ковалентных сшивок
- 134. Макрофибрилла коллагена
- 135. Зрелое коллагеновое волокно толщиной в 1 мм выдерживает нагрузку до 10 кг
- 136. Катаболизм коллагена Коллаген – медленно обменивающийся белок (время полужизни – недели, месяцы). Обмен более активен у
- 137. Регуляция обмена коллагена Аскорбиновая кислота стимулирует синтеза коллагена, протеогликанов и пролиферацию фибробластов Половые гормоны регулируют синтез
- 138. При старении увеличивается количество поперечных сшивок между молекулами тропоколлагена, затрудняющих действие коллагеназы уменьшается соотношение гликозамингликаны /
- 139. Метаболизм коллагена При патологии Нарушение катаболизма приводит к фиброзу органов Усиление распада происходит при аутоиммунных заболеваниях
- 140. Структурные белки соединительной ткани: Эластин Основной компонент волокон тканей, обладающих значительной эластичностью (сосуды, связки, легкие) 70
- 141. Эластин Время полураспада – 75 лет Протеолиз под действием эластазы (нейтрофилы) Нарушения образования эластина (в частности
- 142. Адгезивные белки соединительной ткани: фибронектин Димер (2 цепи сходны, но не идентичны, соединены дисульфидными мостиками), гликопротеин,
- 143. Структура фибронектина
- 144. Адгезивные белки соединительной ткани: ламинин Основной гликопротеин базальных мембран 3 полипептидные цепи, имеющие доменное строение для
- 145. Адгезивные белки соединительной ткани: нидоген Сульфатированный гликопротеин базальных мембран Одна полипептидная цепь Три глобулярных домена: центр
- 146. Кость – минерализованная соединительная ткань Функции: размер и форма тела, защита внутренних органов, минеральный гомеостаз, кроветворение
- 147. Костный обмен Костный обмен (ремоделирование) включает: резорбцию (разрушение) костной ткани костеобразование (обновление, реорганизация органической матрицы и
- 148. Минеральный состав и строение апатитов Ячейка гидроксиапатита Кристалл гидроксиапатита состоит из 2000 ячеек Гидроксиапатит кальция придает
- 149. Строение апатитов Оптимальное расстояние между ионами Са2+ и РО43- задается органической матрицей. Изменение строения компонента матрицы
- 150. Органические вещества кости: коллаген Особенности строения коллагена I типа (остеоколлагена): Больше гидроксипролина, но меньше гидроксилизина Менее
- 151. Органические вещества кости: неколлагеновые белки Большинство неколлагеновых белков – гликопротеины или гликофосфопротеины; присутствуют и в других
- 152. Органические вещества кости: неколлагеновые белки Неколлагеновые белки, содержащие RGD-последовательность: -Арг-Глу-Асп- (комплементарна рецепторам остеокластов): Сиалопротеин, остеопонтин –
- 153. Другие органические вещества кости Факторы роста и дифференцировки (IGF-1, TGF-β, PDGF) Протеогликаны (ГАГ: хондроитин-, дерматан-, кератансульфаты)
- 154. Прикрепление ЩФ к мембране остеобластов с помощью инозитолфосфатного якоря
- 155. Роль щелочной фосфатазы в костной ткани Проявляет активность на стадии минерализации (рН 9,6) Катализирует реакцию дефосфорилирования
- 156. Ремоделирование костной ткани Каждые 10 лет костная ткань заменяется новой (ежегодно обновляется 5-10% скелета) Интенсивность обмена
- 157. Этапы цикла ремоделирования костной ткани 1. Активация: остеобласты под влиянием стимулирующих факторов секретируют фактор RANKL (receptor
- 158. Этапы цикла ремоделирования костной ткани 2. Резорбция: «работа» остеокластов Прикрепление остеокласта к кости Карбоангидразная реакция и
- 159. Активация остеокластов в процессе костной резорбции
- 160. Этапы цикла ремоделирования костной ткани 3. Реверсия Действие локальных ростовых факторов на остеобласты приводит к синтезу
- 161. Регуляция ремоделирования Паракринно-аутокринная (местная) регуляция: цитокины, фаторы роста, простагландины костной ткани Системная регуляция Основные регуляторы: Паратгормон
- 162. Регуляция ремоделирования
- 163. Регуляция ремоделирования
- 164. Регуляция ремоделирования ПК А фосфорилирует белок-переносчик Са2+, что приводит к поступлению Са2+ в цитозоль остеокласта из
- 165. Регуляция ремоделирования Кортизол замедляет костеобразование: подавляет пролиферацию остеобластов, синтез остеоколлагена, остеонектина, протеогликанов Кортизол стимулирует костную резорбцию:
- 166. Регуляция ремоделирования Гормон роста повышает активность остеобластов, которые стимулируют остеокласты, поэтому происходит очаговая деминерализация, затем увеличение
- 167. Регуляция ремоделирования В остеобластах эстрогены стимулируют синтез остеоколлагена, остеонектина, ЩФ. Вызывают раннюю минерализацию, поэтому в период
- 169. Скачать презентацию

План лекции
Нервная ткань: определение понятия, классификация по функциям и клеточному составу.
Нейрон
План лекции
Нервная ткань: определение понятия, классификация по функциям и клеточному составу.
Нейрон

Функции нервной системы:
воспринимает информацию из внешней и внутренней среды;
перерабатывает полученную информацию;
хранит
Функции нервной системы:
воспринимает информацию из внешней и внутренней среды;
перерабатывает полученную информацию;
хранит
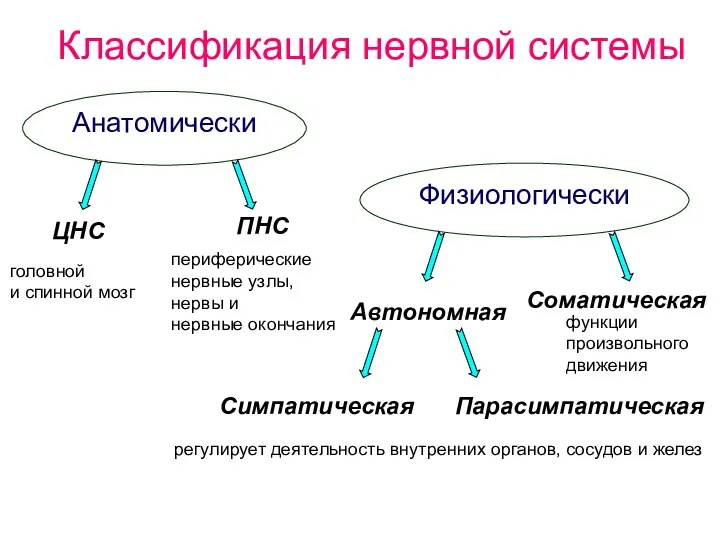
Классификация нервной системы
Анатомически
Физиологически
ЦНС
ПНС
Соматическая
Автономная
Симпатическая
Парасимпатическая
головной
и спинной
Классификация нервной системы
Анатомически
Физиологически
ЦНС
ПНС
Соматическая
Автономная
Симпатическая
Парасимпатическая
головной
и спинной

Функциональной тканью нервной системы является нервная.
Нервная ткань – это высокоспециализированная
Функциональной тканью нервной системы является нервная.
Нервная ткань – это высокоспециализированная

Нейрон
Нейрон

Нейроглия (от греческого glia – клей) это клетки нервной системы, которые
Нейроглия (от греческого glia – клей) это клетки нервной системы, которые
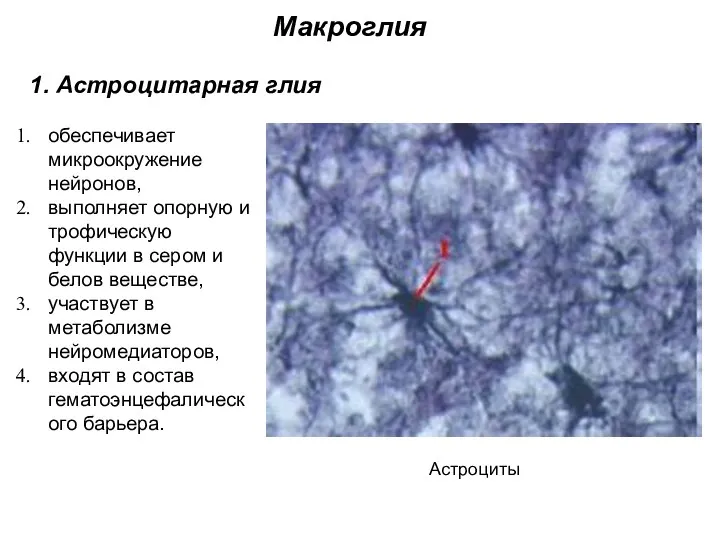
Макроглия
1. Астроцитарная глия
Астроциты
обеспечивает микроокружение нейронов,
выполняет опорную и трофическую функции
Макроглия
1. Астроцитарная глия
Астроциты
обеспечивает микроокружение нейронов,
выполняет опорную и трофическую функции
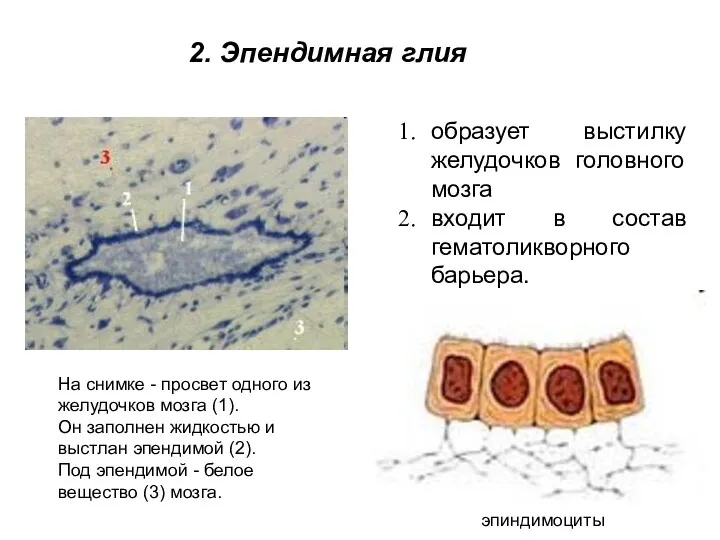
образует выстилку желудочков головного мозга
входит в состав гематоликворного барьера.
На снимке
образует выстилку желудочков головного мозга
входит в состав гематоликворного барьера.
На снимке
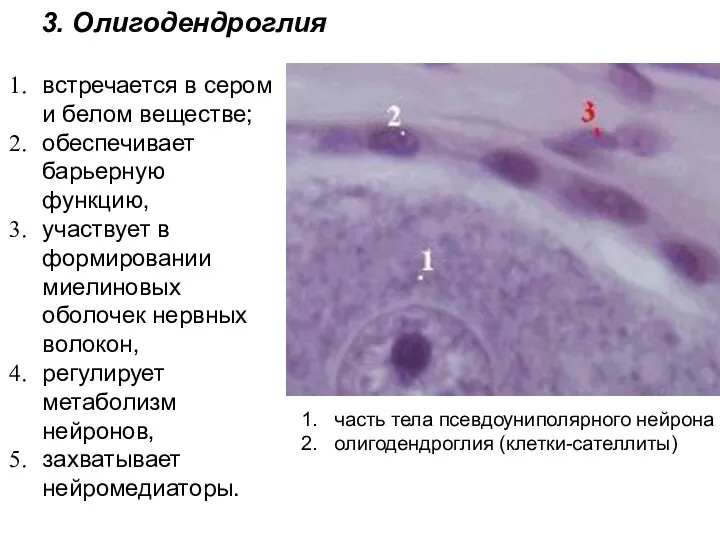
встречается в сером и белом веществе;
обеспечивает барьерную функцию,
участвует в
встречается в сером и белом веществе;
обеспечивает барьерную функцию,
участвует в

Микроглия – специализированные макрофаги ЦНС.
Способны к амёбоидным движениям и фагоцитозу
Активизируются
Микроглия – специализированные макрофаги ЦНС.
Способны к амёбоидным движениям и фагоцитозу
Активизируются

Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии
Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии
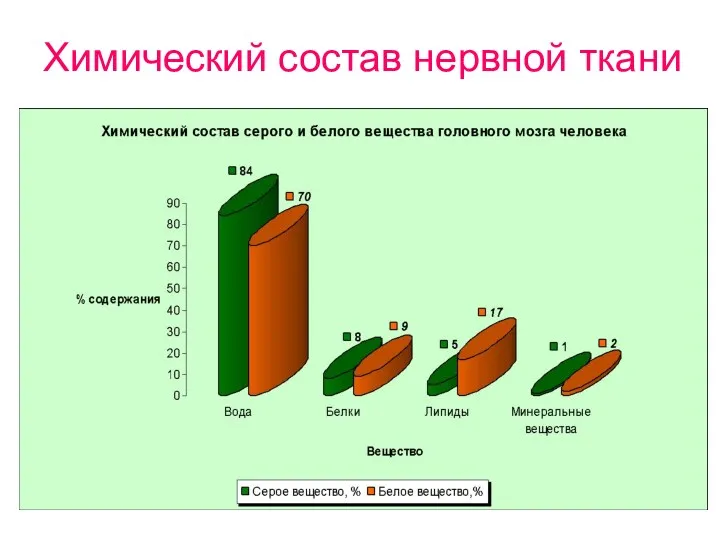
Химический состав нервной ткани
Химический состав нервной ткани
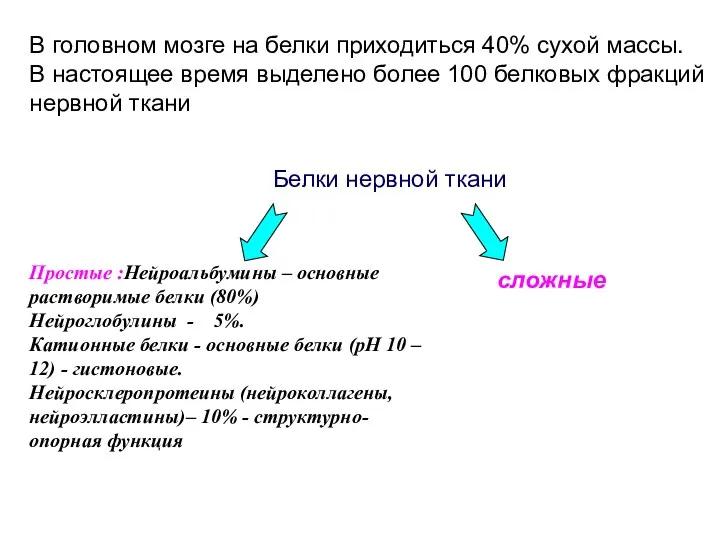
Белки нервной ткани
сложные
Простые :Нейроальбумины – основные растворимые белки (80%)
Нейроглобулины -
Белки нервной ткани
сложные
Простые :Нейроальбумины – основные растворимые белки (80%)
Нейроглобулины -

Сложные белки
Гликопротеины –нейрорецепция
Протеолипиды – структурная ф -я
Нейроспецифические белки
Нейроспецифическая енолаза (Белок
Сложные белки
Гликопротеины –нейрорецепция
Протеолипиды – структурная ф -я
Нейроспецифические белки
Нейроспецифическая енолаза (Белок

Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в
Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в

Опиоидные нейропептиды
Опиоидные нейропептиды
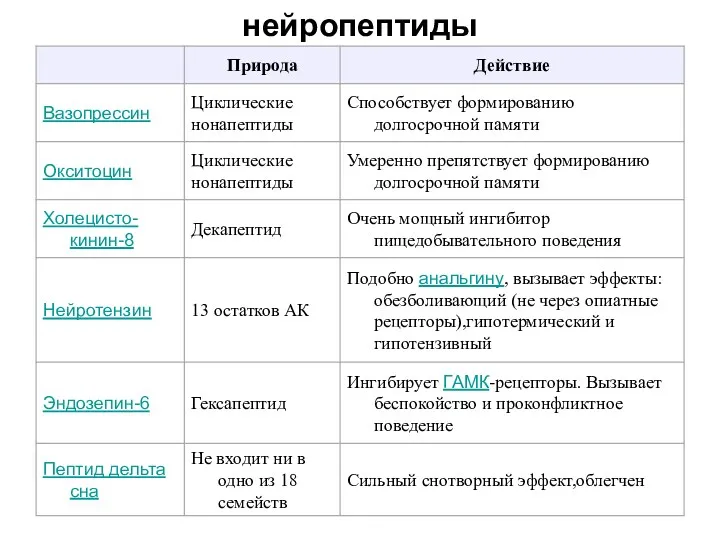
нейропептиды
нейропептиды
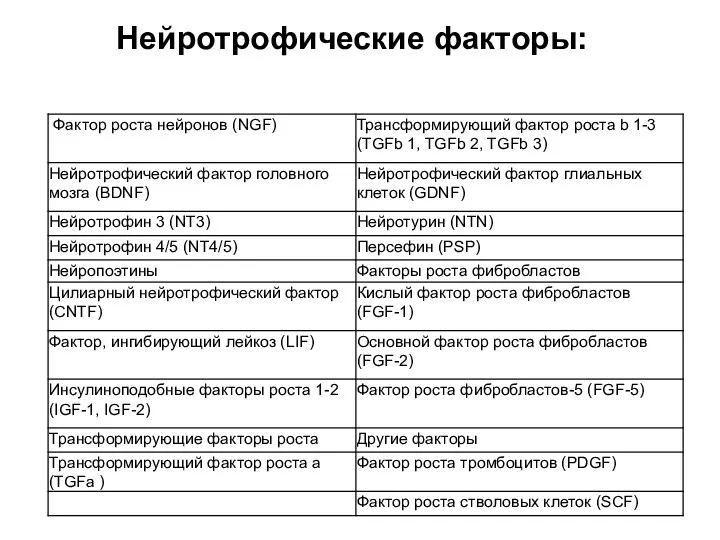
Нейротрофические факторы:
Нейротрофические факторы:

Ферменты
Нейроспецифическая енолаза
ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии),
Ферменты
Нейроспецифическая енолаза
ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии),
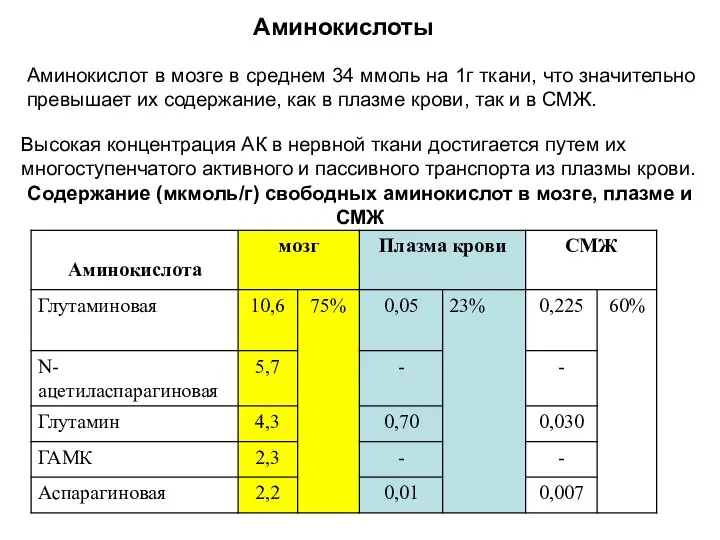
Аминокислоты
Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ
Аминокислот в мозге
Аминокислоты
Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ
Аминокислот в мозге

Липиды нервной ткани
фосфоглицериды в сером веществе составляют более 60% от
Липиды нервной ткани
фосфоглицериды в сером веществе составляют более 60% от

Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез
Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез

Углеводы нервной ткани
много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей
Углеводы нервной ткани
много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей

Нуклеотиды нервной ткани
Клетки Пуркинье мозжечка содержат избыточное количество ДНК.
Необычно короткие нуклеосомные
Нуклеотиды нервной ткани
Клетки Пуркинье мозжечка содержат избыточное количество ДНК.
Необычно короткие нуклеосомные

Макроэргические соединения нервной ткани
Содержание креатина и креатинфосфата более, чем в 2
Макроэргические соединения нервной ткани
Содержание креатина и креатинфосфата более, чем в 2

Минеральные вещества нервной ткани
Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+
Минеральные вещества нервной ткани
Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+

Строение нервного волокна. Миелиновая оболочка
1
2
3
1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои)
Строение нервного волокна. Миелиновая оболочка
1
2
3
1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои)
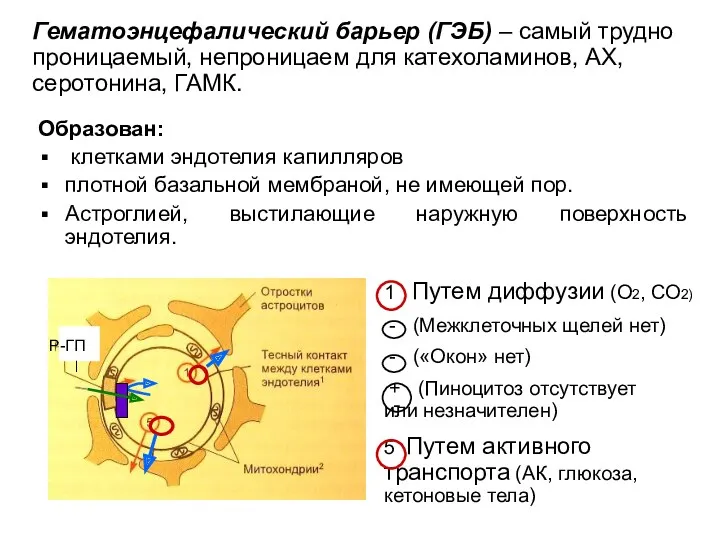
Образован:
клетками эндотелия капилляров
плотной базальной мембраной, не имеющей пор.
Астроглией,
Образован:
клетками эндотелия капилляров
плотной базальной мембраной, не имеющей пор.
Астроглией,

Особенности метаболизма нервной ткани
Для мозга характерна высокая интенсивность энергетического обмена с
Особенности метаболизма нервной ткани
Для мозга характерна высокая интенсивность энергетического обмена с

Основной энергетический субстрат для нервной ткани - глюкоза
За 1 минуту 100
Основной энергетический субстрат для нервной ткани - глюкоза
За 1 минуту 100

Обмен белков и аминокислот нервной ткани
Нервная ткань характеризуется высоким обменом аминокислот
Обмен белков и аминокислот нервной ткани
Нервная ткань характеризуется высоким обменом аминокислот
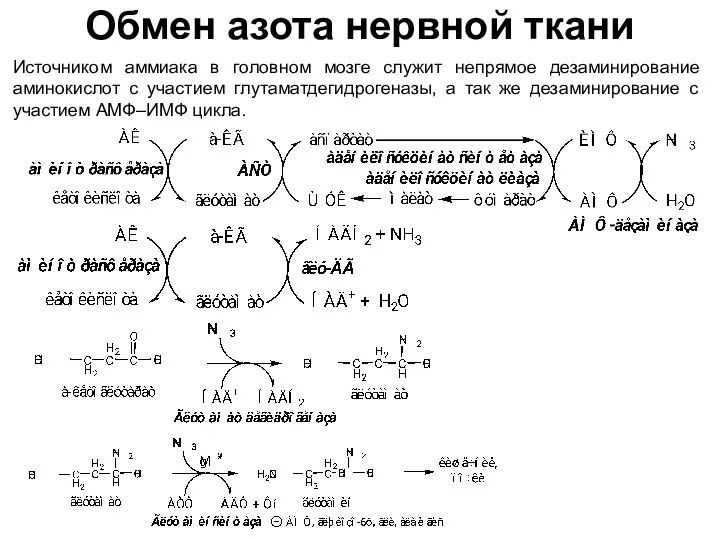
Обмен азота нервной ткани
Источником аммиака в головном мозге служит непрямое
Обмен азота нервной ткани
Источником аммиака в головном мозге служит непрямое

Липидный обмен нервной ткани
Особенность: липиды не используются в качестве энергетического материала,
Липидный обмен нервной ткани
Особенность: липиды не используются в качестве энергетического материала,

Механизмы передачи
нервного импульса
Механизмы передачи
нервного импульса

Механизмы передачи нервного импульса
по нервному волокну
1. Потенциал покоя (-70 мВ)
Na-канал
К-канал
2.
Механизмы передачи нервного импульса
по нервному волокну
1. Потенциал покоя (-70 мВ)
Na-канал
К-канал
2.
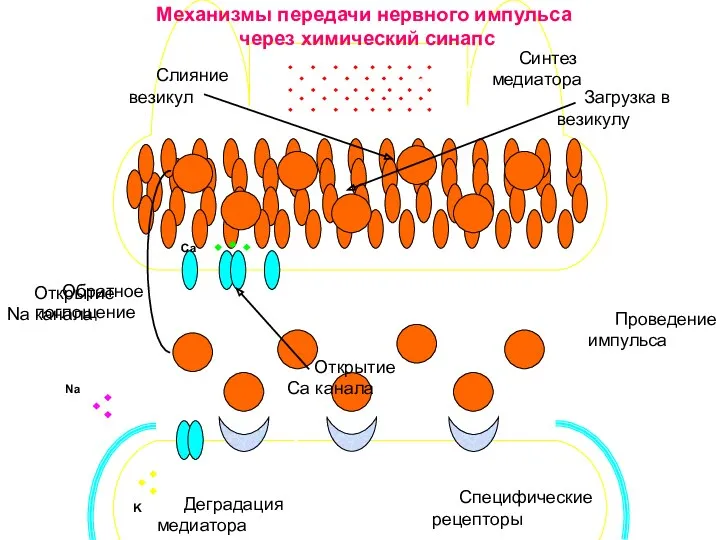
Са
Na
K
Синтез медиатора
Загрузка в везикулу
Слияние везикул
Специфические рецепторы
Открытие Na канала
Проведение импульса
Открытие Сa канала
Деградация
Са
Na
K
Синтез медиатора
Загрузка в везикулу
Слияние везикул
Специфические рецепторы
Открытие Na канала
Проведение импульса
Открытие Сa канала
Деградация

Аминокислотные медиаторы
ингибиторные нейтральные
(ГАМК, глицин, β-аланин и таурин).
возбуждающие кислые
(глутамат и
Аминокислотные медиаторы
ингибиторные нейтральные
(ГАМК, глицин, β-аланин и таурин).
возбуждающие кислые
(глутамат и
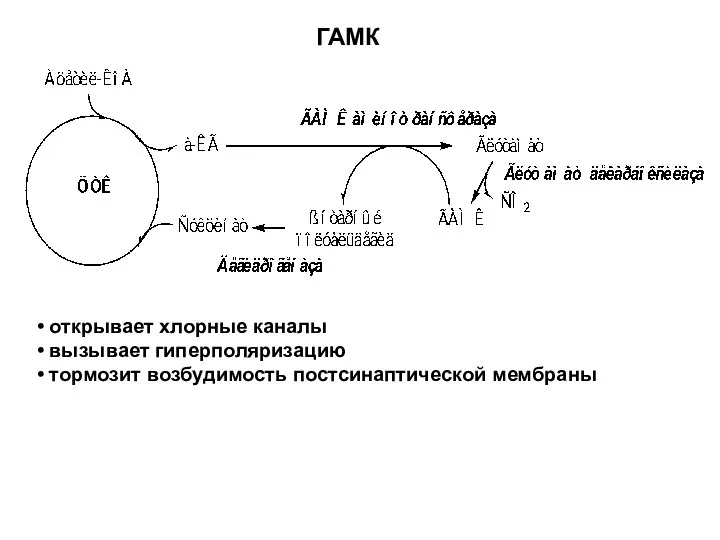
открывает хлорные каналы
вызывает гиперполяризацию
тормозит возбудимость постсинаптической мембраны
открывает хлорные каналы
вызывает гиперполяризацию
тормозит возбудимость постсинаптической мембраны
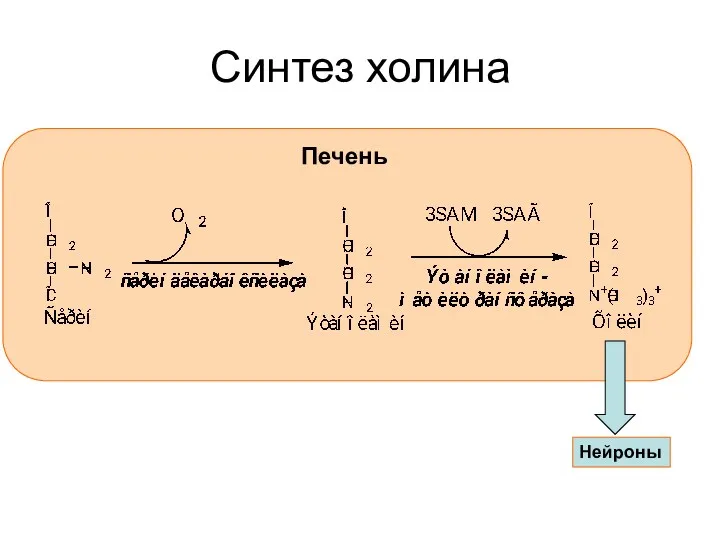
Синтез холина
Печень
Нейроны
Синтез холина
Печень
Нейроны
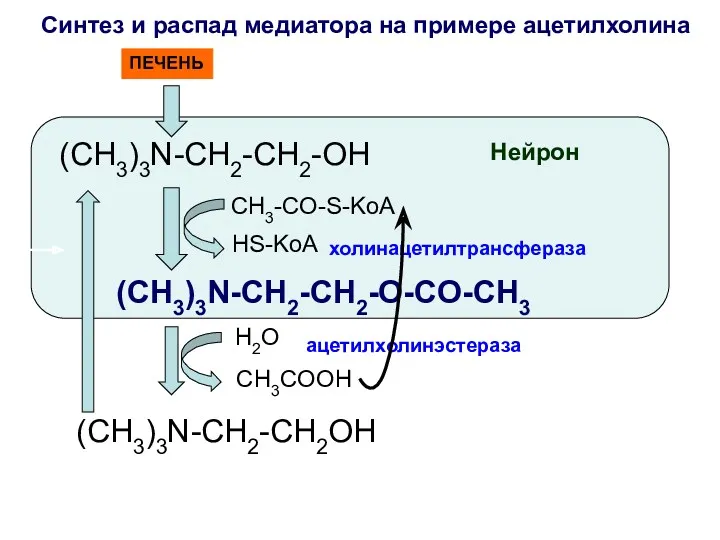
Синтез и распад медиатора на примере ацетилхолина
(CH3)3N-CH2-CH2-OH
холинацетилтрансфераза
(CH3)3N-CH2-CH2-O-CO-CH3
H2O
ацетилхолинэстераза
(CH3)3N-CH2-CH2OH
Нейрон
ПЕЧЕНЬ
CH3-CO-S-KoA
HS-KoA
CH3COOH
Синтез и распад медиатора на примере ацетилхолина
(CH3)3N-CH2-CH2-OH
холинацетилтрансфераза
(CH3)3N-CH2-CH2-O-CO-CH3
H2O
ацетилхолинэстераза
(CH3)3N-CH2-CH2OH
Нейрон
ПЕЧЕНЬ
CH3-CO-S-KoA
HS-KoA
CH3COOH

Рецепторы
Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри
Рецепторы
Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри

Биохимические синдромы в психиатрии
Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина)
Нарушения окислительно-восстановительных процессов (гипоксия,
Биохимические синдромы в психиатрии
Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина)
Нарушения окислительно-восстановительных процессов (гипоксия,

Патобиохимия нервной системы
1) Миастения - уменьшение числа холинорецепторов
3) болезнь Паркинсона -
Патобиохимия нервной системы
1) Миастения - уменьшение числа холинорецепторов
3) болезнь Паркинсона -
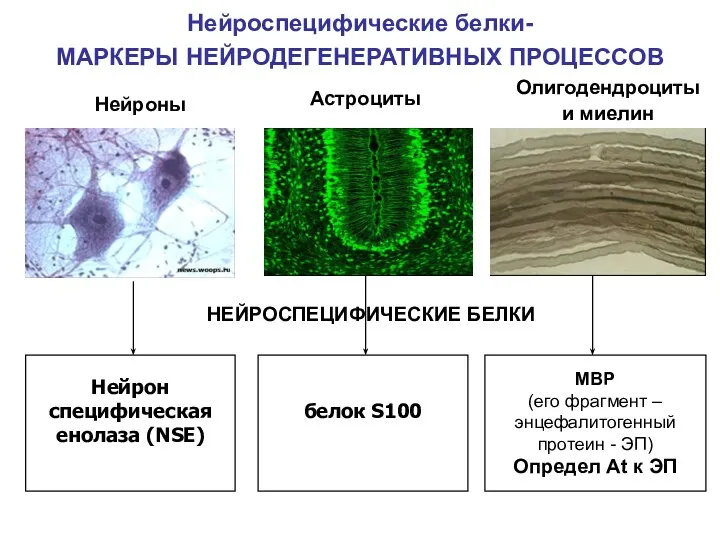
Нейроспецифические белки-
МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ
белок S100
Нейрон
специфическая
енолаза (NSE)
Нейроны
Астроциты
Олигодендроциты
и миелин
MBP
(его
Нейроспецифические белки-
МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ
белок S100
Нейрон
специфическая
енолаза (NSE)
Нейроны
Астроциты
Олигодендроциты
и миелин
MBP
(его

Нейронспецифическая енолаза (NSE)
Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент,
Нейронспецифическая енолаза (NSE)
Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент,

Белок S100
S-100 - кальций-связывающий протеин.
Локализация мембраны, цитоплазма.
участвует в фосфорилировании
Белок S100
S-100 - кальций-связывающий протеин.
Локализация мембраны, цитоплазма.
участвует в фосфорилировании

антитела к S100β являются специфичными маркерами повреждения астроцитарной глии.
Ген S100
Ген S100

Основной белок миелина (MBP)
MBP - щелочной белок, с высоким содержанием (25%)
Основной белок миелина (MBP)
MBP - щелочной белок, с высоким содержанием (25%)

NSE, S100 и MBP – маркеры повреждения мозговой ткани
Нейроспецифические белки в
NSE, S100 и MBP – маркеры повреждения мозговой ткани
Нейроспецифические белки в

NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также
NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также

концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени,
концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени,
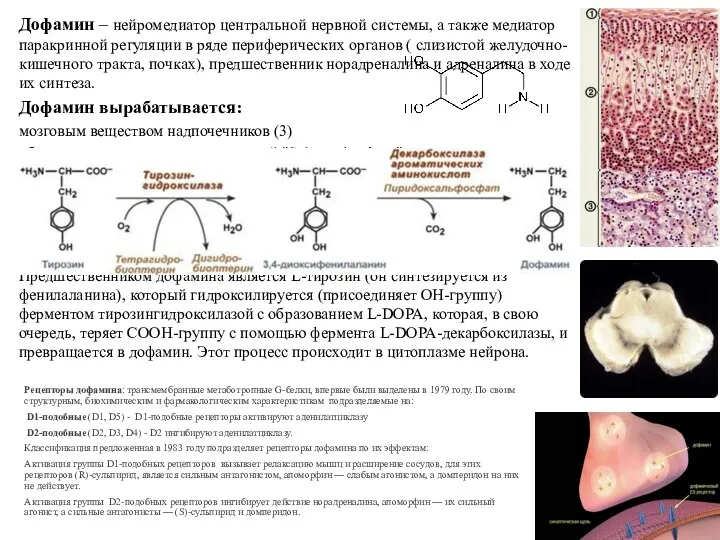
Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции
Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции
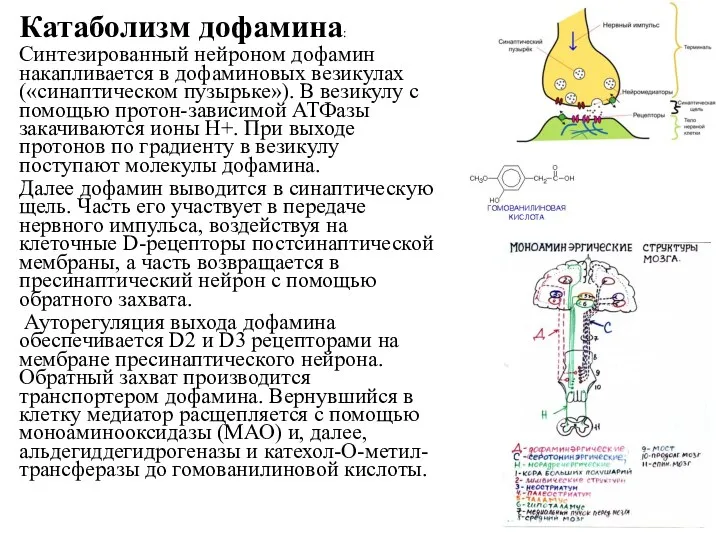
Катаболизм дофамина:
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»).
Катаболизм дофамина:
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»).

ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
Различают семь отдельных подсистем (первые три являются основными):
ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
Различают семь отдельных подсистем (первые три являются основными):
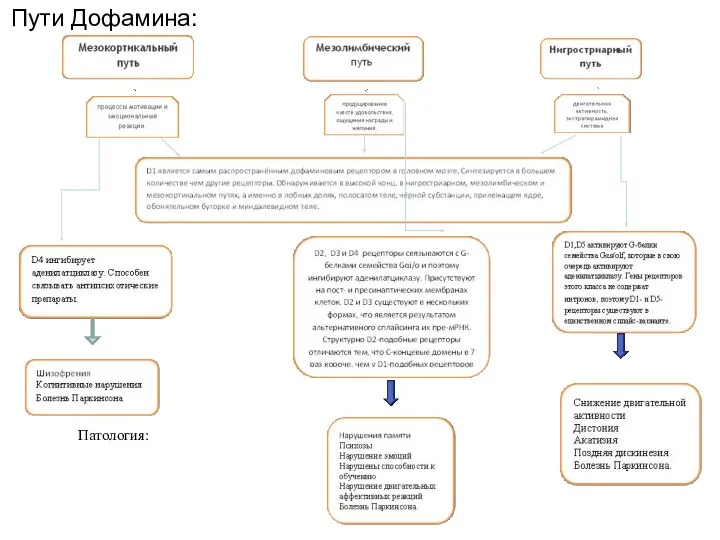
Пути Дофамина:
Патология:
Патология
Патология
Пути Дофамина:
Патология:
Патология
Патология

Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга.
Снижена активность тирозингидроксилазы,
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы,
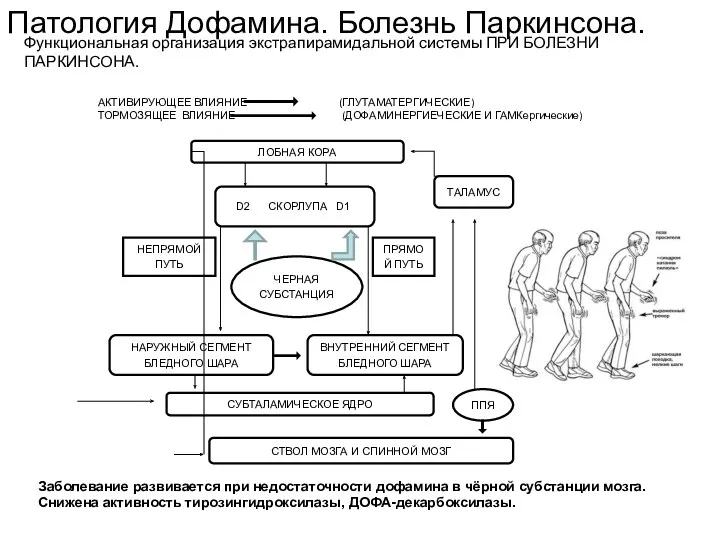
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга.
Снижена активность тирозингидроксилазы,
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы,

План лекции
Функции мышц
Классификация мышечных волокон
Химический состав мышц
Особенности обмена веществ и энергии
План лекции
Функции мышц
Классификация мышечных волокон
Химический состав мышц
Особенности обмена веществ и энергии
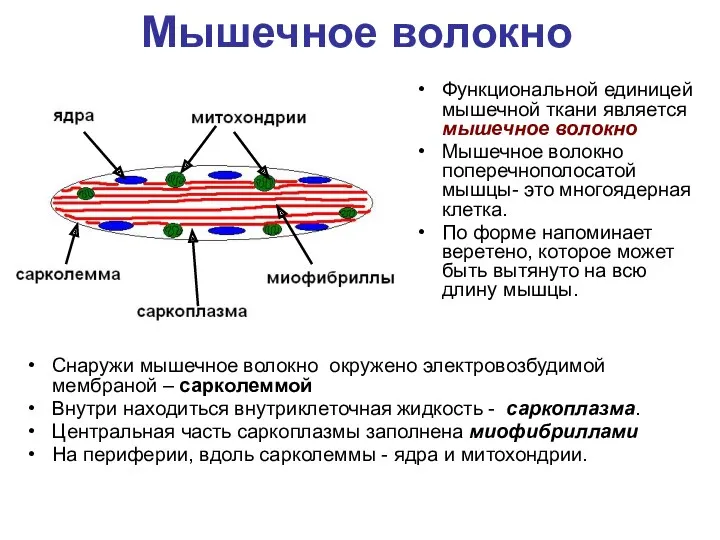
Мышечное волокно
Функциональной единицей мышечной ткани является мышечное волокно
Мышечное волокно поперечнополосатой
Мышечное волокно
Функциональной единицей мышечной ткани является мышечное волокно
Мышечное волокно поперечнополосатой
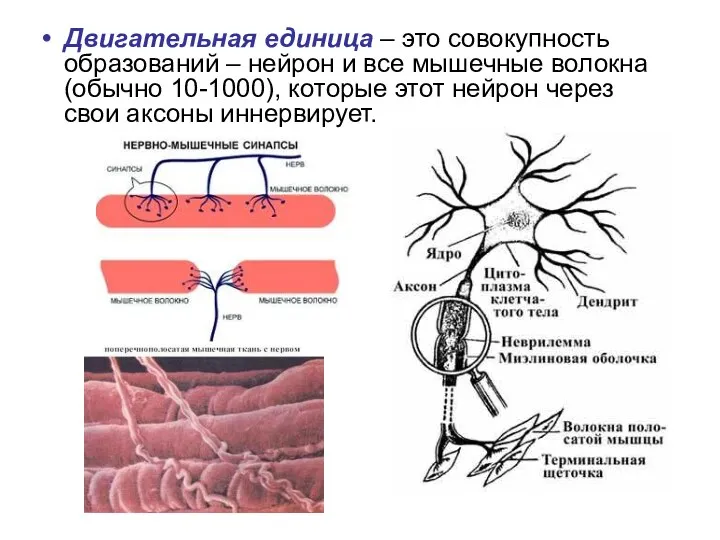
Двигательная единица – это совокупность образований – нейрон и все мышечные
Двигательная единица – это совокупность образований – нейрон и все мышечные

ХИМИЧЕСКИЙ СОСТАВ МЫШЕЧНОЙ ТКАНИ
Вода............................................72—80
Сухой остаток …........................20—28
В том числе:
Белки...........................................16,5—20,9
Гликоген......................................0,3—3,0
Фосфатиды.................................0,4—1,0
Холестерин.................................0,06—0,2
Креатин + креатинфосфат........0,2—0,55
Креатинин....................................0,003—0,005
АТФ...............................................0,25—0,4
Карнозин......................................0,2—0,3
Карнитин......................................0,02—0,05
Анзерин........................................0,09—0,15
Свободные
ХИМИЧЕСКИЙ СОСТАВ МЫШЕЧНОЙ ТКАНИ
Вода............................................72—80
Сухой остаток …........................20—28
В том числе:
Белки...........................................16,5—20,9
Гликоген......................................0,3—3,0
Фосфатиды.................................0,4—1,0
Холестерин.................................0,06—0,2
Креатин + креатинфосфат........0,2—0,55
Креатинин....................................0,003—0,005
АТФ...............................................0,25—0,4
Карнозин......................................0,2—0,3
Карнитин......................................0,02—0,05
Анзерин........................................0,09—0,15
Свободные

1. Сократительные (миофибриллярные) белки
миозин 55%
актин 25%
тропомиозин (во всех
1. Сократительные (миофибриллярные) белки
миозин 55%
актин 25%
тропомиозин (во всех

Углеводы мышечной ткани
Гликоген: 0,3-3,0%
ГАГ
моносахариды глюкоза, фруктоза
Липиды мышечной ткани
Фосфолипиды (в миокарде
Углеводы мышечной ткани
Гликоген: 0,3-3,0%
ГАГ
моносахариды глюкоза, фруктоза
Липиды мышечной ткани
Фосфолипиды (в миокарде

ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ
В МЫШЕЧНОЙ ТКАНИ
Обмен белков и аминокислот
Мышцы характеризуются
ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ
В МЫШЕЧНОЙ ТКАНИ
Обмен белков и аминокислот
Мышцы характеризуются
Мышцы также синтезируют и выделяют много аланина и глутамина. В реакций
Мышцы также синтезируют и выделяют много аланина и глутамина. В реакций
Аминокислоты
Белки
При интенсивной работе для нейтрализации лактата мышцы выделяют аммиак. В мышцах
Аминокислоты
Белки
При интенсивной работе для нейтрализации лактата мышцы выделяют аммиак. В мышцах

Липидный обмен
преобладает катаболизм липидов.
Жирные кислоты, кетоновые тела в аэробных условиях
Липидный обмен
преобладает катаболизм липидов.
Жирные кислоты, кетоновые тела в аэробных условиях

Углеводный обмен
преобладает катаболизм углеводов.
глюкоза окисляется в аэробных или анаэробных условиях
Углеводный обмен
преобладает катаболизм углеводов.
глюкоза окисляется в аэробных или анаэробных условиях

Энергетический обмен
Энергетический обмен в состоянии покоя очень низкий, при интенсивной физической
Энергетический обмен
Энергетический обмен в состоянии покоя очень низкий, при интенсивной физической
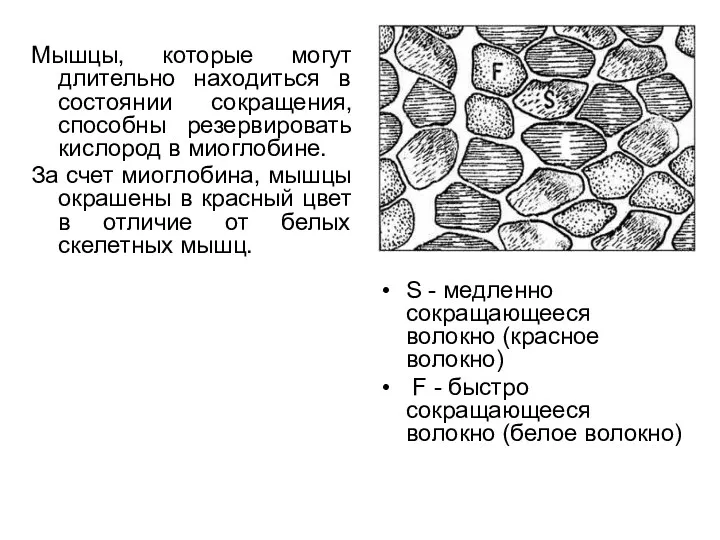
Мышцы, которые могут длительно находиться в состоянии сокращения, способны резервировать кислород
Мышцы, которые могут длительно находиться в состоянии сокращения, способны резервировать кислород

Основной потребитель АТФ - процесс мышечного сокращения. Запасы АТФ быстро истощаются.
Источники
Основной потребитель АТФ - процесс мышечного сокращения. Запасы АТФ быстро истощаются.
Источники
(Окислительное фосфорилирование)
АТФ
АДФ-АТФ-транслоказа(ААТ)
ВНУТРЕННЯЯ МЕМБРАНА МИТОХОНДРИЙ
КРЕАТИН
КРЕАТИН ФОСФАТ
АТФ
АДФ
САРКОЛЕММА
МИОФИБРИЛЛА
КРЕАТИН ФОСФАТ
АДФ
АТФ
КРЕАТИН
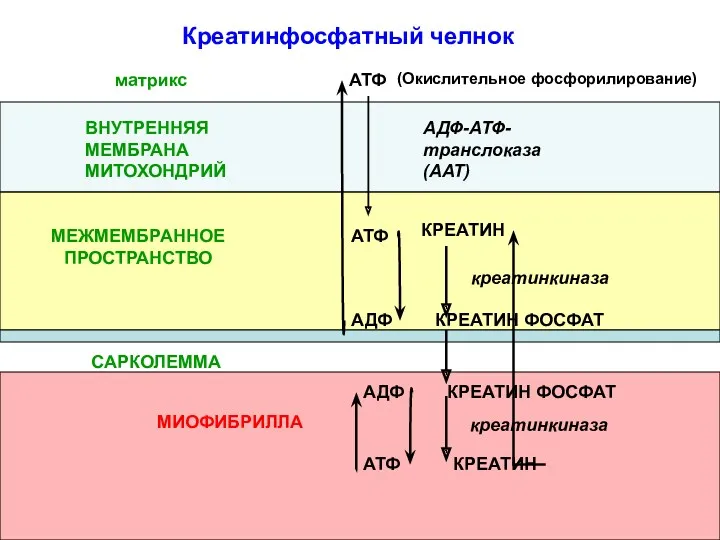
Креатинфосфатный челнок
креатинкиназа
креатинкиназа
матрикс
МЕЖМЕМБРАННОЕ ПРОСТРАНСТВО
(Окислительное фосфорилирование)
АТФ
АДФ-АТФ-транслоказа(ААТ)
ВНУТРЕННЯЯ МЕМБРАНА МИТОХОНДРИЙ
КРЕАТИН
КРЕАТИН ФОСФАТ
АТФ
АДФ
САРКОЛЕММА
МИОФИБРИЛЛА
КРЕАТИН ФОСФАТ
АДФ
АТФ
КРЕАТИН
Креатинфосфатный челнок
креатинкиназа
креатинкиназа
матрикс
МЕЖМЕМБРАННОЕ ПРОСТРАНСТВО

Креатинфосфатный челнок предотвращает быстрое истощение запасов АТФ в мышце за счет:
запаса
Креатинфосфатный челнок предотвращает быстрое истощение запасов АТФ в мышце за счет:
запаса

Характеристика быстрых и медленных скелетных мышц
Характеристика быстрых и медленных скелетных мышц

Миофибрилла
Миофибрилла
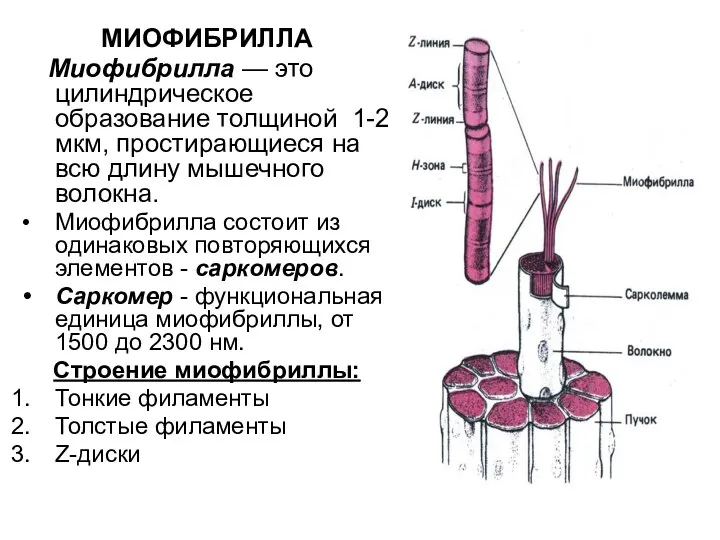
МИОФИБРИЛЛА
Миофибрилла — это цилиндрическое образование толщиной 1-2 мкм, простирающиеся на
МИОФИБРИЛЛА
Миофибрилла — это цилиндрическое образование толщиной 1-2 мкм, простирающиеся на
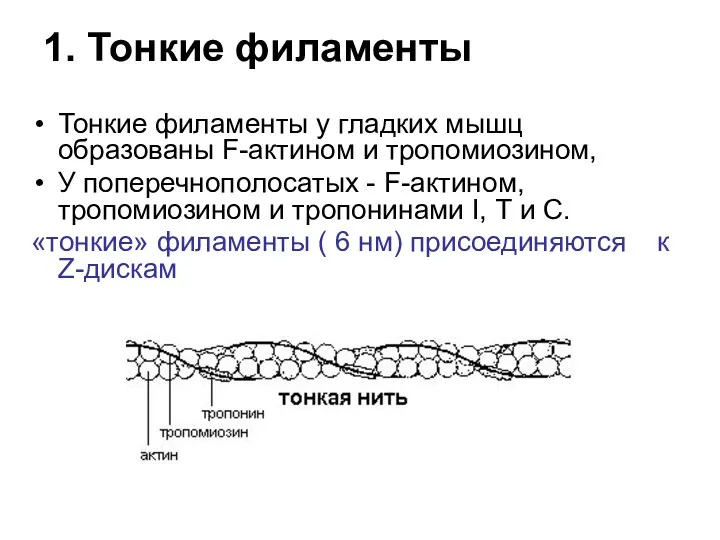
1. Тонкие филаменты
Тонкие филаменты у гладких мышц образованы F-актином и тропомиозином,
1. Тонкие филаменты
Тонкие филаменты у гладких мышц образованы F-актином и тропомиозином,

Актин
G-актин - мономерный (глобулярный) белок с массой 43кДа.
F-актин. При физиологической
Актин
G-актин - мономерный (глобулярный) белок с массой 43кДа.
F-актин. При физиологической

Тропомиозин - есть во всех мышцах: белок, состоящий из а и
Тропомиозин - есть во всех мышцах: белок, состоящий из а и
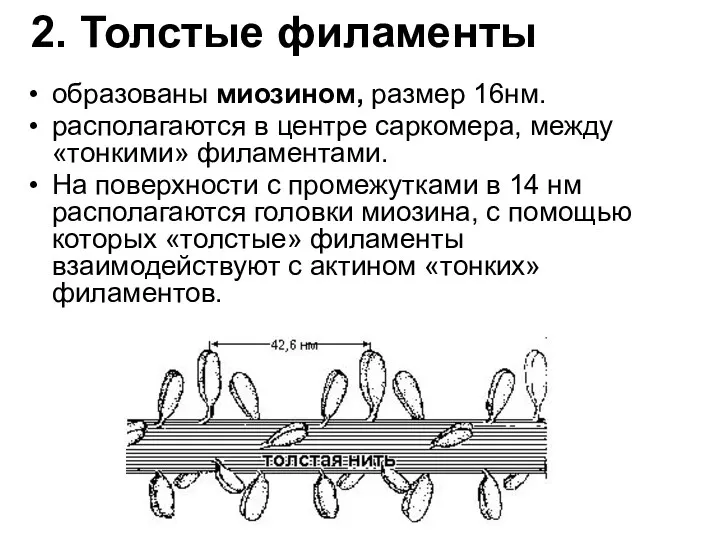
2. Толстые филаменты
образованы миозином, размер 16нм.
располагаются в центре саркомера, между «тонкими»
2. Толстые филаменты
образованы миозином, размер 16нм.
располагаются в центре саркомера, между «тонкими»
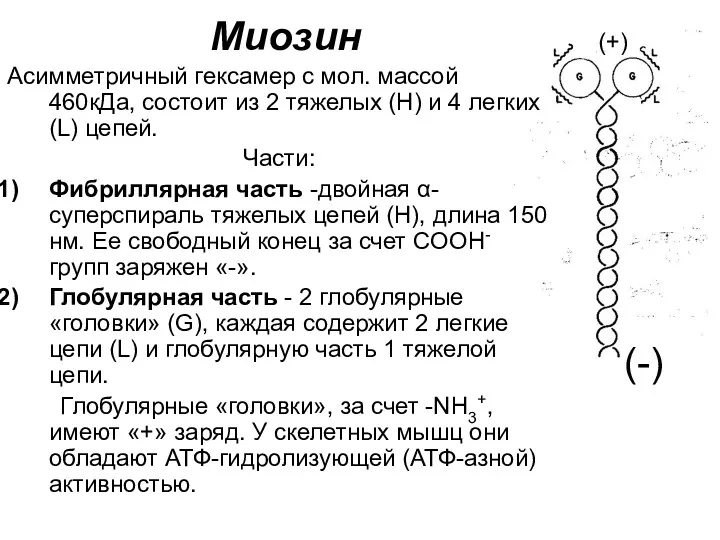
Миозин
Асимметричный гексамер с мол. массой 460кДа, состоит из 2 тяжелых
Миозин
Асимметричный гексамер с мол. массой 460кДа, состоит из 2 тяжелых
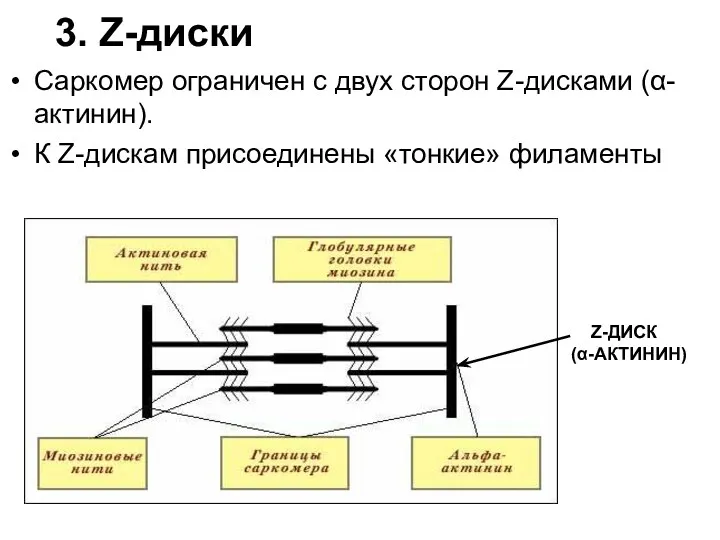
3. Z-диски
Саркомер ограничен с двух сторон Z-дисками (α-актинин).
К Z-дискам присоединены
3. Z-диски
Саркомер ограничен с двух сторон Z-дисками (α-актинин).
К Z-дискам присоединены
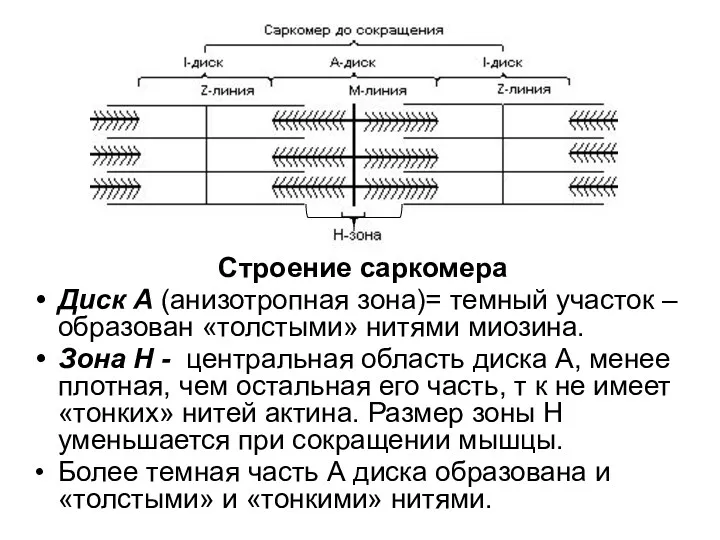
Строение саркомера
Диск А (анизотропная зона)= темный участок –образован «толстыми» нитями миозина.
Строение саркомера
Диск А (анизотропная зона)= темный участок –образован «толстыми» нитями миозина.

Полоса М - пересекает центральную область диска А, образована толстыми нитями,
Полоса М - пересекает центральную область диска А, образована толстыми нитями,
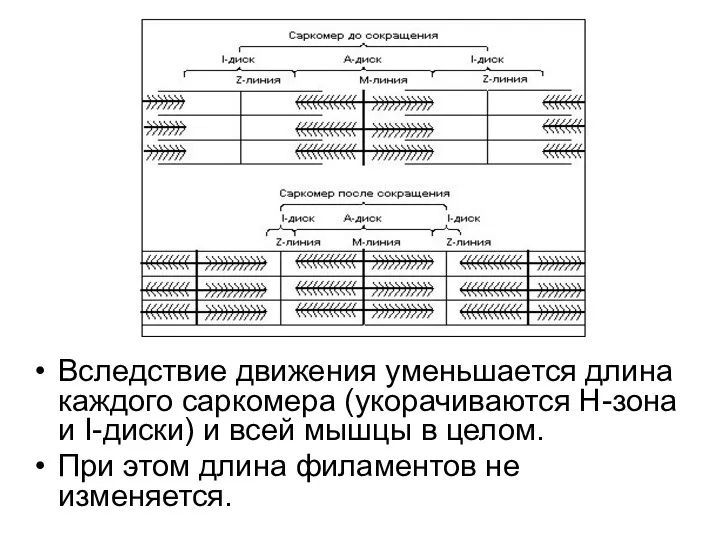
Вследствие движения уменьшается длина каждого саркомера (укорачиваются Н-зона и I-диски) и
Вследствие движения уменьшается длина каждого саркомера (укорачиваются Н-зона и I-диски) и

В скелетной мышечной ткани саркомеры миофибрилл располагаются параллельно.
За счет этого
В скелетной мышечной ткани саркомеры миофибрилл располагаются параллельно.
За счет этого
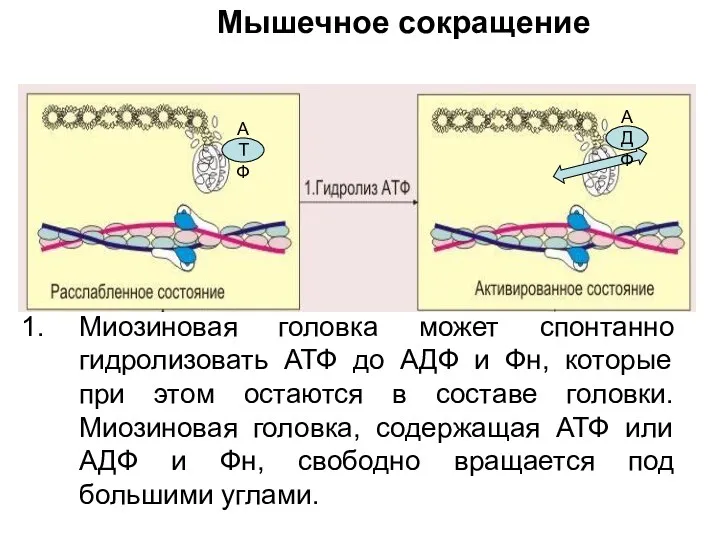
Миозиновая головка может спонтанно гидролизовать АТФ до АДФ и Фн, которые
Миозиновая головка может спонтанно гидролизовать АТФ до АДФ и Фн, которые
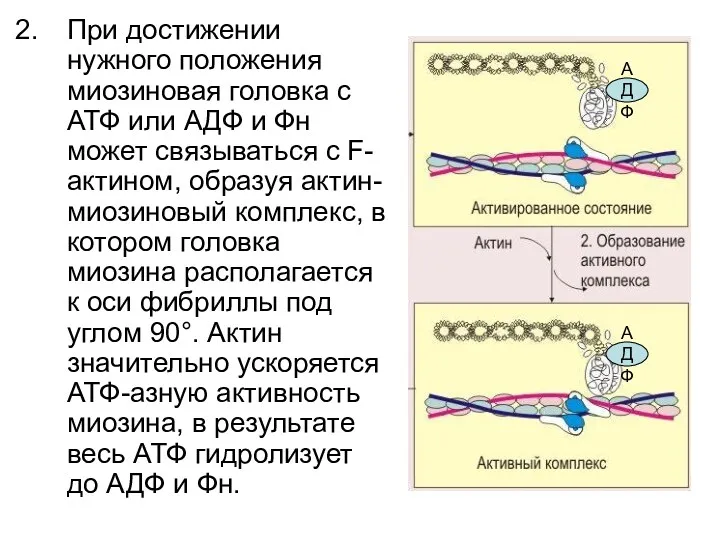
При достижении нужного положения миозиновая головка с АТФ или АДФ и
При достижении нужного положения миозиновая головка с АТФ или АДФ и
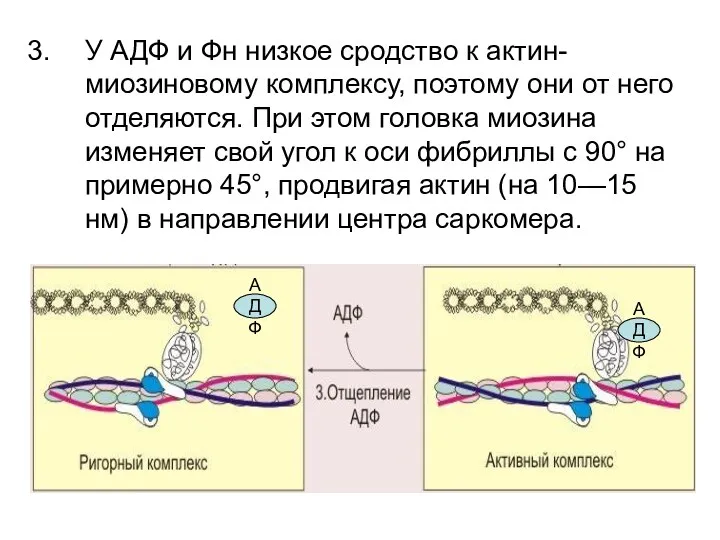
У АДФ и Фн низкое сродство к актин-миозиновому комплексу, поэтому они
У АДФ и Фн низкое сродство к актин-миозиновому комплексу, поэтому они
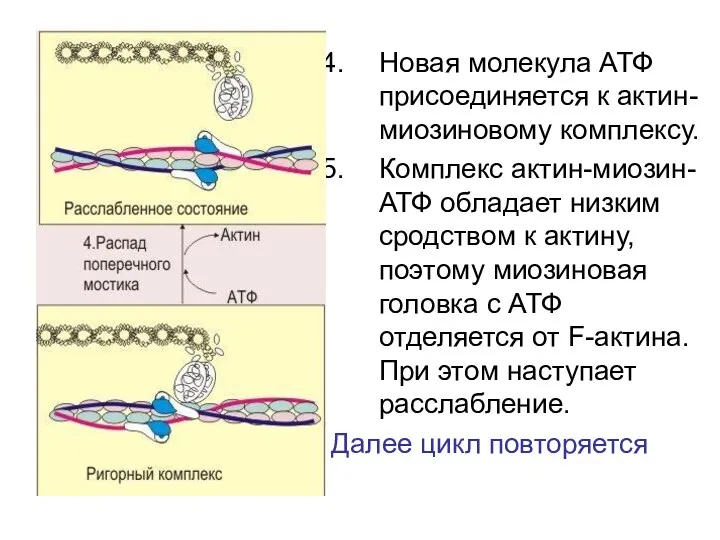
Новая молекула АТФ присоединяется к актин-миозиновому комплексу.
Комплекс актин-миозин-АТФ обладает низким
Новая молекула АТФ присоединяется к актин-миозиновому комплексу.
Комплекс актин-миозин-АТФ обладает низким
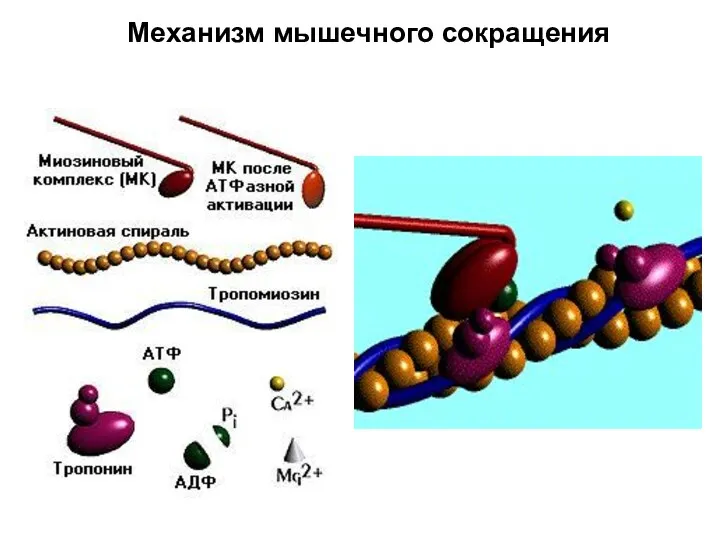
Механизм мышечного сокращения
Механизм мышечного сокращения

Регуляция сокращения и расслабления мышц
Любое мышечное сокращение опосредуется Са2+.
Кальциевые насосы
Регуляция сокращения и расслабления мышц
Любое мышечное сокращение опосредуется Са2+.
Кальциевые насосы
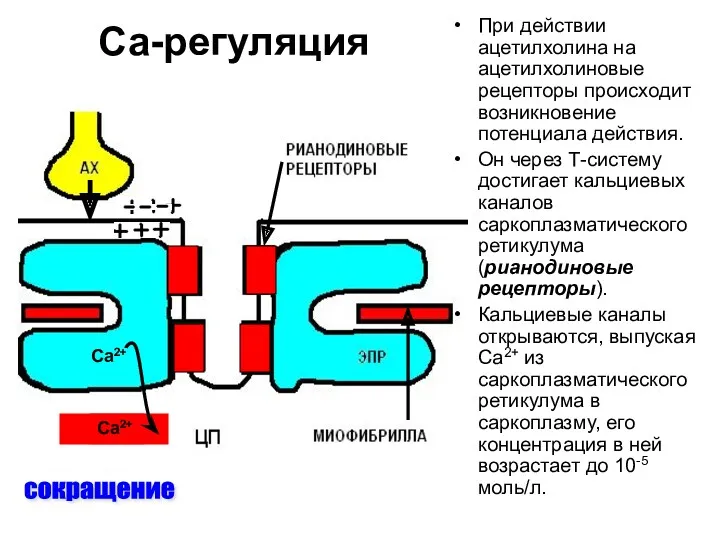
Са-регуляция
сокращение
При действии ацетилхолина на ацетилхолиновые рецепторы происходит возникновение потенциала действия.
Са-регуляция
сокращение
При действии ацетилхолина на ацетилхолиновые рецепторы происходит возникновение потенциала действия.
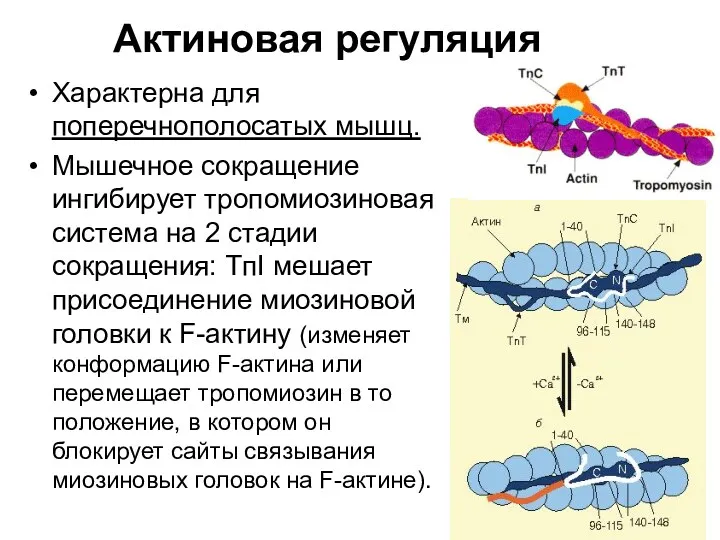
Актиновая регуляция
Характерна для поперечнополосатых мышц.
Мышечное сокращение ингибирует тропомиозиновая система на
Актиновая регуляция
Характерна для поперечнополосатых мышц.
Мышечное сокращение ингибирует тропомиозиновая система на
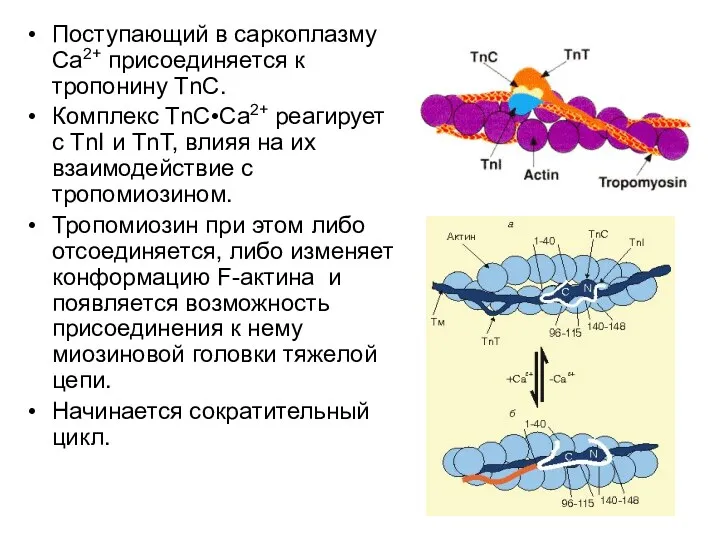
Поступающий в саркоплазму Са2+ присоединяется к тропонину ТnС.
Комплекс ТnС•Са2+ реагирует
Поступающий в саркоплазму Са2+ присоединяется к тропонину ТnС.
Комплекс ТnС•Са2+ реагирует

Расслабление происходит, когда:
1) При перекачке Са2+ в ЭПР его содержание
Расслабление происходит, когда:
1) При перекачке Са2+ в ЭПР его содержание
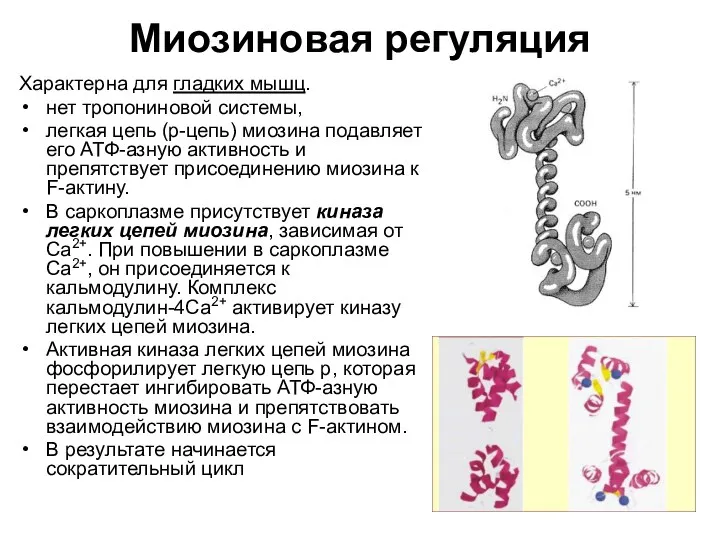
Миозиновая регуляция
Характерна для гладких мышц.
нет тропониновой системы,
легкая цепь (р-цепь) миозина
Миозиновая регуляция
Характерна для гладких мышц.
нет тропониновой системы,
легкая цепь (р-цепь) миозина

Расслабление гладких мышц происходит, когда:
1) содержание ионов Са2+ в саркоплазме
Расслабление гладких мышц происходит, когда:
1) содержание ионов Са2+ в саркоплазме

Биохимические показатели крови и мочи, отражающие функциональное состояние мышечной ткани
Аминотрансферазы -диагностика
Биохимические показатели крови и мочи, отражающие функциональное состояние мышечной ткани
Аминотрансферазы -диагностика

Тропонин Т -маркер инфаркта миокарда в острой и подострой фазе.
Миоглобин
Тропонин Т -маркер инфаркта миокарда в острой и подострой фазе.
Миоглобин

Миопатии
(греч. mys, myos мышца + pathos страдание, болезнь) - нервно-мышечные заболевания,
Миопатии
(греч. mys, myos мышца + pathos страдание, болезнь) - нервно-мышечные заболевания,


БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ В МЫШЦАХ
ПРИ ПАТОЛОГИИ:
снижение содержания миофибриллярных белков
возрастание концентрации
БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ В МЫШЦАХ
ПРИ ПАТОЛОГИИ:
снижение содержания миофибриллярных белков
возрастание концентрации
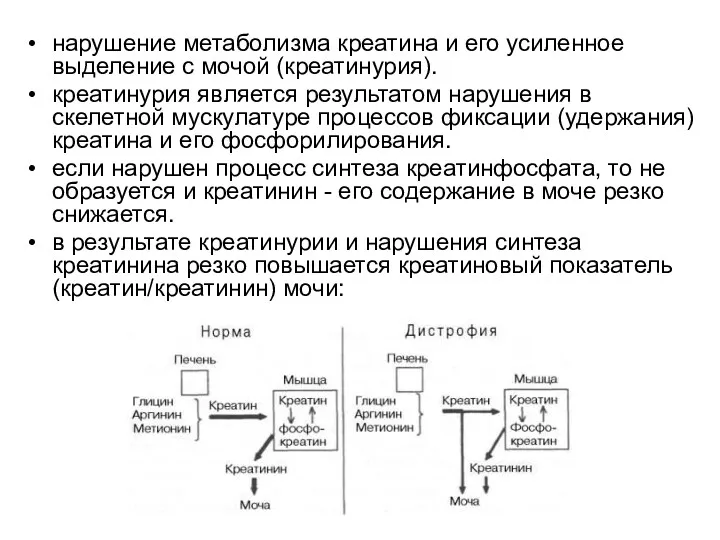
нарушение метаболизма креатина и его усиленное выделение с мочой (креатинурия).
креатинурия
нарушение метаболизма креатина и его усиленное выделение с мочой (креатинурия).
креатинурия

Изменение активности ферментов в мышцах:
уменьшается активность ферментов, локализованных в саркоплазме
Изменение активности ферментов в мышцах:
уменьшается активность ферментов, локализованных в саркоплазме
Ишемическая болезнь сердца
ИБС - патологическое состояние, характеризующееся абсолютным или относительным
Ишемическая болезнь сердца
ИБС - патологическое состояние, характеризующееся абсолютным или относительным
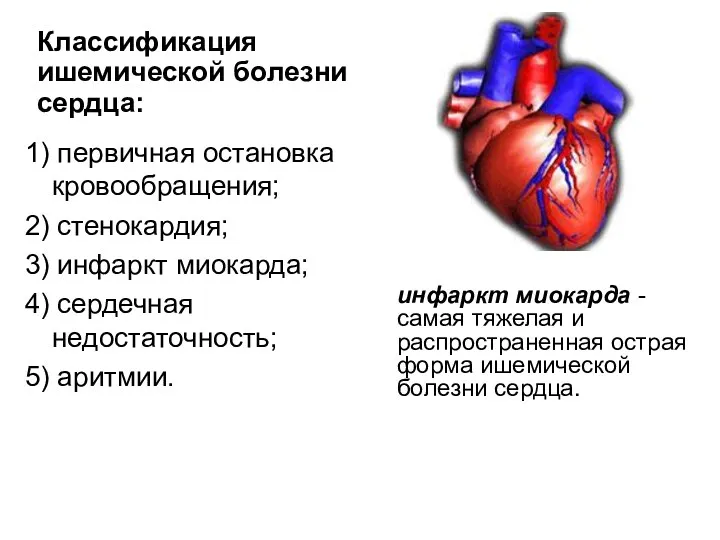
1) первичная остановка кровообращения;
2) стенокардия;
3) инфаркт миокарда;
4) сердечная недостаточность;
1) первичная остановка кровообращения;
2) стенокардия;
3) инфаркт миокарда;
4) сердечная недостаточность;

Причины инфаркта миокарда
Атеросклероз коронарных артерий
Эмболы, тромбы коронарной артерии
Спазм коронарных артерий и
Причины инфаркта миокарда
Атеросклероз коронарных артерий
Эмболы, тромбы коронарной артерии
Спазм коронарных артерий и
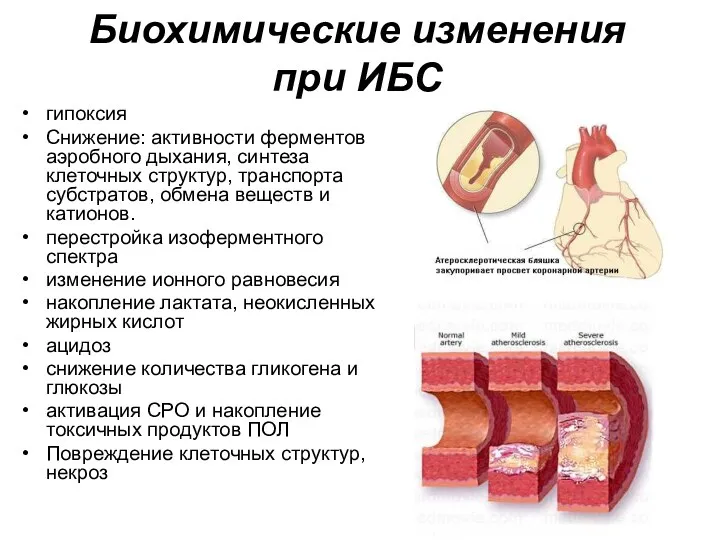
Биохимические изменения
при ИБС
гипоксия
Снижение: активности ферментов аэробного дыхания, синтеза клеточных структур,
Биохимические изменения
при ИБС
гипоксия
Снижение: активности ферментов аэробного дыхания, синтеза клеточных структур,
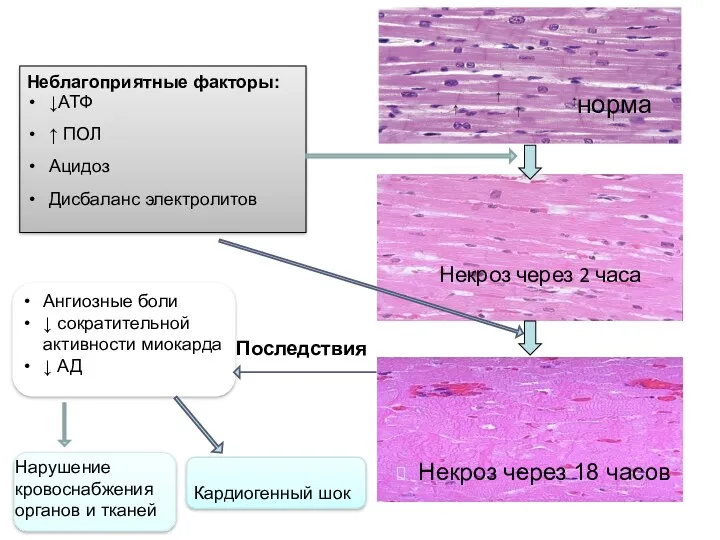
норма
Некроз через 2 часа
Некроз через 18 часов
Неблагоприятные факторы:
↓АТФ
↑ ПОЛ
Ацидоз
Дисбаланс электролитов
Ангиозные боли
↓
норма
Некроз через 2 часа
Некроз через 18 часов
Неблагоприятные факторы:
↓АТФ
↑ ПОЛ
Ацидоз
Дисбаланс электролитов
Ангиозные боли
↓
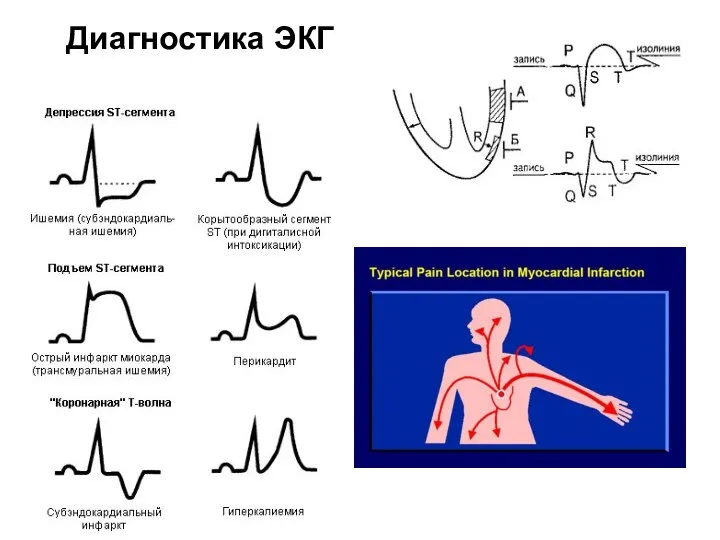
Диагностика ЭКГ
Диагностика ЭКГ

При гипоксии миокард поглощает ТГ из липопротеинов, которые не используются, а
При гипоксии миокард поглощает ТГ из липопротеинов, которые не используются, а

Терапия
Терапия

План лекции
Биохимия межклеточного матрикса
гетерополисахариды
структурные и адгезивные белки
Минерализованная соединительная ткань. Ремоделирование
План лекции
Биохимия межклеточного матрикса
гетерополисахариды
структурные и адгезивные белки
Минерализованная соединительная ткань. Ремоделирование

Межклеточный матрикс
Гетерополисахариды и протеогликаны
Гликозамингликаны
гиалуроновая кислота
хондроитинсульфат
дерматансульфат
гепарансульфат
гепарин
кератансульфат
Протеогликаны (гликозамингликаны + белки)
Белки
Структурные
коллаген
эластин
Межклеточный матрикс
Гетерополисахариды и протеогликаны
Гликозамингликаны
гиалуроновая кислота
хондроитинсульфат
дерматансульфат
гепарансульфат
гепарин
кератансульфат
Протеогликаны (гликозамингликаны + белки)
Белки
Структурные
коллаген
эластин

Гликозамингликаны, или мукополисахариды (ГАГ)
Линейные отрицательно заряженные гетерополисахариды, состоящие из повторяющихся дисахаридных
Гликозамингликаны, или мукополисахариды (ГАГ)
Линейные отрицательно заряженные гетерополисахариды, состоящие из повторяющихся дисахаридных

Дисахаридная единица гиалуроновой кислоты (всего несколько тыс. дисахаридных единиц)
Дисахаридная единица дерматансульфата
Структуру
Дисахаридная единица гиалуроновой кислоты (всего несколько тыс. дисахаридных единиц)
Дисахаридная единица дерматансульфата
Структуру

Особенности строения отдельных ГАГ
Кератансульфаты вместо гексуроновой кислоты содержат галактозу и кроме
Особенности строения отдельных ГАГ
Кератансульфаты вместо гексуроновой кислоты содержат галактозу и кроме

Функции гликозамингликанов и протеогликанов
Структурный компонент межклеточного матрикса
Формируют тургор тканей (гиалуроновая кислота)
«Смазка»
Функции гликозамингликанов и протеогликанов
Структурный компонент межклеточного матрикса
Формируют тургор тканей (гиалуроновая кислота)
«Смазка»

Протеогликаны – основное вещество межклеточного матрикса
Состав протеогликанов:
Гликозамингликаны (кроме гиалуроновой кислоты) –
Протеогликаны – основное вещество межклеточного матрикса
Состав протеогликанов:
Гликозамингликаны (кроме гиалуроновой кислоты) –
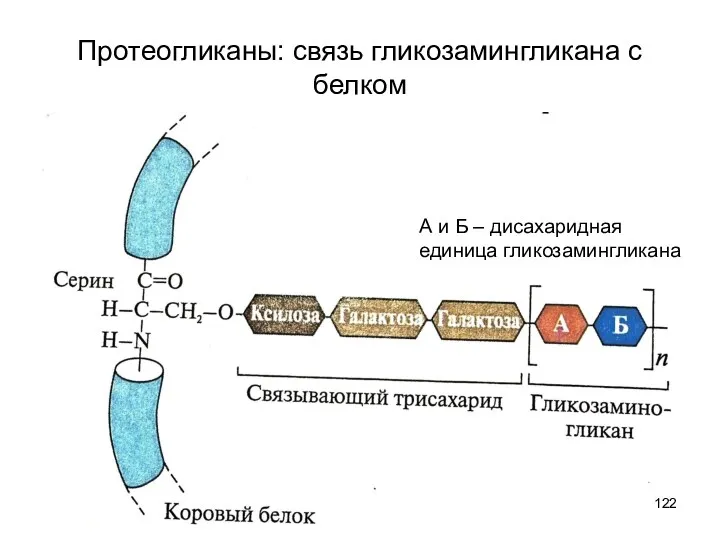
Протеогликаны: связь гликозамингликана с белком
А и Б – дисахаридная единица гликозамингликана
Протеогликаны: связь гликозамингликана с белком
А и Б – дисахаридная единица гликозамингликана

Синтез протеогликанов
1. Синтез белковой части
2. Образование линкерного трисахарида
3. Рост цепи гликозамингликана
Реакции
Синтез протеогликанов
1. Синтез белковой части
2. Образование линкерного трисахарида
3. Рост цепи гликозамингликана
Реакции
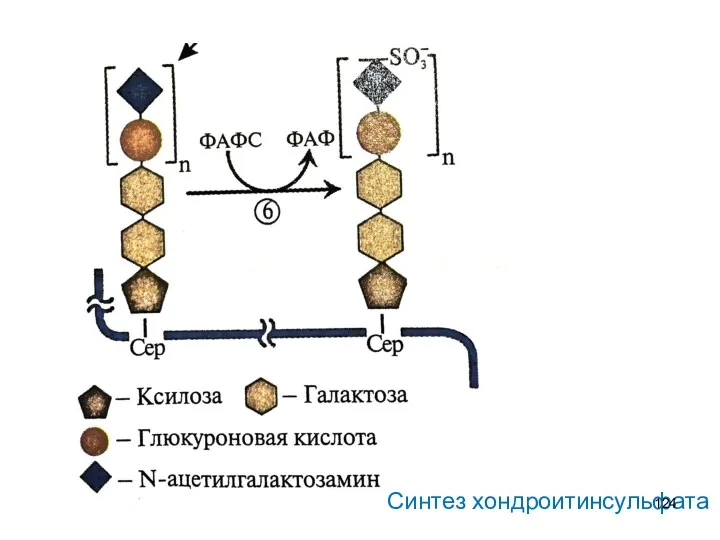
Синтез хондроитинсульфата
Синтез хондроитинсульфата

Катаболизм гликозамингликанов и протеогликанов
Полупериод жизни: 3 – 10 дней (кератансульфат 120
Катаболизм гликозамингликанов и протеогликанов
Полупериод жизни: 3 – 10 дней (кератансульфат 120

Структурные белки соединительной ткани: Коллаген
Коллагены – семейство полиморфных фибриллярных белков (19
Структурные белки соединительной ткани: Коллаген
Коллагены – семейство полиморфных фибриллярных белков (19

Коллаген: особенности строения
Пептидная левозакрученная α-спираль коллагена содержит 600 – 3000 АК
Коллаген: особенности строения
Пептидная левозакрученная α-спираль коллагена содержит 600 – 3000 АК

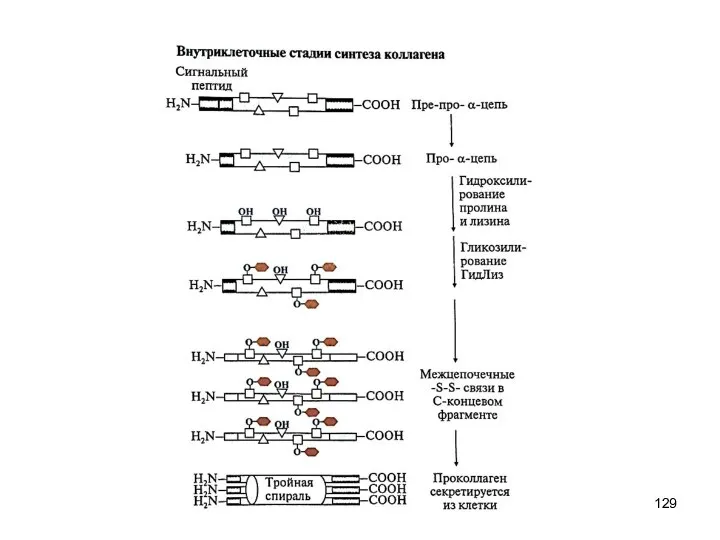
Синтез и созревание коллагена
1. Внутриклеточный этап (фибробласты, хондробласты, остеобласты):
образование пре-про-α-цепи,
Синтез и созревание коллагена
1. Внутриклеточный этап (фибробласты, хондробласты, остеобласты):
образование пре-про-α-цепи,
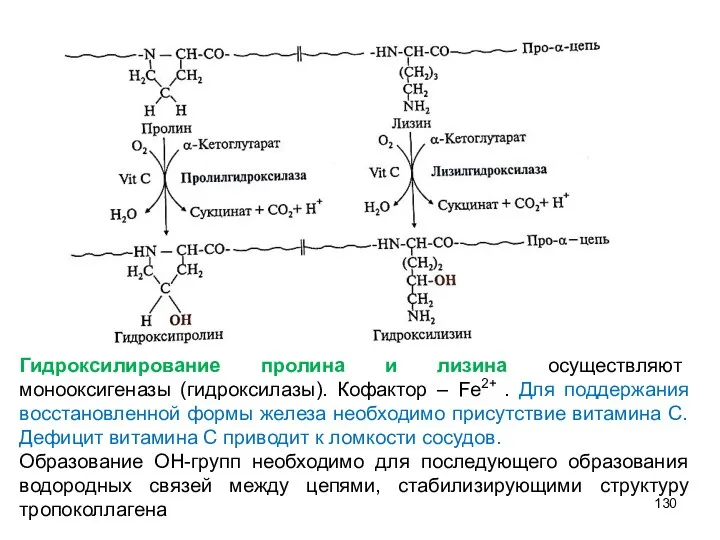
Гидроксилирование пролина и лизина осуществляют монооксигеназы (гидроксилазы). Кофактор – Fe2+ .
Гидроксилирование пролина и лизина осуществляют монооксигеназы (гидроксилазы). Кофактор – Fe2+ .
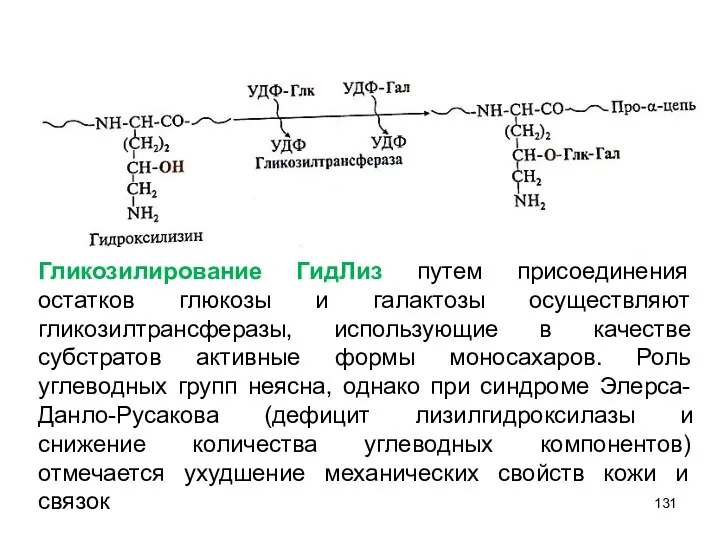
Гликозилирование ГидЛиз путем присоединения остатков глюкозы и галактозы осуществляют гликозилтрансферазы, использующие
Гликозилирование ГидЛиз путем присоединения остатков глюкозы и галактозы осуществляют гликозилтрансферазы, использующие
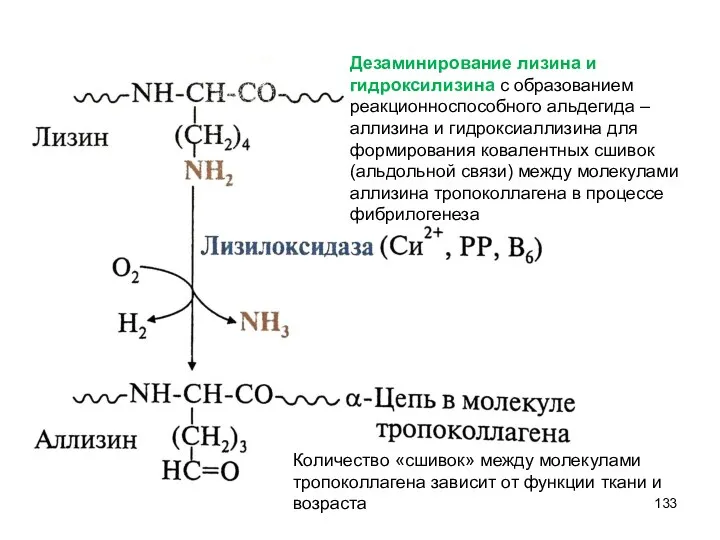
Дезаминирование лизина и гидроксилизина с образованием реакционноспособного альдегида – аллизина и
Дезаминирование лизина и гидроксилизина с образованием реакционноспособного альдегида – аллизина и

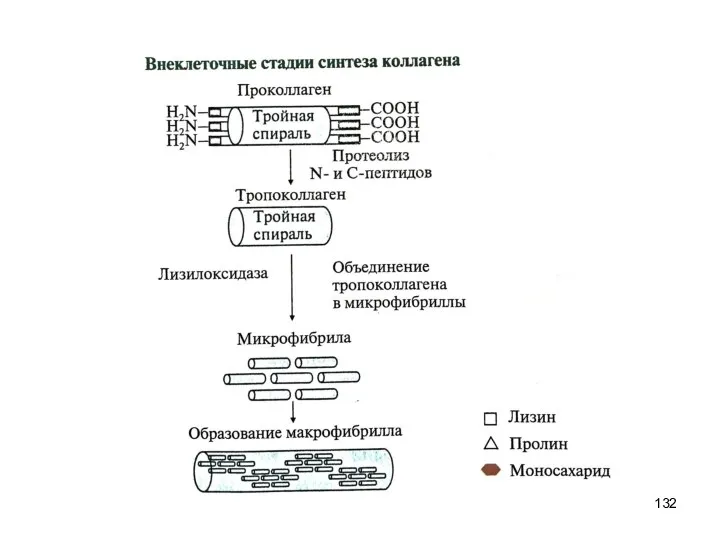
Макрофибрилла коллагена
Макрофибрилла коллагена
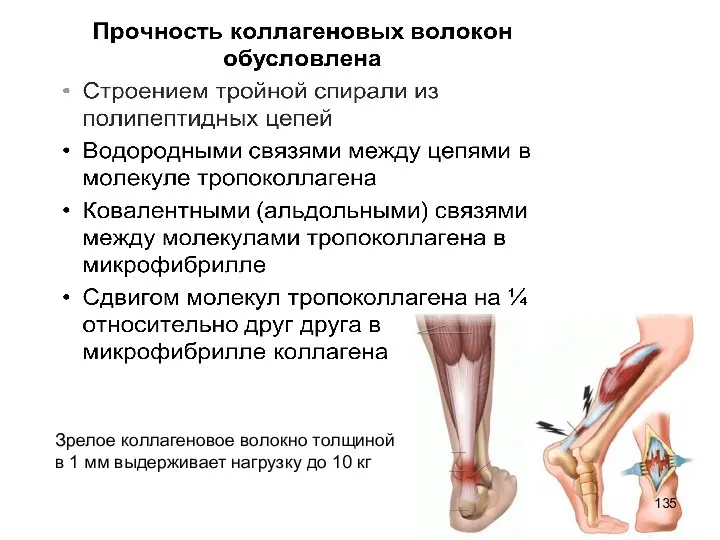
Зрелое коллагеновое волокно толщиной в 1 мм выдерживает нагрузку до 10
Зрелое коллагеновое волокно толщиной в 1 мм выдерживает нагрузку до 10

Катаболизм коллагена
Коллаген – медленно обменивающийся белок (время полужизни – недели, месяцы).
Катаболизм коллагена
Коллаген – медленно обменивающийся белок (время полужизни – недели, месяцы).

Регуляция обмена коллагена
Аскорбиновая кислота стимулирует синтеза коллагена, протеогликанов и пролиферацию фибробластов
Половые
Регуляция обмена коллагена
Аскорбиновая кислота стимулирует синтеза коллагена, протеогликанов и пролиферацию фибробластов
Половые

При старении
увеличивается количество поперечных сшивок между молекулами тропоколлагена, затрудняющих действие
При старении
увеличивается количество поперечных сшивок между молекулами тропоколлагена, затрудняющих действие

Метаболизм коллагена
При патологии
Нарушение катаболизма приводит к фиброзу органов
Усиление распада происходит
Метаболизм коллагена
При патологии
Нарушение катаболизма приводит к фиброзу органов
Усиление распада происходит

Структурные белки соединительной ткани:
Эластин
Основной компонент волокон тканей, обладающих значительной эластичностью (сосуды,
Структурные белки соединительной ткани:
Эластин
Основной компонент волокон тканей, обладающих значительной эластичностью (сосуды,

Эластин
Время полураспада – 75 лет
Протеолиз под действием эластазы (нейтрофилы)
Нарушения образования
Эластин
Время полураспада – 75 лет
Протеолиз под действием эластазы (нейтрофилы)
Нарушения образования

Адгезивные белки соединительной ткани:
фибронектин
Димер (2 цепи сходны, но не идентичны, соединены
Адгезивные белки соединительной ткани:
фибронектин
Димер (2 цепи сходны, но не идентичны, соединены
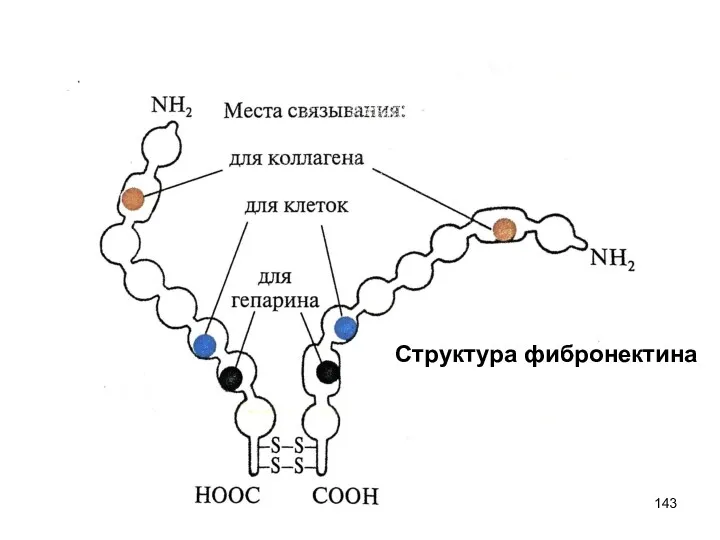
Структура фибронектина
Структура фибронектина
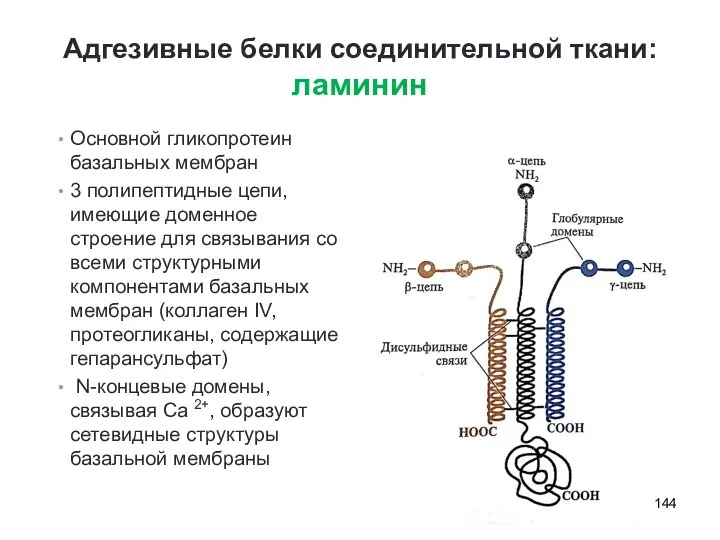
Адгезивные белки соединительной ткани:
ламинин
Основной гликопротеин базальных мембран
3 полипептидные цепи, имеющие доменное
Адгезивные белки соединительной ткани:
ламинин
Основной гликопротеин базальных мембран
3 полипептидные цепи, имеющие доменное

Адгезивные белки соединительной ткани:
нидоген
Сульфатированный гликопротеин базальных мембран
Одна полипептидная цепь
Три глобулярных домена:
Адгезивные белки соединительной ткани:
нидоген
Сульфатированный гликопротеин базальных мембран
Одна полипептидная цепь
Три глобулярных домена:

Кость – минерализованная соединительная ткань
Функции: размер и форма тела, защита внутренних
Кость – минерализованная соединительная ткань
Функции: размер и форма тела, защита внутренних

Костный обмен
Костный обмен (ремоделирование) включает:
резорбцию (разрушение) костной ткани
костеобразование (обновление, реорганизация
Костный обмен
Костный обмен (ремоделирование) включает:
резорбцию (разрушение) костной ткани
костеобразование (обновление, реорганизация

Минеральный состав и строение апатитов
Ячейка гидроксиапатита
Кристалл гидроксиапатита состоит из 2000 ячеек
Гидроксиапатит
Минеральный состав и строение апатитов
Ячейка гидроксиапатита
Кристалл гидроксиапатита состоит из 2000 ячеек
Гидроксиапатит

Строение апатитов
Оптимальное расстояние между ионами Са2+ и РО43- задается органической
Строение апатитов
Оптимальное расстояние между ионами Са2+ и РО43- задается органической

Органические вещества кости: коллаген
Особенности строения коллагена I типа (остеоколлагена):
Больше гидроксипролина, но
Органические вещества кости: коллаген
Особенности строения коллагена I типа (остеоколлагена):
Больше гидроксипролина, но

Органические вещества кости: неколлагеновые белки
Большинство неколлагеновых белков – гликопротеины или гликофосфопротеины;
Органические вещества кости: неколлагеновые белки
Большинство неколлагеновых белков – гликопротеины или гликофосфопротеины;

Органические вещества кости:
неколлагеновые белки
Неколлагеновые белки, содержащие RGD-последовательность: -Арг-Глу-Асп- (комплементарна рецепторам остеокластов):
Сиалопротеин,
Органические вещества кости:
неколлагеновые белки
Неколлагеновые белки, содержащие RGD-последовательность: -Арг-Глу-Асп- (комплементарна рецепторам остеокластов):
Сиалопротеин,

Другие органические вещества кости
Факторы роста и дифференцировки (IGF-1, TGF-β, PDGF)
Протеогликаны (ГАГ:
Другие органические вещества кости
Факторы роста и дифференцировки (IGF-1, TGF-β, PDGF)
Протеогликаны (ГАГ:
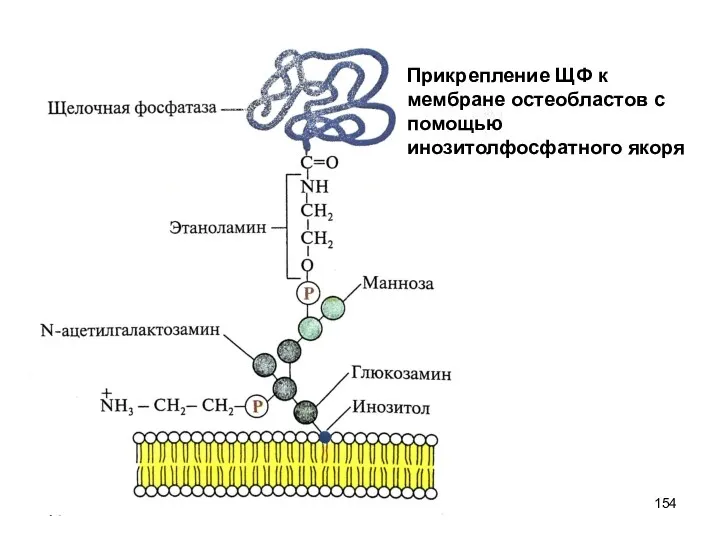
Прикрепление ЩФ к мембране остеобластов с помощью инозитолфосфатного якоря
Прикрепление ЩФ к мембране остеобластов с помощью инозитолфосфатного якоря

Роль щелочной фосфатазы в костной ткани
Проявляет активность на стадии минерализации (рН
Роль щелочной фосфатазы в костной ткани
Проявляет активность на стадии минерализации (рН

Ремоделирование костной ткани
Каждые 10 лет костная ткань заменяется новой (ежегодно обновляется
Ремоделирование костной ткани
Каждые 10 лет костная ткань заменяется новой (ежегодно обновляется

Этапы цикла ремоделирования костной ткани
1. Активация: остеобласты под влиянием стимулирующих факторов
Этапы цикла ремоделирования костной ткани
1. Активация: остеобласты под влиянием стимулирующих факторов

Этапы цикла ремоделирования костной ткани
2. Резорбция: «работа» остеокластов
Прикрепление остеокласта
Этапы цикла ремоделирования костной ткани
2. Резорбция: «работа» остеокластов
Прикрепление остеокласта
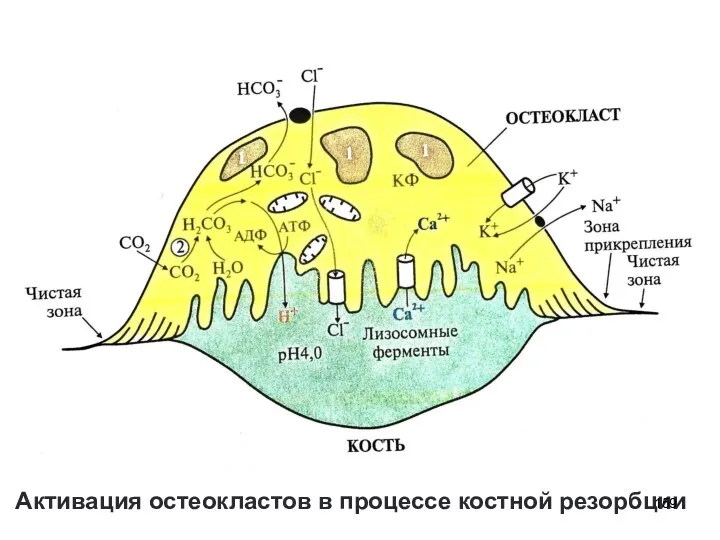
Активация остеокластов в процессе костной резорбции
Активация остеокластов в процессе костной резорбции

Этапы цикла ремоделирования костной ткани
3. Реверсия
Действие локальных ростовых факторов на
Этапы цикла ремоделирования костной ткани
3. Реверсия
Действие локальных ростовых факторов на

Регуляция ремоделирования
Паракринно-аутокринная (местная) регуляция: цитокины, фаторы роста, простагландины костной ткани
Системная регуляция
Регуляция ремоделирования
Паракринно-аутокринная (местная) регуляция: цитокины, фаторы роста, простагландины костной ткани
Системная регуляция
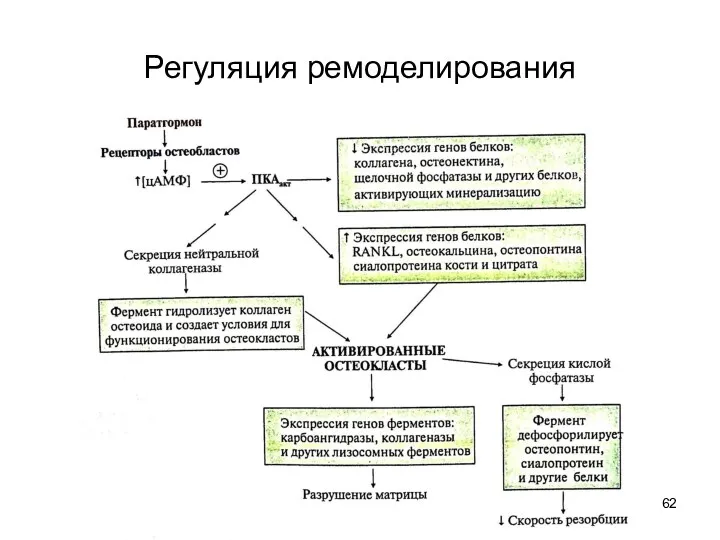
Регуляция ремоделирования
Регуляция ремоделирования
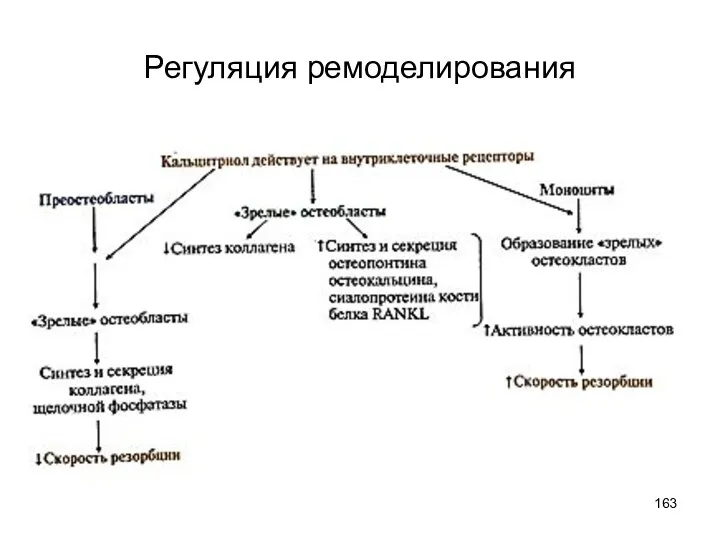
Регуляция ремоделирования
Регуляция ремоделирования

Регуляция ремоделирования
ПК А фосфорилирует белок-переносчик Са2+, что приводит к поступлению Са2+
Регуляция ремоделирования
ПК А фосфорилирует белок-переносчик Са2+, что приводит к поступлению Са2+

Регуляция ремоделирования
Кортизол замедляет костеобразование: подавляет пролиферацию остеобластов, синтез остеоколлагена, остеонектина, протеогликанов
Кортизол
Регуляция ремоделирования
Кортизол замедляет костеобразование: подавляет пролиферацию остеобластов, синтез остеоколлагена, остеонектина, протеогликанов
Кортизол

Регуляция ремоделирования
Гормон роста повышает активность остеобластов, которые стимулируют остеокласты, поэтому происходит
Регуляция ремоделирования
Гормон роста повышает активность остеобластов, которые стимулируют остеокласты, поэтому происходит
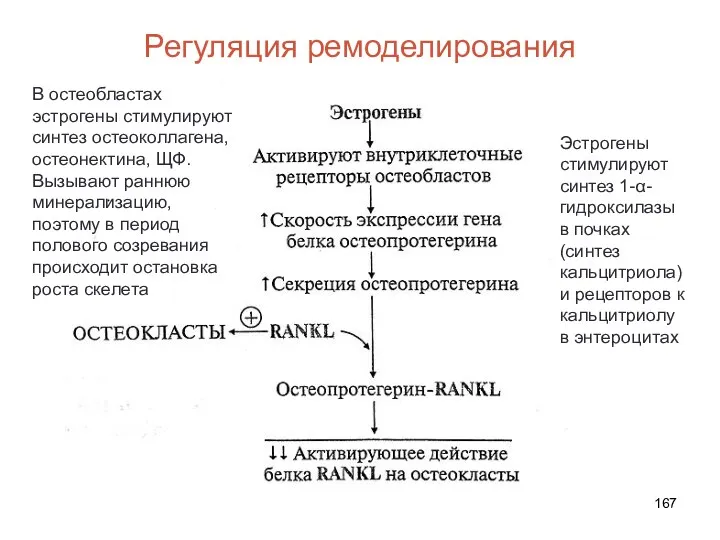
Регуляция ремоделирования
В остеобластах эстрогены стимулируют синтез остеоколлагена, остеонектина, ЩФ. Вызывают раннюю
Регуляция ремоделирования
В остеобластах эстрогены стимулируют синтез остеоколлагена, остеонектина, ЩФ. Вызывают раннюю
 Гели косметические
Гели косметические Популяциялық генетика негіздері. Генетикалық полиморфизм
Популяциялық генетика негіздері. Генетикалық полиморфизм Мембрана и органоиды клетки. 9 класс
Мембрана и органоиды клетки. 9 класс Класс Птицы
Класс Птицы Система инактивации лекарственных средств в организме человека
Система инактивации лекарственных средств в организме человека Формирование экологической компетентности школьников через интеграцию общего и дополнительного образования
Формирование экологической компетентности школьников через интеграцию общего и дополнительного образования Презентация по биологии Отдел Плауновидные и отдел Хвощевидные.
Презентация по биологии Отдел Плауновидные и отдел Хвощевидные. Приспособились ли потомки древних обитателей Земли – бактерии – к жизни на современной планете?
Приспособились ли потомки древних обитателей Земли – бактерии – к жизни на современной планете? Внутриутробное развитие плода. Развитие плода с первых дней беременности
Внутриутробное развитие плода. Развитие плода с первых дней беременности Витамины. Определение, классификация
Витамины. Определение, классификация Плесневые грибы и дрожжи. Грибы-паразиты
Плесневые грибы и дрожжи. Грибы-паразиты Общая спланхнология
Общая спланхнология Слуховой анализатор
Слуховой анализатор Методы цитологии. Клеточная теория
Методы цитологии. Клеточная теория Презентация по биологии для учащихся 8-го класса на тему: Эволюция человека
Презентация по биологии для учащихся 8-го класса на тему: Эволюция человека Размножение организмов
Размножение организмов Растительные сообщества городской системы
Растительные сообщества городской системы Простые и сложные углеводы. Гликемический индекс
Простые и сложные углеводы. Гликемический индекс Антропогенез. Расы. Расизм. Часть 9
Антропогенез. Расы. Расизм. Часть 9 Энергообразующие органоиды (лекция 7)
Энергообразующие органоиды (лекция 7) Приспособленность организмов к условиям внешней среды
Приспособленность организмов к условиям внешней среды Постэмбриональное развитие животных
Постэмбриональное развитие животных Человек как уникальный вид живой природы
Человек как уникальный вид живой природы Открытия в области биологии в эпоху НТР
Открытия в области биологии в эпоху НТР Ткани растений и животных
Ткани растений и животных Отдел Покрытосеменные
Отдел Покрытосеменные Рыбы (7 класс)
Рыбы (7 класс) Тип Моллюски или мягкотелые
Тип Моллюски или мягкотелые