- Биохимия нервной ткани. (Лекция 23)

Содержание
- 2. План лекции Нервная ткань: определение понятия, классификация по функциям и клеточному составу. Нейрон – как основная
- 3. Функции нервной системы: воспринимает информацию из внешней и внутренней среды; перерабатывает полученную информацию; хранит информацию; генерирует
- 4. Классификация нервной системы Анатомически Физиологически ЦНС ПНС Соматическая Автономная Симпатическая Парасимпатическая головной и спинной мозг периферические
- 5. Функциональной тканью нервной системы является нервная. Нервная ткань – это высокоспециализированная ткань, обладающая возбудимостью и проводимостью,
- 6. Нейрон
- 7. Нейроглия (от греческого glia – клей) это клетки нервной системы, которые не проводят нервные импульсы. Глиальные
- 8. Макроглия 1. Астроцитарная глия Астроциты обеспечивает микроокружение нейронов, выполняет опорную и трофическую функции в сером и
- 9. образует выстилку желудочков головного мозга входит в состав гематоликворного барьера. На снимке - просвет одного из
- 10. встречается в сером и белом веществе; обеспечивает барьерную функцию, участвует в формировании миелиновых оболочек нервных волокон,
- 11. Микроглия – специализированные макрофаги ЦНС. Способны к амёбоидным движениям и фагоцитозу Активизируются при воспалительных и дегенеративных
- 12. Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии (астроциты, олигодендроциты) в ЦНС называется
- 13. Химический состав нервной ткани
- 14. Белки нервной ткани сложные простые В головном мозге на белки приходиться 40% сухой массы. В настоящее
- 15. Простые белки Нейроальбумины – основные растворимые белки (80%) Нейроглобулины - 5%. Катионные белки - основные белки
- 16. Сложные белки Гликопротеины –нейрорецепция Протеолипиды – структурная ф -я Нейроспецифические белки Нейроспецифическая енолаза (Белок 14-3-2) -
- 17. Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в астроцитах) и интенсивно нарабатывается в
- 18. НЕЙРОПЕПТИДЫ - эндогенные регуляторы функций ЦНС Осуществляют контроль: за экспрессией вторичных клеточных мессенджеров, цитокинов и других
- 19. Опиоидные нейропептиды
- 20. нейропептиды
- 21. Нейротрофические факторы:
- 22. Ферменты Нейроспецифическая енолаза ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии), АСТ, альдолаза, креатинкиназа (ВВ),
- 23. Аминокислоты Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ Аминокислот в мозге в среднем 34ммоль
- 24. Липиды нервной ткани фосфоглицериды в сером веществе составляют более 60% от всех липидов, а в белом
- 25. Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез ганглиозидов связан с дифференциацией нейронов.
- 26. Углеводы нервной ткани много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей индивидуальность и специфичность. мало
- 27. Нуклеотиды нервной ткани Клетки Пуркинье мозжечка содержат избыточное количество ДНК. Необычно короткие нуклеосомные единицы, наличие редких
- 28. Макроэргические соединения нервной ткани Содержание креатина и креатинфосфата более, чем в 2 раза превышает количество адениновых
- 29. Минеральные вещества нервной ткани Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+ распределены в головном мозге
- 30. Строение нервного волокна. Миелиновая оболочка 1 2 3 1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои) 5-липиды;
- 31. Образован: клетками эндотелия капилляров плотной базальной мембраной, не имеющей пор. Астроглией, выстилающие наружную поверхность эндотелия. Р-ГП
- 32. Особенности метаболизма нервной ткани Для мозга характерна высокая интенсивность энергетического обмена с преобладанием аэробных процессов. Головной
- 33. Основной энергетический субстрат для нервной ткани - глюкоза За 1 минуту 100 г ткани мозга потребляют
- 34. Обмен белков и аминокислот нервной ткани Нервная ткань характеризуется высоким обменом аминокислот и белков. Максимальная скорость
- 35. Обмен азота нервной ткани Источником аммиака в головном мозге служит непрямое дезаминирование аминокислот с участием глутаматдегидрогеназы,
- 36. Липидный обмен нервной ткани Особенность: липиды не используются в качестве энергетического материала, а в основном идут
- 37. Механизмы передачи нервного импульса
- 38. Механизмы передачи нервного импульса по нервному волокну 1. Потенциал покоя (-70 мВ) Na-канал К-канал 2. Потенциал
- 39. Са Na K Синтез медиатора Загрузка в везикулу Слияние везикул Специфические рецепторы Открытие Na канала Проведение
- 40. Аминокислотные медиаторы ингибиторные нейтральные (ГАМК, глицин, β-аланин и таурин). возбуждающие кислые (глутамат и аспартат) Глицин открывает
- 41. открывает хлорные каналы вызывает гиперполяризацию тормозит возбудимость постсинаптической мембраны ГАМК
- 42. Синтез холина Печень Нейроны
- 43. Синтез и распад медиатора на примере ацетилхолина (CH3)3N-CH2-CH2-OH холинацетилтрансфераза (CH3)3N-CH2-CH2-O-CO-CH3 H2O ацетилхолинэстераза (CH3)3N-CH2-CH2OH Нейрон ПЕЧЕНЬ CH3-CO-S-KoA
- 44. Рецепторы Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри клетки, которые, взаимодействуя с
- 45. Биохимические синдромы в психиатрии Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина) Нарушения окислительно-восстановительных процессов (гипоксия, накопление молочной к-ты,
- 46. Патобиохимия нервной системы 1) Миастения - уменьшение числа холинорецепторов 3) болезнь Паркинсона - дегенерация дофамин содержащих
- 47. Нейроспецифические белки- МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ белок S100 Нейрон специфическая енолаза (NSE) Нейроны Астроциты Олигодендроциты и миелин
- 48. Нейронспецифическая енолаза (NSE) Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент, катализирующий превращение 2-фосфоглицерата в
- 49. Белок S100 S-100 - кальций-связывающий протеин. Локализация мембраны, цитоплазма. участвует в фосфорилировании белка, обеспечивает работу цитоскелета
- 50. антитела к S100β являются специфичными маркерами повреждения астроцитарной глии. Ген S100 находиться в длинном плече 21-й
- 51. Основной белок миелина (MBP) MBP - щелочной белок, с высоким содержанием (25%) основных аминокислот (аргинина, лизина,
- 52. NSE, S100 и MBP – маркеры повреждения мозговой ткани Нейроспецифические белки в нормальных условиях обнаруживаются в
- 53. NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также других опухолей нейроэктодермального или нейроэндокринного
- 54. концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени, чем у женщин NSE, S100
- 55. Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции в ряде периферических органов (
- 56. Катаболизм дофамина: Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»). В везикулу с помощью протон-зависимой
- 57. ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА Различают семь отдельных подсистем (первые три являются основными): 1. Нигростриатная 2. Мезокортикальная
- 58. Пути Дофамина: Патология: Патология Патология
- 59. Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы, ДОФА-декарбоксилазы. Патология Дофамина. Болезнь
- 60. Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы, ДОФА-декарбоксилазы. Патология Дофамина. Болезнь
- 61. 1.Журнал экспериментальная и клиническая фармакология 2007 г. том 70. №4 с.11 «изучение эффектов острого и хронического
- 63. Скачать презентацию

План лекции
Нервная ткань: определение понятия, классификация по функциям и клеточному составу.
Нейрон
План лекции
Нервная ткань: определение понятия, классификация по функциям и клеточному составу.
Нейрон

Функции нервной системы:
воспринимает информацию из внешней и внутренней среды;
перерабатывает полученную информацию;
хранит
Функции нервной системы:
воспринимает информацию из внешней и внутренней среды;
перерабатывает полученную информацию;
хранит
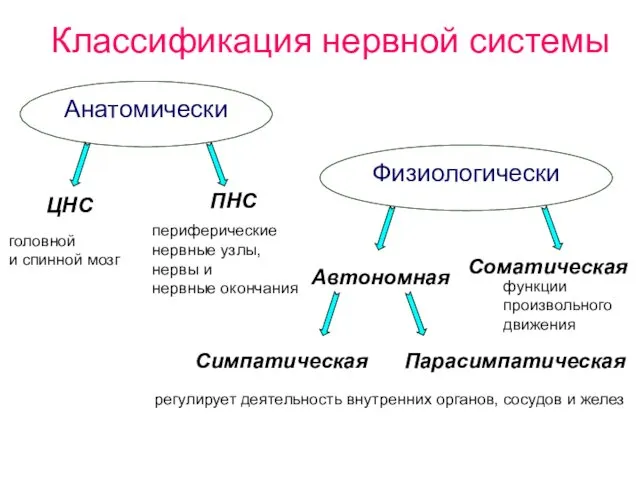
Классификация нервной системы
Анатомически
Физиологически
ЦНС
ПНС
Соматическая
Автономная
Симпатическая
Парасимпатическая
головной
и спинной
Классификация нервной системы
Анатомически
Физиологически
ЦНС
ПНС
Соматическая
Автономная
Симпатическая
Парасимпатическая
головной
и спинной

Функциональной тканью нервной системы является нервная.
Нервная ткань – это высокоспециализированная
Функциональной тканью нервной системы является нервная.
Нервная ткань – это высокоспециализированная
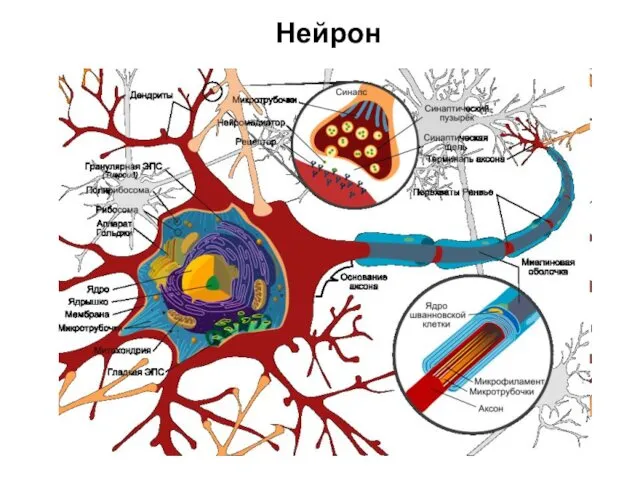
Нейрон
Нейрон

Нейроглия (от греческого glia – клей) это клетки нервной системы, которые
Нейроглия (от греческого glia – клей) это клетки нервной системы, которые

Макроглия
1. Астроцитарная глия
Астроциты
обеспечивает микроокружение нейронов,
выполняет опорную и трофическую функции
Макроглия
1. Астроцитарная глия
Астроциты
обеспечивает микроокружение нейронов,
выполняет опорную и трофическую функции
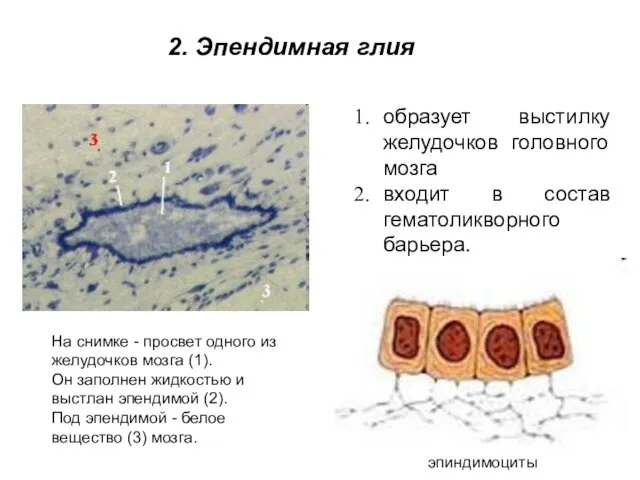
образует выстилку желудочков головного мозга
входит в состав гематоликворного барьера.
На снимке
образует выстилку желудочков головного мозга
входит в состав гематоликворного барьера.
На снимке

встречается в сером и белом веществе;
обеспечивает барьерную функцию,
участвует в
встречается в сером и белом веществе;
обеспечивает барьерную функцию,
участвует в

Микроглия – специализированные макрофаги ЦНС.
Способны к амёбоидным движениям и фагоцитозу
Активизируются
Микроглия – специализированные макрофаги ЦНС.
Способны к амёбоидным движениям и фагоцитозу
Активизируются

Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии
Серое вещество образовано скоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии

Химический состав нервной ткани
Химический состав нервной ткани
Белки нервной ткани
сложные
простые
В головном мозге на белки приходиться 40% сухой
Белки нервной ткани
сложные
простые
В головном мозге на белки приходиться 40% сухой

Простые белки
Нейроальбумины – основные растворимые белки (80%)
Нейроглобулины - 5%.
Катионные
Простые белки
Нейроальбумины – основные растворимые белки (80%)
Нейроглобулины - 5%.
Катионные

Сложные белки
Гликопротеины –нейрорецепция
Протеолипиды – структурная ф -я
Нейроспецифические белки
Нейроспецифическая енолаза (Белок
Сложные белки
Гликопротеины –нейрорецепция
Протеолипиды – структурная ф -я
Нейроспецифические белки
Нейроспецифическая енолаза (Белок

Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в
Белок S100 - гетерогенный кислый Сa-связывающий белок, локализуется в нейроглии (в

НЕЙРОПЕПТИДЫ - эндогенные регуляторы функций ЦНС
Осуществляют контроль:
за экспрессией
НЕЙРОПЕПТИДЫ - эндогенные регуляторы функций ЦНС
Осуществляют контроль:
за экспрессией

Опиоидные нейропептиды
Опиоидные нейропептиды
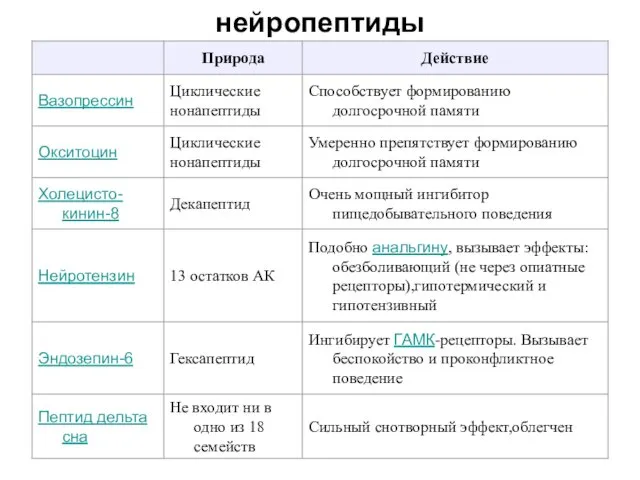
нейропептиды
нейропептиды
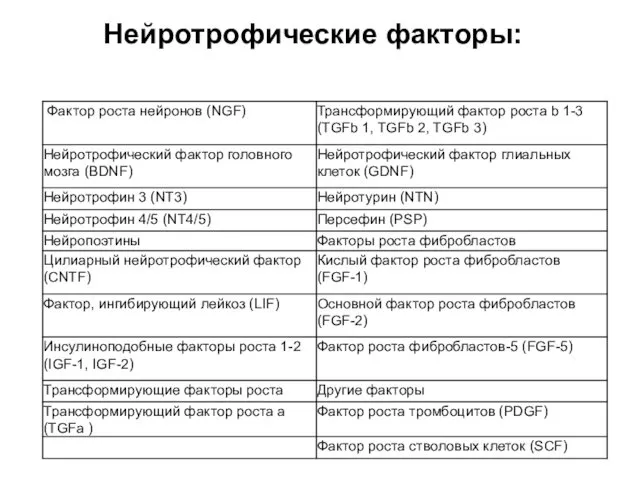
Нейротрофические факторы:
Нейротрофические факторы:

Ферменты
Нейроспецифическая енолаза
ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии),
Ферменты
Нейроспецифическая енолаза
ЛДГ (ЛДГ1, ЛДГ2 в нейронах, ЛДГ5 - в глии),
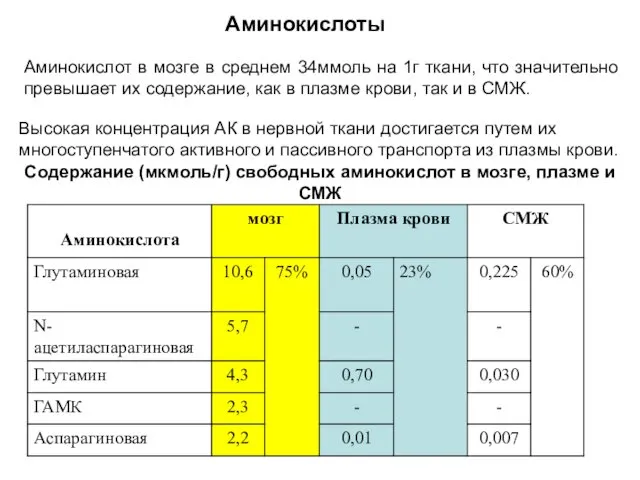
Аминокислоты
Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ
Аминокислот в мозге
Аминокислоты
Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ
Аминокислот в мозге

Липиды нервной ткани
фосфоглицериды в сером веществе составляют более 60% от
Липиды нервной ткани
фосфоглицериды в сером веществе составляют более 60% от

Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез
Ганглиозиды - в сером веществе - Gм1, GD1a, GD1b, GT1. Синтез

Углеводы нервной ткани
много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей
Углеводы нервной ткани
много олигосахаров (составляют 2-10% массы плазматической мембраны) придают ей

Нуклеотиды нервной ткани
Клетки Пуркинье мозжечка содержат избыточное количество ДНК.
Необычно короткие нуклеосомные
Нуклеотиды нервной ткани
Клетки Пуркинье мозжечка содержат избыточное количество ДНК.
Необычно короткие нуклеосомные

Макроэргические соединения нервной ткани
Содержание креатина и креатинфосфата более, чем в 2
Макроэргические соединения нервной ткани
Содержание креатина и креатинфосфата более, чем в 2

Минеральные вещества нервной ткани
Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+
Минеральные вещества нервной ткани
Na+, K+, Cu2+, Fe2+, Ca2+, Mg2+ и Mn2+
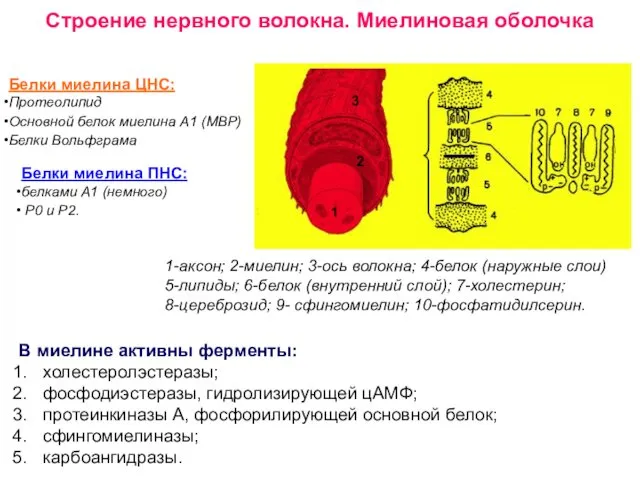
Строение нервного волокна. Миелиновая оболочка
1
2
3
1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои)
Строение нервного волокна. Миелиновая оболочка
1
2
3
1-аксон; 2-миелин; 3-ось волокна; 4-белок (наружные слои)
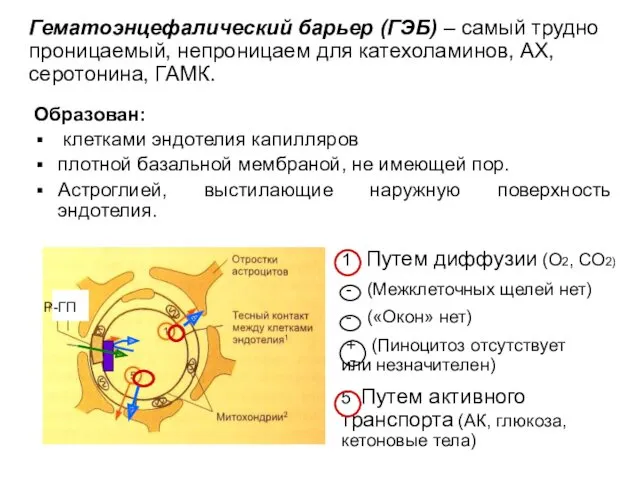
Образован:
клетками эндотелия капилляров
плотной базальной мембраной, не имеющей пор.
Астроглией,
Образован:
клетками эндотелия капилляров
плотной базальной мембраной, не имеющей пор.
Астроглией,

Особенности метаболизма нервной ткани
Для мозга характерна высокая интенсивность энергетического обмена с
Особенности метаболизма нервной ткани
Для мозга характерна высокая интенсивность энергетического обмена с

Основной энергетический субстрат для нервной ткани - глюкоза
За 1 минуту 100
Основной энергетический субстрат для нервной ткани - глюкоза
За 1 минуту 100

Обмен белков и аминокислот нервной ткани
Нервная ткань характеризуется высоким обменом аминокислот
Обмен белков и аминокислот нервной ткани
Нервная ткань характеризуется высоким обменом аминокислот
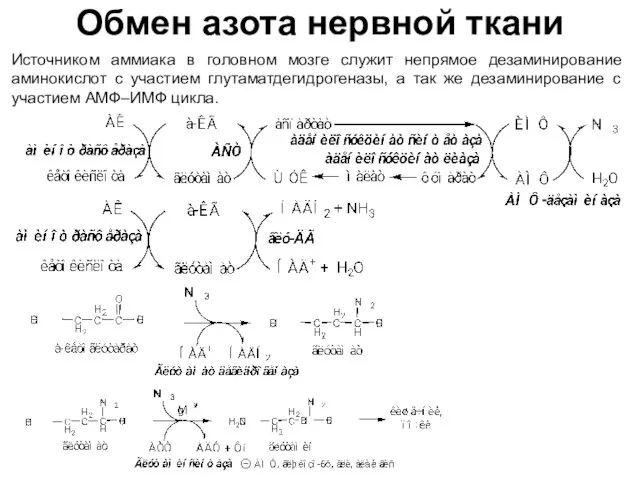
Обмен азота нервной ткани
Источником аммиака в головном мозге служит непрямое
Обмен азота нервной ткани
Источником аммиака в головном мозге служит непрямое

Липидный обмен нервной ткани
Особенность: липиды не используются в качестве энергетического материала,
Липидный обмен нервной ткани
Особенность: липиды не используются в качестве энергетического материала,

Механизмы передачи
нервного импульса
Механизмы передачи
нервного импульса

Механизмы передачи нервного импульса
по нервному волокну
1. Потенциал покоя (-70 мВ)
Na-канал
К-канал
2.
Механизмы передачи нервного импульса
по нервному волокну
1. Потенциал покоя (-70 мВ)
Na-канал
К-канал
2.
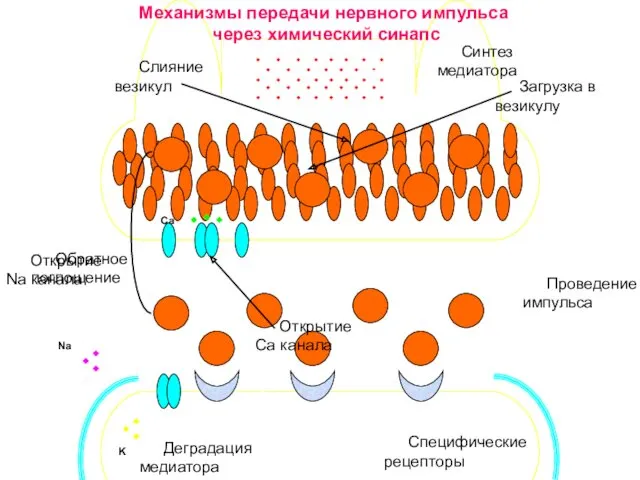
Са
Na
K
Синтез медиатора
Загрузка в везикулу
Слияние везикул
Специфические рецепторы
Открытие Na канала
Проведение импульса
Открытие Сa канала
Деградация
Са
Na
K
Синтез медиатора
Загрузка в везикулу
Слияние везикул
Специфические рецепторы
Открытие Na канала
Проведение импульса
Открытие Сa канала
Деградация
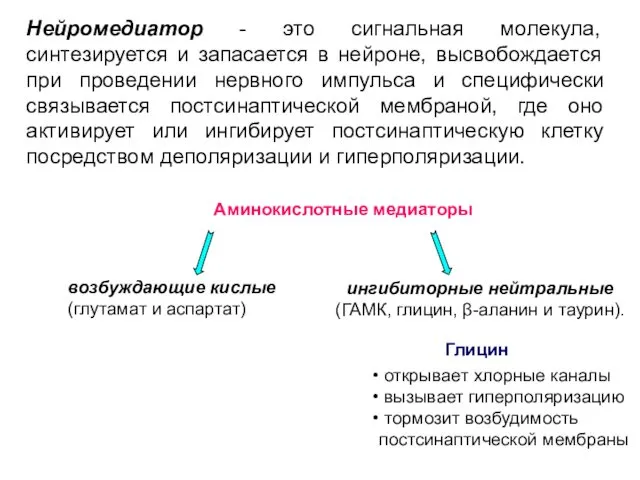
Аминокислотные медиаторы
ингибиторные нейтральные
(ГАМК, глицин, β-аланин и таурин).
возбуждающие кислые
(глутамат и
Аминокислотные медиаторы
ингибиторные нейтральные
(ГАМК, глицин, β-аланин и таурин).
возбуждающие кислые
(глутамат и
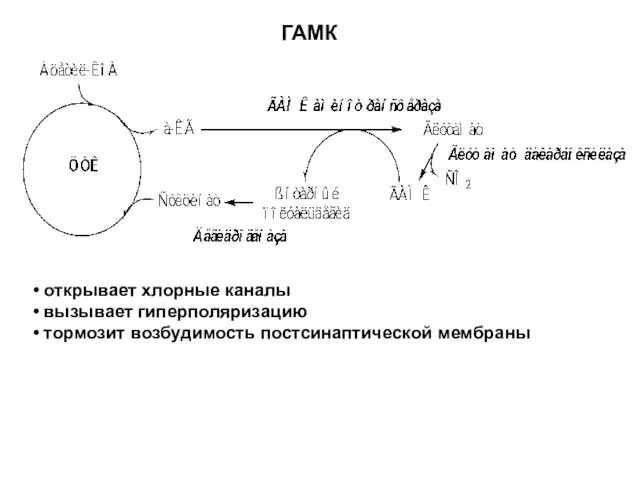
открывает хлорные каналы
вызывает гиперполяризацию
тормозит возбудимость постсинаптической мембраны
открывает хлорные каналы
вызывает гиперполяризацию
тормозит возбудимость постсинаптической мембраны
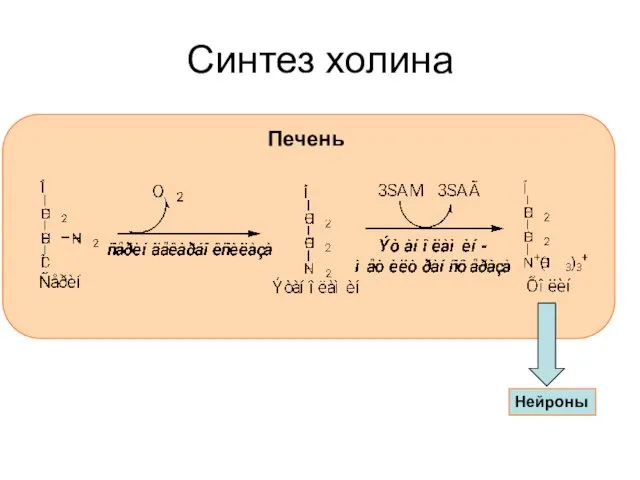
Синтез холина
Печень
Нейроны
Синтез холина
Печень
Нейроны
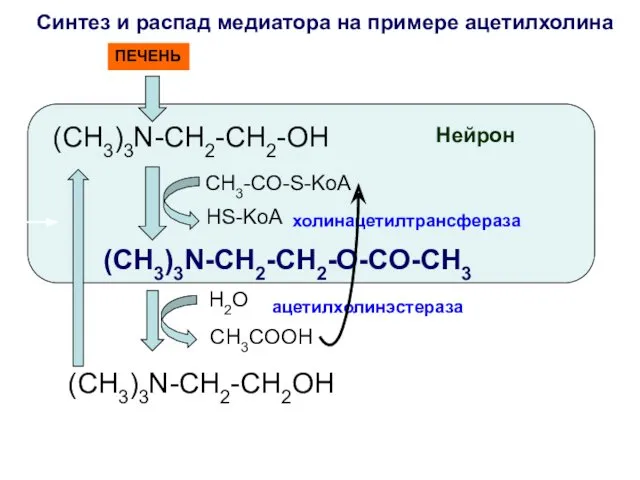
Синтез и распад медиатора на примере ацетилхолина
(CH3)3N-CH2-CH2-OH
холинацетилтрансфераза
(CH3)3N-CH2-CH2-O-CO-CH3
H2O
ацетилхолинэстераза
(CH3)3N-CH2-CH2OH
Нейрон
ПЕЧЕНЬ
CH3-CO-S-KoA
HS-KoA
CH3COOH
Синтез и распад медиатора на примере ацетилхолина
(CH3)3N-CH2-CH2-OH
холинацетилтрансфераза
(CH3)3N-CH2-CH2-O-CO-CH3
H2O
ацетилхолинэстераза
(CH3)3N-CH2-CH2OH
Нейрон
ПЕЧЕНЬ
CH3-CO-S-KoA
HS-KoA
CH3COOH

Рецепторы
Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри
Рецепторы
Рецепторы - это белки, встроенные в клеточную мембрану или находящиеся внутри

Биохимические синдромы в психиатрии
Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина)
Нарушения окислительно-восстановительных процессов (гипоксия,
Биохимические синдромы в психиатрии
Интоксикации (аминокислоты, кетокислоты, аммиак, мочевина)
Нарушения окислительно-восстановительных процессов (гипоксия,

Патобиохимия нервной системы
1) Миастения - уменьшение числа холинорецепторов
3) болезнь Паркинсона -
Патобиохимия нервной системы
1) Миастения - уменьшение числа холинорецепторов
3) болезнь Паркинсона -

Нейроспецифические белки-
МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ
белок S100
Нейрон
специфическая
енолаза (NSE)
Нейроны
Астроциты
Олигодендроциты
и миелин
MBP
(его
Нейроспецифические белки-
МАРКЕРЫ НЕЙРОДЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ
белок S100
Нейрон
специфическая
енолаза (NSE)
Нейроны
Астроциты
Олигодендроциты
и миелин
MBP
(его

Нейронспецифическая енолаза (NSE)
Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент,
Нейронспецифическая енолаза (NSE)
Нейроспецифическая енолаза (антиген 14-3-2) NSE – это гликолитический фермент,

Белок S100
S-100 - кальций-связывающий протеин.
Локализация мембраны, цитоплазма.
участвует в фосфорилировании
Белок S100
S-100 - кальций-связывающий протеин.
Локализация мембраны, цитоплазма.
участвует в фосфорилировании

антитела к S100β являются специфичными маркерами повреждения астроцитарной глии.
Ген S100
Ген S100

Основной белок миелина (MBP)
MBP - щелочной белок, с высоким содержанием (25%)
Основной белок миелина (MBP)
MBP - щелочной белок, с высоким содержанием (25%)

NSE, S100 и MBP – маркеры повреждения мозговой ткани
Нейроспецифические белки в
NSE, S100 и MBP – маркеры повреждения мозговой ткани
Нейроспецифические белки в

NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также
NSE является высокоспецифичным маркером мелкоклеточного рака легкого и нейробластомы, а также

концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени,
концентрация белка НСБ увеличивается с возрастом, у мужчин в большей степени,
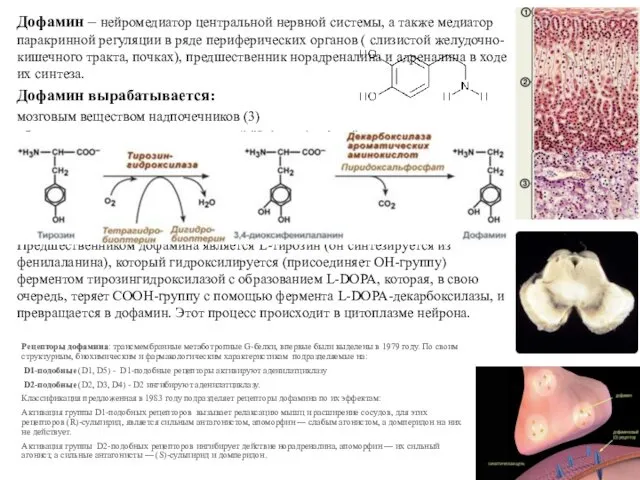
Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции
Дофамин – нейромедиатор центральной нервной системы, а также медиатор паракринной регуляции
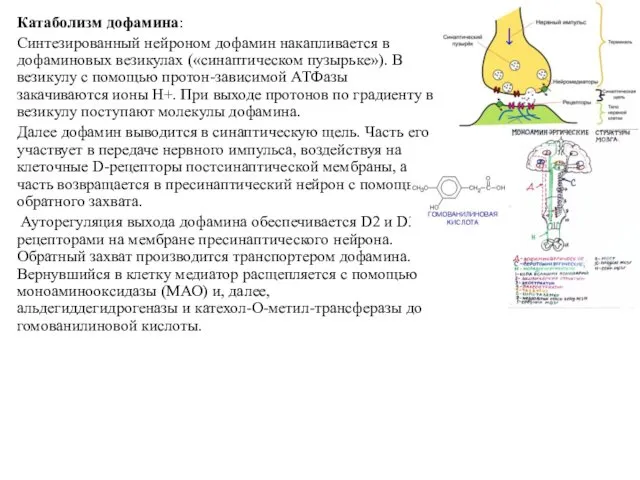
Катаболизм дофамина:
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»).
Катаболизм дофамина:
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах («синаптическом пузырьке»).
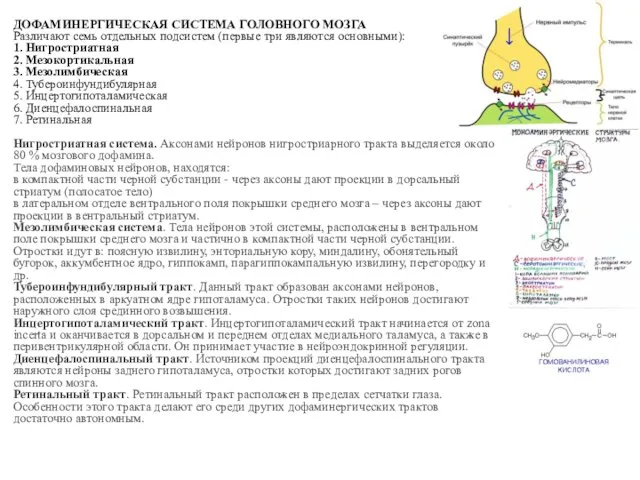
ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
Различают семь отдельных подсистем (первые три являются основными):
ДОФАМИНЕРГИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
Различают семь отдельных подсистем (первые три являются основными):
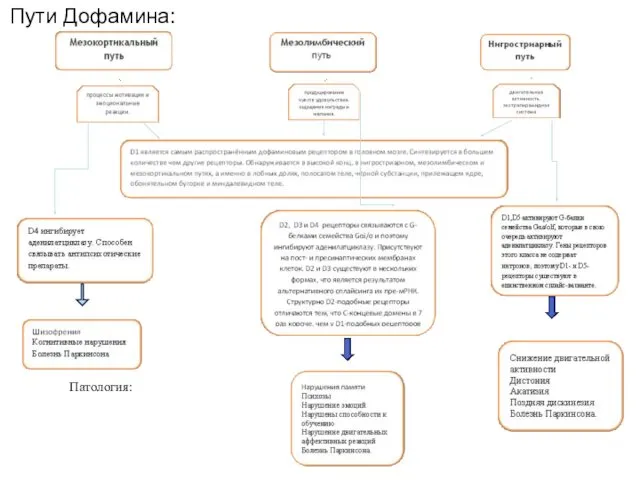
Пути Дофамина:
Патология:
Патология
Патология
Пути Дофамина:
Патология:
Патология
Патология
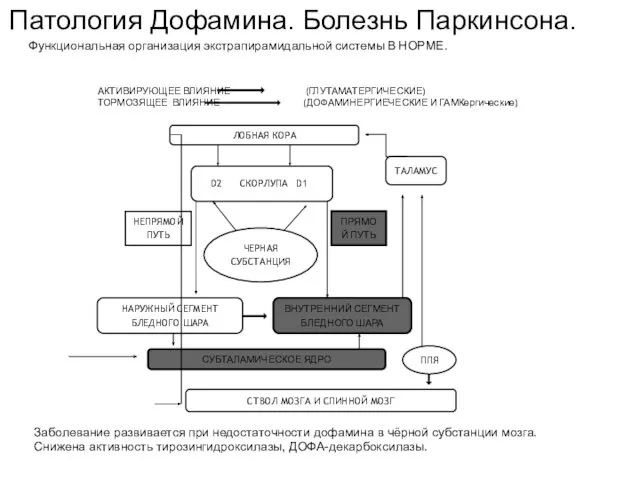
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга.
Снижена активность тирозингидроксилазы,
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы,

Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга.
Снижена активность тирозингидроксилазы,
Заболевание развивается при недостаточности дофамина в чёрной субстанции мозга. Снижена активность тирозингидроксилазы,

1.Журнал экспериментальная и клиническая фармакология 2007 г. том 70. №4 с.11
1.Журнал экспериментальная и клиническая фармакология 2007 г. том 70. №4 с.11
 Трихология. Волосы и кожа головы
Трихология. Волосы и кожа головы Ч.Дарвиннің. Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары,эволюцияның синтетикалық теориясы
Ч.Дарвиннің. Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары,эволюцияның синтетикалық теориясы Слуховой анализатор. Структура слухового анализатора, возрастные особенности
Слуховой анализатор. Структура слухового анализатора, возрастные особенности Биотические взаимоотношения организмов
Биотические взаимоотношения организмов Основные этапы эволюции человека
Основные этапы эволюции человека Транскрипция, посттранскрипционные модификации
Транскрипция, посттранскрипционные модификации Иван Владимирович Мичурин
Иван Владимирович Мичурин Влияние воды на прорастание семян
Влияние воды на прорастание семян Решение задач на моногибридное скрещивание и взаимодействие аллельных генов
Решение задач на моногибридное скрещивание и взаимодействие аллельных генов Царство Грибов. 5 класс
Царство Грибов. 5 класс Дикие и домашние животные
Дикие и домашние животные Роль микроорганизмов в природе и жизни человека
Роль микроорганизмов в природе и жизни человека Многообразие мира растений
Многообразие мира растений Неклеточные формы жизни. Вирусы
Неклеточные формы жизни. Вирусы Приспособленность живых организмов к среде обитания
Приспособленность живых организмов к среде обитания Общие свойства живых организмов
Общие свойства живых организмов Кожа. Железы кожи, волосы, ногти
Кожа. Железы кожи, волосы, ногти Морфология, систематика и физиология микроорганизмов
Морфология, систематика и физиология микроорганизмов Продуктивність екосистем. Ланцюги живлення
Продуктивність екосистем. Ланцюги живлення Подснежники
Подснежники Паразитические простейшие
Паразитические простейшие Биополимерлер және олардың құрылымдық құрам бөліктері
Биополимерлер және олардың құрылымдық құрам бөліктері Влияние витаминов на здоровье человека
Влияние витаминов на здоровье человека Строение волос
Строение волос Азбука здоровья от А до Я
Азбука здоровья от А до Я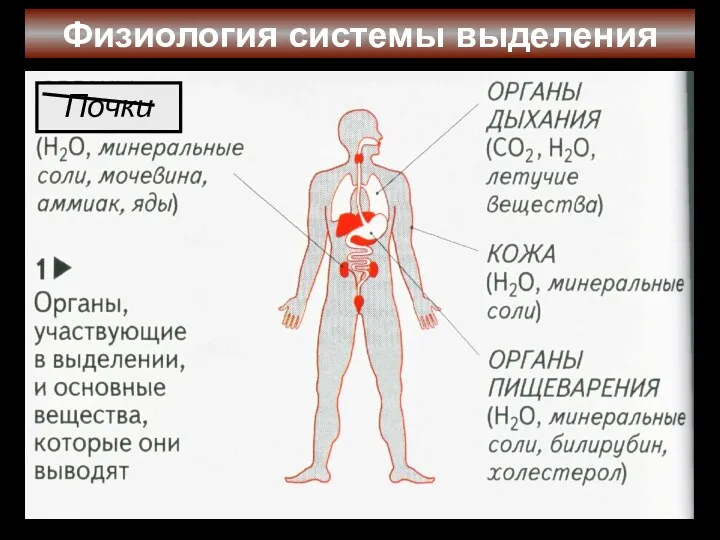 Физиология системы выделения. Почки
Физиология системы выделения. Почки животный мир австралии
животный мир австралии Мозговой ствол. Продолговатый мозг. Варолиев мост. Средний мозг
Мозговой ствол. Продолговатый мозг. Варолиев мост. Средний мозг