- Биологическая подвижность

Содержание
- 2. Механохимические процессы мышечное сокращение; движение клеток с помощью жгутиков и ресничек; все движения в митозе и
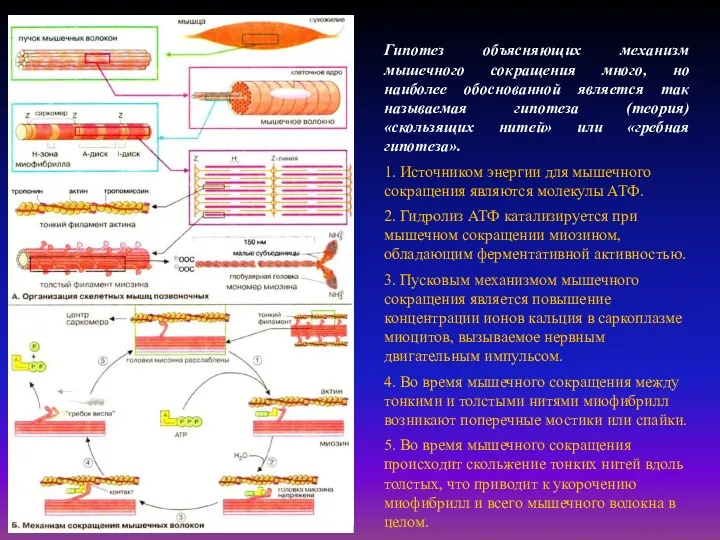
- 12. Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так называемая гипотеза (теория) «скользящих нитей»
- 14. Толстая нить состоит из большого числа молекул миозина, собранных в пучок. Каждая молекула миозина длиной 155
- 15. Благодаря огромному количеству работ посвященных мышечному сокращению [Бэгшоу С, 1985, Леднев В. В., 1983, Подлубная 3.
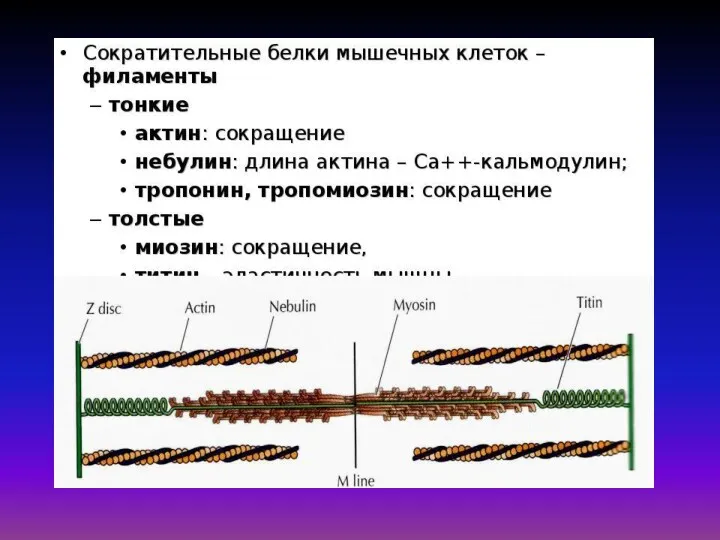
- 16. Актин — водорастворимый глобулярный белок (М 42 000), состоящий из 376 аминокислотных остатков (G-актин). С каждой
- 20. В клетках поперечнополосатых мышц в состав тонких нитей миофибрилл кроме актина и тропомиозина входит ещё и
- 21. Молекула тропонина обладает высоким сродством к кальцию. Под его влиянием она изменяет положение тропомиозиновой нити на
- 22. Структура и функционирование поперечнополосатой мышцы позвоночных. Диск I включает в себя только тонкие нити, а диск
- 23. Схема строения миофибрилл Схематическое изображение микрофотографии продольного среза мышцы: 1. саркомер 2. I диск 3. А-диск
- 24. Микроструктура саркомера
- 25. Модель мышечного сокращения Хаксли 1 – актиновый филамент, 2 – центр связывания, 3 – миозиновый филамент,
- 26. Молекулярный механизм мышечного сокращения. Модель скользящих нитей.
- 27. Основные положения модели скользящих нитей 1) Длина нитей актина и миозина в ходе сокращения не меняется.
- 28. Структурные элементы сократительного аппарата мышечных волокон (длина около 5 см)
- 29. Схема структурного взаимоотношения миофибриллы и саркоплазматического ретикулума мышечного волокна (Кальциевый насос) М и МТ – плазматическая
- 30. Схема временной последовательности развития потенциала действия, освобождения ионов Са2+ и развития изометрического сокращения мышцы ПД
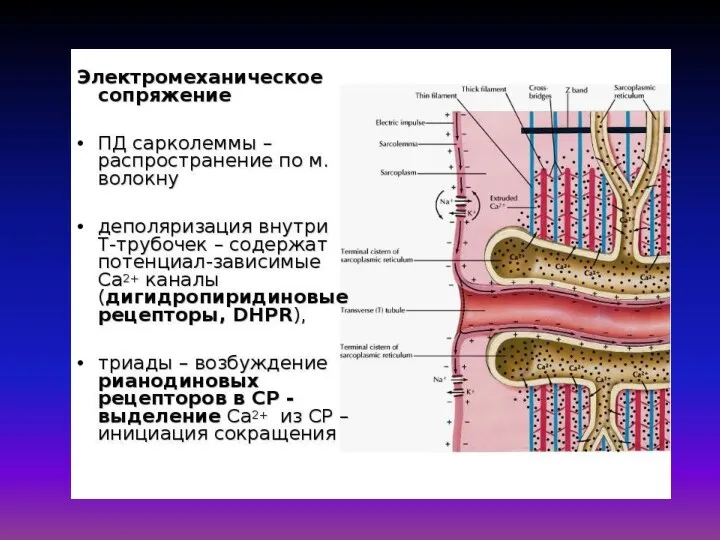
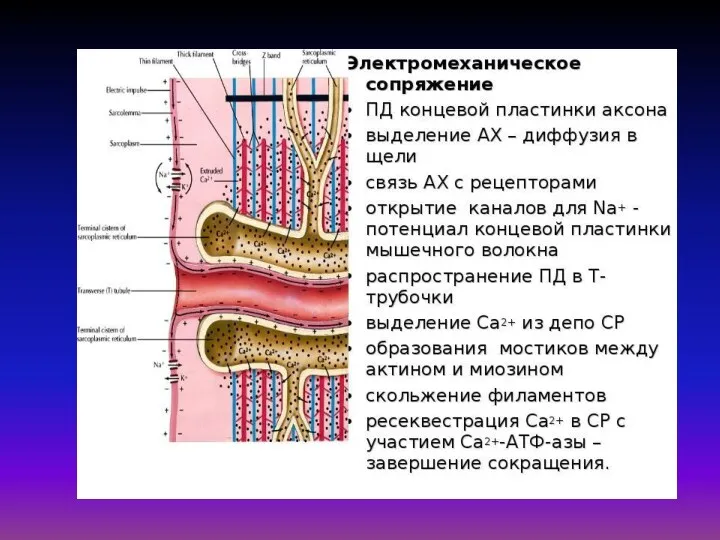
- 32. «Электромеханическое сопряжение» – это совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция
- 53. Математическая модель Дещеревского основана на модели «скользящих нитей» Хаксли В кинетической теории В. И. Дещеревского рассмотрена
- 54. Выведено английским физиологом А. В. Хиллом в 1938, выражает изменение скорости сокращения мышцы в зависимости от
- 55. Микротрубочки создают направление упорядоченного перемещения веществ в клетке. Встречаются в свободном состоянии в цитоплазме клеток или
- 57. Эксперименты показали, что движение ресничек и жгутиков является результатом химически обусловленного "хождения" ответвлений динеина с одной
- 58. Белки-моторы, ассоциированные с микротрубочками, участвуют в формировании митотического веретена и осуществляют направленный транспорт вдоль микротрубочек интерфазной
- 59. Схема перемещения молекулы кинезина вдоль микротрубочки, состоящей из мономеров тубулина Буквами Т и D обозначены головки
- 60. Бактериальный жгутик - это сложный наномеханизм, в чьем создании принимают участие около 240 различных белков. Он
- 63. Скачать презентацию

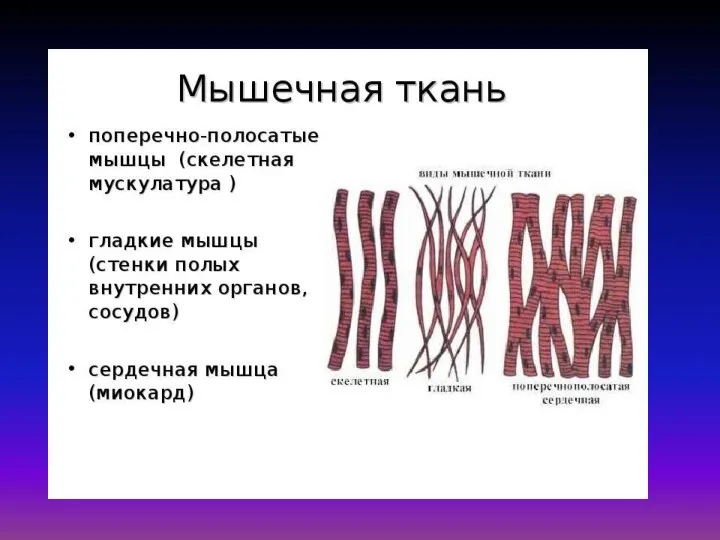
Механохимические процессы
мышечное сокращение;
движение клеток с помощью жгутиков и ресничек;
все движения в
Механохимические процессы
мышечное сокращение;
движение клеток с помощью жгутиков и ресничек;
все движения в
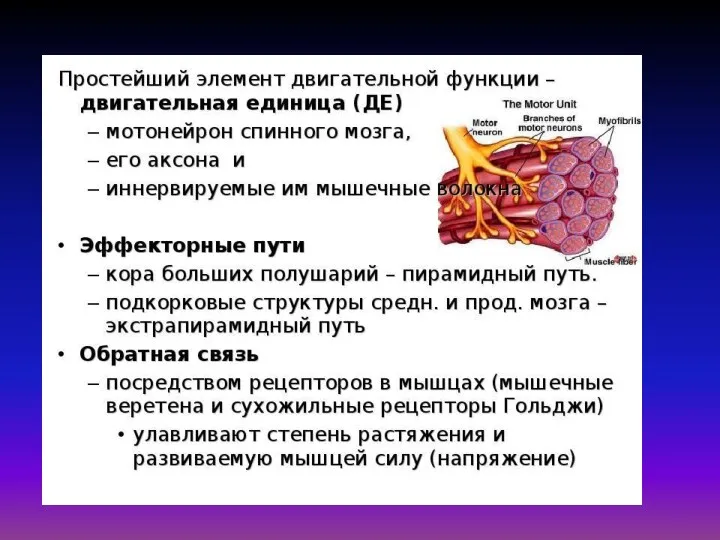
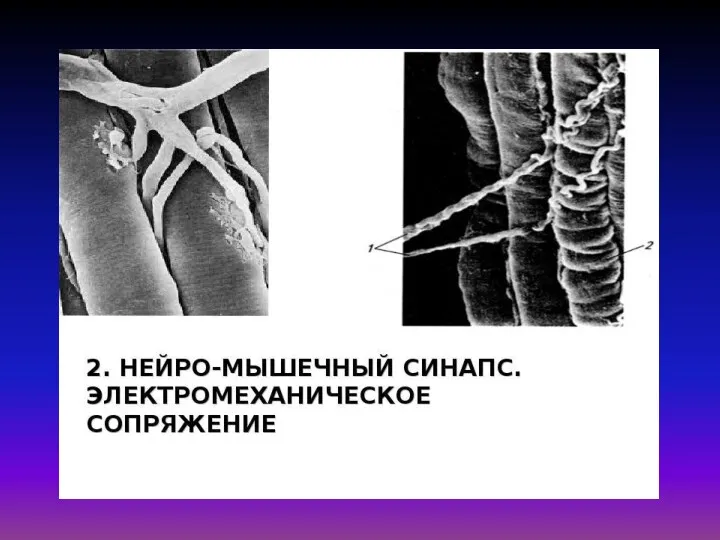


Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так
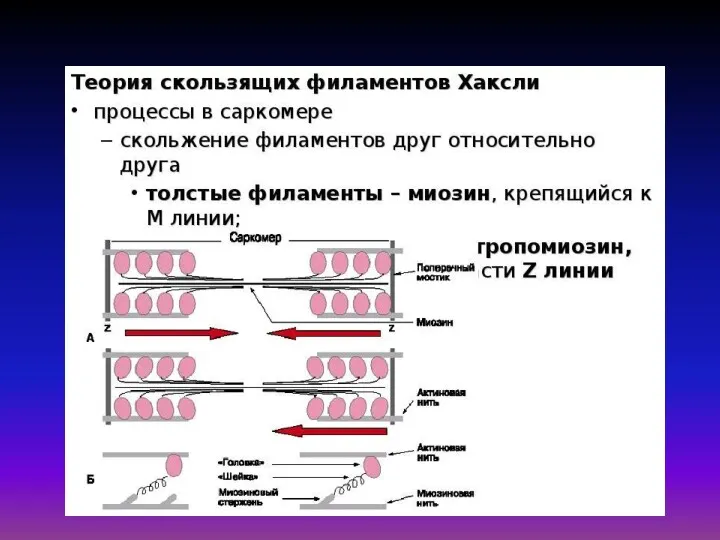
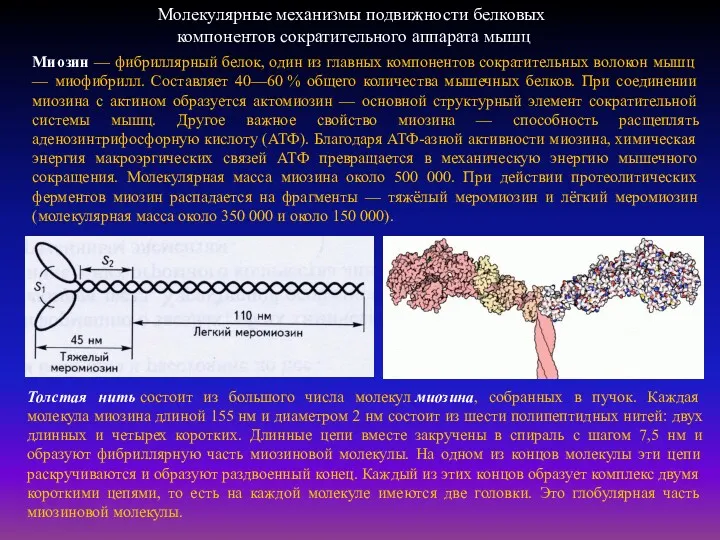
Толстая нить состоит из большого числа молекул миозина, собранных в пучок. Каждая молекула
Толстая нить состоит из большого числа молекул миозина, собранных в пучок. Каждая молекула

Благодаря огромному количеству работ посвященных мышечному сокращению [Бэгшоу С, 1985, Леднев
Благодаря огромному количеству работ посвященных мышечному сокращению [Бэгшоу С, 1985, Леднев
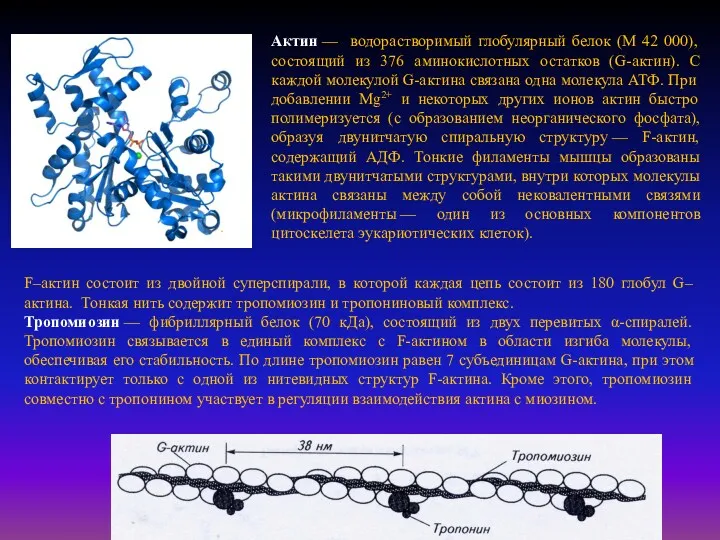
Актин — водорастворимый глобулярный белок (М 42 000), состоящий из 376 аминокислотных
Актин — водорастворимый глобулярный белок (М 42 000), состоящий из 376 аминокислотных
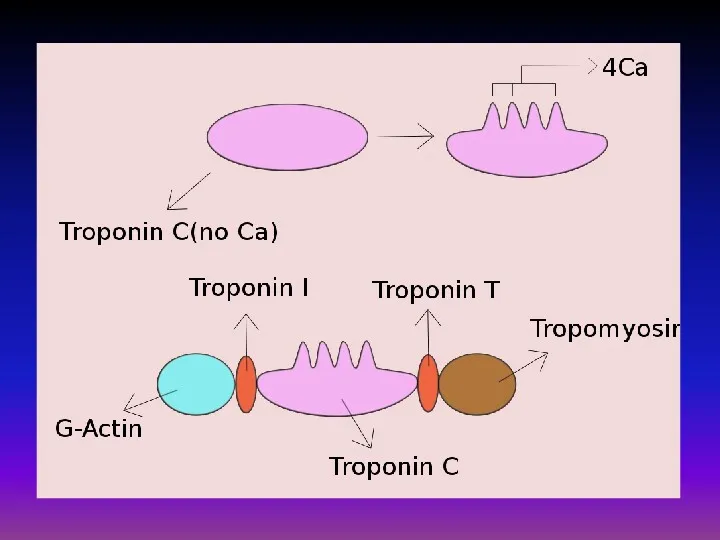
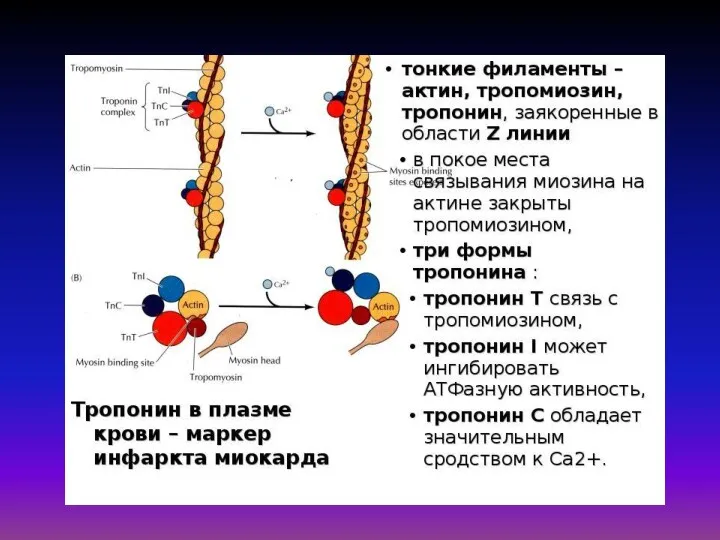

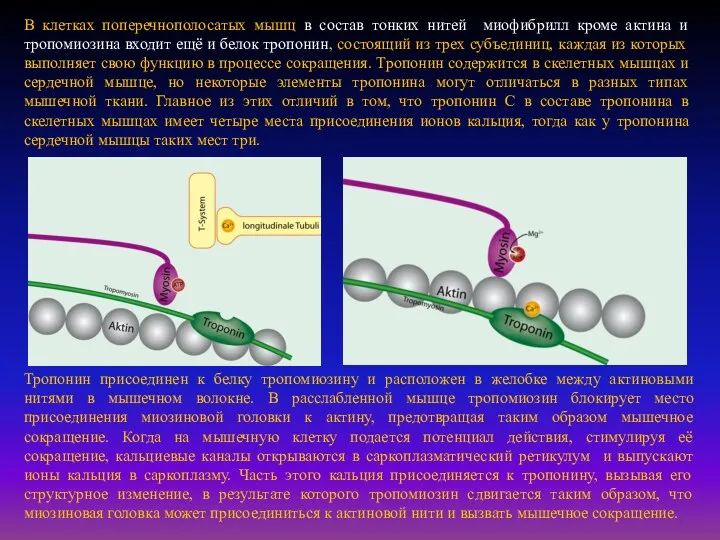
В клетках поперечнополосатых мышц в состав тонких нитей миофибрилл кроме актина
В клетках поперечнополосатых мышц в состав тонких нитей миофибрилл кроме актина
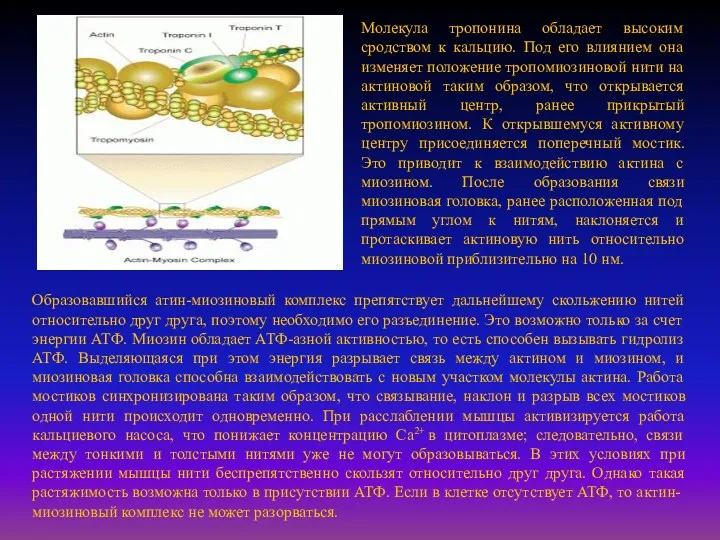
Молекула тропонина обладает высоким сродством к кальцию. Под его влиянием она
Молекула тропонина обладает высоким сродством к кальцию. Под его влиянием она

Структура и функционирование
поперечнополосатой мышцы позвоночных.
Диск I включает в себя
Структура и функционирование
поперечнополосатой мышцы позвоночных.
Диск I включает в себя
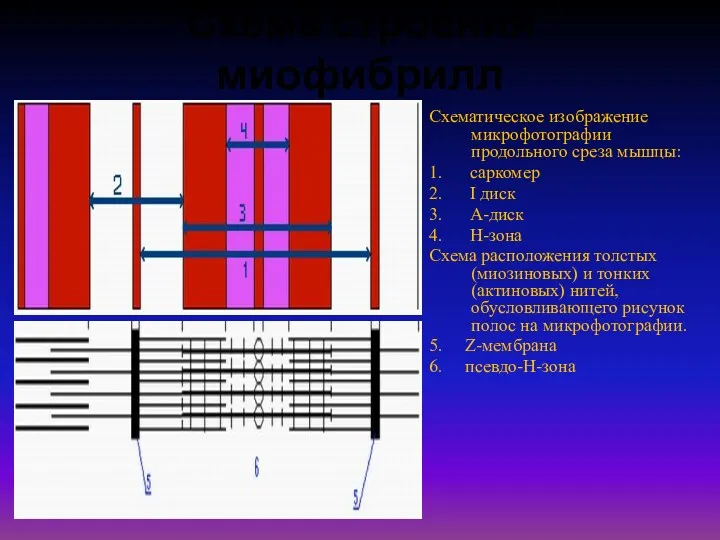
Схема строения миофибрилл
Схематическое изображение микрофотографии продольного среза мышцы:
1. саркомер
2. I
Схема строения миофибрилл
Схематическое изображение микрофотографии продольного среза мышцы:
1. саркомер
2. I
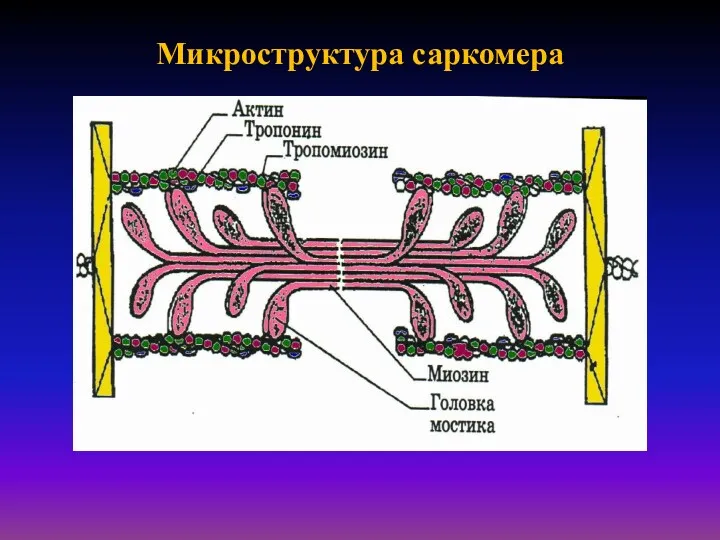
Микроструктура саркомера
Микроструктура саркомера

Модель мышечного сокращения Хаксли
1 – актиновый филамент, 2 – центр связывания,
Модель мышечного сокращения Хаксли
1 – актиновый филамент, 2 – центр связывания,
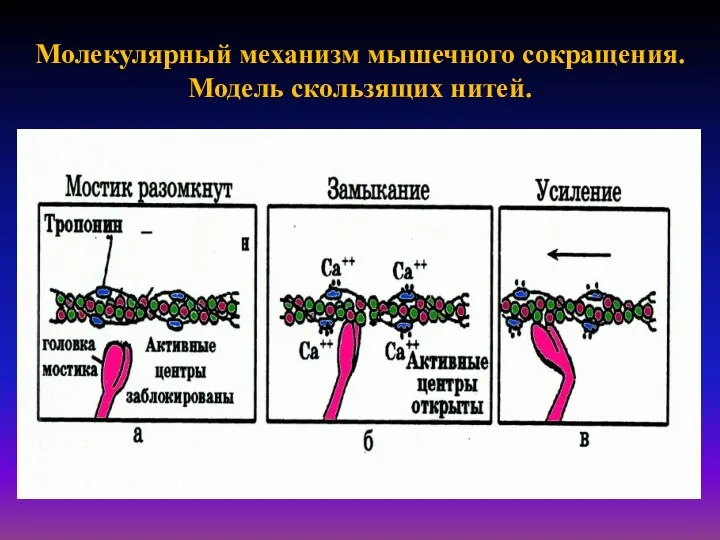
Молекулярный механизм мышечного сокращения.
Модель скользящих нитей.
Молекулярный механизм мышечного сокращения.
Модель скользящих нитей.

Основные положения модели скользящих нитей
1) Длина нитей актина и миозина в
Основные положения модели скользящих нитей
1) Длина нитей актина и миозина в
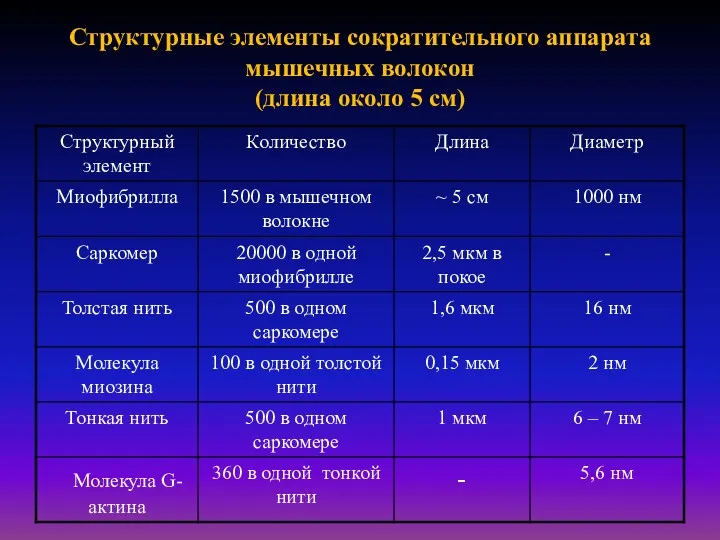
Структурные элементы сократительного аппарата мышечных волокон
(длина около 5 см)
Структурные элементы сократительного аппарата мышечных волокон
(длина около 5 см)
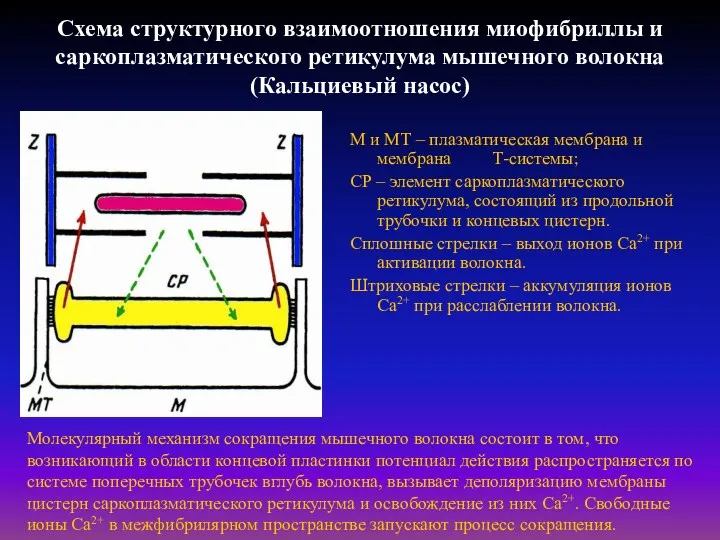
Схема структурного взаимоотношения миофибриллы и саркоплазматического ретикулума мышечного волокна (Кальциевый насос)
М
Схема структурного взаимоотношения миофибриллы и саркоплазматического ретикулума мышечного волокна (Кальциевый насос)
М
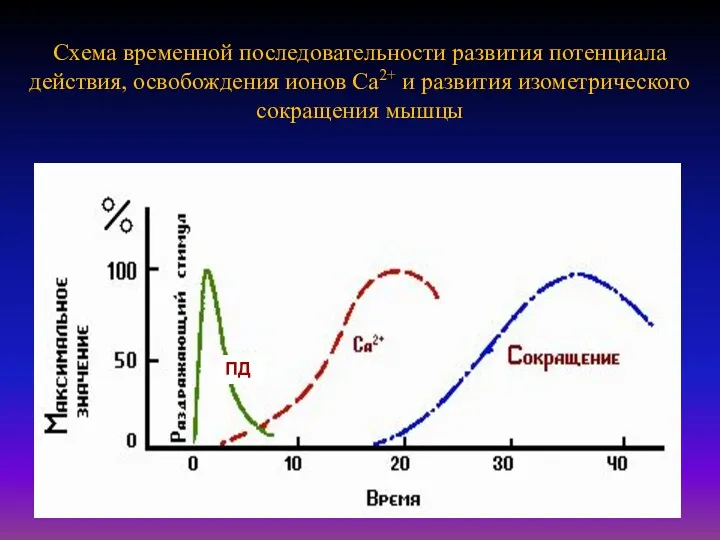
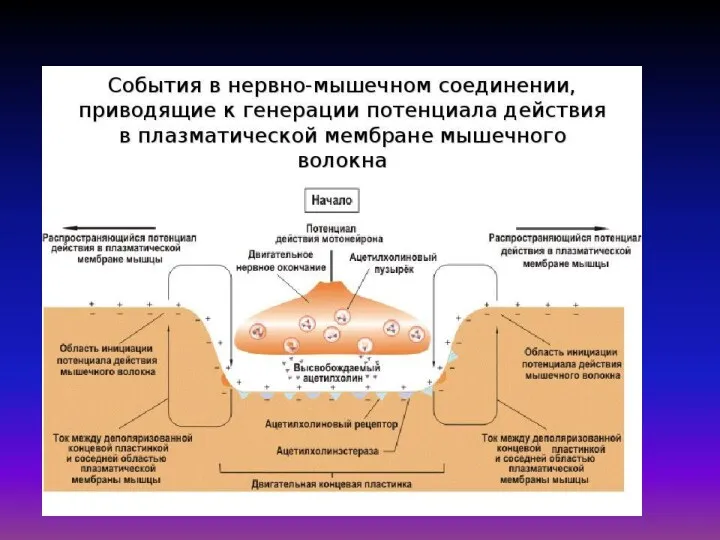
Схема временной последовательности развития потенциала действия, освобождения ионов Са2+ и развития
Схема временной последовательности развития потенциала действия, освобождения ионов Са2+ и развития

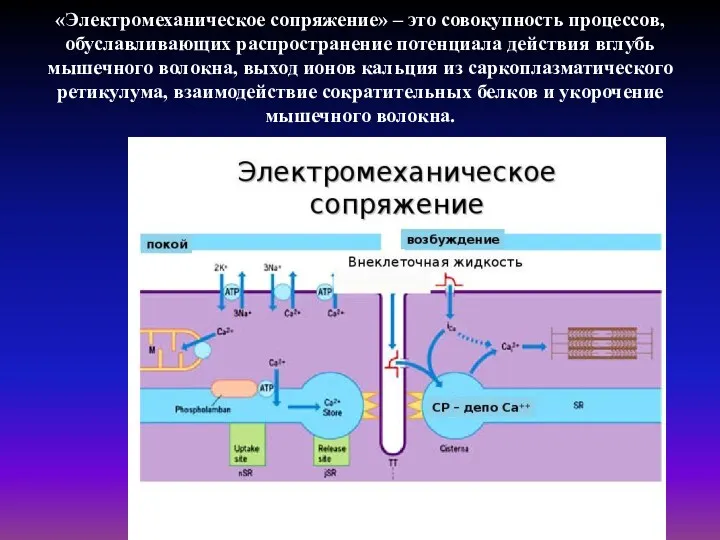
«Электромеханическое сопряжение» – это совокупность процессов, обуславливающих распространение потенциала действия вглубь
«Электромеханическое сопряжение» – это совокупность процессов, обуславливающих распространение потенциала действия вглубь
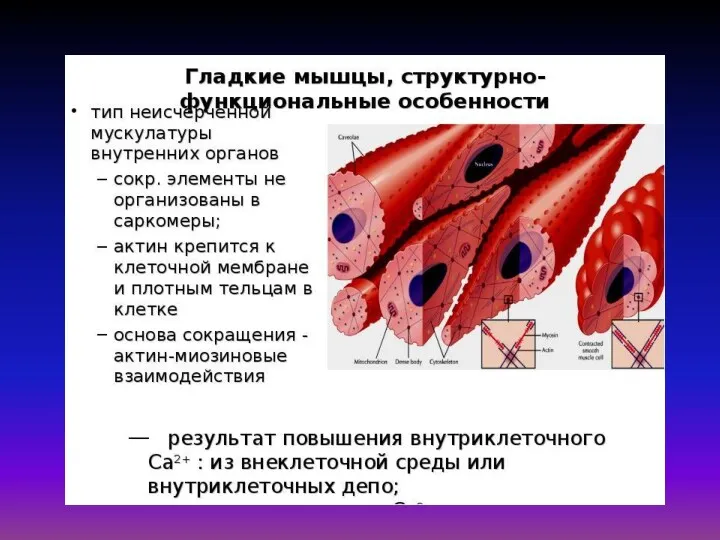
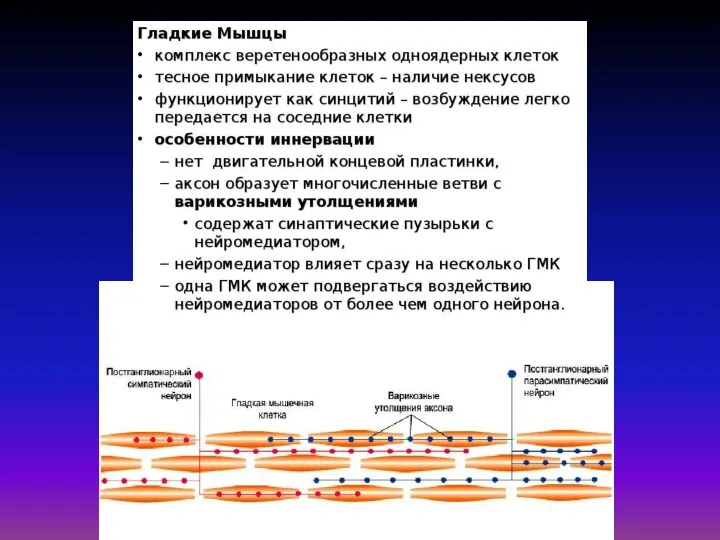



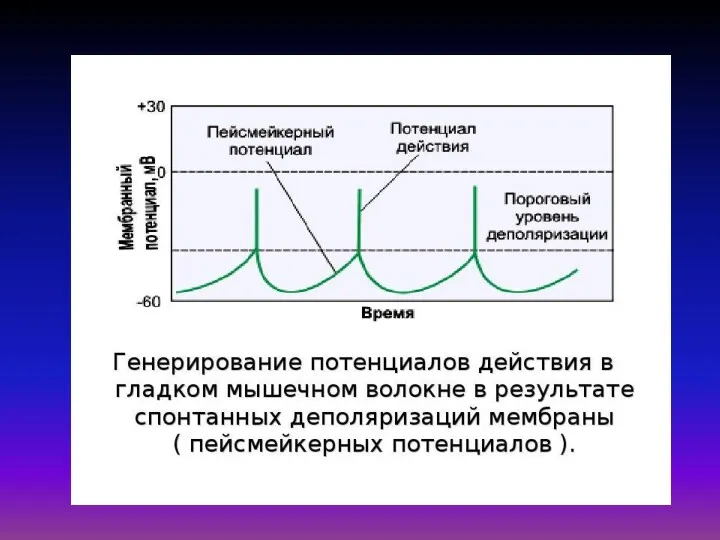


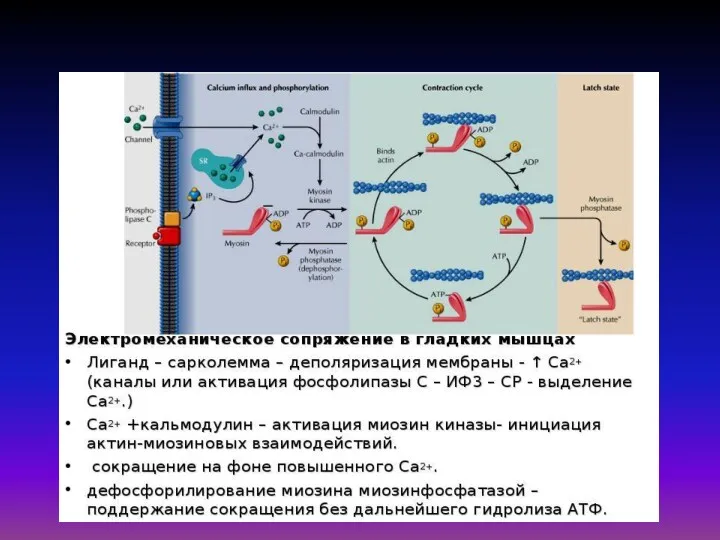



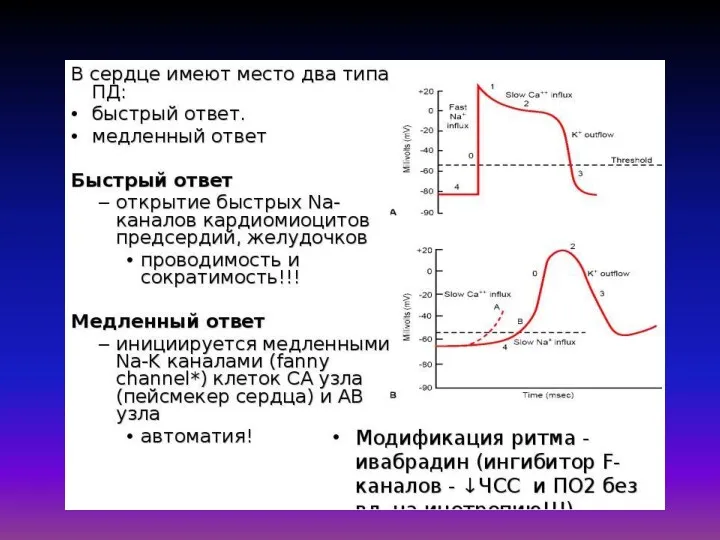
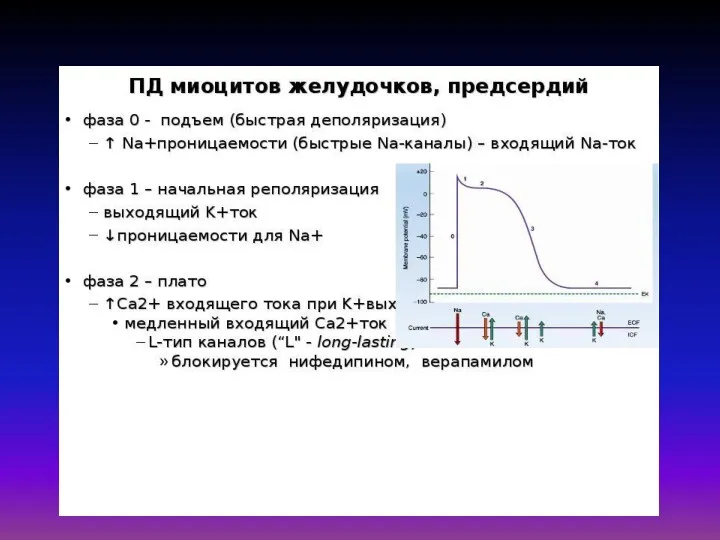




Математическая модель Дещеревского основана на модели «скользящих нитей» Хаксли
В кинетической теории
Математическая модель Дещеревского основана на модели «скользящих нитей» Хаксли
В кинетической теории

Выведено английским физиологом А. В. Хиллом в 1938, выражает изменение скорости
Выведено английским физиологом А. В. Хиллом в 1938, выражает изменение скорости

Микротрубочки создают направление упорядоченного перемещения веществ в клетке. Встречаются в свободном
Микротрубочки создают направление упорядоченного перемещения веществ в клетке. Встречаются в свободном

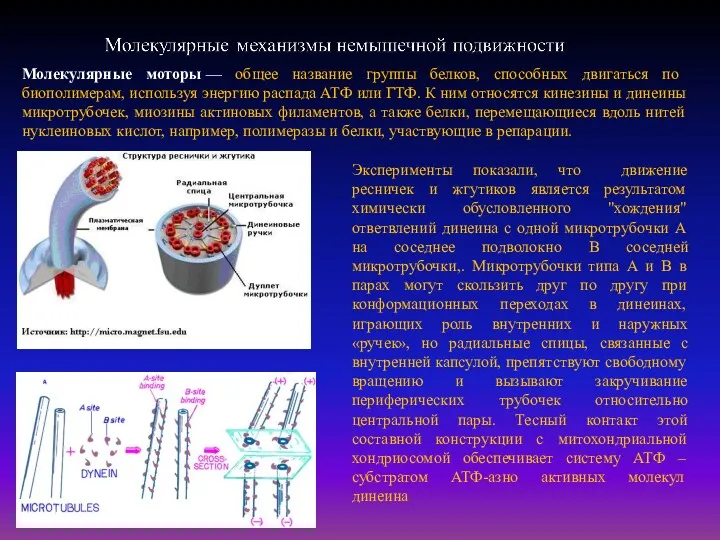
Эксперименты показали, что движение ресничек и жгутиков является результатом химически обусловленного
Эксперименты показали, что движение ресничек и жгутиков является результатом химически обусловленного
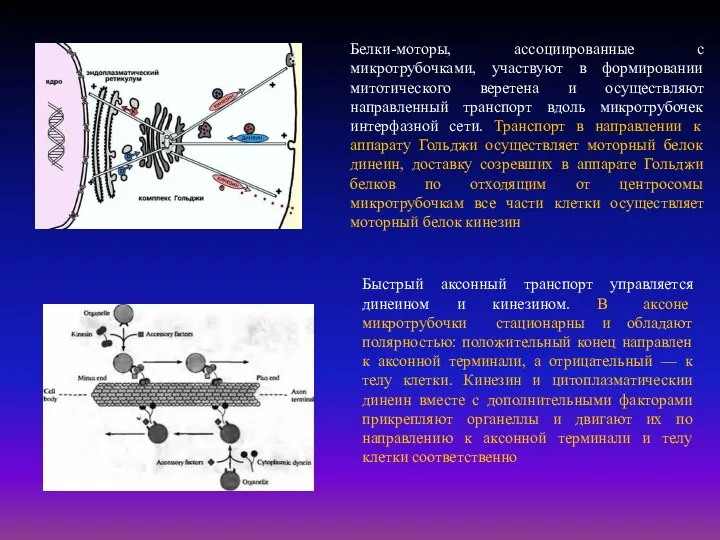
Белки-моторы, ассоциированные с микротрубочками, участвуют в формировании митотического веретена и осуществляют
Белки-моторы, ассоциированные с микротрубочками, участвуют в формировании митотического веретена и осуществляют
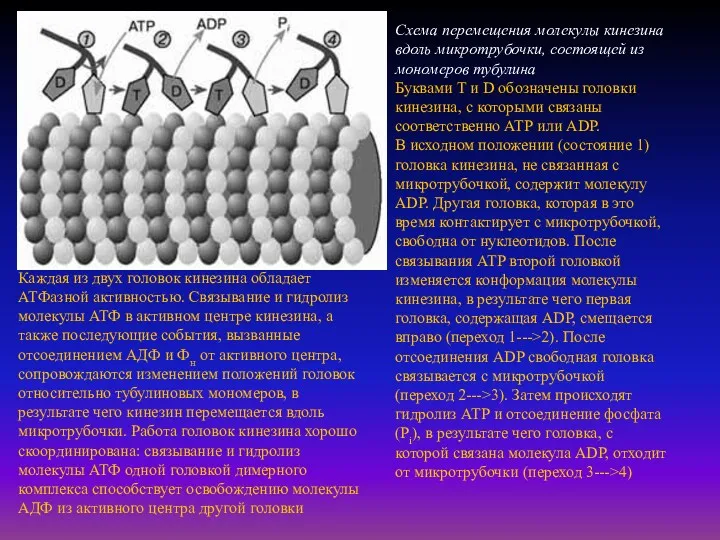
Схема перемещения молекулы кинезина вдоль микротрубочки, состоящей из мономеров тубулина
Буквами
Схема перемещения молекулы кинезина вдоль микротрубочки, состоящей из мономеров тубулина
Буквами
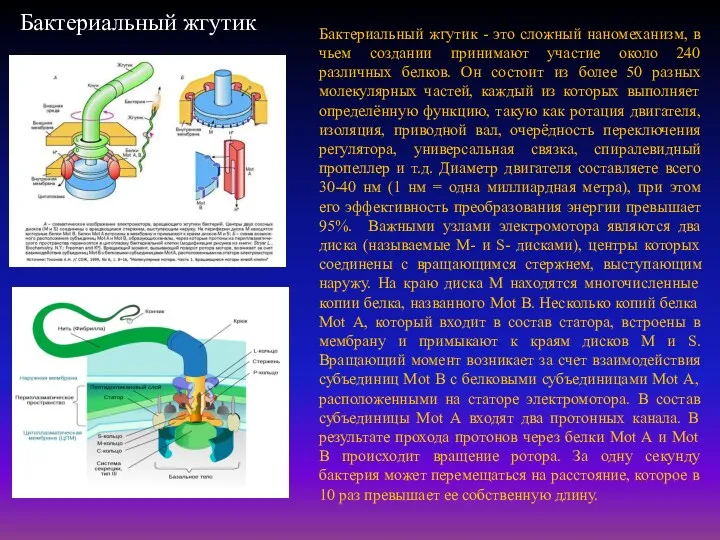
Бактериальный жгутик - это сложный наномеханизм, в чьем создании принимают участие
Бактериальный жгутик - это сложный наномеханизм, в чьем создании принимают участие

 Железы внутренней и смешанной секреции
Железы внутренней и смешанной секреции Введение в ветеринарную паразитологию. Биологические основы паразитизма. Лекция 1
Введение в ветеринарную паразитологию. Биологические основы паразитизма. Лекция 1 Животные. Суслики
Животные. Суслики Иван Владимирович Мичурин
Иван Владимирович Мичурин Разнообразие клеток. Клеточная теория. Эукариоты и прокариоты
Разнообразие клеток. Клеточная теория. Эукариоты и прокариоты Тварини. Характерні особливості тварин
Тварини. Характерні особливості тварин Исторические этапы развития микробиологии
Исторические этапы развития микробиологии В гостях у природы
В гостях у природы Популяционно-статистические методы
Популяционно-статистические методы Введение в миологию
Введение в миологию Особенности строения представителей надкласса Рыбы. 2
Особенности строения представителей надкласса Рыбы. 2 Лук от семи недуг. Польза лука
Лук от семи недуг. Польза лука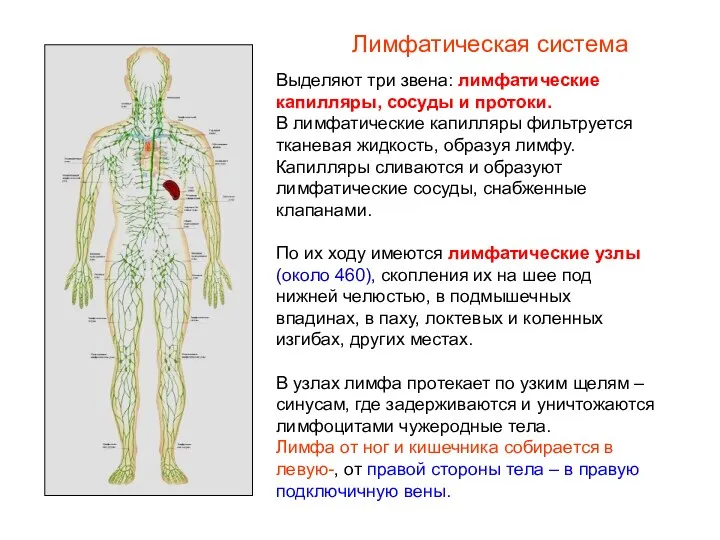 Лимфатическая система в организме человека
Лимфатическая система в организме человека Физиология возбудимых тканей
Физиология возбудимых тканей Энергетический обмен. (10 класс)
Энергетический обмен. (10 класс) Типи поживних речовин
Типи поживних речовин Надтип Amera. Тип Моллюски, или мягкотелые MOLLUSCA
Надтип Amera. Тип Моллюски, или мягкотелые MOLLUSCA Поздняя палеозойская эра
Поздняя палеозойская эра Исследовательский проект Пластиковая болезнь цивилизации
Исследовательский проект Пластиковая болезнь цивилизации Урок по кубановедению Кубань - степная дочь России (6 класс)
Урок по кубановедению Кубань - степная дочь России (6 класс) Теории возникновения жизни
Теории возникновения жизни Хомяки. Особенности содержания в домашних условиях
Хомяки. Особенности содержания в домашних условиях Жири, як компоненти їжі, їх біологічна роль
Жири, як компоненти їжі, їх біологічна роль Раздел Билатерально симметричные
Раздел Билатерально симметричные Биоэтика как мировоззрение. Принципы биоэтики, ее философские и правовые основы
Биоэтика как мировоззрение. Принципы биоэтики, ее философские и правовые основы Покрытосеменные (цветковые) растения
Покрытосеменные (цветковые) растения Микробиологическая характеристика биопленки, использование в биотехнологических процессах
Микробиологическая характеристика биопленки, использование в биотехнологических процессах Органические вещества клетки. Белки
Органические вещества клетки. Белки