- Физиология возбудимых тканей

Содержание
- 2. Возбудимые ткани. Основные понятия Возбудимые ткани – ткани способные в ответ на действие раздражителя переходить из
- 3. К невозбудимым тканям относятся эпителиальная и соединительная (собственно соединительная), ретикулярная, жировая, хрящевая, костная, кровь. Клетки этих
- 4. Для возбудимых тканей характерны такие свойства, как: Раздражимость – это общее свойство клеток отвечать на внешнее
- 5. Частным случаем раздражимости является возбудимость. Возбудимость - способность ткани при действии раздражителей переходить в активное состояние,
- 6. Раздражитель – это любое изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающее ответную реакцию.
- 7. Раздражители по физиологическому действию делятся:
- 8. СОСТОЯНИЕ КЛЕТКИ Клетка может находится в двух состояниях: Для нервной и мышечной ткани характерно возбуждение и
- 9. Биологические мембраны Биологические мембраны являются функционально-активными структурами клеток, ограничивающими цитоплазму и большинство внутриклеточных образований. Мембрана, ограничивает
- 10. Строение мембраны нервной клетки
- 11. Биологические мембраны выполняют следующие функции: Роль механического барьера. В клетке обеспечивается нормальное осмотическое и гидростатическое давление.
- 12. Метаболическая функция мембран. Эта функция определяется двумя факторами: объединением большого количества ферментов и ферментных систем с
- 13. Биоэлектрические явления в возбудимых тканях. Методы исследования При возбуждении в клетки происходят различные изменения: Структурные –
- 14. Животное электричество было открыто физиологом Л. Гальвани и физиком А. Вольта ( 1791- 1797). Эти ученые
- 15. Для регистрации биопотенциалов используется микроэлектродная техника. Микроэлектрод представляет собой стеклянную микропепетку (диаметр кончика 0,5 мкм), которая
- 16. Схема установки для регистрации биопотенциалов
- 17. А-В. Внутриклеточная регистрация мембранного потенциала. А. Клетка помещена в камеру, заполненную плазмой крови (или физ. раствором).
- 18. Происхождение потенциалов покоя Теория объясняющая происхождение мембранного потенциала покоя базируется на следующих основных положениях: Мембрана избирательно
- 19. На внутренней поверхности возникает избыток крупных молекул органических анионов. Благодаря электростатическим силам, вышедшие катионы калия не
- 20. Потенциал действия При действии раздражителей проницаемость мембраны изменяется, открываются мембранные каналы, пропускающие в клетку ионы натрия.
- 21. Фазы потенциала действия; временной ход потенциала действия в нерве
- 22. Восходящая часть пика – деполяризация, нисходящая – реполяризация. Имеется овершут – перезарядка мембраны (или перескок –
- 23. Если потенциал покоя присущ всем живым клеткам, то потенциал действия характерен в основном для нервных и
- 24. Возникновение ПД связано с изменением ионной проницаемости клеточной мембраны при ее возбуждении. В фазу деполяризации проницаемость
- 25. Ионное неравновесие Na+, K+ и др. ионов в системе «клетка – межклеточное вещество» восстанавливается за счет
- 26. Методы измерения возбудимости Возбудимость измеряется: А) по силе раздражения; Б) по длительности. Мерилом возбудимости является порог
- 27. РАЗДРАЖИТЕЛИ
- 28. Зависимость ответа ткани от силы раздражения
- 29. Зависимость между силой и длительностью порогового раздражения Пороговая сила любого раздражителя зависит от его длительности. Эта
- 30. Кривая «силы – времени» (по Лапику и др., 1926): 1 – реобаза, 2 – удвоенная реобаза;
- 31. Минимальная сила тока способная вызвать возбуждение была названа реобазой. Реобаза равна порогу. Это порог раздражения для
- 32. При раздражении целого нерва или мышцы используется метод хронаксометри который позволяет определить возбудимость нервно-мышечного препарата человека.
- 33. Изменение возбудимости ткани при прохождении волны возбуждения Возбудимость ткани при прохождении волны возбуждения меняется и не
- 34. Соотношение фаз возбудимости с фазами потенциала действия
- 35. Пику потенциала действия (мембрана полностью деполяризована) соответствует потеря возбудимости (возбудимость = 0). Эта фаза получила название
- 36. Быстрая реполяризация – восстановление потенциала покоя соответствует период относительной рефрактерности. В эту фазу возбудимость постепенно возрастает.
- 37. Отрицательному следовому потенциалу соответствует фаза повышенной возбудимости (супернормальной), то есть возбудимость выше нормы. В эту фазу
- 38. Значение рефрактерности: Обеспечивает осуществление реакции без помех (сокращение сердца). Защищает ткань от избыточного расхода энергии. Возбуждение
- 39. Физиологическая лабильность и ее измерение Лабильность - скорость протекания электрических и биохимических процессов, обеспечивающих функциональную деятельность
- 40. Лабильность нервного волокна составляет – 450 – 1000 имп/с; Мышечного волокна – 250 – 400 имп/с
- 41. РИТМ РАЗДРАЖЕНИЯ РИТМ РАЗДРАЖЕНИЯ Оптимальный Максимальный Пессимальный Частота ответов = частоте раздражения (длительно) Частота ответов соответствует
- 42. При определенных условиях процесс возбуждения переходит в свою противоположность. Таким образом, процесс возбуждения и торможения являются
- 44. Скачать презентацию

Возбудимые ткани. Основные понятия
Возбудимые ткани – ткани способные в ответ на
Возбудимые ткани. Основные понятия
Возбудимые ткани – ткани способные в ответ на

К невозбудимым тканям относятся эпителиальная и соединительная (собственно соединительная), ретикулярная, жировая,
К невозбудимым тканям относятся эпителиальная и соединительная (собственно соединительная), ретикулярная, жировая,

Для возбудимых тканей характерны такие свойства, как:
Раздражимость – это общее
Для возбудимых тканей характерны такие свойства, как:
Раздражимость – это общее

Частным случаем раздражимости является возбудимость.
Возбудимость - способность ткани при действии раздражителей
Частным случаем раздражимости является возбудимость.
Возбудимость - способность ткани при действии раздражителей

Раздражитель – это любое изменение внешней или внутренней среды организма, воспринимаемое
Раздражитель – это любое изменение внешней или внутренней среды организма, воспринимаемое
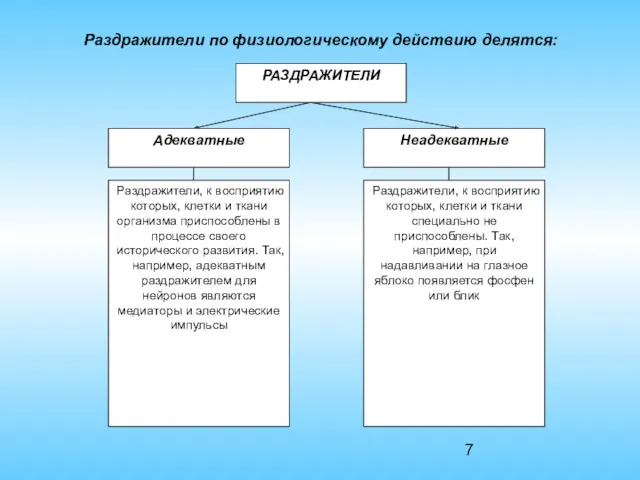
Раздражители по физиологическому действию делятся:
Раздражители по физиологическому действию делятся:
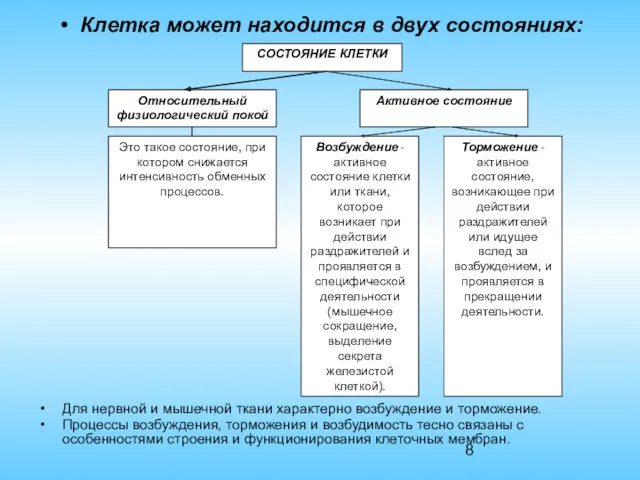
СОСТОЯНИЕ КЛЕТКИ
Клетка может находится в двух состояниях:
Для нервной и мышечной ткани
СОСТОЯНИЕ КЛЕТКИ
Клетка может находится в двух состояниях:
Для нервной и мышечной ткани

Биологические мембраны
Биологические мембраны являются функционально-активными структурами клеток, ограничивающими цитоплазму и большинство
Биологические мембраны
Биологические мембраны являются функционально-активными структурами клеток, ограничивающими цитоплазму и большинство
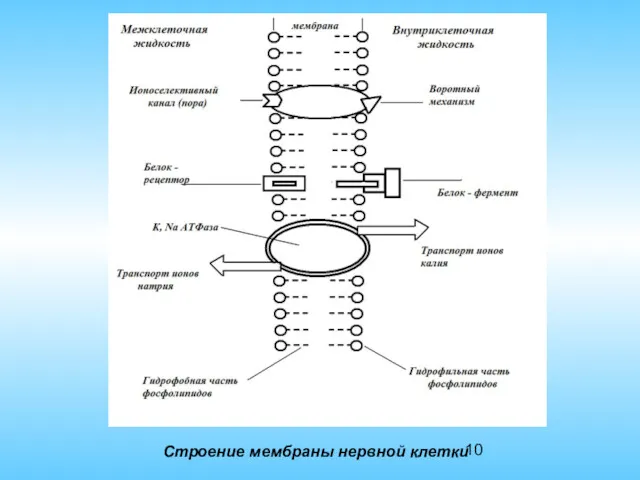
Строение мембраны нервной клетки
Строение мембраны нервной клетки

Биологические мембраны выполняют следующие функции:
Роль механического барьера. В клетке обеспечивается нормальное
Биологические мембраны выполняют следующие функции:
Роль механического барьера. В клетке обеспечивается нормальное

Метаболическая функция мембран. Эта функция определяется двумя факторами: объединением большого количества
Метаболическая функция мембран. Эта функция определяется двумя факторами: объединением большого количества

Биоэлектрические явления в возбудимых тканях. Методы исследования
При возбуждении в клетки происходят
Биоэлектрические явления в возбудимых тканях. Методы исследования
При возбуждении в клетки происходят

Животное электричество было открыто физиологом Л. Гальвани и физиком А. Вольта
Животное электричество было открыто физиологом Л. Гальвани и физиком А. Вольта

Для регистрации биопотенциалов используется микроэлектродная техника. Микроэлектрод представляет собой стеклянную микропепетку
Для регистрации биопотенциалов используется микроэлектродная техника. Микроэлектрод представляет собой стеклянную микропепетку
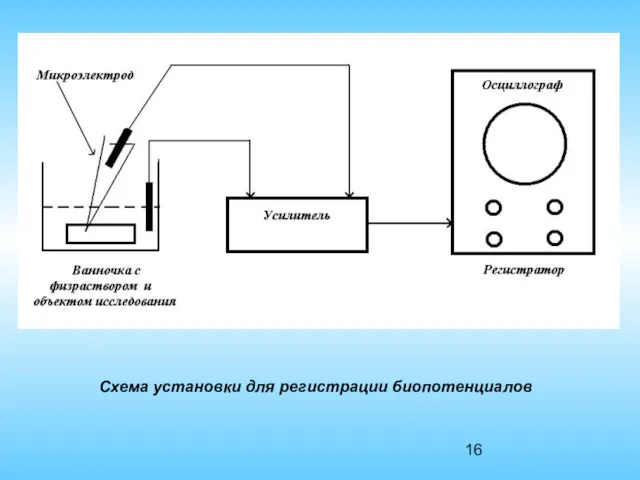
Схема установки для регистрации биопотенциалов
Схема установки для регистрации биопотенциалов
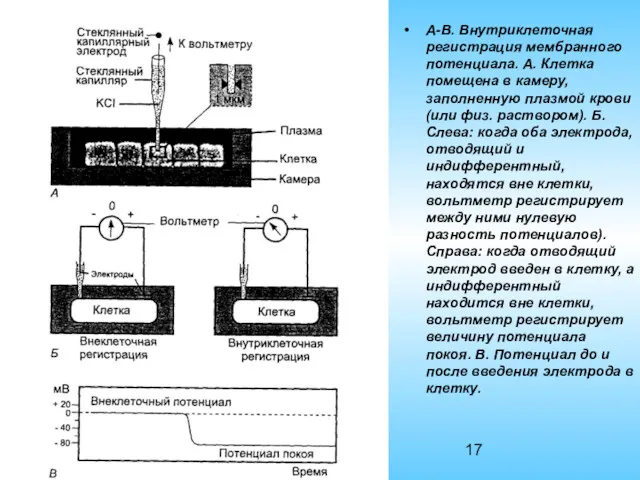
А-В. Внутриклеточная регистрация мембранного потенциала. А. Клетка помещена в камеру, заполненную
А-В. Внутриклеточная регистрация мембранного потенциала. А. Клетка помещена в камеру, заполненную

Происхождение потенциалов покоя
Теория объясняющая происхождение мембранного потенциала покоя базируется на следующих
Происхождение потенциалов покоя
Теория объясняющая происхождение мембранного потенциала покоя базируется на следующих

На внутренней поверхности возникает избыток крупных молекул органических анионов. Благодаря электростатическим
На внутренней поверхности возникает избыток крупных молекул органических анионов. Благодаря электростатическим

Потенциал действия
При действии раздражителей проницаемость мембраны изменяется, открываются мембранные каналы, пропускающие
Потенциал действия
При действии раздражителей проницаемость мембраны изменяется, открываются мембранные каналы, пропускающие
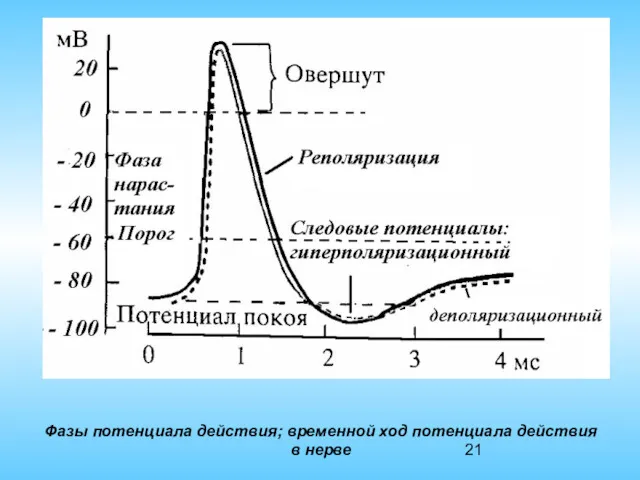
Фазы потенциала действия; временной ход потенциала действия в нерве
Фазы потенциала действия; временной ход потенциала действия в нерве

Восходящая часть пика – деполяризация, нисходящая – реполяризация. Имеется овершут –
Восходящая часть пика – деполяризация, нисходящая – реполяризация. Имеется овершут –

Если потенциал покоя присущ всем живым клеткам, то потенциал действия характерен
Если потенциал покоя присущ всем живым клеткам, то потенциал действия характерен

Возникновение ПД связано с изменением ионной проницаемости клеточной мембраны при ее
Возникновение ПД связано с изменением ионной проницаемости клеточной мембраны при ее

Ионное неравновесие Na+, K+ и др. ионов в системе «клетка –
Ионное неравновесие Na+, K+ и др. ионов в системе «клетка –

Методы измерения возбудимости
Возбудимость измеряется:
А) по силе раздражения;
Б) по длительности.
Мерилом
Методы измерения возбудимости
Возбудимость измеряется:
А) по силе раздражения;
Б) по длительности.
Мерилом
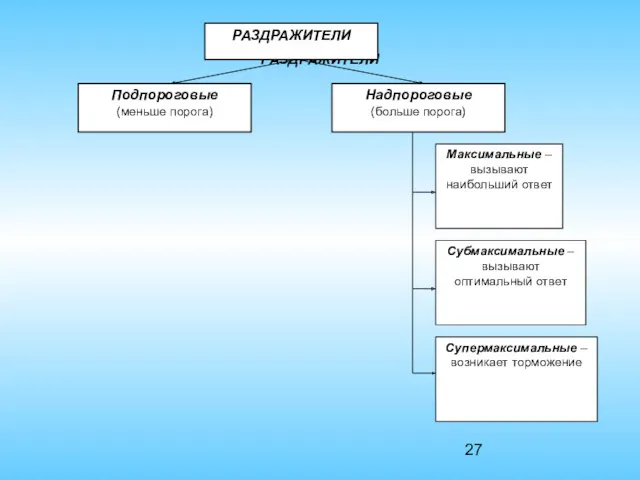
РАЗДРАЖИТЕЛИ
РАЗДРАЖИТЕЛИ
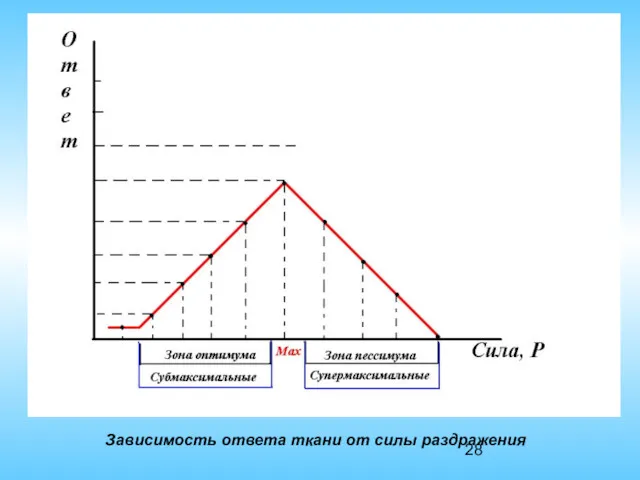
Зависимость ответа ткани от силы раздражения
Зависимость ответа ткани от силы раздражения

Зависимость между силой и длительностью порогового раздражения
Пороговая сила любого раздражителя зависит
Зависимость между силой и длительностью порогового раздражения
Пороговая сила любого раздражителя зависит
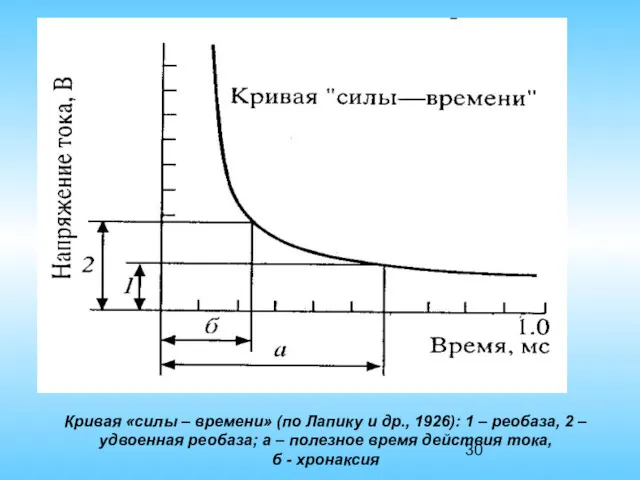
Кривая «силы – времени» (по Лапику и др., 1926): 1 –
Кривая «силы – времени» (по Лапику и др., 1926): 1 –

Минимальная сила тока способная вызвать возбуждение была названа реобазой. Реобаза равна
Минимальная сила тока способная вызвать возбуждение была названа реобазой. Реобаза равна

При раздражении целого нерва или мышцы используется метод хронаксометри который позволяет
При раздражении целого нерва или мышцы используется метод хронаксометри который позволяет

Изменение возбудимости ткани при прохождении волны возбуждения
Возбудимость ткани при прохождении волны
Изменение возбудимости ткани при прохождении волны возбуждения
Возбудимость ткани при прохождении волны
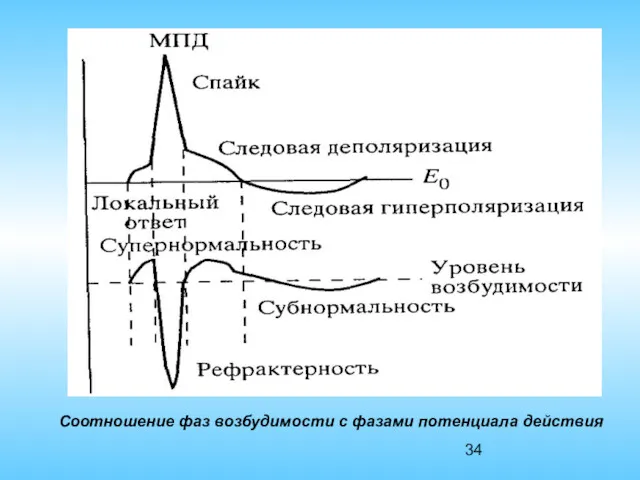
Соотношение фаз возбудимости с фазами потенциала действия
Соотношение фаз возбудимости с фазами потенциала действия

Пику потенциала действия (мембрана полностью деполяризована) соответствует потеря возбудимости (возбудимость =
Пику потенциала действия (мембрана полностью деполяризована) соответствует потеря возбудимости (возбудимость =

Быстрая реполяризация – восстановление потенциала покоя соответствует период относительной рефрактерности.
В эту
Быстрая реполяризация – восстановление потенциала покоя соответствует период относительной рефрактерности.
В эту

Отрицательному следовому потенциалу соответствует фаза повышенной возбудимости (супернормальной), то есть возбудимость
Отрицательному следовому потенциалу соответствует фаза повышенной возбудимости (супернормальной), то есть возбудимость

Значение рефрактерности:
Обеспечивает осуществление реакции без помех (сокращение сердца).
Защищает ткань от избыточного
Значение рефрактерности:
Обеспечивает осуществление реакции без помех (сокращение сердца).
Защищает ткань от избыточного

Физиологическая лабильность
и ее измерение
Лабильность - скорость протекания электрических и биохимических
Физиологическая лабильность
и ее измерение
Лабильность - скорость протекания электрических и биохимических

Лабильность нервного волокна составляет – 450 – 1000 имп/с;
Мышечного волокна –
Лабильность нервного волокна составляет – 450 – 1000 имп/с;
Мышечного волокна –
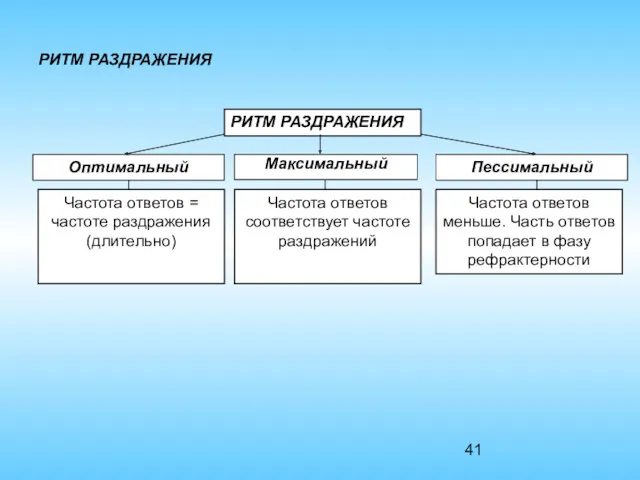
РИТМ РАЗДРАЖЕНИЯ
РИТМ РАЗДРАЖЕНИЯ
Оптимальный
Максимальный
Пессимальный
Частота ответов = частоте раздражения (длительно)
Частота ответов соответствует частоте
РИТМ РАЗДРАЖЕНИЯ
РИТМ РАЗДРАЖЕНИЯ
Оптимальный
Максимальный
Пессимальный
Частота ответов = частоте раздражения (длительно)
Частота ответов соответствует частоте

При определенных условиях процесс возбуждения переходит в свою противоположность.
Таким образом,
При определенных условиях процесс возбуждения переходит в свою противоположность.
Таким образом,
 Красная Книга Ростовской области
Красная Книга Ростовской области Механизмы биоэлектрогенеза
Механизмы биоэлектрогенеза Структура спинного мозга
Структура спинного мозга Обеспечение клеток энергией
Обеспечение клеток энергией Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9)
Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9) Физиология сердца. (Лекция 5.1)
Физиология сердца. (Лекция 5.1) Отряд Киты
Отряд Киты Межвидовые отношения организмов
Межвидовые отношения организмов Эволюция ОДС (опорно-двигательной системы)
Эволюция ОДС (опорно-двигательной системы) Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян
Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян Предмет и структура ихтиологической науки
Предмет и структура ихтиологической науки Цветы в легендах, поэзии, музыке
Цветы в легендах, поэзии, музыке ВНД человека. Вторая сигнальная система
ВНД человека. Вторая сигнальная система Морфология. Общие понятия
Морфология. Общие понятия Конструкции, обогрев и эксплуатация сооружений защищенного грунта
Конструкции, обогрев и эксплуатация сооружений защищенного грунта Жизнедеятельность клетки
Жизнедеятельность клетки Natural selection of human population. Darwin’s theory of evolution by natural selection
Natural selection of human population. Darwin’s theory of evolution by natural selection Антропогенез. Расы. Расизм. Часть 4
Антропогенез. Расы. Расизм. Часть 4 Бактерии
Бактерии Потребности и интересы. Уровень развития способностей человека
Потребности и интересы. Уровень развития способностей человека Динозавры
Динозавры Пищеварительная система
Пищеварительная система Физиология спорта. Физиологические основы классификации физических упражнений
Физиология спорта. Физиологические основы классификации физических упражнений Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования
Презентация к выступлению на научной конференции Совершенствование естественнонаучного школьного образования Клубеньковые долгоносики. Гороховая зерновка
Клубеньковые долгоносики. Гороховая зерновка Типы кожи лица. Уход за сухой кожей
Типы кожи лица. Уход за сухой кожей Как животные пустынь получают воду
Как животные пустынь получают воду