- Биологические основы клеточной сигнализации

Содержание
- 2. На лекции рассматриваются следующие вопросы: понятие о клеточной сигнализации и ее типы; внутриклеточные рецепторы и их
- 3. Клеточная сигнализация – это процесс получения и обработки клеткой информации, поступающей из окружающей среды в виде
- 4. Клеточная сигнализация осуществляется сигнальными системами, материальную основу которых на молекулярном уровне составляют рецепторы, элементы сигнал передающих
- 5. 1 2 3 Ответ
- 6. Общий принцип функционирования сигнальных систем состоит в том, что внешний сигнал, взаимодействуя с рецептором клеток-мишеней, вызывает
- 7. Клеточная сигнализация играет большую роль в формировании адаптивных реакций про- и эукариотических организмов в ответ на
- 8. Клеточная сигнализация у прокариотических клеток
- 9. Типы клеточной сигнализации с использованием химических сигнальных молекул: - паракринная сигнализация характеризуется выделением клеткой химических веществ,
- 10. - синаптическая сигнализация, встречается лишь у животных, имеющих нервную систему. Характеризуется секрецией нейронами в синаптическую щель
- 11. Типы клеточной сигнализации А-паракринная сигнализация; Б-синаптическая сигнализация; В-эндокринная сигнализация 1- секретируемые сигнальные молекулы нейромедиатор 1 1
- 12. Характеристика химических сигнальных молекул
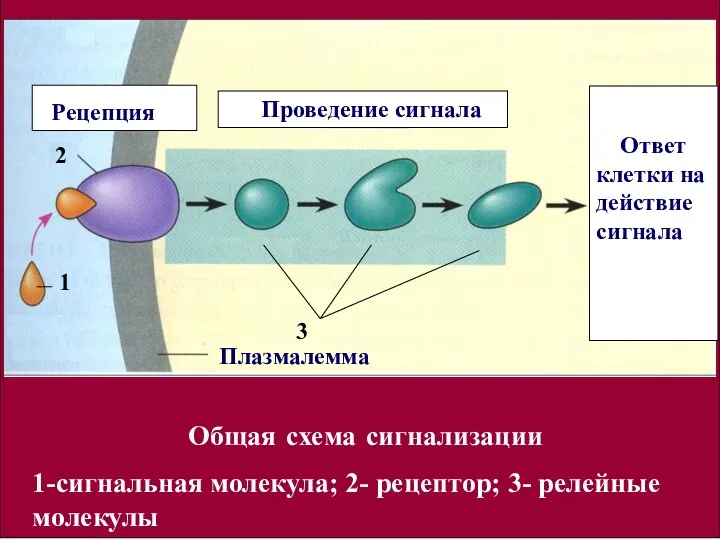
- 13. Выделяют три стадии клеточной сигнализации: рецепция; проведение сигнала к эффекторным молекулам; ответ клетки на действие сигнальных
- 15. Клетки-мишени распознают сигнальные молекулы с помощью рецепторов. Механизм распознавания сигналов осуществляется на основе конформационного соответствия молекул
- 16. Рецепторы по локализации их в клетке подразделяют на две группы: • внутриклеточные рецепторы; • рецепторы клеточной
- 17. Внутриклеточные рецепторы расположены в цитозоле клетки или ядре. Химическими сигналами для них служат молекулы, которые переносятся
- 18. Внутриклеточные рецепторы не только распознают и связывают сигнальные молекулы, но и сами участвуют в передаче сигнала,
- 19. Передача сигнала внутриклеточным рецептором
- 20. Рецепторы, расположенные в плазматической мембране, или рецепторы клеточной поверхности, относятся к интегральным мембранным белкам. Они взаимодействуют
- 21. Выделяют три главных типа рецепторов клеточной поверхности: рецепторы ионных каналов; рецепторы, сопряженные с G- белками; каталитические
- 22. Рецепторы ионных каналов представляют собой регулируемые лигандами ионные каналы. Сигнальные молекулы, связываясь с белком рецептором, изменяют
- 24. Передача сигнала никотиновым ацетилхолиновым рецептором 1-2-3-4 – последовательность открытия ионных каналов.
- 26. Механизм синаптической передачи сигналов между электрически возбудимыми клетками с помощью рецепторов ионных каналов заключается в преобразовании
- 27. Рецепторы, сопряженные с G-белками, функционируют в тесной кооперации с регуляторными G-белками. Они встречаются у всех представителей
- 28. Структура рецептора, сопряженного с G-белками 1- сигнал-связывающий участок; 2-участок , взаимодействую- щий с G-белком
- 29. Рецепторы, сопряженные с G-белками, передают сигнал путем активации или инактивации определенных ферментов или ионных каналов, связанных
- 30. G-белки имеют центры связывания гуаниловых нуклеотидов: гуанозиндифосфата (ГДФ) и гуанозинтрифосфата (ГТФ). G-белки обладают собственной фосфатазной активностью,
- 32. Принцип работы рецептора, сопряженного с G-белком - связывание сигнальной молекулы с рецептором; - активация рецептора; -
- 33. Каталитические рецепторы обладают собственной ферментативной активностью. Сигнальные системы с участием каталитических рецепторов играют большую роль в
- 34. Тирозинкиназный рецептор 1 - плазмалемма; 2 - сигнал-связывающий участок; 3 - мембранный домен; 4,5 - тирозин-киназные
- 35. Принцип работы тирозинкиназного рецептора - связывание сигнальной молекулы с рецептором; агрегация двух полипептидных цепей рецептора с
- 36. Передача сигнала от рецепторов клеточной поверхности к эффекторным молекулам включает цепь молекулярных взаимодействий релейных белковых молекул,
- 37. Многие релейные белки, участвующие в проведении сигнала от рецепторов клеточной поверхности к эффекторным белкам представляют собой
- 38. Наряду с протеинкиназами большую роль в передаче сигналов играют ферменты протеинфосфатазы, которые быстро удаляют фосфатные группы
- 39. сигнальная молекула 1 1а 2а 3 3а 2 Клеточный ответ 4 5 рецептор Каскад реакций фофорилирования
- 40. Система, обеспечивающая фосфорилирование и дефосфорилирование релейных белков в клетке, действует как «молекулярный выключатель», который активирует или
- 41. 1-сигнальная молекула (фактор роста);2-активный тирозинкиназнывй рецептор; 3-фосфорилированые остатки тирозина цитоплазматических доменов; 5- Grb-белок; 6- SOS-белок; 7-Ras
- 42. Во многих сигнальных путях в проведении сигнала, наряду с белками, участвуют малые небелковые молекулы и ионы,
- 43. Превращения циклического аденозинмонофосфата
- 44. Схема проведения сигнала при взаимодействии адреналина с β- адренергическим рецептором клетки печени.
- 45. Общая схема проведения сигнала в клетке печени. при взаимодействии адреналина с рецептором, сопряженным с G- белком:
- 46. Действие адреналина сопровождается усилением распада гликогена и прекращением его синтеза , что способствует максимальной мобилизации энергетических
- 47. Использование в сигнальных системах ионов Са2+ в качестве вторичных посредников определяется двумя особенностями указанных ионов :
- 48. Схема транспорта ионов Са++ из цитозоля клетки в окружающуюсреду эпс
- 49. В механизме проведения сигнала с участием ионов Са++, участвуют два других вторичных посредника: инозитолтрифосфат и диацилглицерол.
- 51. Инозитол трифосфатный путь проведения сигнала 1- рецептор; 2 - G-белок; 3 - фосфолипаза С; 4- фосфатидилинозитол-4,5
- 52. Общая схема проведения сигнала с участием ионов кальция: - связывание сигнальной молекулы с рецептором; - активация
- 53. Диацилглицерол активирует фермент протеинкиназу С, связанный с цитоплазматической мембраной клетки. Протеинкиназа С участвует во многих процессах,
- 54. Схема проведения сигнала при связывании АДФ с рецепторами тромбоцитов 1-АДФ; 2- рецептор; 3,4 G - белок;
- 55. Наличие нескольких релейных молекул, вовлеченных в передачу сигнала, обеспечивает : • усиление сигнала; • возможность взаимодействия
- 56. Схема усиления сигнала при связывании адреналина с β- адренергическим рецептором клетки печени.
- 57. Ответ 1 Ответ 2 Схема взаимодействия сигналов, идущих от разных рецепторов Схема «ветвления» сигнальных путей, идущих
- 58. Ответные реакции клеток на действие сигнальных молекул отличаются высокой специфичностью. Это означает, что одни и те
- 59. Специфичность ответа клетки на действие сигнальных молекул объясняется: способностью одних и тех же сигнальных молекул связываться
- 61. Скачать презентацию

На лекции рассматриваются
следующие вопросы:
понятие о клеточной сигнализации и ее типы;
На лекции рассматриваются
следующие вопросы:
понятие о клеточной сигнализации и ее типы;

Клеточная сигнализация – это процесс получения и обработки клеткой информации, поступающей
Клеточная сигнализация – это процесс получения и обработки клеткой информации, поступающей

Клеточная сигнализация осуществляется сигнальными системами, материальную основу которых на молекулярном
Клеточная сигнализация осуществляется сигнальными системами, материальную основу которых на молекулярном
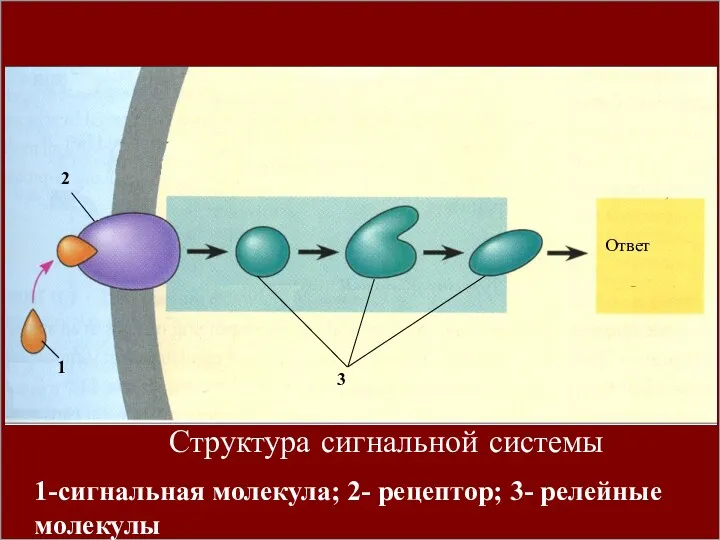
1
2
3
Ответ
1
2
3
Ответ

Общий принцип функционирования сигнальных систем состоит в том, что внешний
Общий принцип функционирования сигнальных систем состоит в том, что внешний

Клеточная сигнализация играет большую роль в формировании адаптивных реакций про-
Клеточная сигнализация играет большую роль в формировании адаптивных реакций про-
Клеточная сигнализация у прокариотических клеток
Клеточная сигнализация у прокариотических клеток

Типы клеточной сигнализации с использованием химических сигнальных молекул:
- паракринная
Типы клеточной сигнализации с использованием химических сигнальных молекул:
- паракринная

- синаптическая сигнализация, встречается лишь у животных, имеющих нервную систему.
- синаптическая сигнализация, встречается лишь у животных, имеющих нервную систему.
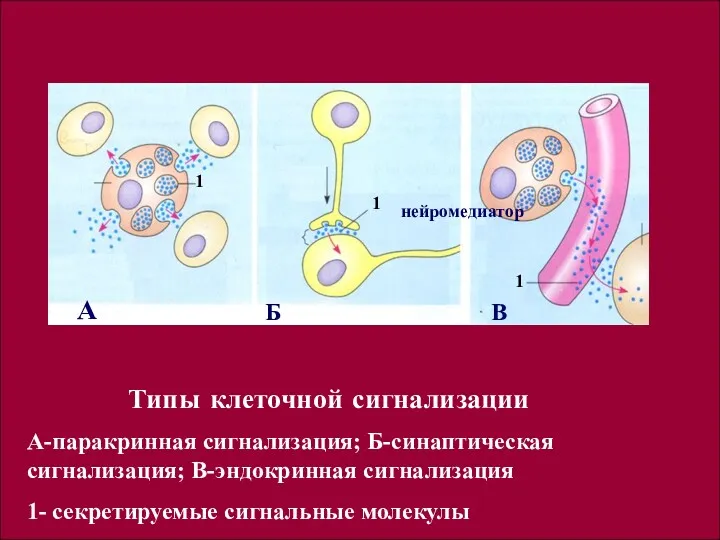
Типы клеточной сигнализации
А-паракринная сигнализация; Б-синаптическая сигнализация; В-эндокринная сигнализация
1- секретируемые сигнальные
Типы клеточной сигнализации
А-паракринная сигнализация; Б-синаптическая сигнализация; В-эндокринная сигнализация
1- секретируемые сигнальные

Характеристика химических сигнальных молекул
Характеристика химических сигнальных молекул

Выделяют
три стадии клеточной сигнализации:
рецепция;
проведение сигнала к эффекторным
три стадии клеточной сигнализации:
рецепция;
проведение сигнала к эффекторным

Клетки-мишени распознают сигнальные молекулы с помощью рецепторов.
Механизм распознавания
Клетки-мишени распознают сигнальные молекулы с помощью рецепторов.
Механизм распознавания

Рецепторы по локализации их в клетке подразделяют на две группы:
• внутриклеточные
Рецепторы по локализации их в клетке подразделяют на две группы:
• внутриклеточные

Внутриклеточные рецепторы расположены в цитозоле клетки или ядре. Химическими сигналами для

Внутриклеточные рецепторы не только распознают и связывают сигнальные молекулы, но и
Внутриклеточные рецепторы не только распознают и связывают сигнальные молекулы, но и
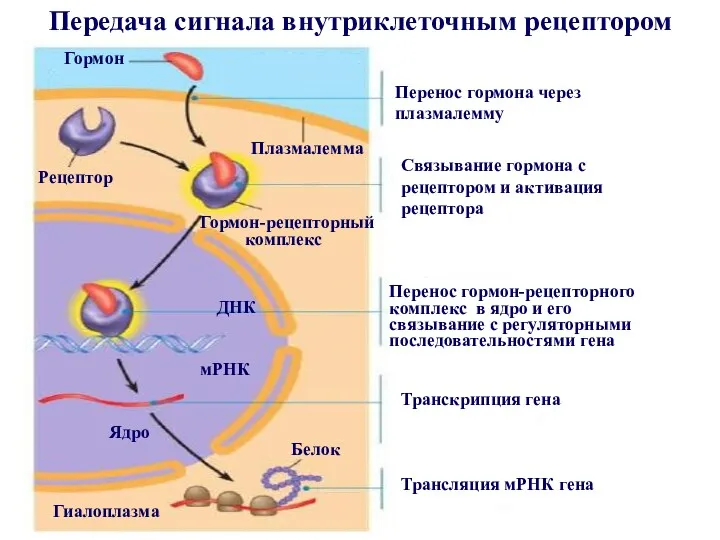
Передача сигнала внутриклеточным рецептором
Передача сигнала внутриклеточным рецептором

Рецепторы, расположенные в плазматической мембране, или рецепторы клеточной поверхности, относятся к
Рецепторы, расположенные в плазматической мембране, или рецепторы клеточной поверхности, относятся к

Выделяют три главных типа рецепторов клеточной поверхности:
рецепторы ионных каналов;
Выделяют три главных типа рецепторов клеточной поверхности:
рецепторы ионных каналов;

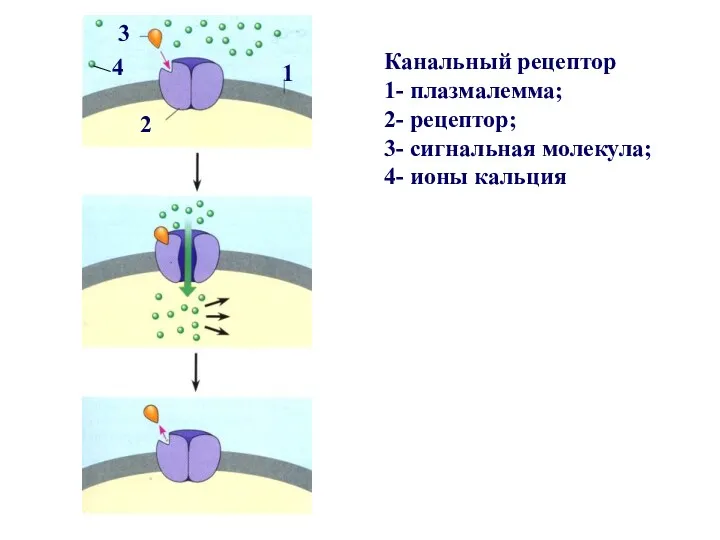
Рецепторы ионных каналов представляют собой регулируемые лигандами ионные каналы. Сигнальные молекулы,
Рецепторы ионных каналов представляют собой регулируемые лигандами ионные каналы. Сигнальные молекулы,
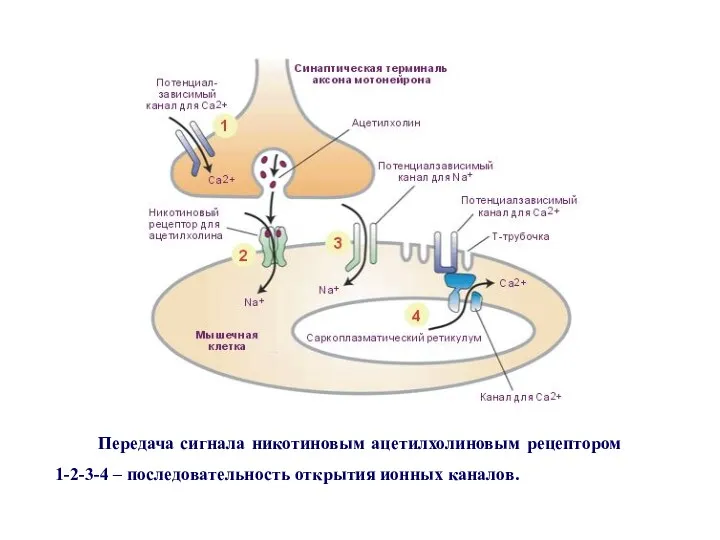
Передача сигнала никотиновым ацетилхолиновым рецептором
1-2-3-4 – последовательность открытия ионных каналов.
Передача сигнала никотиновым ацетилхолиновым рецептором
1-2-3-4 – последовательность открытия ионных каналов.


Механизм синаптической передачи сигналов между электрически возбудимыми клетками с помощью
Механизм синаптической передачи сигналов между электрически возбудимыми клетками с помощью

Рецепторы, сопряженные с G-белками, функционируют в тесной кооперации с регуляторными G-белками.
Рецепторы, сопряженные с G-белками, функционируют в тесной кооперации с регуляторными G-белками.
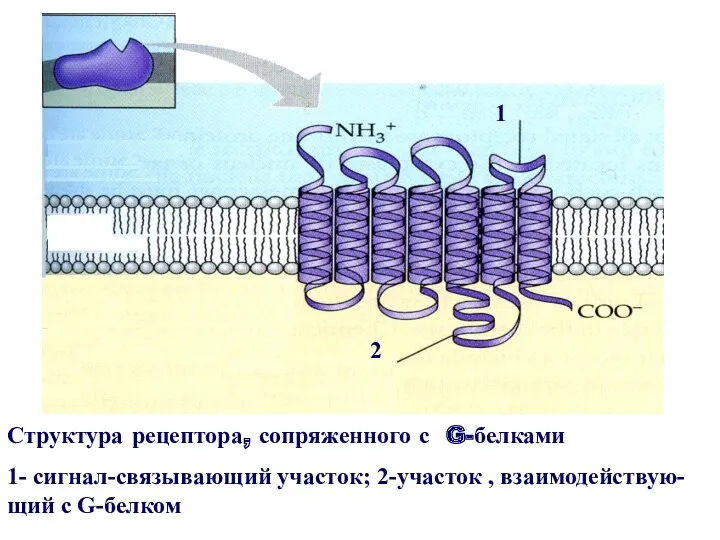
Структура рецептора, сопряженного с G-белками
1- сигнал-связывающий участок; 2-участок , взаимодействую- щий
Структура рецептора, сопряженного с G-белками
1- сигнал-связывающий участок; 2-участок , взаимодействую- щий

Рецепторы, сопряженные с G-белками, передают сигнал путем активации или инактивации
Рецепторы, сопряженные с G-белками, передают сигнал путем активации или инактивации

G-белки имеют центры связывания гуаниловых нуклеотидов: гуанозиндифосфата (ГДФ) и гуанозинтрифосфата
G-белки имеют центры связывания гуаниловых нуклеотидов: гуанозиндифосфата (ГДФ) и гуанозинтрифосфата
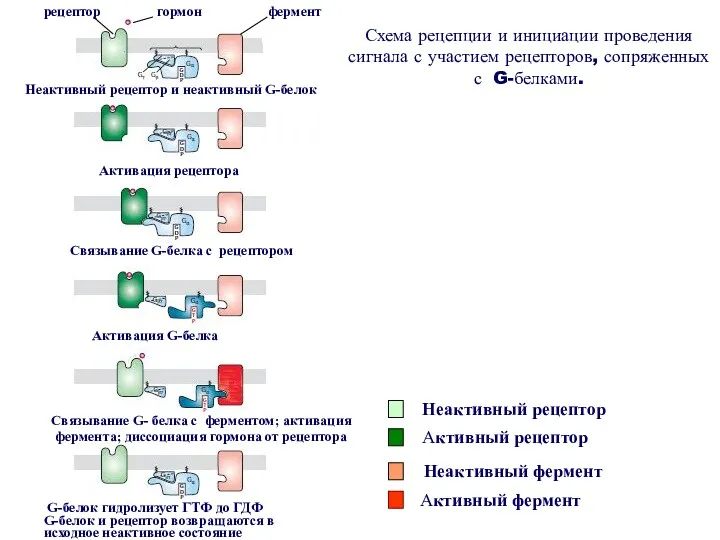

Принцип работы рецептора, сопряженного с G-белком
- связывание сигнальной молекулы с рецептором;
Принцип работы рецептора, сопряженного с G-белком
- связывание сигнальной молекулы с рецептором;

Каталитические рецепторы обладают собственной ферментативной активностью.
Сигнальные системы с участием каталитических
Каталитические рецепторы обладают собственной ферментативной активностью.
Сигнальные системы с участием каталитических
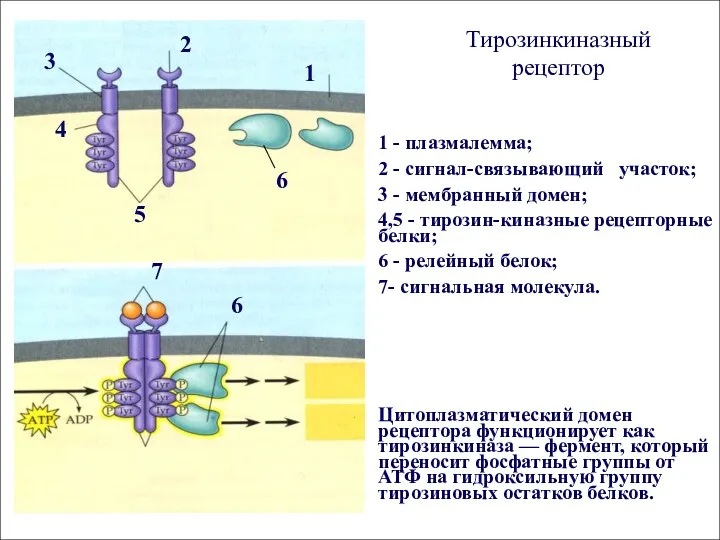
Тирозинкиназный рецептор
1 - плазмалемма;
2 - сигнал-связывающий участок;
3 - мембранный домен;
4,5 -
Тирозинкиназный рецептор
1 - плазмалемма;
2 - сигнал-связывающий участок;
3 - мембранный домен;
4,5 -

Принцип работы тирозинкиназного рецептора
- связывание сигнальной молекулы с рецептором;
агрегация
Принцип работы тирозинкиназного рецептора
- связывание сигнальной молекулы с рецептором;
агрегация

Передача сигнала от рецепторов клеточной поверхности к эффекторным молекулам включает
Передача сигнала от рецепторов клеточной поверхности к эффекторным молекулам включает

Многие релейные белки, участвующие в проведении сигнала от рецепторов клеточной
Многие релейные белки, участвующие в проведении сигнала от рецепторов клеточной

Наряду с протеинкиназами большую роль в передаче сигналов играют ферменты
Наряду с протеинкиназами большую роль в передаче сигналов играют ферменты
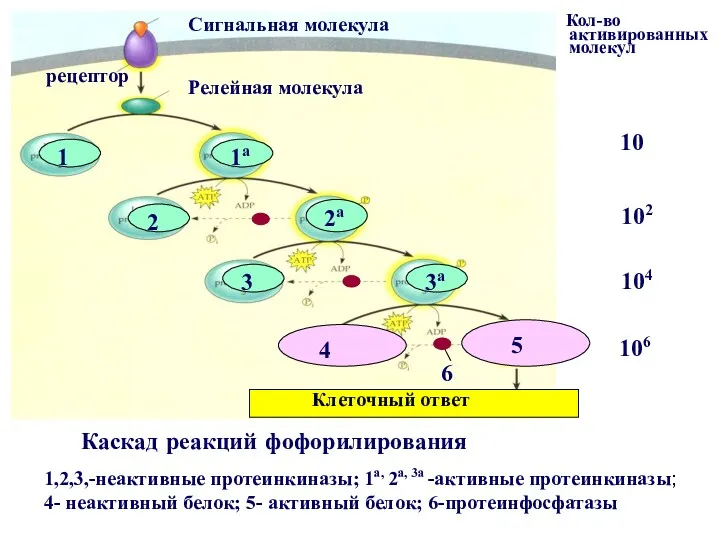
сигнальная молекула
1
1а
2а
3
3а
2
Клеточный ответ
4
5
рецептор
Каскад реакций фофорилирования
Сигнальная молекула
Релейная
сигнальная молекула
1
1а
2а
3
3а
2
Клеточный ответ
4
5
рецептор
Каскад реакций фофорилирования
Сигнальная молекула
Релейная

Система, обеспечивающая фосфорилирование и дефосфорилирование релейных белков в клетке, действует
Система, обеспечивающая фосфорилирование и дефосфорилирование релейных белков в клетке, действует
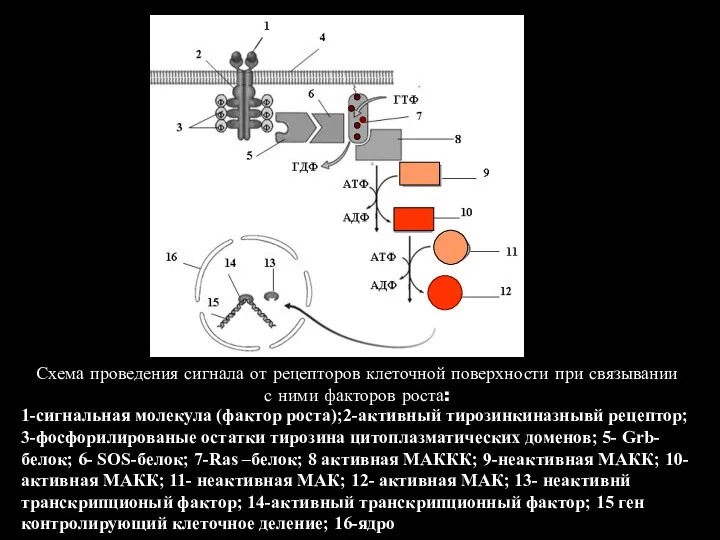
1-сигнальная молекула (фактор роста);2-активный тирозинкиназнывй рецептор; 3-фосфорилированые остатки тирозина цитоплазматических
1-сигнальная молекула (фактор роста);2-активный тирозинкиназнывй рецептор; 3-фосфорилированые остатки тирозина цитоплазматических

Во многих сигнальных путях в проведении сигнала, наряду с белками,
Во многих сигнальных путях в проведении сигнала, наряду с белками,
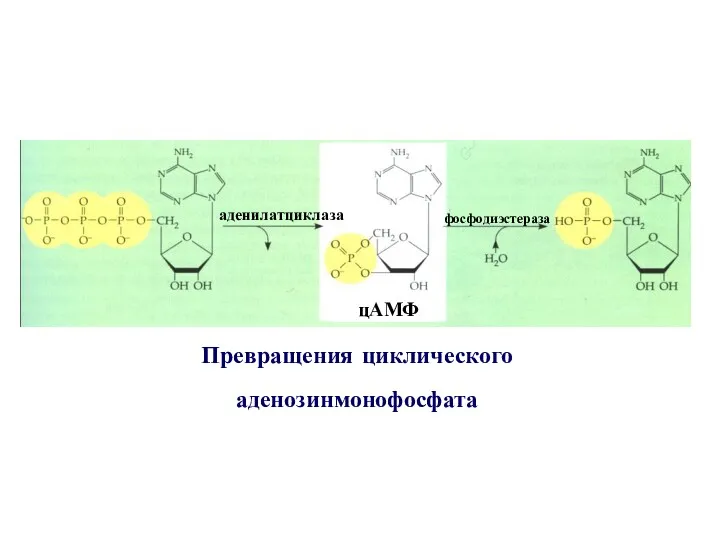
Превращения циклического
аденозинмонофосфата
Превращения циклического
аденозинмонофосфата
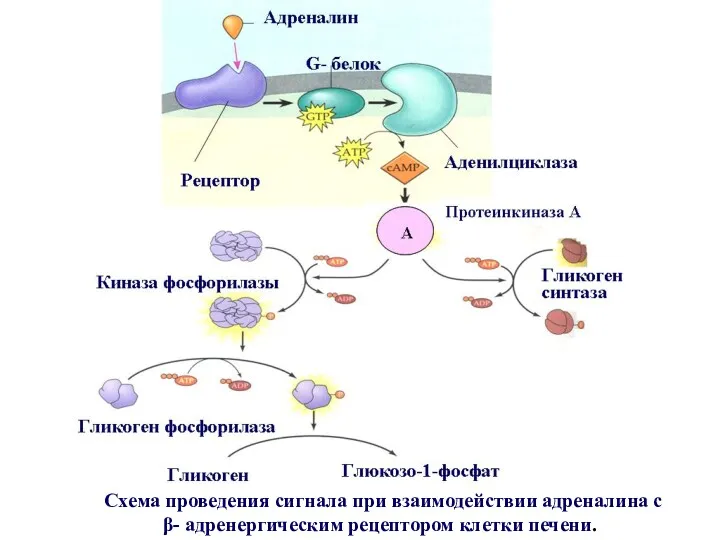
Схема проведения сигнала при взаимодействии адреналина с β- адренергическим рецептором
Схема проведения сигнала при взаимодействии адреналина с β- адренергическим рецептором

Общая схема проведения сигнала в клетке печени.
при взаимодействии адреналина с
при взаимодействии адреналина с

Действие адреналина сопровождается усилением распада гликогена и прекращением его синтеза ,
Действие адреналина сопровождается усилением распада гликогена и прекращением его синтеза ,

Использование в сигнальных системах ионов Са2+ в качестве вторичных посредников определяется
Использование в сигнальных системах ионов Са2+ в качестве вторичных посредников определяется
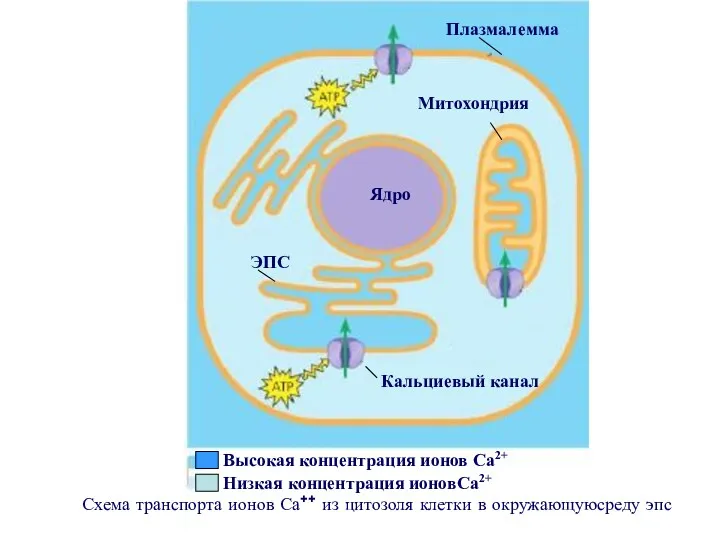
Схема транспорта ионов Са++ из цитозоля клетки в окружающуюсреду эпс
Схема транспорта ионов Са++ из цитозоля клетки в окружающуюсреду эпс

В механизме проведения сигнала с участием ионов Са++, участвуют два других
В механизме проведения сигнала с участием ионов Са++, участвуют два других
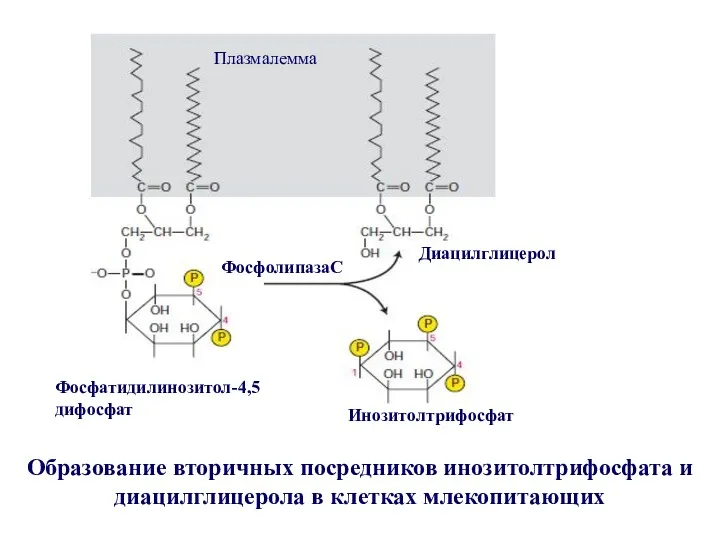
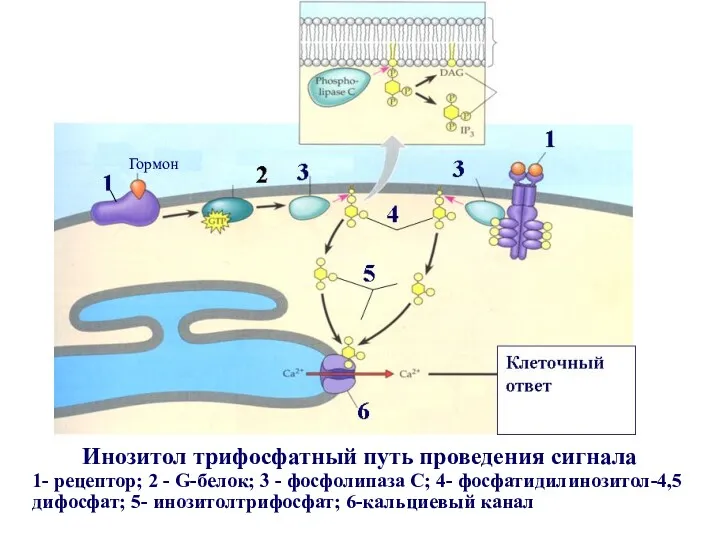
Инозитол трифосфатный путь проведения сигнала
1- рецептор; 2 - G-белок; 3 -
1- рецептор; 2 - G-белок; 3 -

Общая схема проведения сигнала с участием ионов кальция:
- связывание сигнальной молекулы
Общая схема проведения сигнала с участием ионов кальция:
- связывание сигнальной молекулы

Диацилглицерол активирует фермент протеинкиназу С, связанный с цитоплазматической мембраной клетки.
Диацилглицерол активирует фермент протеинкиназу С, связанный с цитоплазматической мембраной клетки.
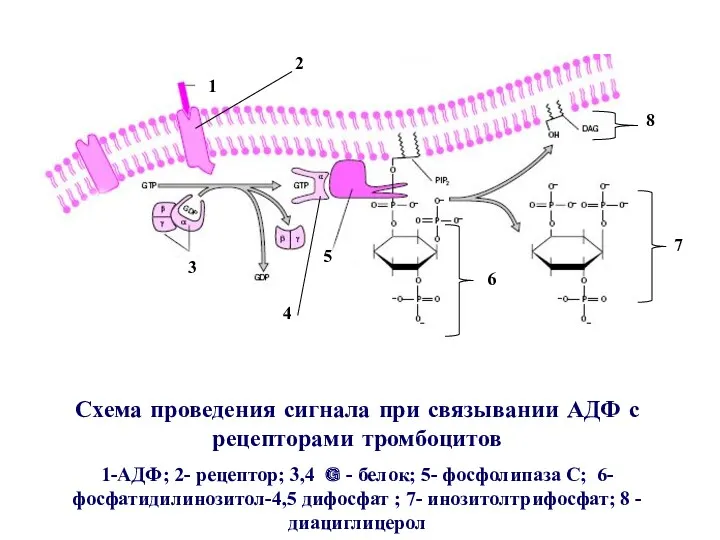
Схема проведения сигнала при связывании АДФ с рецепторами тромбоцитов
1-АДФ; 2- рецептор;
1-АДФ; 2- рецептор;

Наличие нескольких релейных молекул, вовлеченных в передачу сигнала, обеспечивает :
• усиление
Наличие нескольких релейных молекул, вовлеченных в передачу сигнала, обеспечивает :
• усиление
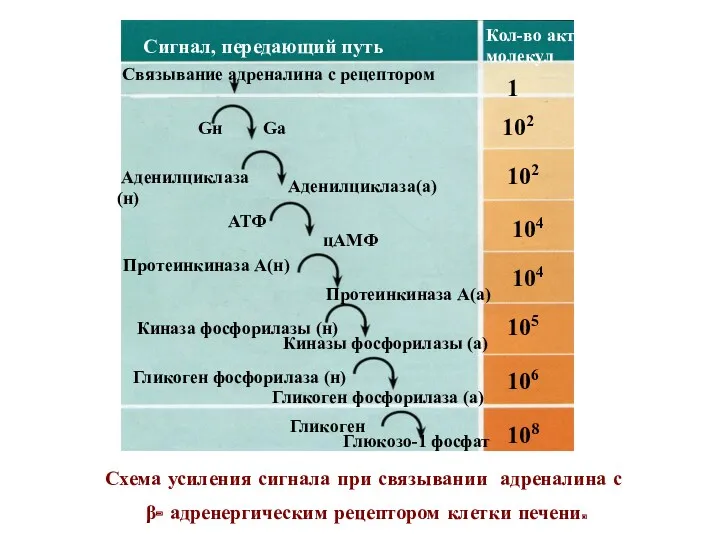
Схема усиления сигнала при связывании адреналина с
β- адренергическим
Схема усиления сигнала при связывании адреналина с
β- адренергическим
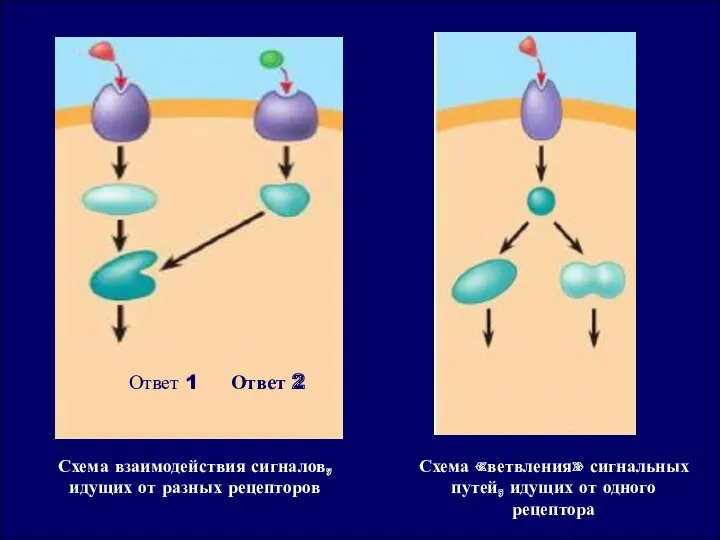
Ответ 1
Ответ 2
Схема взаимодействия сигналов, идущих от разных рецепторов
Схема «ветвления» сигнальных
Ответ 1
Ответ 2
Схема взаимодействия сигналов, идущих от разных рецепторов
Схема «ветвления» сигнальных

Ответные реакции клеток на действие сигнальных молекул отличаются высокой специфичностью.
Это
Ответные реакции клеток на действие сигнальных молекул отличаются высокой специфичностью.
Это

Специфичность ответа клетки на действие сигнальных молекул объясняется:
способностью одних и
Специфичность ответа клетки на действие сигнальных молекул объясняется:
способностью одних и
 Естественный отбор
Естественный отбор Луг и его обитатели. Растения
Луг и его обитатели. Растения Сенсорні системи рівноваги, руху, дотику, температури, болю
Сенсорні системи рівноваги, руху, дотику, температури, болю Класс Головоногие моллюски
Класс Головоногие моллюски Редкие древесные растения Приморского края
Редкие древесные растения Приморского края Как на Земле появился человек: от дриопитека к человеку прямоходящему
Как на Земле появился человек: от дриопитека к человеку прямоходящему Визначення особливостей зовнішньої будови хребетних тварин у зв’язку з пристосуванням до різних умов існування. ЛР №2
Визначення особливостей зовнішньої будови хребетних тварин у зв’язку з пристосуванням до різних умов існування. ЛР №2 Типы развития насекомых
Типы развития насекомых Использование ИКТ на уроках биологии
Использование ИКТ на уроках биологии Гендік инженерия
Гендік инженерия Слуховой анализатор. Строение
Слуховой анализатор. Строение Животноводство
Животноводство Слуховий аналізатор
Слуховий аналізатор Одномембранні органели
Одномембранні органели Организация и проведение современного урока биологии
Организация и проведение современного урока биологии Вид. Критерии вида. 9 класс
Вид. Критерии вида. 9 класс Строение нейрона. Синапс
Строение нейрона. Синапс Хромосомадан тыс тұқымқуалау заңдылықтары. Хромосомалық тұқым қуалау өзгешелігі
Хромосомадан тыс тұқымқуалау заңдылықтары. Хромосомалық тұқым қуалау өзгешелігі Вирусы. Гипотезы о происхождении вирусов
Вирусы. Гипотезы о происхождении вирусов Тип Иглокожие
Тип Иглокожие Wildlife The United Kingdom of Great Britain and Northern Ireland
Wildlife The United Kingdom of Great Britain and Northern Ireland Иван Петрович Павлов (1849-1936)
Иван Петрович Павлов (1849-1936) Фізіологічні основи формування рухових навичок в спортивному та оздоровчому тренуванні. (Лекція 5)
Фізіологічні основи формування рухових навичок в спортивному та оздоровчому тренуванні. (Лекція 5) Радиационные поражения при радиоактивном заражении
Радиационные поражения при радиоактивном заражении Animales en el zoológico de Madrid
Animales en el zoológico de Madrid Карпеченко Георгий Дмитриевич (1899-1942) - русский цитогенетик
Карпеченко Георгий Дмитриевич (1899-1942) - русский цитогенетик Группа веществ, изолируемых из биологического материала экстракцией и сорбцией. (Лекция 6)
Группа веществ, изолируемых из биологического материала экстракцией и сорбцией. (Лекция 6) Китообразные
Китообразные