- Строение нейрона. Синапс

Содержание
- 2. Динамометр кистевой
- 3. Что такое нейрон? Нейрон — это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко
- 4. Строение нейрона Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим
- 5. Отростки нейронов Аксон — обычно длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты —
- 6. Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии. Дендриты не имеют
- 7. Миелинизированное нервное волокно Миелиновая оболочка — электроизолирующая оболочка, покрывающая аксоны многих нейронов. Миелиновую оболочку образуют глиальные
- 8. Формирование миелиновой оболочки Миелиновая оболочка формируется из плоского выроста тела глиальной клетки, многократно оборачивающего аксон подобно
- 9. Немиелиновое волокно Безмякотные нервные волокна построены примитивнее, нежели мякотные нервные волокна. В состав безмякотного нервного волокна
- 10. Классификация нейронов Чувствительные нейроны Двигательные нейроны Вставочные нейроны
- 11. Возникновение ПД в нейронах и НВ аналогично ранее описанным, но имеются свои особенности : Возбудимость НВ
- 13. 2 способа проведения возбуждения: В безмякотных НВ возбуждения (ПД) проводятся беспрерывно, декрементно (затухая). По мякотным НВ
- 14. Проведение возбуждение по НВ осуществляется по 3-законам: Анатомическая и физиологическая целостность. Двустороннее проведение. Изолированное проведение Относительной
- 15. Проведение возбуждения (ПД) в НВ осуществляется посредством электротонического распространения ПД вдоль волокна от возбужденного (-) к
- 16. Скорость проведения возбуждения по НВ зависит от : Скорости нарастания ПД Константы длины НВ. Диаметра НВ.
- 17. Амплитуда ПД НВ не зависит от силы раздражения, она определяется биологическими особенностями мембраны. Амплитуда ПД нерва
- 18. При регистрации возбуждений нервного ствола на определенном расстоянии от место раздражения наблюдается ряд последовательных ПД с
- 19. Функциональная классификация нервных волокон Волокна Аα (толстые миелиновые); Аβ; Аγ; Аδ; В (миелин, преганглионарные); С (безмиел.
- 20. При проведении возбуждении утомление НВ практически не наблюдается, так как обмен веществ в них очень низок
- 21. Парабиоз нерва (около жизни) – состояние, возникающее при воздействии на НВ химических агентов (аммиак,спирт, новокаин), характеризующееся
- 22. Синапс Синапс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой.
- 23. Классификация синапсов По морфологическому принципу: • нейро-мышечные • нейро-секреторные • нейро-нейрональные • аксо-соматические • аксо-аксональные •
- 24. Конец аксона теряет миелиновую оболочку и образует небольшое утолщение (синаптическую бляшку). Часть, контактирующая с иннервируемой клеткой,
- 25. В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче
- 26. Классификация синапсов в зависимости от механизма передачи нервного импульса: химические; электрические; смешанные синапсы;
- 27. Рис. 1 Особенности ультраструктуры электрического синапса и химического синапса
- 28. Основные характеристики, отличающие химическую синаптическую передачу от электрической В химическом синапсе постсинаптический ток генерируется за счет
- 29. Механизм передачи возбуждения в электрическом синапсе Аналогичен механизму проведения возбуждения в НВ. Во время развития ПД
- 30. Свойства электрических и химических синапсов
- 31. Мионевральный синапс — эффекторное нервное окончание на скелетном мышечном волокне; зона контакта двигательного окончания и мышечного
- 32. Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. По химической структуре
- 33. Факторы, определяющие концентрацию нейромедиатора в синаптической щели: Количество нейромедиатора, высвобождаемого пресинаптическим нервным окончанием. Пассивная диффузия медиатора
- 34. Синаптическая задержка Время от прихода нервного импульса до развития постсинаптического ответа в химическом синапсе составляет около
- 35. Медиаторы могут (ацетилхолин) синтезоватся в цитоплазме синаптического окончания, и депонируются в синаптических пузырьках. Ферменты, для синтеза
- 36. Медиатора могут выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении пузырька в синаптичекую щель выбрасывается порция
- 37. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+
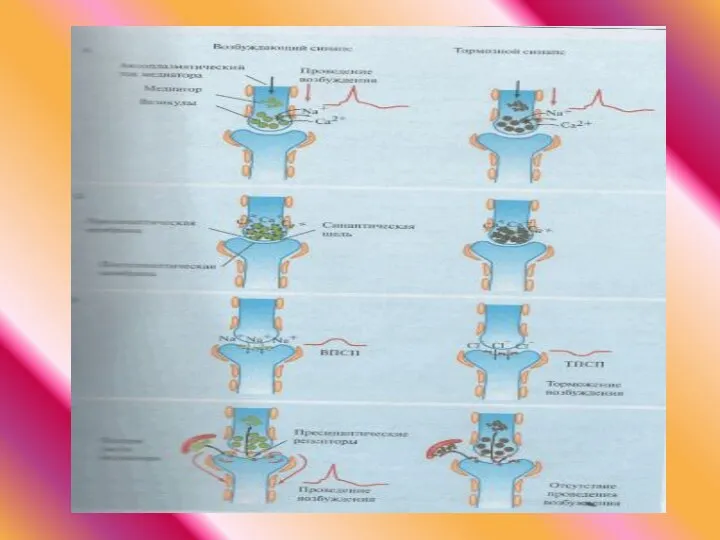
- 38. Возбуждающий постсинаптический потенциал (ВПСП) локальный потенциал, который электротонически распространяется по мембране. С помощью внутриклеточных микроэлектродов и
- 39. Синаптическим торможением обозначают влияние пресинаптической нервной клетки, прекращающее или предотвращающее возбуждение постсинаптической нервной клетки.
- 40. Тормозной постсинаптический потенциал (ТПСП) имеет вид гиперполяризации развитие ТПСП связано со значительным увеличением ионной проводимости развивается
- 42. Временная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, исходящих из одного и того же
- 43. Пространственная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, поступающих одновременно из разных пресинаптических нейронов,
- 44. Явление суммации: А — пространственная суммация в результате одновременно наносимых раздражении: а — передача возбуждения с
- 45. Ацетилхолин Возбуждающий медиатор: медиатор α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру, ретикулярной формации, гипоталамусе. Обнаружены M- и
- 46. Адреналин (эпинефрин) (L-1(3,4-Диоксифенил)-2-метиламиноэтанол)
- 47. γ-Аминомасляная кислота (ГАМК, GABA) Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы человека и млекопитающих. Аминомасляная кислота
- 48. Глици́н (аминоуксусная кислота) является нейромедиаторной аминокислотой. Рецепторы к глицину имеются во многих участках головного мозга и
- 49. Серотонин, 5-гидрокситриптамин, 5-НТ важный нейромедиатор и гормон. По химическому строению он относится к биогенным аминам, к
- 50. Глутаминовая кислота является нейромедиаторной аминокислотой одним из важных представителей класса «возбуждающих аминокислот». Связывание аниона глутамата со
- 51. Лекарства и яды, вызывающие мышечную слабость и паралич Нейротоксин ботулин – ингибирует высвобождение нейромедиатора из везикул
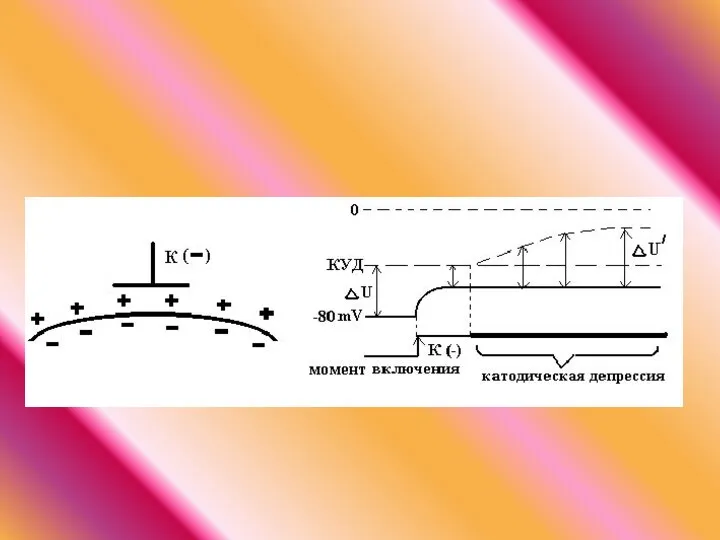
- 52. ЭЛЕКТРОТОН это пассивное изменение функциональных характеристик тканей (МПП) в области полюсов постоянного тока.
- 53. Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока
- 57. Скачать презентацию

Динамометр кистевой
Динамометр кистевой
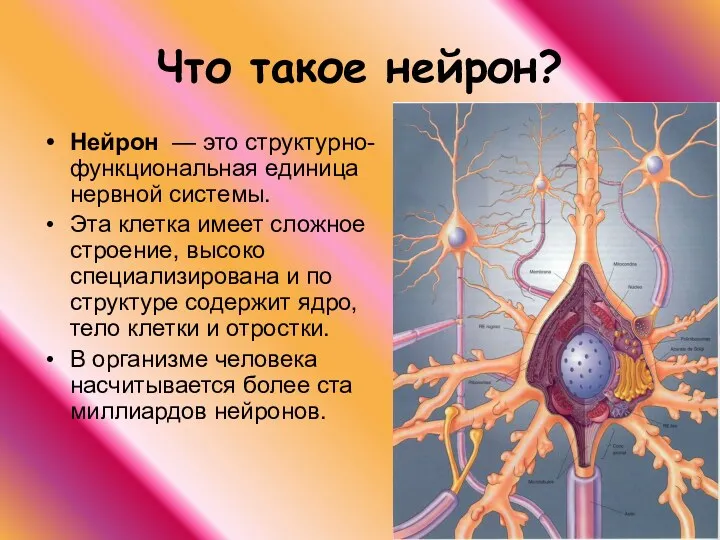
Что такое нейрон?
Нейрон — это структурно-функциональная единица нервной системы.
Эта клетка
Что такое нейрон?
Нейрон — это структурно-функциональная единица нервной системы.
Эта клетка
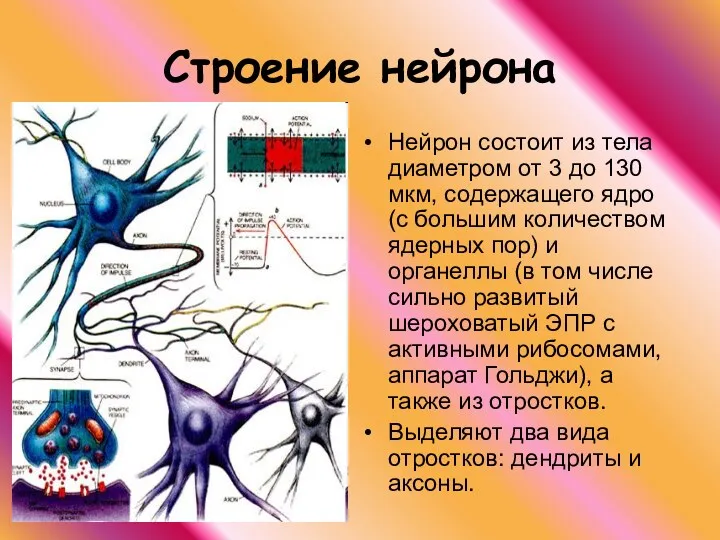
Строение нейрона
Нейрон состоит из тела диаметром от 3 до 130 мкм,
Строение нейрона
Нейрон состоит из тела диаметром от 3 до 130 мкм,
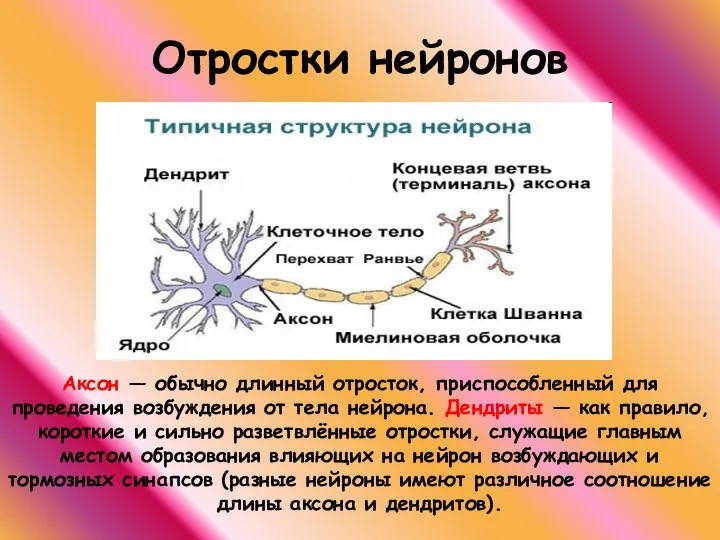
Отростки нейронов
Аксон — обычно длинный отросток, приспособленный для проведения возбуждения от тела
Отростки нейронов
Аксон — обычно длинный отросток, приспособленный для проведения возбуждения от тела
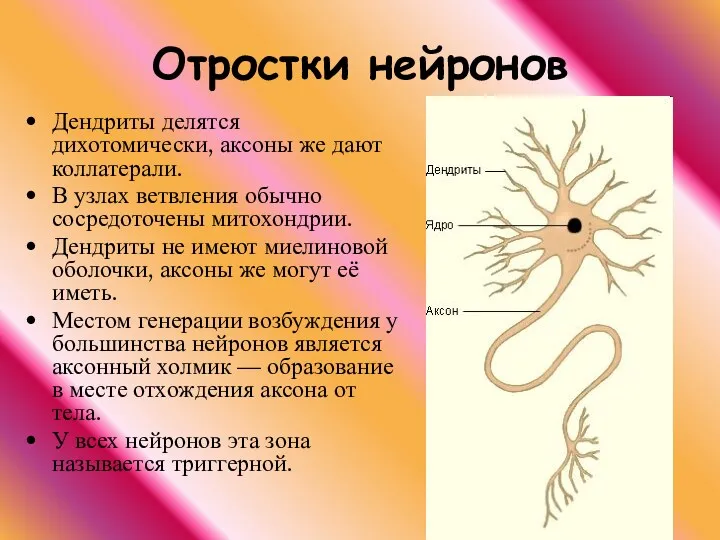
Дендриты делятся дихотомически, аксоны же дают коллатерали.
В узлах ветвления обычно
Дендриты делятся дихотомически, аксоны же дают коллатерали.
В узлах ветвления обычно
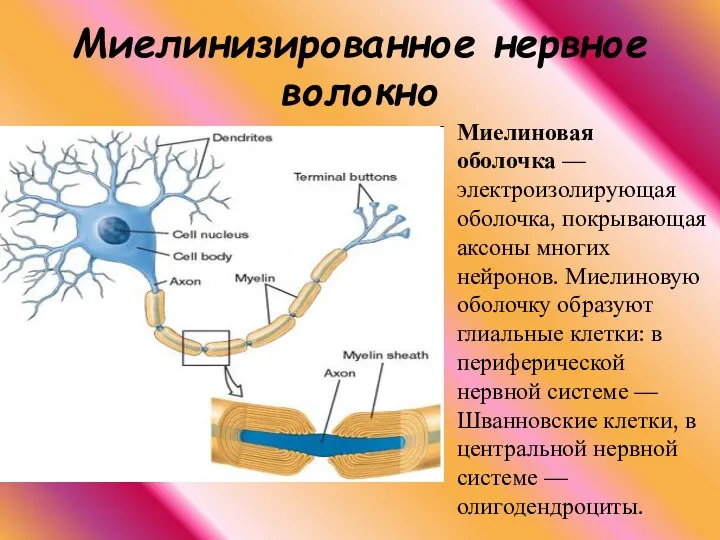
Миелинизированное нервное волокно
Миелиновая оболочка — электроизолирующая оболочка, покрывающая аксоны многих нейронов. Миелиновую
Миелинизированное нервное волокно
Миелиновая оболочка — электроизолирующая оболочка, покрывающая аксоны многих нейронов. Миелиновую
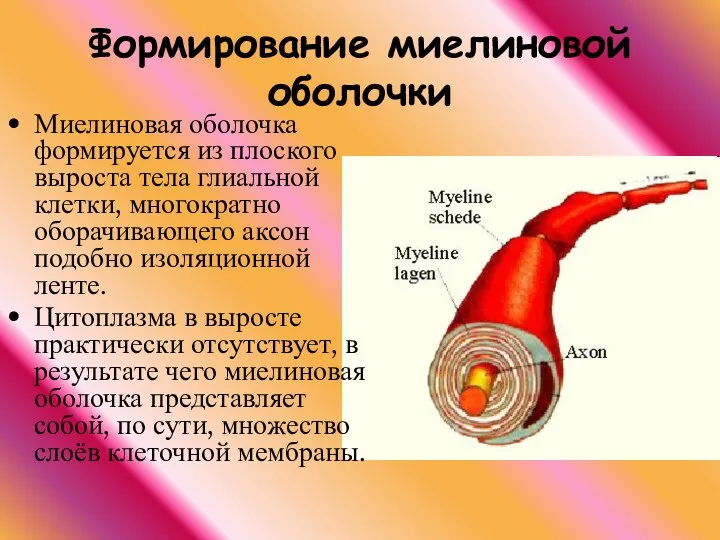
Формирование миелиновой оболочки
Миелиновая оболочка формируется из плоского выроста тела глиальной клетки,
Формирование миелиновой оболочки
Миелиновая оболочка формируется из плоского выроста тела глиальной клетки,

Немиелиновое волокно
Безмякотные нервные волокна построены примитивнее, нежели мякотные нервные волокна.
В состав
Немиелиновое волокно
Безмякотные нервные волокна построены примитивнее, нежели мякотные нервные волокна.
В состав
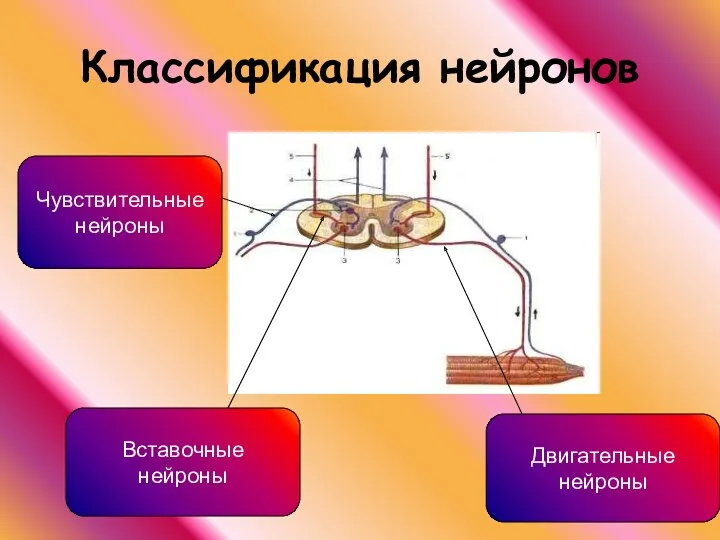
Классификация нейронов
Чувствительные
нейроны
Двигательные
нейроны
Вставочные
нейроны
Классификация нейронов
Чувствительные
нейроны
Двигательные
нейроны
Вставочные
нейроны

Возникновение ПД в нейронах и НВ аналогично ранее описанным, но имеются
Возникновение ПД в нейронах и НВ аналогично ранее описанным, но имеются
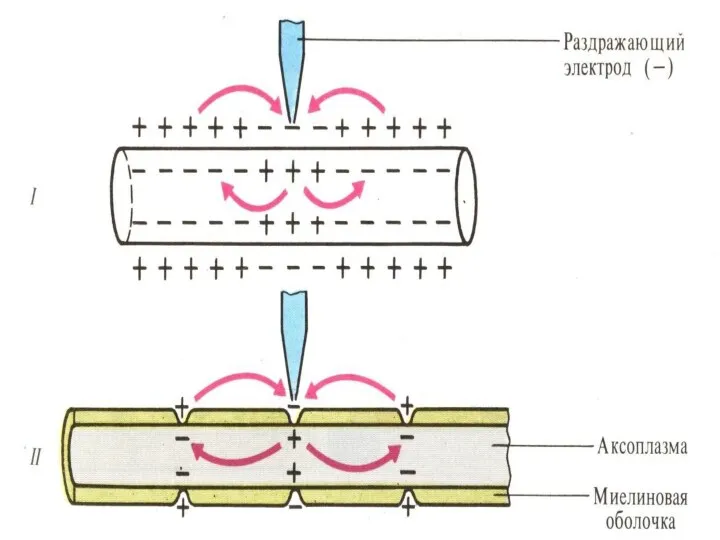

2 способа проведения возбуждения:
В безмякотных НВ возбуждения (ПД) проводятся беспрерывно, декрементно
2 способа проведения возбуждения:
В безмякотных НВ возбуждения (ПД) проводятся беспрерывно, декрементно

Проведение возбуждение по НВ осуществляется по 3-законам:
Анатомическая и физиологическая целостность.
Двустороннее проведение.
Изолированное
Проведение возбуждение по НВ осуществляется по 3-законам:
Анатомическая и физиологическая целостность.
Двустороннее проведение.
Изолированное

Проведение возбуждения (ПД) в НВ осуществляется посредством электротонического распространения ПД вдоль
Проведение возбуждения (ПД) в НВ осуществляется посредством электротонического распространения ПД вдоль

Скорость проведения возбуждения по НВ зависит от :
Скорости нарастания ПД
Константы
Скорость проведения возбуждения по НВ зависит от :
Скорости нарастания ПД
Константы

Амплитуда ПД НВ не зависит от силы раздражения, она определяется биологическими
Амплитуда ПД НВ не зависит от силы раздражения, она определяется биологическими

При регистрации возбуждений нервного ствола на определенном расстоянии от место раздражения
При регистрации возбуждений нервного ствола на определенном расстоянии от место раздражения

Функциональная классификация нервных волокон
Волокна Аα (толстые миелиновые);
Аβ;
Аγ;
Аδ;
В (миелин, преганглионарные);
С (безмиел. постганглионарные).
Функциональная классификация нервных волокон
Волокна Аα (толстые миелиновые);
Аβ;
Аγ;
Аδ;
В (миелин, преганглионарные);
С (безмиел. постганглионарные).

При проведении возбуждении утомление НВ практически не наблюдается, так как обмен
При проведении возбуждении утомление НВ практически не наблюдается, так как обмен

Парабиоз нерва (около жизни) – состояние, возникающее при воздействии на НВ
Парабиоз нерва (около жизни) – состояние, возникающее при воздействии на НВ
Синапс
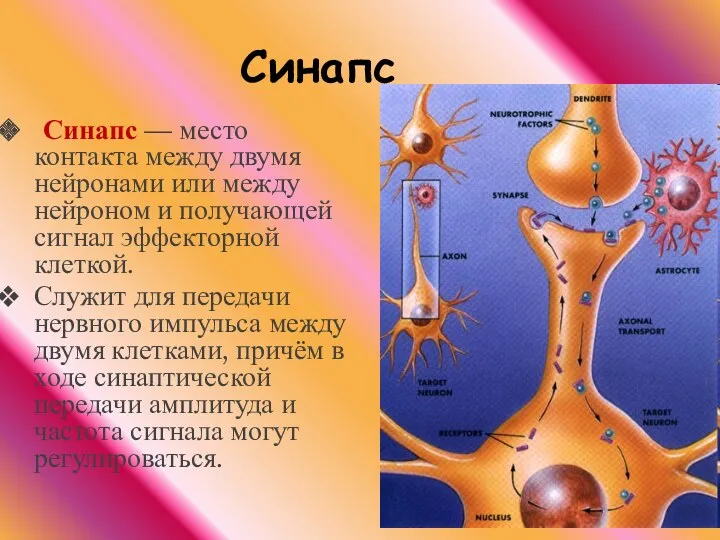
Синапс — место контакта между двумя нейронами или между нейроном и
Синапс
Синапс — место контакта между двумя нейронами или между нейроном и
Классификация синапсов
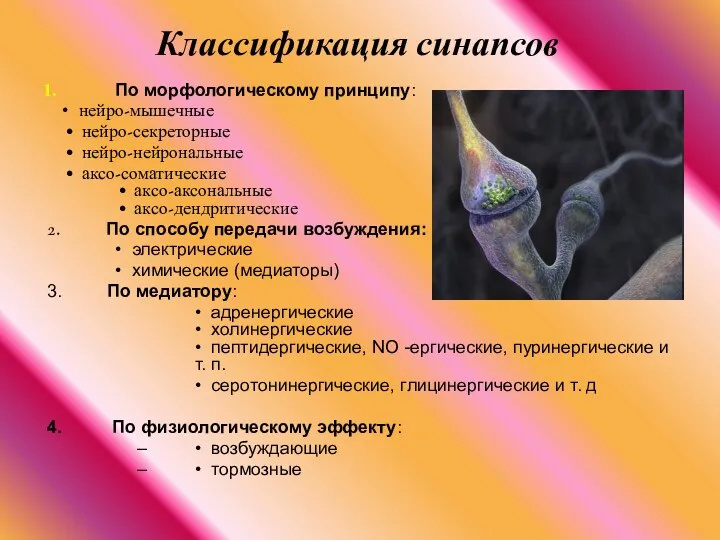
По морфологическому принципу:
• нейро-мышечные
• нейро-секреторные
Классификация синапсов
По морфологическому принципу:
• нейро-мышечные
• нейро-секреторные

Конец аксона теряет миелиновую оболочку и образует небольшое утолщение (синаптическую бляшку).
Конец аксона теряет миелиновую оболочку и образует небольшое утолщение (синаптическую бляшку).
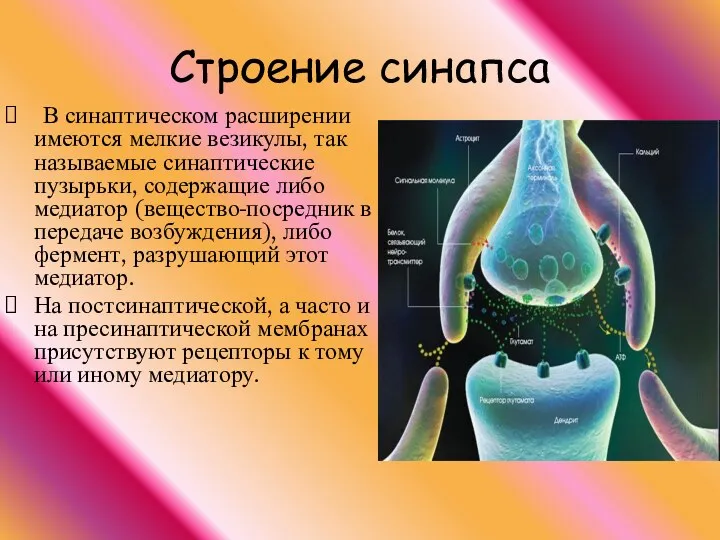
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие
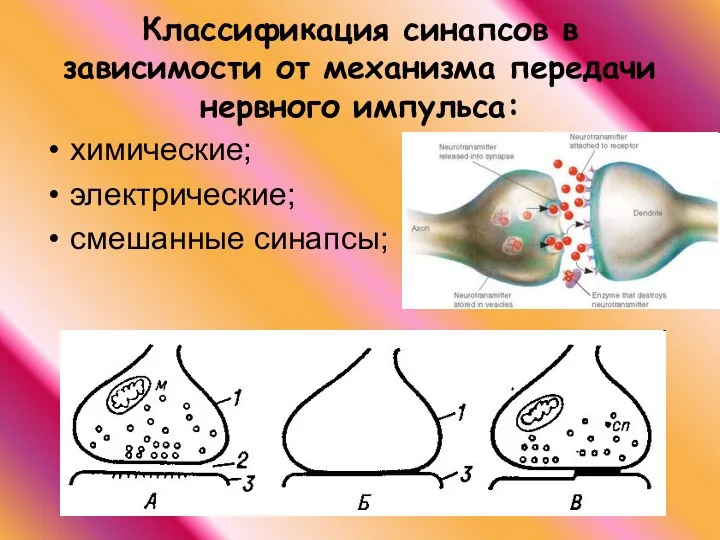
Классификация синапсов в зависимости от механизма передачи нервного импульса:
химические;
электрические;
смешанные синапсы;
Классификация синапсов в зависимости от механизма передачи нервного импульса:
химические;
электрические;
смешанные синапсы;
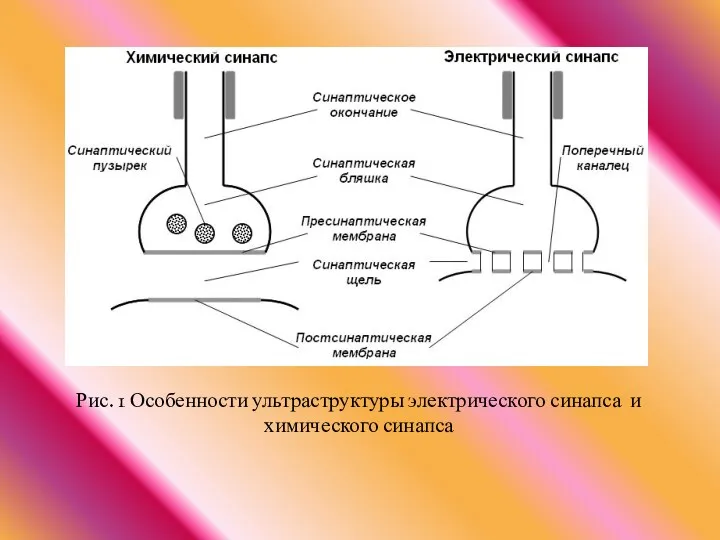
Рис. 1 Особенности ультраструктуры электрического синапса и химического синапса
Рис. 1 Особенности ультраструктуры электрического синапса и химического синапса

Основные характеристики, отличающие химическую синаптическую передачу от электрической
В химическом синапсе
Основные характеристики, отличающие химическую синаптическую передачу от электрической
В химическом синапсе

Механизм передачи возбуждения в электрическом синапсе
Аналогичен механизму проведения возбуждения в
Механизм передачи возбуждения в электрическом синапсе
Аналогичен механизму проведения возбуждения в

Свойства электрических и химических синапсов
Свойства электрических и химических синапсов

Мионевральный синапс — эффекторное нервное окончание на скелетном мышечном волокне; зона

Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в

Факторы, определяющие концентрацию нейромедиатора в синаптической щели:
Количество нейромедиатора, высвобождаемого пресинаптическим нервным
Факторы, определяющие концентрацию нейромедиатора в синаптической щели:
Количество нейромедиатора, высвобождаемого пресинаптическим нервным

Синаптическая задержка
Время от прихода нервного импульса до развития постсинаптического ответа в
Синаптическая задержка
Время от прихода нервного импульса до развития постсинаптического ответа в

Медиаторы могут (ацетилхолин) синтезоватся в цитоплазме синаптического окончания, и депонируются в
Медиаторы могут (ацетилхолин) синтезоватся в цитоплазме синаптического окончания, и депонируются в

Медиатора могут выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении пузырька

Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где

Возбуждающий постсинаптический потенциал (ВПСП)
локальный потенциал, который электротонически распространяется по мембране. С
Возбуждающий постсинаптический потенциал (ВПСП)
локальный потенциал, который электротонически распространяется по мембране. С

Синаптическим торможением
обозначают влияние пресинаптической нервной клетки, прекращающее или предотвращающее возбуждение
Синаптическим торможением
обозначают влияние пресинаптической нервной клетки, прекращающее или предотвращающее возбуждение

Тормозной постсинаптический потенциал (ТПСП)
имеет вид гиперполяризации
развитие ТПСП связано со значительным увеличением
Тормозной постсинаптический потенциал (ТПСП)
имеет вид гиперполяризации
развитие ТПСП связано со значительным увеличением

Временная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, исходящих
Временная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, исходящих

Пространственная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, поступающих
Пространственная суммация представляет собой суммарное воздействие нескольких ВПСП или ТПСП, поступающих
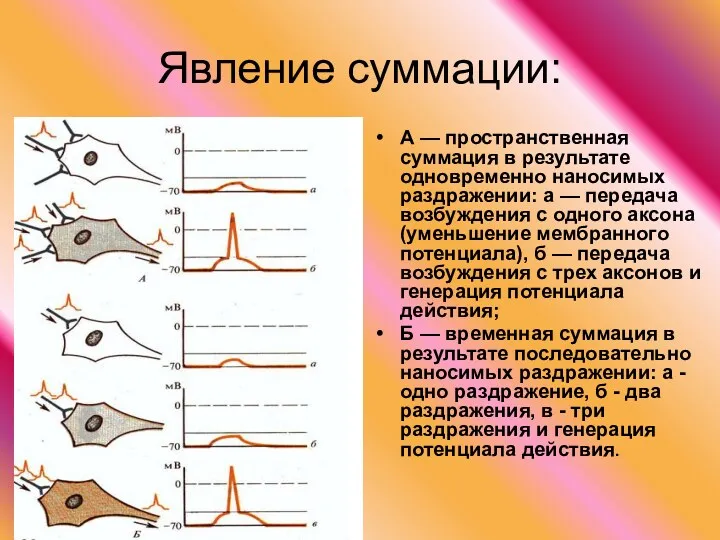
Явление суммации:
А — пространственная суммация в результате одновременно наносимых раздражении:
Явление суммации:
А — пространственная суммация в результате одновременно наносимых раздражении:

Ацетилхолин
Возбуждающий медиатор: медиатор α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру, ретикулярной формации,
Ацетилхолин
Возбуждающий медиатор: медиатор α-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру, ретикулярной формации,
Адреналин (эпинефрин) (L-1(3,4-Диоксифенил)-2-метиламиноэтанол)
Адреналин (эпинефрин) (L-1(3,4-Диоксифенил)-2-метиламиноэтанол)
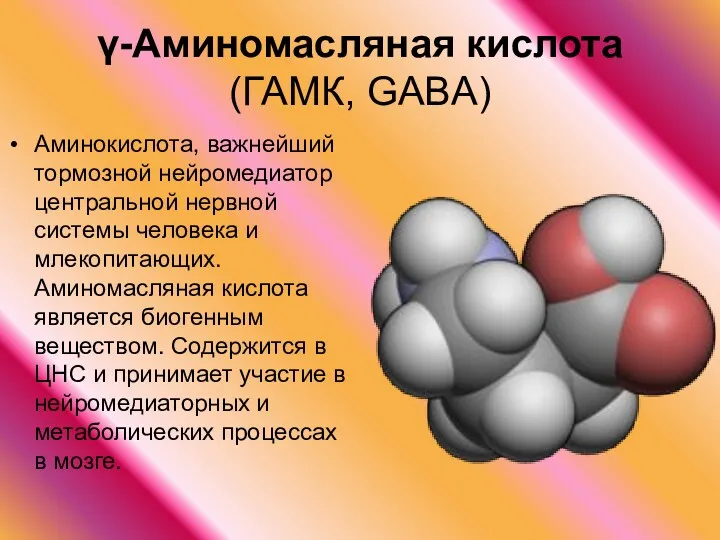
γ-Аминомасляная кислота (ГАМК, GABA)
Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы
γ-Аминомасляная кислота (ГАМК, GABA)
Аминокислота, важнейший тормозной нейромедиатор центральной нервной системы
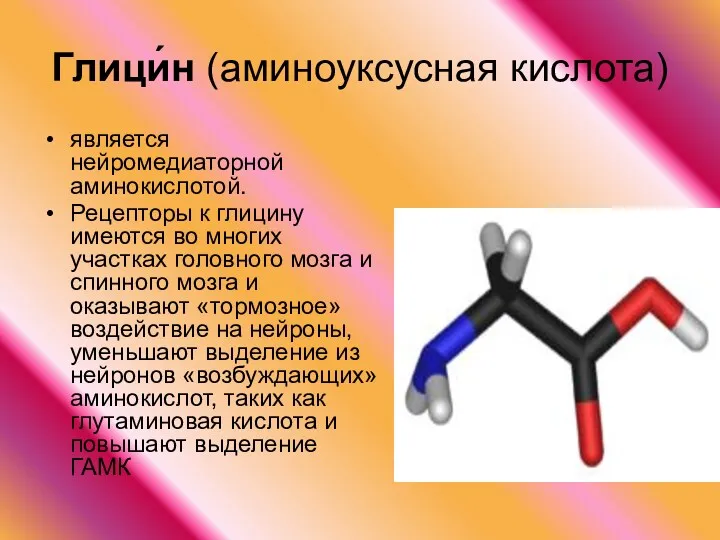
Глици́н (аминоуксусная кислота)
является нейромедиаторной аминокислотой.
Рецепторы к глицину имеются во многих
Глици́н (аминоуксусная кислота)
является нейромедиаторной аминокислотой.
Рецепторы к глицину имеются во многих
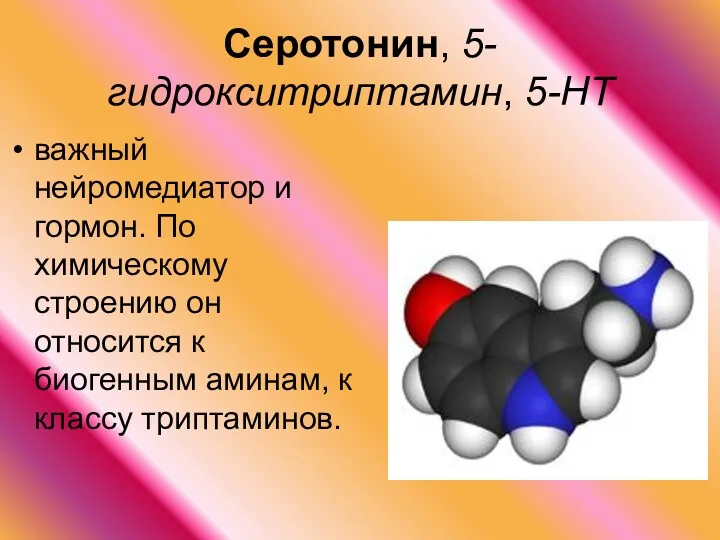
Серотонин, 5-гидрокситриптамин, 5-НТ
важный нейромедиатор и гормон. По химическому строению он относится к
Серотонин, 5-гидрокситриптамин, 5-НТ
важный нейромедиатор и гормон. По химическому строению он относится к

Глутаминовая кислота
является нейромедиаторной аминокислотой одним из важных представителей класса «возбуждающих аминокислот».
Глутаминовая кислота
является нейромедиаторной аминокислотой одним из важных представителей класса «возбуждающих аминокислот».

Лекарства и яды, вызывающие мышечную слабость и паралич
Нейротоксин ботулин – ингибирует
Лекарства и яды, вызывающие мышечную слабость и паралич
Нейротоксин ботулин – ингибирует

ЭЛЕКТРОТОН это пассивное изменение функциональных характеристик тканей (МПП) в области
ЭЛЕКТРОТОН это пассивное изменение функциональных характеристик тканей (МПП) в области

Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее
Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее
 Царство Грибы. Общая характеристика и значение
Царство Грибы. Общая характеристика и значение Проводящие пути центральной нервной системы
Проводящие пути центральной нервной системы Биосинтез белка. Биосинтез углеводов
Биосинтез белка. Биосинтез углеводов Строение тела человека. Окружающий мир. 2 класс
Строение тела человека. Окружающий мир. 2 класс Черенкование комнатных растений
Черенкование комнатных растений Новые подходы к ремедиации почв, загрязненных пестицидом Гезагард
Новые подходы к ремедиации почв, загрязненных пестицидом Гезагард Фотосинтездiң жарық сатысы
Фотосинтездiң жарық сатысы Отряд перепончатокрылые. Пчелы
Отряд перепончатокрылые. Пчелы Жири, склад жирів, їх утворення. Гідроліз та гідрування жирів. Жири в природі. Біологічна роль жирів
Жири, склад жирів, їх утворення. Гідроліз та гідрування жирів. Жири в природі. Біологічна роль жирів Ұлу қабыршағы
Ұлу қабыршағы Роль бактерії в екосистемі
Роль бактерії в екосистемі Нуклеиновые кислоты. Биология. 10 класс. Профильный уровень
Нуклеиновые кислоты. Биология. 10 класс. Профильный уровень Вакцинация животных
Вакцинация животных Хронобиология и биоритмы человека
Хронобиология и биоритмы человека Идентификационные признаки семейства тресковых
Идентификационные признаки семейства тресковых Этология с основами зоопсихологии
Этология с основами зоопсихологии Тварина минулого - мамонт
Тварина минулого - мамонт Испарение воды растениями. Листопад
Испарение воды растениями. Листопад Семейства кипрейные и дербенниковые
Семейства кипрейные и дербенниковые Биоэнергетика. Современное представление о биологическом окислении
Биоэнергетика. Современное представление о биологическом окислении Семейство крестоцветные
Семейство крестоцветные Естественный отбор и борьба за существование
Естественный отбор и борьба за существование Презентация Лекарственные растения
Презентация Лекарственные растения Строение и жизненные циклы первичнополостных червей. Тип nematoda
Строение и жизненные циклы первичнополостных червей. Тип nematoda Поле чудес
Поле чудес Ферменты. Свойства и строение ферментов
Ферменты. Свойства и строение ферментов Сон-трава или прострел раскрытый
Сон-трава или прострел раскрытый Метод классификации организмов, применение двойных названий организмов. Биология. 5 класс
Метод классификации организмов, применение двойных названий организмов. Биология. 5 класс