- Биотехнология в селекции растений. Часть 2. Вспомогательные технологии in vitro

Содержание
- 2. Основные селекционные задачи: Создание нового исходного материала для селекции Ускорение селекционного процесса Повышение эффективности отбора ценных
- 3. Генетическая неоднородность клеток в культуре Соматические мутации Генетические мутации Полиплоидия и анеуплоидия Физические мутагены – рентгеновское,
- 4. Соматическая гибридизация, слияние протопластов (этиленгликоль, высокая концентрация двухвалентных ионов, высокое значение рН, лазерное облучение) Низкий выход
- 5. ИСПОЛЬЗОВАНИЕ МЕТОДОВ IN VITRO В СЕЛЕКЦИИ: Преодоление прогамной несовместимости Преодоление постгамной несовместимости Клональное микроразмножение отдаленных гибридов
- 6. Оплодотворение in vitro – преодоление прогамной несовместимости Причины: Физиологические (несоответствие во времени созревания) Морфологические (короткая пыльцевая
- 7. Модификации метода опыления и оплодотворения in vitro - прививка столбика с рыльцем совместимого гинецея в завязь
- 8. Преодоление постгамной несовместимости Отделенная гибридизация Эмбриокультура (одно- или двухфазная) Пшенично-ржаные гибриды (доращивание незрелых зародышей) Межвидовая гибридизация
- 9. Исследования по культуре незрелых зародышей: оптимальный состав питательной среды - жидкие и твердые питательные среды -
- 10. Показана возможность увеличения выхода пшенично-ржаных гибридов путем доращивания незрелых зародышей, эмбриокультуры; Получены триплоидные растения ржи от
- 11. Клональное микроразмножение отдаленных гибридов Активация развития меристемы пазушных почек (черенкование стерильных побегов) Адвентивными почками Регенерация растений
- 12. Получение гаплоидов in vitro Быстрый поиск комбинаций - Сокращение срока селекции Сокращение объемов питомников Расширение генетического
- 13. Получение гаплоидов in vitro Стимуляция деления неоплодотворенной яйцеклетки опылением убитой пыльцой Задержка опыления Способы получения: Андрогенез
- 14. Партеногенез – из гибридного зародыша, потерявшего отцовские хромосомы, метод гаплопродюссера H. bulbosum для H. vulgare Penissetum
- 15. Сдерживающие факторы использования гаплоидных технологий: 1. Эмпирический подход к поиску генотипов, отзывчивых на условия культивирования пыльников
- 16. Методы создания гаплоидных растений отдаленная гибридизация внутривидовая гибридизация температурные воздействия метод ионизирующей радиации метод близнецов химический
- 17. Температурные воздействия Индукция гаплоидии у кукурузы путем обработки материнских растений через сутки после опыления температурой +43
- 18. Метод близнецов Близнецы – пары или группы самостоятельных растений, полученных из одного семени. Закладка в одной
- 19. Повышение частоты возникновения гаплоидов при обработке рылец ИУК (до 1: 1550) и НУК (до 1:913) –
- 20. Спонтанный эмбриогенез в культуре изолированных репродуктивных органов имеет низкий выход гаплоидов 1-4 %, результаты плохо воспроизводимы
- 21. Важно выявлять корреляцию между морфологическим показателями генеративных органов растения и стадией микроспоро- и гаметогенеза Длина пыльника
- 22. Частота образования гаплоидов зависит от генотипа исходного растения, видовой и сортовой принадлежности (у Пасленовых легче получить
- 23. N. tabacum +3-5 °C во время первого митоза – возникновение двух одинаковых дочерних ядер Температурный шок
- 26. Nicotiana: предварительное инкубирование цветочных почек при 7 °С 7 сут. Картофель: 4 °С в течение 48-72
- 27. Микроспоры продуцируют и выделяют в культуральную среду кондиционирующие факторы, индуцирующие спорофитный морфогенез в культуре микроспор После
- 28. Воздействие повышенных температур на изолированные экспланты Микроспоры рапса +32,5 °С 4 сут. - переход на эмбриогенный
- 29. Повышенные температуры вызывают распад микротрубочек, нарушение веретена деления, ненормальное деление ядер микроспор, образование эмбриоидов и/или каллусов
- 30. Синтез этилена блокирует нитрат серебра (рис, капуста, пшеница и др.) Обработка колосьев яровой пшеницы кинетином, 2,4-Д,
- 31. Влияние компонентов питательной среды: - поддерживающие жизнедеятельность: макро- и микроэлементы, витамины, углеродное и азотное питание, агар
- 32. Среды для различных таксономических групп различны Видоспецифичность: соотношение азотсодержащих солей, аминокислот и гормонов Соотношение нитратного (повышение)
- 33. Жидкие питательные среды имеют преимущества для культивирования пыльников Агар целесообразно заменять агарозой Для уплотнения среды можно
- 34. Гормононезависимые растения – гормоны поступают из стенки пыльника или из самой пыльцы – Пасленовые Гормонозависимые растения
- 35. Ячмень: 2,4-Д 1,5 мг/л + кинетин 0,5 мг/л, ИУК 1,0 мг/л + 6-БАП – 1,0 мг/л
- 36. Источник углеродного питания сахароза, глюкоза, мальтоза Сахароза: пшеница 9 %, рис 6 %, кукуруза и ячмень
- 37. Для понимания закономерностей андрогенеза in vitro важно изучение путей развития микроспор, формирования андрогенных структур и регенерации
- 38. А и В ведут к образованию гаплоидных каллусов и эмбриоидов, С – формированию диплоидных и полиплоидных
- 39. При культивировании пыльников необходимо учитывать особенности всех тканей. В образовании эмбриоидов могут принимать участие и соматические
- 40. Доказательства гаплоидной природы: Подсчет числа хлоропластов в замыкающих клетках устьиц подсчет числа хромосом в меристеме корня
- 41. Отбор из первого гибридного поколения дигаплоидов невозможен, растения депрессированы колхицинированием. У дигаплоидов может проявляться «инбредная» депрессия
- 42. Использование при отдаленной гибридизации форм с разным уровнем плоидности (например, культурный картофель × дикий картофель) Использование
- 43. Foroughi-Wehr and Wenzel (1990) рекуррентная селекция ячменя с отбором ценных генотипов на уровне удвоенных гаплоидов Скрещивание
- 44. Получение гаплоидов in vitro Культура пыльников (пшеница, ячмень, кукуруза, озимая рожь, картофель): эмбриогенез в пыльцевых зернах,
- 45. Япония – табак, устойчивый к вилту и с высоким качеством сырья Высокопродуктивные сорта риса, пшеницы, табака,
- 46. Криосохранение растений Жидкий азот, -196 °С Сохранение генофонда и обеспечение селекционера генотипами: Пыльца для проведение гибридизации
- 47. Криосохранение растений Обработка клеток перед замораживанием (маннит, сорбит, пролин, γ-аминомасляная кислота) Применение криопротекторов (ДМСО, глицерин, ПВП,
- 49. Скачать презентацию

Основные селекционные задачи:
Создание нового исходного материала для селекции
Ускорение селекционного процесса
Повышение эффективности
Основные селекционные задачи:
Создание нового исходного материала для селекции
Ускорение селекционного процесса
Повышение эффективности

Генетическая неоднородность клеток в культуре
Соматические мутации
Генетические мутации
Полиплоидия и анеуплоидия
Физические мутагены –
Генетическая неоднородность клеток в культуре
Соматические мутации
Генетические мутации
Полиплоидия и анеуплоидия
Физические мутагены –

Соматическая гибридизация, слияние протопластов (этиленгликоль, высокая концентрация двухвалентных ионов, высокое значение
Соматическая гибридизация, слияние протопластов (этиленгликоль, высокая концентрация двухвалентных ионов, высокое значение

ИСПОЛЬЗОВАНИЕ МЕТОДОВ IN VITRO В СЕЛЕКЦИИ:
Преодоление прогамной несовместимости
Преодоление постгамной несовместимости
Клональное
ИСПОЛЬЗОВАНИЕ МЕТОДОВ IN VITRO В СЕЛЕКЦИИ:
Преодоление прогамной несовместимости
Преодоление постгамной несовместимости
Клональное

Оплодотворение in vitro – преодоление прогамной несовместимости
Причины:
Физиологические (несоответствие во времени
Оплодотворение in vitro – преодоление прогамной несовместимости
Причины:
Физиологические (несоответствие во времени

Модификации метода опыления и оплодотворения in vitro
- прививка столбика с рыльцем
Модификации метода опыления и оплодотворения in vitro
- прививка столбика с рыльцем
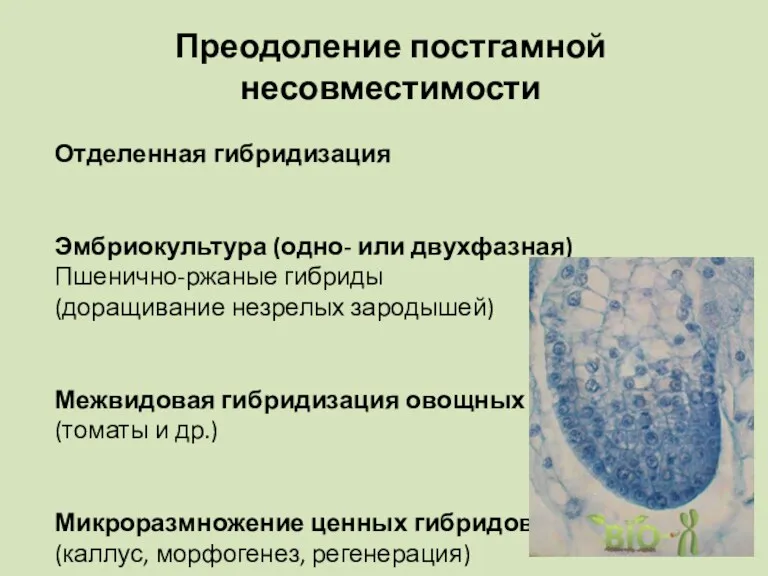
Преодоление постгамной несовместимости
Отделенная гибридизация
Эмбриокультура (одно- или двухфазная)
Пшенично-ржаные гибриды
(доращивание незрелых зародышей)
Межвидовая
Преодоление постгамной несовместимости
Отделенная гибридизация
Эмбриокультура (одно- или двухфазная)
Пшенично-ржаные гибриды
(доращивание незрелых зародышей)
Межвидовая
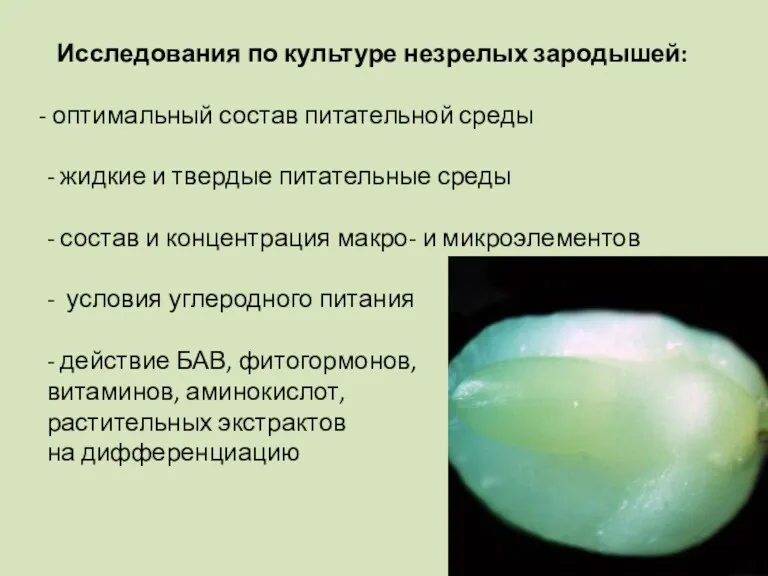
Исследования по культуре незрелых зародышей:
оптимальный состав питательной среды
- жидкие и
Исследования по культуре незрелых зародышей:
оптимальный состав питательной среды
- жидкие и

Показана возможность увеличения выхода пшенично-ржаных гибридов путем доращивания незрелых зародышей, эмбриокультуры;
Показана возможность увеличения выхода пшенично-ржаных гибридов путем доращивания незрелых зародышей, эмбриокультуры;

Клональное микроразмножение отдаленных гибридов
Активация развития меристемы пазушных почек (черенкование стерильных побегов)
Адвентивными
Клональное микроразмножение отдаленных гибридов
Активация развития меристемы пазушных почек (черенкование стерильных побегов)
Адвентивными

Получение гаплоидов in vitro
Быстрый поиск комбинаций - Сокращение срока селекции
Сокращение объемов
Получение гаплоидов in vitro
Быстрый поиск комбинаций - Сокращение срока селекции
Сокращение объемов

Получение гаплоидов in vitro
Стимуляция деления неоплодотворенной яйцеклетки опылением убитой пыльцой
Задержка опыления
Способы
Получение гаплоидов in vitro
Стимуляция деления неоплодотворенной яйцеклетки опылением убитой пыльцой
Задержка опыления
Способы

Партеногенез – из гибридного зародыша, потерявшего отцовские хромосомы, метод гаплопродюссера
H. bulbosum
Партеногенез – из гибридного зародыша, потерявшего отцовские хромосомы, метод гаплопродюссера
H. bulbosum

Сдерживающие факторы использования гаплоидных технологий:
1. Эмпирический подход к поиску генотипов, отзывчивых
Сдерживающие факторы использования гаплоидных технологий:
1. Эмпирический подход к поиску генотипов, отзывчивых

Методы создания гаплоидных растений
отдаленная гибридизация
внутривидовая гибридизация
температурные воздействия
метод
Методы создания гаплоидных растений
отдаленная гибридизация
внутривидовая гибридизация
температурные воздействия
метод

Температурные воздействия
Индукция гаплоидии у кукурузы путем обработки материнских растений через сутки
Температурные воздействия
Индукция гаплоидии у кукурузы путем обработки материнских растений через сутки

Метод близнецов
Близнецы – пары или группы самостоятельных растений, полученных из одного
Метод близнецов
Близнецы – пары или группы самостоятельных растений, полученных из одного

Повышение частоты возникновения гаплоидов при обработке рылец ИУК (до 1: 1550)
Повышение частоты возникновения гаплоидов при обработке рылец ИУК (до 1: 1550)
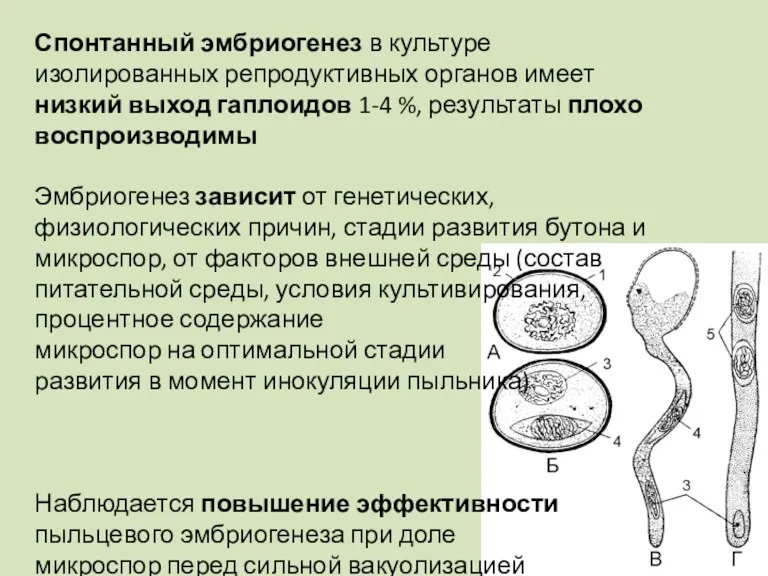
Спонтанный эмбриогенез в культуре изолированных репродуктивных органов имеет низкий выход гаплоидов
Спонтанный эмбриогенез в культуре изолированных репродуктивных органов имеет низкий выход гаплоидов

Важно выявлять корреляцию между морфологическим показателями генеративных органов растения и стадией
Важно выявлять корреляцию между морфологическим показателями генеративных органов растения и стадией

Частота образования гаплоидов зависит от генотипа исходного растения, видовой и сортовой
Частота образования гаплоидов зависит от генотипа исходного растения, видовой и сортовой

N. tabacum +3-5 °C во время первого митоза – возникновение двух
N. tabacum +3-5 °C во время первого митоза – возникновение двух
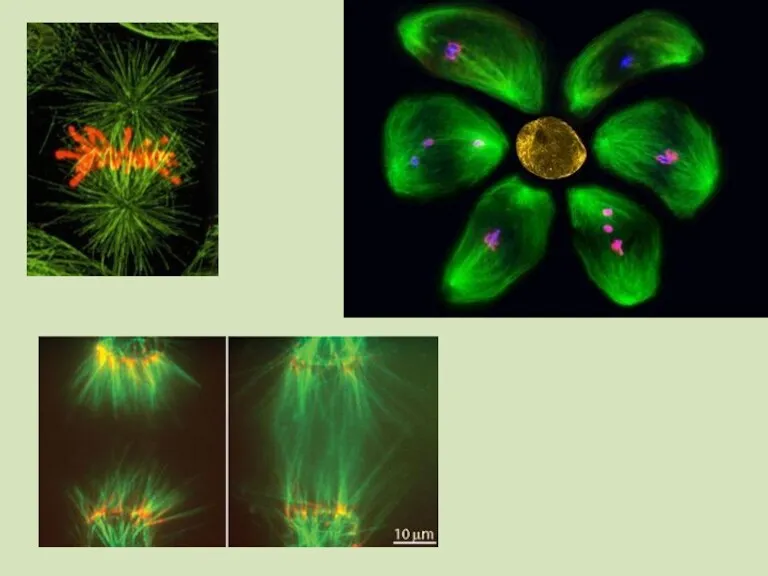

Nicotiana: предварительное инкубирование цветочных почек при 7 °С 7 сут.
Картофель:
Nicotiana: предварительное инкубирование цветочных почек при 7 °С 7 сут.
Картофель:
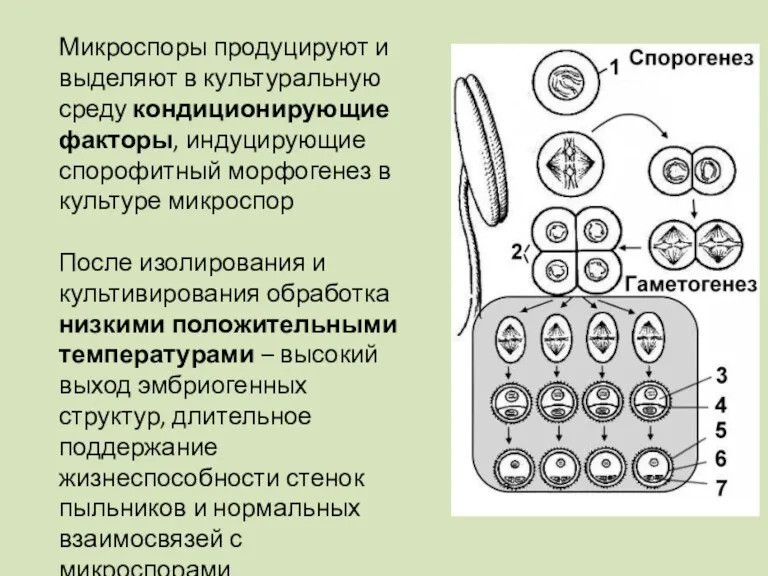
Микроспоры продуцируют и выделяют в культуральную среду кондиционирующие факторы, индуцирующие спорофитный
Микроспоры продуцируют и выделяют в культуральную среду кондиционирующие факторы, индуцирующие спорофитный

Воздействие повышенных температур на изолированные экспланты
Микроспоры рапса +32,5 °С 4 сут.
Воздействие повышенных температур на изолированные экспланты
Микроспоры рапса +32,5 °С 4 сут.

Повышенные температуры вызывают распад микротрубочек, нарушение веретена деления, ненормальное деление ядер
Повышенные температуры вызывают распад микротрубочек, нарушение веретена деления, ненормальное деление ядер

Синтез этилена блокирует нитрат серебра (рис, капуста, пшеница и др.)
Обработка колосьев
Синтез этилена блокирует нитрат серебра (рис, капуста, пшеница и др.)
Обработка колосьев

Влияние компонентов питательной среды:
- поддерживающие жизнедеятельность: макро- и микроэлементы, витамины, углеродное
Влияние компонентов питательной среды:
- поддерживающие жизнедеятельность: макро- и микроэлементы, витамины, углеродное
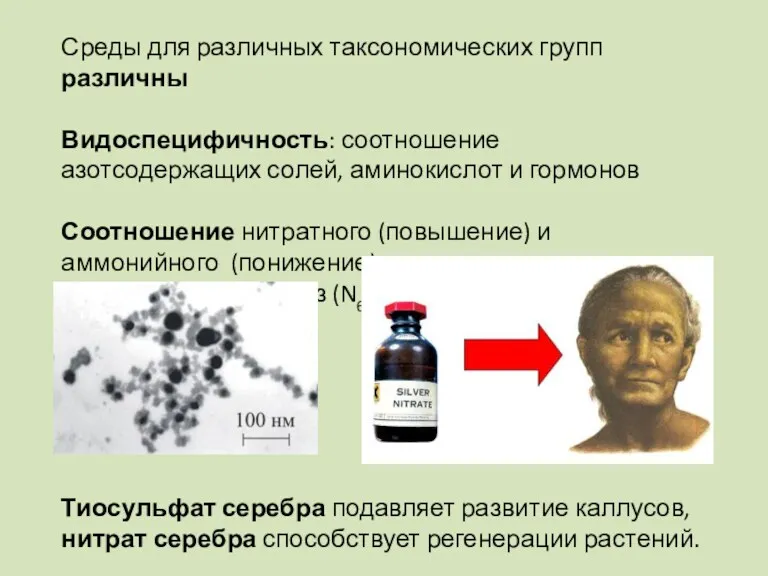
Среды для различных таксономических групп различны
Видоспецифичность: соотношение азотсодержащих солей, аминокислот и
Среды для различных таксономических групп различны
Видоспецифичность: соотношение азотсодержащих солей, аминокислот и

Жидкие питательные среды имеют преимущества для культивирования пыльников
Агар целесообразно заменять агарозой
Для
Жидкие питательные среды имеют преимущества для культивирования пыльников
Агар целесообразно заменять агарозой
Для

Гормононезависимые растения – гормоны поступают из стенки пыльника или из самой
Гормононезависимые растения – гормоны поступают из стенки пыльника или из самой

Ячмень:
2,4-Д 1,5 мг/л + кинетин 0,5 мг/л,
ИУК 1,0 мг/л
Ячмень:
2,4-Д 1,5 мг/л + кинетин 0,5 мг/л,
ИУК 1,0 мг/л

Источник углеродного питания
сахароза, глюкоза, мальтоза
Сахароза: пшеница 9 %, рис 6 %,
Источник углеродного питания
сахароза, глюкоза, мальтоза
Сахароза: пшеница 9 %, рис 6 %,

Для понимания закономерностей андрогенеза in vitro важно изучение путей развития микроспор,
Для понимания закономерностей андрогенеза in vitro важно изучение путей развития микроспор,

А и В ведут к образованию гаплоидных каллусов и эмбриоидов, С
А и В ведут к образованию гаплоидных каллусов и эмбриоидов, С

При культивировании пыльников необходимо учитывать особенности всех тканей. В образовании эмбриоидов
При культивировании пыльников необходимо учитывать особенности всех тканей. В образовании эмбриоидов

Доказательства гаплоидной природы:
Подсчет числа хлоропластов в замыкающих клетках устьиц
подсчет числа
Доказательства гаплоидной природы:
Подсчет числа хлоропластов в замыкающих клетках устьиц
подсчет числа

Отбор из первого гибридного поколения дигаплоидов невозможен, растения депрессированы колхицинированием.
У дигаплоидов
Отбор из первого гибридного поколения дигаплоидов невозможен, растения депрессированы колхицинированием.
У дигаплоидов

Использование при отдаленной гибридизации форм с разным уровнем плоидности (например, культурный
Использование при отдаленной гибридизации форм с разным уровнем плоидности (например, культурный

Foroughi-Wehr and Wenzel (1990)
рекуррентная селекция ячменя с отбором ценных генотипов на
Foroughi-Wehr and Wenzel (1990)
рекуррентная селекция ячменя с отбором ценных генотипов на

Получение гаплоидов in vitro
Культура пыльников (пшеница, ячмень, кукуруза, озимая рожь, картофель):
Получение гаплоидов in vitro
Культура пыльников (пшеница, ячмень, кукуруза, озимая рожь, картофель):

Япония – табак, устойчивый к вилту и с высоким качеством сырья
Высокопродуктивные
Япония – табак, устойчивый к вилту и с высоким качеством сырья
Высокопродуктивные

Криосохранение растений
Жидкий азот, -196 °С
Сохранение генофонда и обеспечение
селекционера генотипами:
Пыльца для
Криосохранение растений
Жидкий азот, -196 °С
Сохранение генофонда и обеспечение
селекционера генотипами:
Пыльца для

Криосохранение растений
Обработка клеток перед замораживанием (маннит, сорбит, пролин, γ-аминомасляная кислота)
Применение криопротекторов
Криосохранение растений
Обработка клеток перед замораживанием (маннит, сорбит, пролин, γ-аминомасляная кислота)
Применение криопротекторов
 Проблема ГМО-продуктів – проблема вибору
Проблема ГМО-продуктів – проблема вибору Нервная система: нервы, нервные узлы, спинной мозг. Тема 14
Нервная система: нервы, нервные узлы, спинной мозг. Тема 14 Презентация Организм растения - единое целое
Презентация Организм растения - единое целое Мотивация. Биологическая мотивация. Общие свойства различных видов мотивации. Мотивация как доминанта
Мотивация. Биологическая мотивация. Общие свойства различных видов мотивации. Мотивация как доминанта ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Рыбы. Приспособленность к обитанию в водной среде . 7 класс (В семи частях)
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Рыбы. Приспособленность к обитанию в водной среде . 7 класс (В семи частях) Животный организм и его особенности
Животный организм и его особенности Водоросли. Общая характеристика. Классификация
Водоросли. Общая характеристика. Классификация Карповые монтажи
Карповые монтажи Эндокринная система
Эндокринная система Насекомые
Насекомые Бактериологическая разведка и индикация бактериологического (биологического) оружия
Бактериологическая разведка и индикация бактериологического (биологического) оружия Тип моллюски
Тип моллюски Красная книга Тверской области
Красная книга Тверской области Проект Геном человека - занятие по биохимии
Проект Геном человека - занятие по биохимии Критерии вида (биологические задачи)
Критерии вида (биологические задачи) Мейоз. Фазы мейоза
Мейоз. Фазы мейоза Растения смешанного леса
Растения смешанного леса Итоги ОГЭ по биологии 2015 г.
Итоги ОГЭ по биологии 2015 г. Первый и второй законы Менделя
Первый и второй законы Менделя Технология выращивания комнатных растений
Технология выращивания комнатных растений Водорастворимые витамины
Водорастворимые витамины Розмноження птахів
Розмноження птахів Спектр белков и ферментов ротовой жидкости
Спектр белков и ферментов ротовой жидкости ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Пути и направления эволюции. 11 класс (В двух частях)
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Пути и направления эволюции. 11 класс (В двух частях) Выделение. 6 класс
Выделение. 6 класс Биохимия витаминов 2. (Лекция 26)
Биохимия витаминов 2. (Лекция 26) Ферменти
Ферменти Выявление ароморфозов и идиоадаптаций у растений
Выявление ароморфозов и идиоадаптаций у растений