- Ферменти

Содержание
- 2. Усі без винятку хімічні процеси в клітині каталізуються спеціальними біологічними каталізаторами, що називаються ферментами або ензимами.
- 3. Загальні властивості каталізаторів: Збільшують швидкість реакції Знижують енергію активації Після реакції виділяються в незмінному вигляді Прискорюють
- 4. Відмінності ферментів від небіологічних каталізаторів: Висока ефективність.У присутності ферментів хімічні реакції протікають набагато швидше, ніж при
- 5. Структура ферментів Холофермент Апофермент Кофактор Кофермент (нековалентно зв'язаний); Простетична група (ковалентно) Ферменти є прості і складні.
- 6. Кофермент А (коензим А), Для його позначення використовується скорочена форма — КоА—SН, Коензим А складається із
- 7. Коферменти У вигляді кофактора вітамін N (ліпоева кислота) входить до складу поліферментних комплексів, які каталізують окисне
- 8. Структура коферментів похідних водорозчинних вітамінів – НАД+ Нікотинамідні коферменти — це найбільш поширена і численна група
- 9. Простетичні групи – ФМН, ФAД Ферменти, які містять флавінові простетичні групи беруть участь в окисно-відновних реакціях
- 11. Специфічність дії ферментів Кожен фермент каталізує певну хімічну реакцію або тип хімічної реакції. Апофермент зумовлює специфічність
- 12. Види специфічності: Відносна (каталізує групу подібних реакцій з певним типом зв′язку (естерази)) Абсолютная (каталізує перетворення тільки
- 13. АКТИВНІСТЬ ФЕРМЕНТІВ Активність фермента характеризується швидкістю хімічних реакцій, виражається в Каталах (кат): 1 кат — це
- 14. Рекомендовано вимірювати активність фермента при температурі 25°С, оптимумі рН та концентрації субстрата, що перевищує концентрацію насичення.
- 15. В харчовій промисловості активність називають здатністю: Мальтазна здатність (М3) - це кількість ферменту, яка розщеплює 1
- 16. Мультиферменти Це надмолекулярні комплекси з різних ферментів та коферментів просторово зближені та певним чином організовані, що
- 17. Ізоферменти. Майже всі ферменти існують в декількох формах. Ізоферменти – це генетично детерміновані форми одного й
- 18. Ізоферментні форми лактатдегідрогенази За зміною вмісту ізоферментів в сировотці крові можна судити як про місце патологічного
- 19. Механізм дії ферментів. Причиною всіх цих унікальних властивостей ферментів є їхня просторова будова. Усі ферменти являють
- 20. Ферменти мають дуже високою специфічністю. Фішер (Fischer) в 1890 р. висловив припущення, що ця специфічність обумовлюється
- 21. Стадії ферментативного каталізу Взаємодія ферменту з субстратом відбувається за законом комплементарності конфігурації активних центрів, конформації та
- 22. Стадії ферментативного каталізу Поняття про “енергію активації” та “енергетичний бар’єр” Ферменти прискорюють хімічну реакцію за рахунок
- 23. Теорія ферментативного каталізу S - початковий субстрат; Р - продукт; ΔЕнф - енергія активації неферментативної реакції;
- 24. ЕФЕКТОРИ Інгібітори – (білкові осаджувачі — ССl3СООН, танін, концентровані кислоти, солі важких металів та ін.). Активатори
- 25. Види інгібування Інгібірування буває оборотним і необоротним. Оборотне – це коли інгібітор утворює з ферментом слабкий
- 26. Інгібітор має структуру, подібну до субстрату і конкурує з ним
- 28. Кінетика ферментативних реакцій Розділ ензимології, який вивчає швидкість ферментативних реакцій та їх залежність від дії різних
- 29. Концентрація субстрату Фактори, що впливають на активність ферментів За дуже низьких концентрацій субстрату швидкість реакції є
- 30. З рівняння Міхаеліса–Ментен витікає, що: за високої концентрації субстрату та низькому значенні Ks швидкість реакції є
- 31. pH Залежність ферментативної активності від рН середовища. Більшість ферментів найактивніші в діапазоні рН 6-8. Екстремальні значення
- 32. Температура Термолабільність – відповідно до своєї білкової природи ферменти є термочутливими. Оптимум 37-45°С. Підвищення температури на
- 33. Номенклатура ферментів Способи номенклатури: Тривіальна назва (пепсин, трипсин и т.д.) Рабоча назва: Назва субстрату+ тип реакції
- 34. Класифікація ферментів 1. Оксидоредуктази. Окислювально-відновні процеси в клітині можуть здійснюватися трьома шляхами: переносом іона водню, приєднанням
- 35. Характеристика окремих класів ферментів 1 КЛАС Оксидоредуктази каталізують окисно-відновні процеси -(дегідрогенази, оксигенази, оксидази, пероксидази, гідроксилази, та
- 36. Згідно з міжнородною класифікацією оксидоредуктаз виділяють 22 підкласи: КФ 1.1 — ферменти, що взаємодіють з CH—OH—групою
- 37. Небілковою частиною є нікотинамідаденіндинуклеотид (НАД) або нікотинамідаденіндинуклеотидфосфат (НАДФ). 1.1.1 Піридинові дегідрогенази Реакції що відбуваються за участі
- 38. 1.1.2 Флавінові дегідрогенази Небілковою частиною є флавінмононуклеотид (ФМН) або флавінаденіндинуклеотид (ФАД). Флавінові дегідрогенази містять похідні вітаміну
- 39. 1.2 Оксигенази - ферменти, за допомогою яких відбувається пряме приєднання одного або двох атомів кисню до
- 40. 1.3 Оксидази – коли акцептором водню служить O2повітря. Ліпоксигеназа (ліпоксидаза) широко розповсюджена у рослинах, каталізує окислення
- 41. 1.4 Гідрогенази – ферменти, що каталізують оборотне окислення водню за участю ферредоксину, НАД+ або цитохрому а3.
- 42. 2 клас Трансферази: каталізують перенесення хімічних групи атомів, від яких і беруть назву (метил-, сульфо-, аміно-,
- 43. Глікозилтрансферази каталізують перенесення залишків вуглеводів До цього підкласу належать ферменти фосфорилази, які каталізують перетворення крохмалю чи
- 44. Ацилтрансферази беруть участь в перенесенні залишків органічних кислот, головним чином залишку оцтової кислоти - ацетилу (СН3СО-).
- 45. Підрозділяється на 13 підкласів в залежності від типу зв'язку, що гідролізується. КФ 3.1 складноефірний зв'язок (естерази:
- 46. Найбільше значення мають естерази, глікозидази, пептидгідролази, амідази: 3.1. Естерази здійснюють гідроліз та синтез складних ефірів. R-CO-O-R1
- 47. Глікозидази (карбогідрази) розщеплюють глікозидні зв’язки, здійснюючи гідроліз вуглеводів та інших глікозидів. R-O-R1 + HOH ↔ ROH
- 48. 2 а Олігази β-фруктозидаза (сахараза, інвертаза) розщеплює β-фруктозидний зв’язок в сахарозі та рафінозі: Препарат інвертази (інвертин)
- 49. α-галактозидаза (мелібіаза) розщеплює α-галактозидний зв’язок в мелібіозі (D-Гал-α(1 → 6)-D-Глю) та рафінозі. Олігази, окрім гідролазної активності,
- 50. - амілази - група ферментів, що гідролізують крохмаль. Встановлена наявність трьох амілаз. Фермент α-амілаза (тваринна амілаза,
- 51. Пептидгідролази (протеази) - це ферменти, які гідролізують пептиди і білки, розщеплюючи в них пептидні зв'язки. Вони
- 52. амінопептидази – розщепляють пептидний зв’язок, що знаходиться поряд із вільною аміногрупою. Для дії амінопептидаз необхідна наявність
- 53. Амідази – каталізують гідроліз амідів (уреаза, аргіназа, аспарагіназа, глютаміназа). уреаза – розщеплює сечовину на амоніак та
- 54. 4 клас ЛІАЗИ Ліази каталізують реакції негідролітичного розщеплення без участі води. (декарбоксилази, альдолази, тощо.) Розчеплюють С-С
- 55. Згідно з міжнародною класифікацією ферментів ліази віднесені до класу (КФ 4). Клас, в свою чергу, підрозділяється
- 56. піруватдекарбоксилаза альдолаза енолаза 2-Фосфогліцеринова кислота Фосфоенол ПВК ЛІАЗИ Приклади:
- 57. Ізомерази — ферменти, що каталізують структурні або просторові зміни в молекулі субстрату. КФ 5.1 ферменти, що
- 58. 5.1. Рацемази – наприклад бактеріальна рацемаза перетворює L-амінокислоти на D-амінокислоти. (аланінрацемаза, метионінрацемаза, гідроксипролінрацемаза, лактатрацемаза та ін.)
- 59. глюкозофосфат-ізмераза каталізує взаємне перетворення глюкопіранозо-6-фосфату та фруктофуранозо-6-фосфату. (рибозофосфатізомераза, ксилозоізомераза, глюкозамінфосфат-ізомераза, еноїл-КоАізомераза та ін.), 5.4. Фосфогліцератмутаза перетворює
- 60. 6 клас Лігази (від лат. ligāre — «зшивати», «зв'язувати») — клас ферментів (КФ 6), здатних каталізувати
- 61. У класифікації міжнародної комісії з ферментів, лігази класифікуються як КФ 6 та поділяються на 6 підгруп:
- 62. 6.1. Лігази, що утворюють C–O-зв’язок – ферменти, що каталізують активацію та приєднання залишків амінокислот до т-РНК
- 63. Лігази 6.3. Лігази, що утворюють C–N-зв’язок. глютамінсинтетаза – каталізує реакцію синтезу глютаміну з глютамінової кислоти та
- 64. Ферменти, які використовуються в промисловості отримують з сировини тваринного, рослинного або мікробного походження. Різні способи виробництва
- 65. Застосування в промисловості Хлібопекарська промисловість амілази протеази ліпоксидази Для відновлення смакових якостей і запаху У виробництві
- 66. Целюлазу, пектиназу і нарингіназу (виробництво цитрусових соків, прибирає гіркий смак) застосовують в різних процесах при переробці
- 67. Панкреатин (ренін, пепсин, трипсин, амілаза і ліпаза) - для лікування диспепсії та шлунково-кишкових розладів. Хімотрипсин -
- 68. ІМОБІЛІЗОВАНІ ФЕРМЕНТИ Ферменти, що закріплені на полімерному носію. Фізична або хімічна мобілізація. Переваги: Підвищення стабільності фермента
- 70. Скачать презентацию

Усі без винятку хімічні процеси в клітині каталізуються спеціальними біологічними каталізаторами,
Усі без винятку хімічні процеси в клітині каталізуються спеціальними біологічними каталізаторами,

Загальні властивості каталізаторів:
Збільшують швидкість реакції
Знижують енергію активації
Після реакції виділяються в незмінному
Загальні властивості каталізаторів:
Збільшують швидкість реакції
Знижують енергію активації
Після реакції виділяються в незмінному

Відмінності ферментів від небіологічних каталізаторів:
Висока ефективність.У присутності ферментів хімічні реакції протікають
Відмінності ферментів від небіологічних каталізаторів:
Висока ефективність.У присутності ферментів хімічні реакції протікають
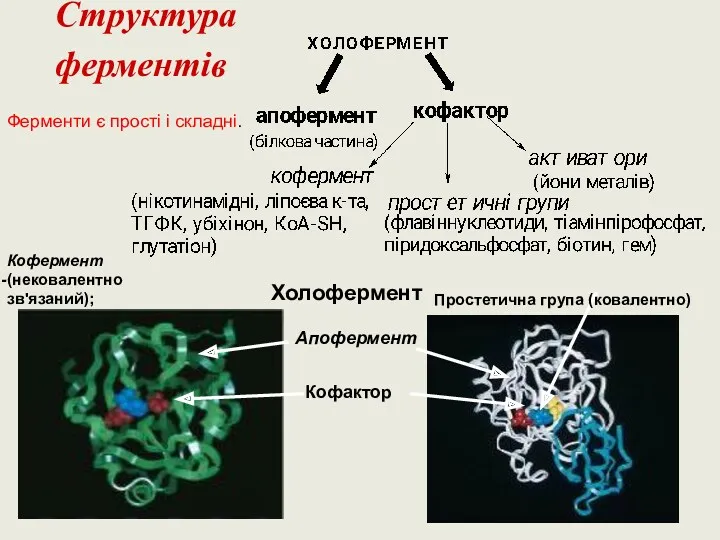
Структура
ферментів
Холофермент
Апофермент
Кофактор
Кофермент
(нековалентно зв'язаний);
Простетична група (ковалентно)
Ферменти є прості і складні.
Структура
ферментів
Холофермент
Апофермент
Кофактор
Кофермент
(нековалентно зв'язаний);
Простетична група (ковалентно)
Ферменти є прості і складні.
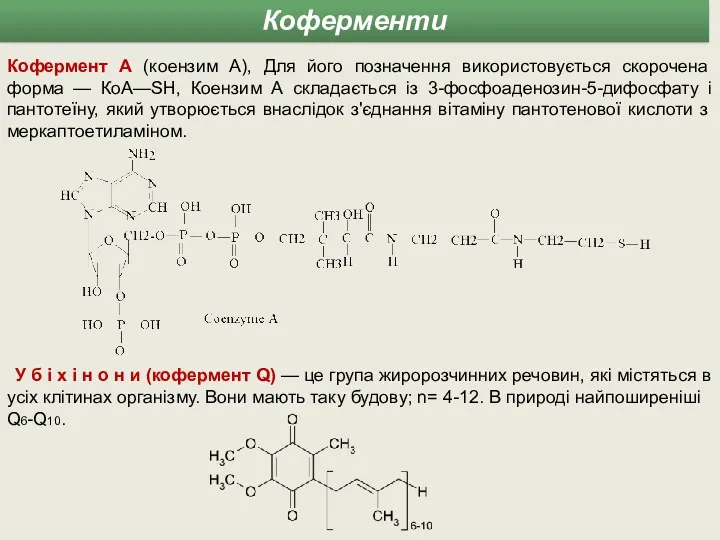
Кофермент А (коензим А), Для його позначення використовується скорочена форма —
Кофермент А (коензим А), Для його позначення використовується скорочена форма —
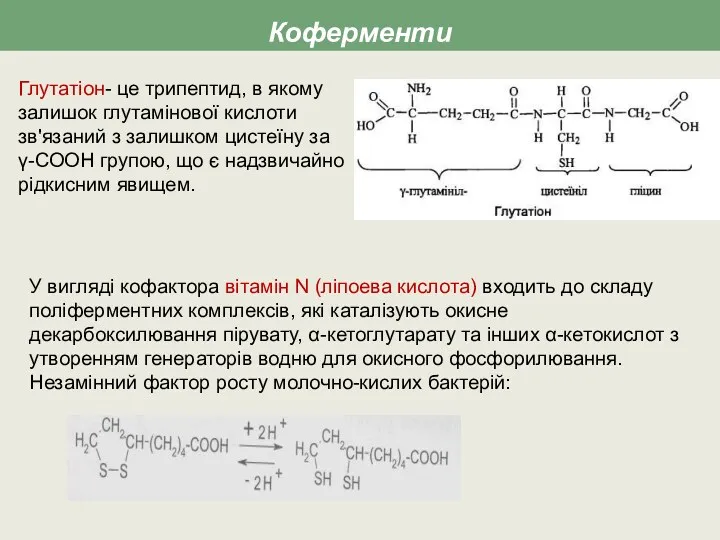
Коферменти
У вигляді кофактора вітамін N (ліпоева кислота) входить до складу поліферментних
Коферменти
У вигляді кофактора вітамін N (ліпоева кислота) входить до складу поліферментних
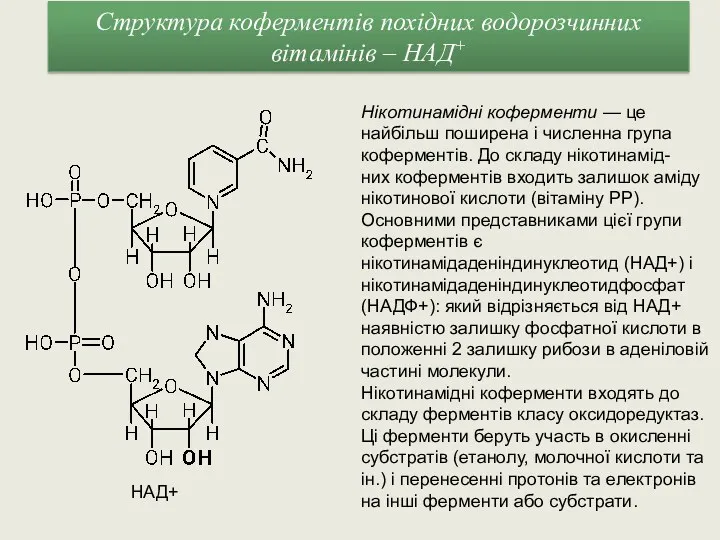
Структура коферментів похідних водорозчинних вітамінів – НАД+
Нікотинамідні коферменти — це найбільш
Структура коферментів похідних водорозчинних вітамінів – НАД+
Нікотинамідні коферменти — це найбільш
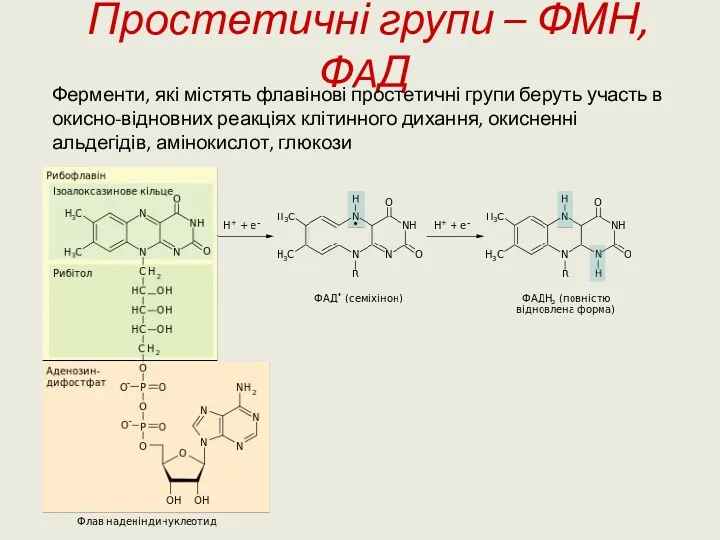
Простетичні групи – ФМН, ФAД
Ферменти, які містять флавінові простетичні групи
Простетичні групи – ФМН, ФAД
Ферменти, які містять флавінові простетичні групи

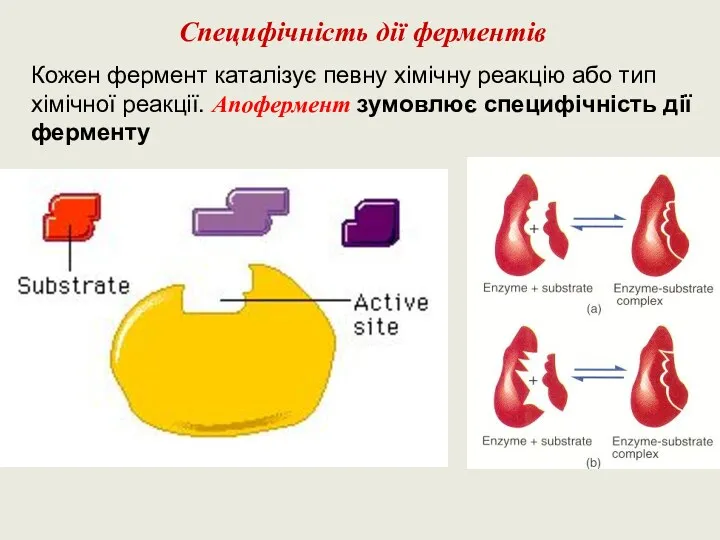
Специфічність дії ферментів
Кожен фермент каталізує певну хімічну реакцію або тип
Специфічність дії ферментів
Кожен фермент каталізує певну хімічну реакцію або тип
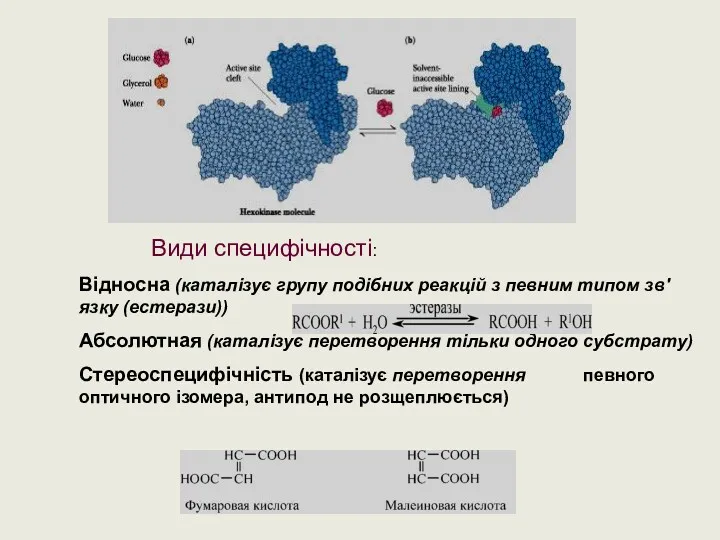
Види специфічності:
Відносна (каталізує групу подібних реакцій з певним типом зв′язку (естерази))
Види специфічності:
Відносна (каталізує групу подібних реакцій з певним типом зв′язку (естерази))

АКТИВНІСТЬ ФЕРМЕНТІВ
Активність фермента характеризується швидкістю хімічних реакцій, виражається в Каталах (кат):
АКТИВНІСТЬ ФЕРМЕНТІВ
Активність фермента характеризується швидкістю хімічних реакцій, виражається в Каталах (кат):

Рекомендовано вимірювати активність фермента при температурі 25°С, оптимумі рН та концентрації
Рекомендовано вимірювати активність фермента при температурі 25°С, оптимумі рН та концентрації

В харчовій промисловості
активність називають здатністю:
Мальтазна здатність (М3) - це кількість
В харчовій промисловості
активність називають здатністю:
Мальтазна здатність (М3) - це кількість

Мультиферменти
Це надмолекулярні комплекси з різних ферментів та коферментів просторово зближені та
Мультиферменти
Це надмолекулярні комплекси з різних ферментів та коферментів просторово зближені та
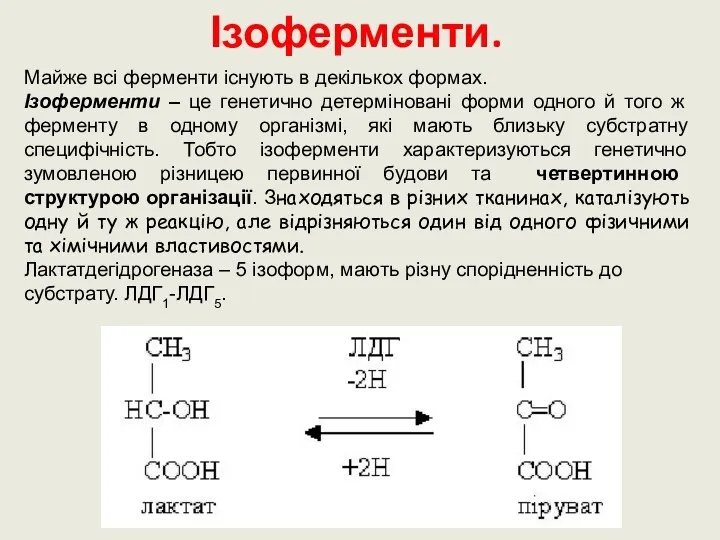
Ізоферменти.
Майже всі ферменти існують в декількох формах.
Ізоферменти – це генетично детерміновані
Ізоферменти.
Майже всі ферменти існують в декількох формах.
Ізоферменти – це генетично детерміновані

Ізоферментні форми лактатдегідрогенази
За зміною вмісту ізоферментів в сировотці крові можна судити
Ізоферментні форми лактатдегідрогенази
За зміною вмісту ізоферментів в сировотці крові можна судити

Механізм дії ферментів.
Причиною всіх цих унікальних властивостей ферментів є їхня просторова
Механізм дії ферментів.
Причиною всіх цих унікальних властивостей ферментів є їхня просторова
Ферменти мають дуже високою специфічністю. Фішер (Fischer) в 1890 р. висловив припущення,
Ферменти мають дуже високою специфічністю. Фішер (Fischer) в 1890 р. висловив припущення,

Стадії ферментативного каталізу
Взаємодія ферменту з субстратом відбувається за законом комплементарності конфігурації
Стадії ферментативного каталізу
Взаємодія ферменту з субстратом відбувається за законом комплементарності конфігурації

Стадії ферментативного каталізу
Поняття про “енергію активації” та “енергетичний бар’єр”
Ферменти прискорюють хімічну
Стадії ферментативного каталізу
Поняття про “енергію активації” та “енергетичний бар’єр”
Ферменти прискорюють хімічну
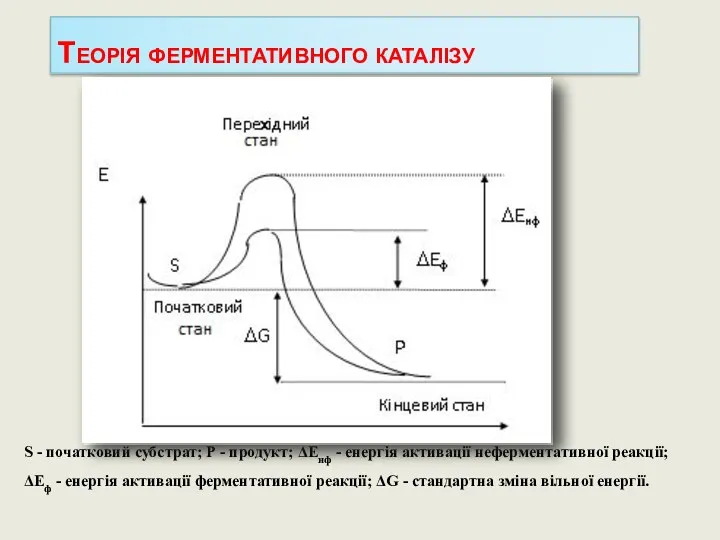
Теорія ферментативного каталізу
S - початковий субстрат; Р - продукт; ΔЕнф -
Теорія ферментативного каталізу
S - початковий субстрат; Р - продукт; ΔЕнф -

ЕФЕКТОРИ
Інгібітори – (білкові осаджувачі — ССl3СООН, танін, концентровані кислоти, солі важких
ЕФЕКТОРИ
Інгібітори – (білкові осаджувачі — ССl3СООН, танін, концентровані кислоти, солі важких
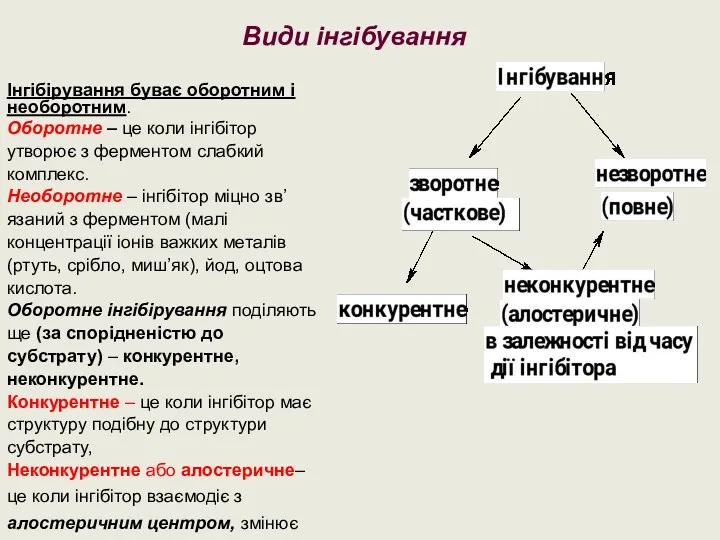
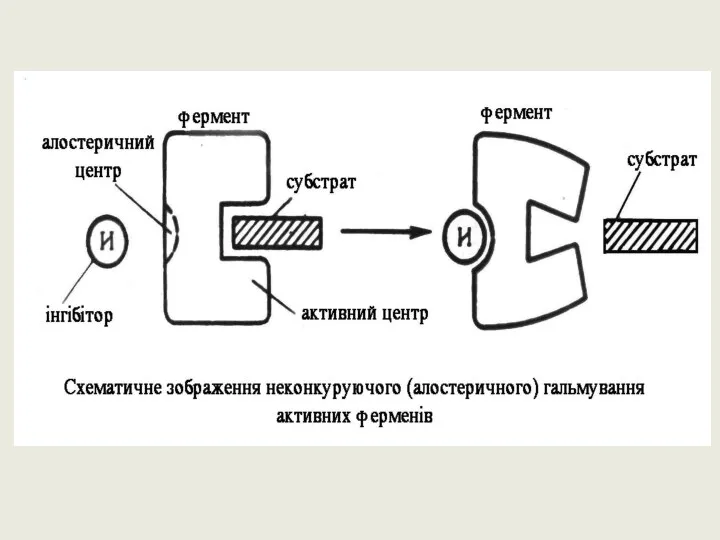
Види інгібування
Інгібірування буває оборотним і необоротним.
Оборотне – це коли інгібітор утворює
Види інгібування
Інгібірування буває оборотним і необоротним.
Оборотне – це коли інгібітор утворює
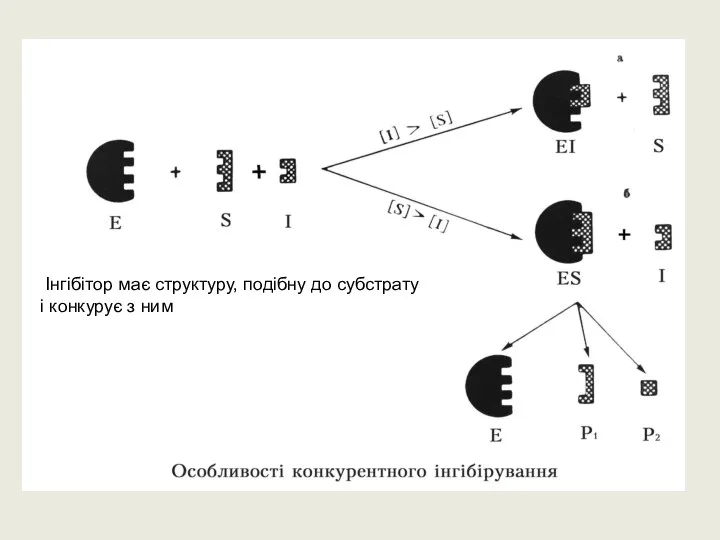
Інгібітор має структуру, подібну до субстрату і конкурує з ним
Інгібітор має структуру, подібну до субстрату і конкурує з ним

Кінетика ферментативних реакцій
Розділ ензимології, який вивчає швидкість ферментативних реакцій та їх
Кінетика ферментативних реакцій
Розділ ензимології, який вивчає швидкість ферментативних реакцій та їх

Концентрація субстрату
Фактори, що впливають на активність ферментів
За дуже низьких концентрацій субстрату
Концентрація субстрату
Фактори, що впливають на активність ферментів
За дуже низьких концентрацій субстрату

З рівняння Міхаеліса–Ментен витікає, що:
за високої концентрації субстрату та низькому значенні
З рівняння Міхаеліса–Ментен витікає, що:
за високої концентрації субстрату та низькому значенні
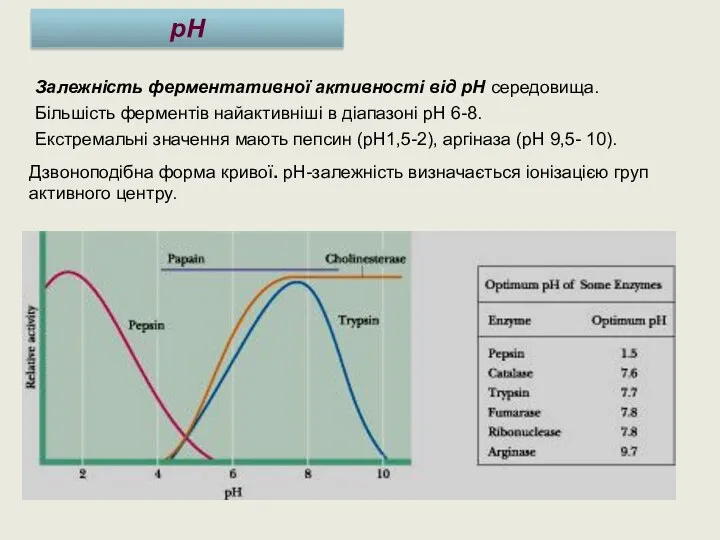
pH
Залежність ферментативної активності від рН середовища.
Більшість ферментів найактивніші в
pH
Залежність ферментативної активності від рН середовища.
Більшість ферментів найактивніші в
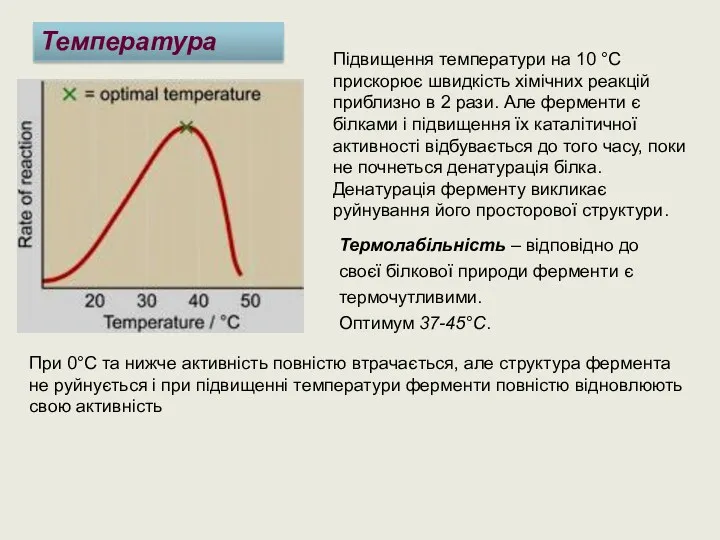
Температура
Термолабільність – відповідно до своєї білкової природи ферменти є термочутливими.
Оптимум
Температура
Термолабільність – відповідно до своєї білкової природи ферменти є термочутливими.
Оптимум

Номенклатура ферментів
Способи номенклатури:
Тривіальна назва (пепсин, трипсин и т.д.)
Рабоча назва:
Назва субстрату+
Номенклатура ферментів
Способи номенклатури:
Тривіальна назва (пепсин, трипсин и т.д.)
Рабоча назва:
Назва субстрату+

Класифікація ферментів
1. Оксидоредуктази. Окислювально-відновні процеси в клітині можуть здійснюватися трьома шляхами:
Класифікація ферментів
1. Оксидоредуктази. Окислювально-відновні процеси в клітині можуть здійснюватися трьома шляхами:

Характеристика окремих класів ферментів
1 КЛАС
Оксидоредуктази каталізують окисно-відновні процеси -(дегідрогенази, оксигенази, оксидази,
Характеристика окремих класів ферментів
1 КЛАС
Оксидоредуктази каталізують окисно-відновні процеси -(дегідрогенази, оксигенази, оксидази,

Згідно з міжнородною класифікацією оксидоредуктаз виділяють 22 підкласи:
КФ 1.1 — ферменти,
Згідно з міжнородною класифікацією оксидоредуктаз виділяють 22 підкласи:
КФ 1.1 — ферменти,
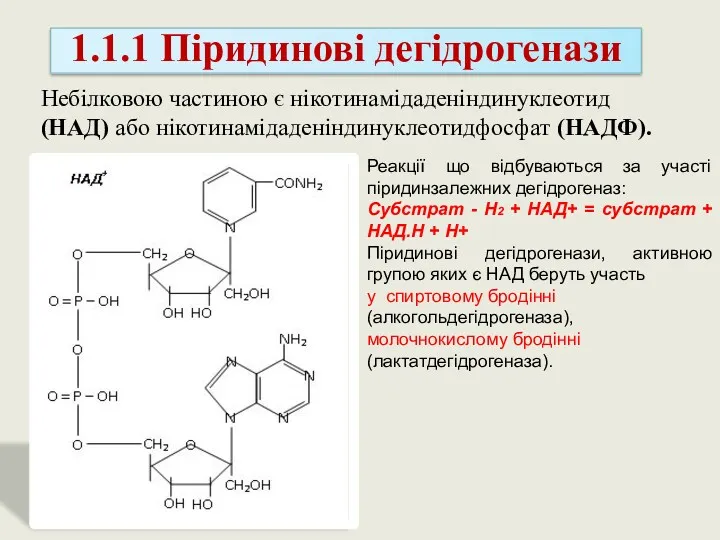
Небілковою частиною є нікотинамідаденіндинуклеотид (НАД) або нікотинамідаденіндинуклеотидфосфат (НАДФ).
1.1.1 Піридинові дегідрогенази
Реакції що
Небілковою частиною є нікотинамідаденіндинуклеотид (НАД) або нікотинамідаденіндинуклеотидфосфат (НАДФ).
1.1.1 Піридинові дегідрогенази
Реакції що
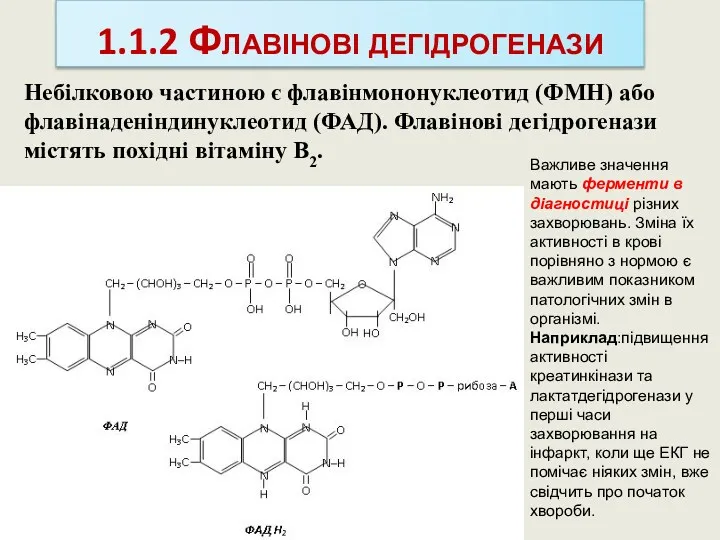
1.1.2 Флавінові дегідрогенази
Небілковою частиною є флавінмононуклеотид (ФМН) або флавінаденіндинуклеотид (ФАД).
1.1.2 Флавінові дегідрогенази
Небілковою частиною є флавінмононуклеотид (ФМН) або флавінаденіндинуклеотид (ФАД).

1.2 Оксигенази - ферменти, за допомогою яких відбувається пряме приєднання одного
1.2 Оксигенази - ферменти, за допомогою яких відбувається пряме приєднання одного

1.3 Оксидази – коли акцептором водню служить O2повітря.
Ліпоксигеназа (ліпоксидаза) широко розповсюджена
1.3 Оксидази – коли акцептором водню служить O2повітря.
Ліпоксигеназа (ліпоксидаза) широко розповсюджена

1.4 Гідрогенази – ферменти, що каталізують оборотне окислення водню за участю
1.4 Гідрогенази – ферменти, що каталізують оборотне окислення водню за участю

2 клас
Трансферази: каталізують перенесення хімічних групи атомів, від яких і беруть
2 клас
Трансферази: каталізують перенесення хімічних групи атомів, від яких і беруть
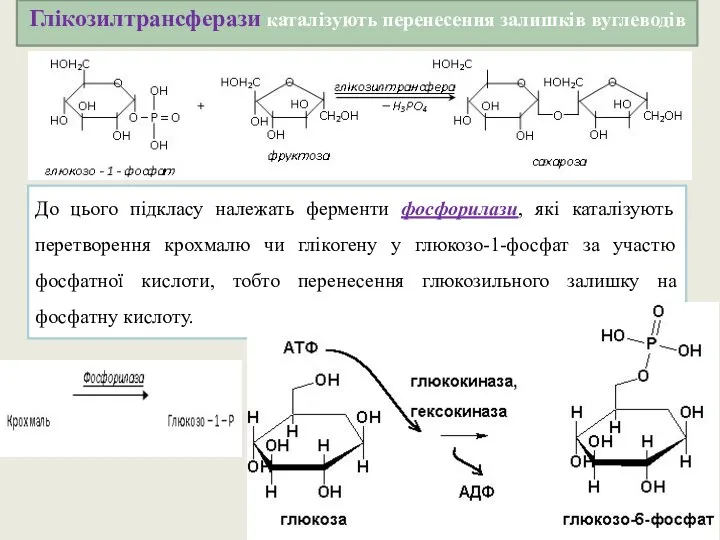
Глікозилтрансферази каталізують перенесення залишків вуглеводів
До цього підкласу належать ферменти фосфорилази,
Глікозилтрансферази каталізують перенесення залишків вуглеводів
До цього підкласу належать ферменти фосфорилази,

Ацилтрансферази беруть участь в перенесенні залишків органічних кислот, головним чином залишку
Ацилтрансферази беруть участь в перенесенні залишків органічних кислот, головним чином залишку

Підрозділяється на 13 підкласів в залежності від типу зв'язку, що гідролізується.
КФ
Підрозділяється на 13 підкласів в залежності від типу зв'язку, що гідролізується.
КФ

Найбільше значення мають естерази, глікозидази, пептидгідролази, амідази:
3.1. Естерази здійснюють
Найбільше значення мають естерази, глікозидази, пептидгідролази, амідази:
3.1. Естерази здійснюють

Глікозидази (карбогідрази) розщеплюють глікозидні зв’язки, здійснюючи гідроліз вуглеводів та інших
Глікозидази (карбогідрази) розщеплюють глікозидні зв’язки, здійснюючи гідроліз вуглеводів та інших

2 а Олігази
β-фруктозидаза (сахараза, інвертаза) розщеплює β-фруктозидний зв’язок в сахарозі
2 а Олігази
β-фруктозидаза (сахараза, інвертаза) розщеплює β-фруктозидний зв’язок в сахарозі

α-галактозидаза (мелібіаза) розщеплює α-галактозидний зв’язок в мелібіозі (D-Гал-α(1 → 6)-D-Глю) та
α-галактозидаза (мелібіаза) розщеплює α-галактозидний зв’язок в мелібіозі (D-Гал-α(1 → 6)-D-Глю) та

- амілази - група ферментів, що гідролізують крохмаль. Встановлена наявність трьох
- амілази - група ферментів, що гідролізують крохмаль. Встановлена наявність трьох

Пептидгідролази (протеази) - це ферменти, які гідролізують пептиди і білки,
Пептидгідролази (протеази) - це ферменти, які гідролізують пептиди і білки,

амінопептидази – розщепляють пептидний зв’язок, що знаходиться поряд із вільною аміногрупою.
амінопептидази – розщепляють пептидний зв’язок, що знаходиться поряд із вільною аміногрупою.

Амідази – каталізують гідроліз амідів (уреаза, аргіназа, аспарагіназа, глютаміназа).
уреаза –
Амідази – каталізують гідроліз амідів (уреаза, аргіназа, аспарагіназа, глютаміназа).
уреаза –

4 клас ЛІАЗИ
Ліази каталізують реакції негідролітичного розщеплення без участі води. (декарбоксилази,
4 клас ЛІАЗИ
Ліази каталізують реакції негідролітичного розщеплення без участі води. (декарбоксилази,

Згідно з міжнародною класифікацією ферментів ліази віднесені до класу (КФ 4).
Згідно з міжнародною класифікацією ферментів ліази віднесені до класу (КФ 4).
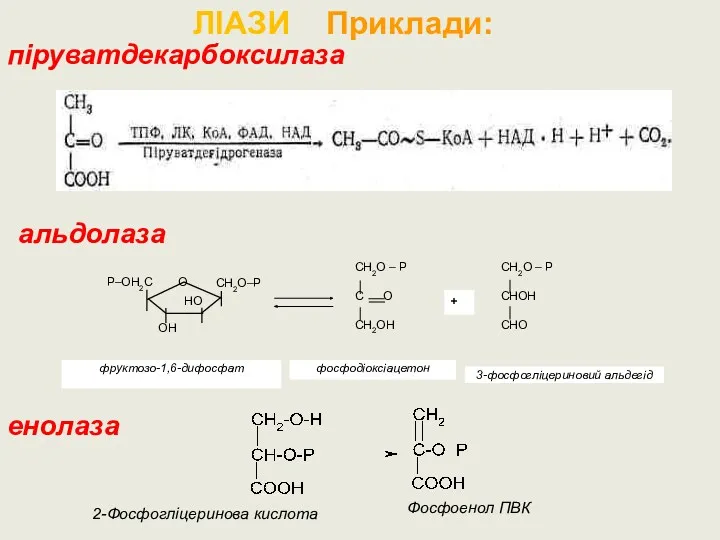
піруватдекарбоксилаза
альдолаза
енолаза
2-Фосфогліцеринова кислота
Фосфоенол ПВК
ЛІАЗИ Приклади:
піруватдекарбоксилаза
альдолаза
енолаза
2-Фосфогліцеринова кислота
Фосфоенол ПВК
ЛІАЗИ Приклади:

Ізомерази — ферменти, що каталізують структурні або просторові зміни в молекулі
Ізомерази — ферменти, що каталізують структурні або просторові зміни в молекулі
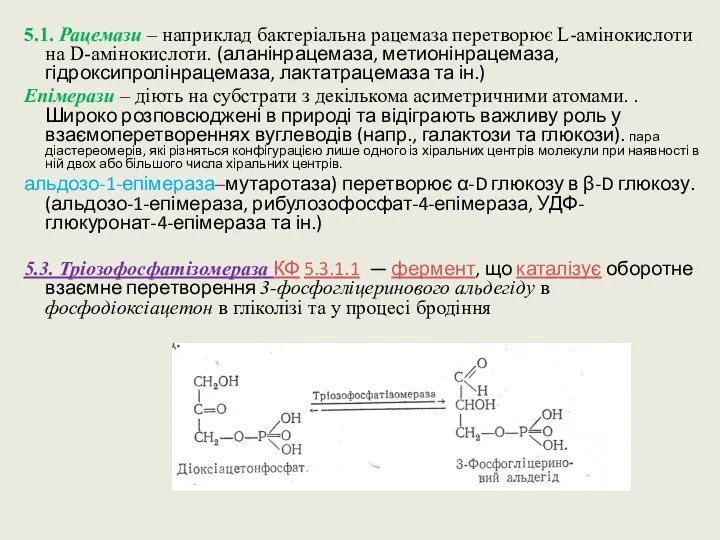
5.1. Рацемази – наприклад бактеріальна рацемаза перетворює L-амінокислоти на D-амінокислоти. (аланінрацемаза,
5.1. Рацемази – наприклад бактеріальна рацемаза перетворює L-амінокислоти на D-амінокислоти. (аланінрацемаза,
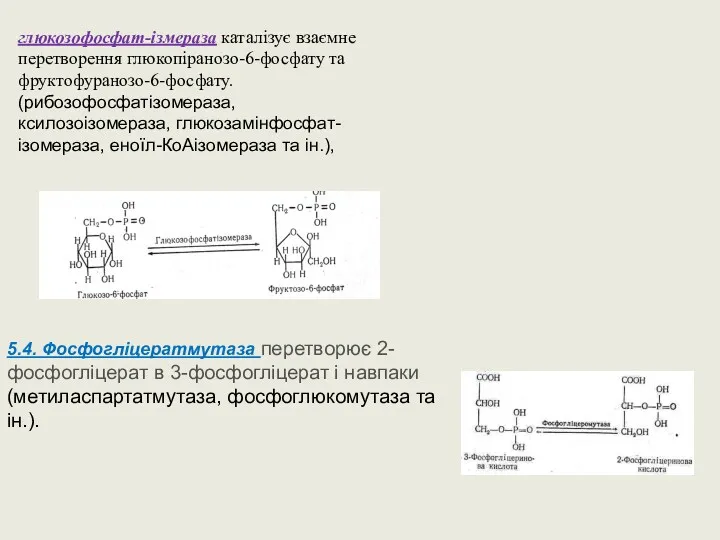
глюкозофосфат-ізмераза каталізує взаємне перетворення глюкопіранозо-6-фосфату та фруктофуранозо-6-фосфату. (рибозофосфатізомераза, ксилозоізомераза, глюкозамінфосфат-ізомераза, еноїл-КоАізомераза
глюкозофосфат-ізмераза каталізує взаємне перетворення глюкопіранозо-6-фосфату та фруктофуранозо-6-фосфату. (рибозофосфатізомераза, ксилозоізомераза, глюкозамінфосфат-ізомераза, еноїл-КоАізомераза

6 клас
Лігази (від лат. ligāre — «зшивати», «зв'язувати») — клас ферментів
6 клас
Лігази (від лат. ligāre — «зшивати», «зв'язувати») — клас ферментів

У класифікації міжнародної комісії з ферментів, лігази класифікуються як КФ 6
У класифікації міжнародної комісії з ферментів, лігази класифікуються як КФ 6
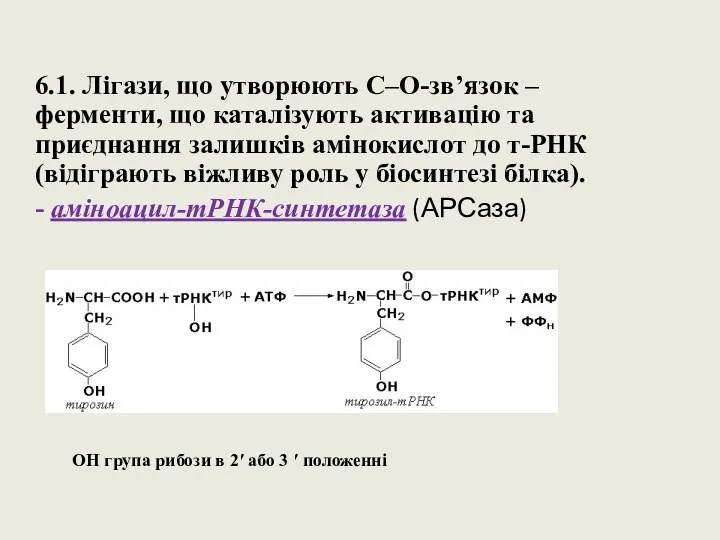
6.1. Лігази, що утворюють C–O-зв’язок – ферменти, що каталізують активацію та
6.1. Лігази, що утворюють C–O-зв’язок – ферменти, що каталізують активацію та

Лігази
6.3. Лігази, що утворюють C–N-зв’язок.
глютамінсинтетаза – каталізує реакцію синтезу глютаміну з
Лігази
6.3. Лігази, що утворюють C–N-зв’язок.
глютамінсинтетаза – каталізує реакцію синтезу глютаміну з

Ферменти, які використовуються в промисловості отримують з сировини тваринного, рослинного або
Ферменти, які використовуються в промисловості отримують з сировини тваринного, рослинного або

Застосування в промисловості
Хлібопекарська промисловість
амілази
протеази
ліпоксидази
Для відновлення смакових якостей і запаху
Застосування в промисловості
Хлібопекарська промисловість
амілази
протеази
ліпоксидази
Для відновлення смакових якостей і запаху

Целюлазу, пектиназу і нарингіназу (виробництво цитрусових соків, прибирає гіркий смак) застосовують
Целюлазу, пектиназу і нарингіназу (виробництво цитрусових соків, прибирає гіркий смак) застосовують
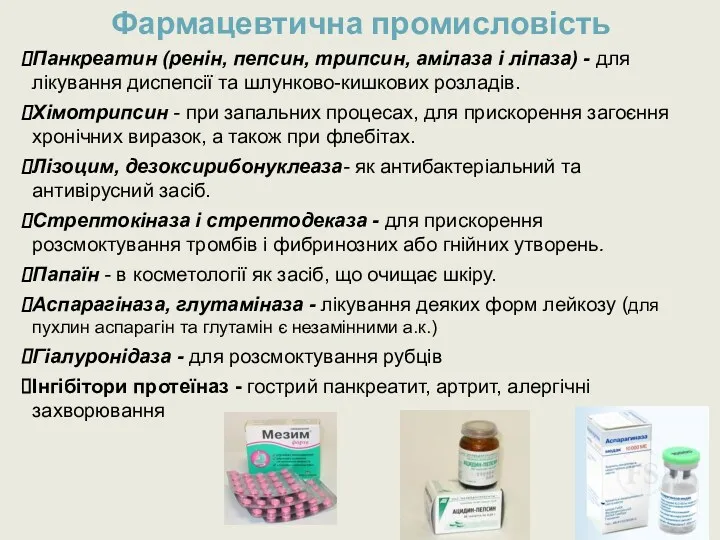
Панкреатин (ренін, пепсин, трипсин, амілаза і ліпаза) - для лікування диспепсії
Панкреатин (ренін, пепсин, трипсин, амілаза і ліпаза) - для лікування диспепсії

ІМОБІЛІЗОВАНІ ФЕРМЕНТИ
Ферменти, що закріплені на полімерному носію. Фізична або хімічна мобілізація.
Переваги:
Підвищення
ІМОБІЛІЗОВАНІ ФЕРМЕНТИ
Ферменти, що закріплені на полімерному носію. Фізична або хімічна мобілізація.
Переваги:
Підвищення
 Мезозойская эра. Эра средней жизни
Мезозойская эра. Эра средней жизни Иммунная система
Иммунная система Цветы нашей Родины
Цветы нашей Родины Одноклітинні організми. Лабораторне дослідження
Одноклітинні організми. Лабораторне дослідження Царства живой природы
Царства живой природы Нервная система рыб
Нервная система рыб Ленточные черви
Ленточные черви Anatomia sistemului nervos
Anatomia sistemului nervos Нуклеиновые кислоты: состав, строение, функции. Биология. 10 класс
Нуклеиновые кислоты: состав, строение, функции. Биология. 10 класс Экология растений. Экологические факторы и группы растений (Лекция №4)
Экология растений. Экологические факторы и группы растений (Лекция №4) Деление клетки. Митоз.
Деление клетки. Митоз. Особенности внешнего строения дождевого червя, связанные с жизнью в почве
Особенности внешнего строения дождевого червя, связанные с жизнью в почве Семейства и признаки покрытосеменных
Семейства и признаки покрытосеменных Отряд Рукокрылые
Отряд Рукокрылые Отдел Мхи
Отдел Мхи Функциональная анатомия мышц туловища и конечностей
Функциональная анатомия мышц туловища и конечностей Молекулярно-генетичні механізми онтогенезу. Порушення онтогенезу та їх місце в патології людини
Молекулярно-генетичні механізми онтогенезу. Порушення онтогенезу та їх місце в патології людини Выдающиеся ученые инфекционисты и эпидемиологи
Выдающиеся ученые инфекционисты и эпидемиологи Регуляция обмена веществ
Регуляция обмена веществ Непрямой онтогенез
Непрямой онтогенез Борьба за существование и ее формы. 11 класс
Борьба за существование и ее формы. 11 класс Вегетативная нервная система
Вегетативная нервная система Организм человека как единая биологическая система
Организм человека как единая биологическая система Человек как объект генетики. Методы изучения генетики человека
Человек как объект генетики. Методы изучения генетики человека What Is Down Syndrome?common Symptoms
What Is Down Syndrome?common Symptoms Імунна система. Імунітет. Імунізація. Алергія
Імунна система. Імунітет. Імунізація. Алергія Естественный отбор и борьба за существование
Естественный отбор и борьба за существование Царство грибы
Царство грибы