- Біофізика фотобіологічних процесів

Содержание
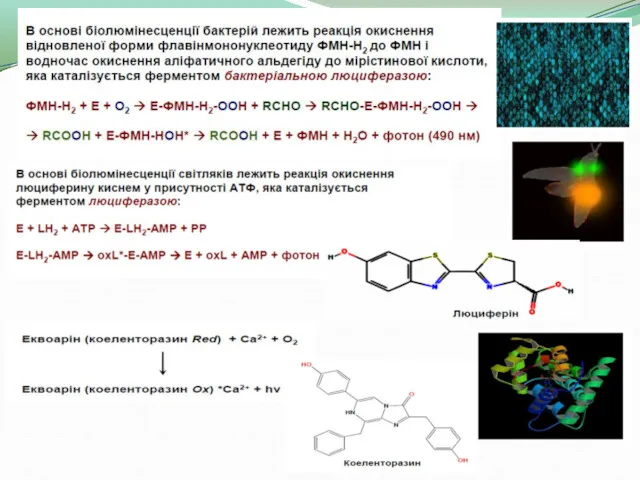
- 2. Деякі процеси, що протікають у біологічних системах, супроводжуються явищем люмінесценції (світіння). Світіння ряду організмів (деяких бактерій,
- 3. Люмінесценція супроводжує ряд біохімічних реакцій, найбільш поширеним і вивченим прикладом яких є реакції перекисного окислення ліпідів.
- 4. Походження явища Особливістю біолюмінесцентних систем є те, що вони не закріплювалися в філогенезі (тобто еволюційно). Більшість
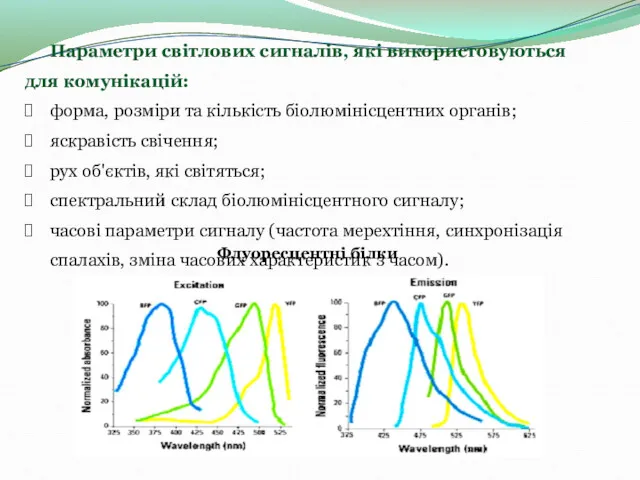
- 6. Параметри світлових сигналів, які використовуються для комунікацій: форма, розміри та кількість біолюмінісцентних органів; яскравість свічення; рух
- 7. Біохемілюмінесценція – світіння, що супроводжує хімічні реакції біологічних об’єктів ( свідчення гнилих дерев, світлячків , тощо).
- 8. Значення: Вимір інтенсивності люмінесценції використовується : в медицині; в біохімічних лабораторій (напр. для визначення експресії генів);
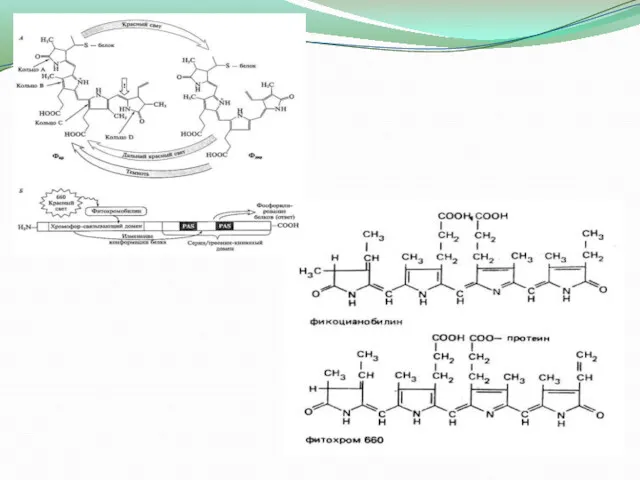
- 9. Фітохром вперше було виявлено у 1950-х роках американськими дослідниками А. Бортвіком, Б. Хендріксом і У. Паркером
- 10. Цей пігмент відіграє важливу роль в ряді процесів, таких як: *цвітіння і проростання насіння; *допомагає встановлювати
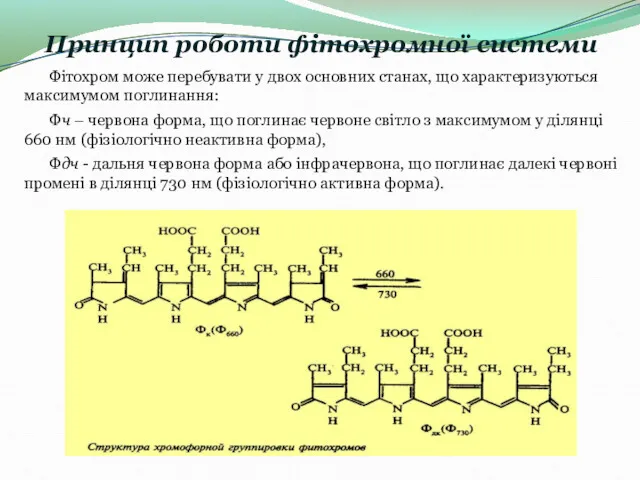
- 12. Принцип роботи фітохромної системи Фітохром може перебувати у двох основних станах, що характеризуються максимумом поглинання: Фч
- 13. Фітохімічні реакції в білках, ліпідах і нуклеїнових кислотах. Загалом можна сказати, що механізм дії фітохрому вивчений
- 14. Фотосинтез - це комплекс процесів асиміляції фотонів і хімічних субстратів в рослинному організмі, які призводять до
- 15. Структурна основа для фотосинтетичних процесів Фотосинтетичні процеси рослинах локалізовані в спеціальних органелах –хлоропластах. Внутрішній простір хлоропласта
- 16. Фізико-хімічна основа фотосинтезу Основна продукція фотосинтезу - це сполуки з високим відновним потенціалом - вуглеводи. Процес
- 17. Видно, що продуктом світлової стадії є відновлені переносники НАДФН2 і АТФ, які в темновій стадії відновлення
- 18. Перша стадія фотосинтезу протікає на світлі за допомогою фотокаталізатора. Фотокаталізатором в рослинах є хлорофіл - a,
- 19. Рис. Розташування електрон-транспортних комплексів (ФС1, ФС2, b/f-комплексу) та їх взаємодія в тилакоїдній мембрані.
- 20. Фізичний сенс фотосинтезу полягає в тому, що в цьому процесі відбувається електронна перебудова молекул. Загалом, фотосинтез
- 21. Рис. Використання в метаболізмі клітини енергетичного та відновного потен-ціалів, що вини-кають у фото-хімічних реакціях. Редокс-енергія в
- 22. Перетворення енергії в фотосинтетичних системах Основні етапи перетворення енергії включають в себе такі процеси: 1. Поглинання
- 23. Перенесення електронів по ЕТЛ супроводжується зниженням енергії. Цей процес можна уподібнити мимовільному руху тіла по похилій
- 24. Основні властивості фотосинтетичного апарату рослин 1. Насичення світлової кривої фотосинтезу із зростанням освітленості: Рис. По вертикальній
- 25. 3. Зміна окисно-відновного стану цитохромів b і f при освітленні світлом з різними довжинами хвиль. При
- 26. 5. Затримана флуоресценція. Слідом за загасанням флуоресценції молекули хлорофілу (декілька наносекунд) в живому листку спостерігається слабке
- 27. Отже, фотосинтез є основним джерелом біологічної енергії, фотосинтезуючі автотрофи використовують її для утворення органічних речовин з
- 29. Скачать презентацию
Деякі процеси, що протікають у біологічних системах, супроводжуються явищем люмінесценції (світіння).
Світіння
Деякі процеси, що протікають у біологічних системах, супроводжуються явищем люмінесценції (світіння).
Світіння

Люмінесценція супроводжує ряд біохімічних реакцій, найбільш поширеним і вивченим прикладом яких є реакції
Люмінесценція супроводжує ряд біохімічних реакцій, найбільш поширеним і вивченим прикладом яких є реакції

Походження явища
Особливістю біолюмінесцентних систем є те, що
вони не закріплювалися в філогенезі (тобто
Походження явища
Особливістю біолюмінесцентних систем є те, що
вони не закріплювалися в філогенезі (тобто

Параметри світлових сигналів, які використовуються для комунікацій:
форма, розміри та кількість біолюмінісцентних органів;
яскравість свічення;
рух
Параметри світлових сигналів, які використовуються для комунікацій:
форма, розміри та кількість біолюмінісцентних органів;
яскравість свічення;
рух
Біохемілюмінесценція – світіння, що супроводжує хімічні реакції біологічних об’єктів ( свідчення гнилих дерев,
Біохемілюмінесценція – світіння, що супроводжує хімічні реакції біологічних об’єктів ( свідчення гнилих дерев,

Значення:
Вимір інтенсивності люмінесценції використовується :
в медицині;
в біохімічних лабораторій (напр. для визначення експресії генів);
при
Значення:
Вимір інтенсивності люмінесценції використовується :
в медицині;
в біохімічних лабораторій (напр. для визначення експресії генів);
при

Фітохром вперше було виявлено у 1950-х роках американськими дослідниками А. Бортвіком, Б. Хендріксом
Фітохром вперше було виявлено у 1950-х роках американськими дослідниками А. Бортвіком, Б. Хендріксом

Цей пігмент відіграє важливу роль в ряді процесів, таких як:
*цвітіння і проростання насіння;
*допомагає
Цей пігмент відіграє важливу роль в ряді процесів, таких як:
*цвітіння і проростання насіння;
*допомагає

Принцип роботи фітохромної системи
Фітохром може перебувати у двох основних станах, що характеризуються максимумом
Принцип роботи фітохромної системи
Фітохром може перебувати у двох основних станах, що характеризуються максимумом
Фітохімічні реакції в білках, ліпідах і
нуклеїнових кислотах.
Загалом можна сказати, що механізм дії
Фітохімічні реакції в білках, ліпідах і
нуклеїнових кислотах.
Загалом можна сказати, що механізм дії

Фотосинтез - це комплекс процесів асиміляції фотонів і хімічних субстратів в рослинному організмі,
Фотосинтез - це комплекс процесів асиміляції фотонів і хімічних субстратів в рослинному організмі,

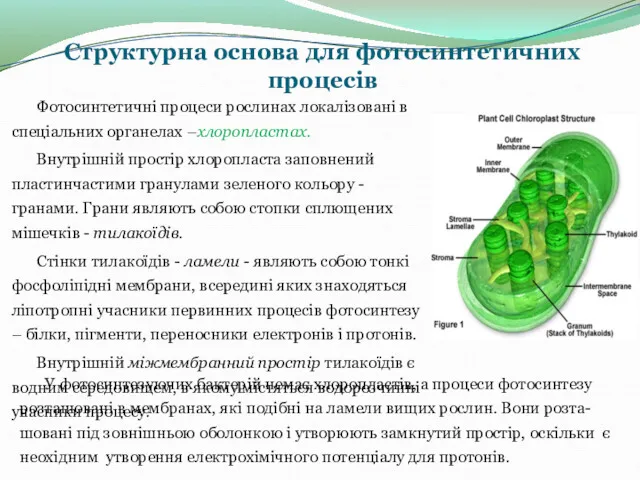
Структурна основа для фотосинтетичних процесів
Фотосинтетичні процеси рослинах локалізовані в спеціальних органелах –хлоропластах.
Внутрішній
Структурна основа для фотосинтетичних процесів
Фотосинтетичні процеси рослинах локалізовані в спеціальних органелах –хлоропластах.
Внутрішній
Фізико-хімічна основа фотосинтезу
Основна продукція фотосинтезу - це сполуки з високим відновним потенціалом -
Фізико-хімічна основа фотосинтезу
Основна продукція фотосинтезу - це сполуки з високим відновним потенціалом -

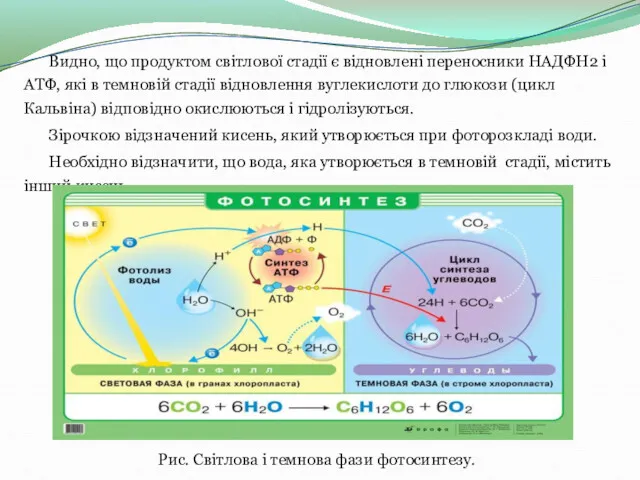
Видно, що продуктом світлової стадії є відновлені переносники НАДФН2 і АТФ, які в
Видно, що продуктом світлової стадії є відновлені переносники НАДФН2 і АТФ, які в
Перша стадія фотосинтезу протікає на світлі за допомогою фотокаталізатора. Фотокаталізатором в рослинах є
Перша стадія фотосинтезу протікає на світлі за допомогою фотокаталізатора. Фотокаталізатором в рослинах є

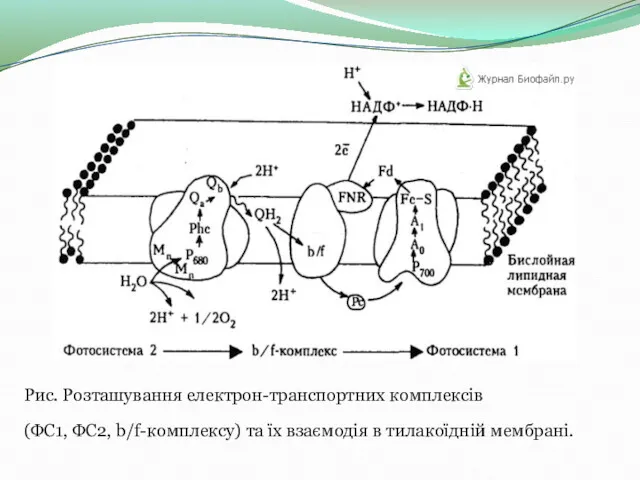
Рис. Розташування електрон-транспортних комплексів
(ФС1, ФС2, b/f-комплексу) та їх взаємодія в тилакоїдній мембрані.
Рис. Розташування електрон-транспортних комплексів
(ФС1, ФС2, b/f-комплексу) та їх взаємодія в тилакоїдній мембрані.
Фізичний сенс фотосинтезу полягає в тому, що в цьому процесі відбувається електронна перебудова
Фізичний сенс фотосинтезу полягає в тому, що в цьому процесі відбувається електронна перебудова

Рис. Використання в метаболізмі клітини енергетичного та
відновного потен-ціалів, що вини-кають у фото-хімічних
Рис. Використання в метаболізмі клітини енергетичного та
відновного потен-ціалів, що вини-кають у фото-хімічних

Перетворення енергії
в фотосинтетичних системах
Основні етапи перетворення енергії включають в себе такі процеси:
1.
Перетворення енергії
в фотосинтетичних системах
Основні етапи перетворення енергії включають в себе такі процеси:
1.

Перенесення електронів по ЕТЛ супроводжується зниженням енергії. Цей процес можна уподібнити мимовільному руху
Перенесення електронів по ЕТЛ супроводжується зниженням енергії. Цей процес можна уподібнити мимовільному руху

Основні властивості фотосинтетичного апарату рослин
1. Насичення світлової кривої фотосинтезу із зростанням освітленості:
Рис. По
Основні властивості фотосинтетичного апарату рослин
1. Насичення світлової кривої фотосинтезу із зростанням освітленості:
Рис. По

3. Зміна окисно-відновного стану цитохромів b і f при освітленні світлом з різними
3. Зміна окисно-відновного стану цитохромів b і f при освітленні світлом з різними

5. Затримана флуоресценція.
Слідом за загасанням флуоресценції молекули хлорофілу (декілька наносекунд) в живому
5. Затримана флуоресценція.
Слідом за загасанням флуоресценції молекули хлорофілу (декілька наносекунд) в живому

Отже, фотосинтез є основним джерелом біологічної енергії, фотосинтезуючі автотрофи використовують її для утворення
Отже, фотосинтез є основним джерелом біологічної енергії, фотосинтезуючі автотрофи використовують її для утворення

 урок по теме Размножение и оплодотворение растений 5 класс
урок по теме Размножение и оплодотворение растений 5 класс Научно-исследовательская работа. Наблюдение за аквариумными рыбками
Научно-исследовательская работа. Наблюдение за аквариумными рыбками электронная презентация по теме Птицы
электронная презентация по теме Птицы Животные тундры. 4 класс
Животные тундры. 4 класс Өсімдіктер әлемі
Өсімдіктер әлемі Эмбриональное развитие организма
Эмбриональное развитие организма Деревья плодовые, простые и кусты
Деревья плодовые, простые и кусты Класс Хрящевые рыбы
Класс Хрящевые рыбы КВН Живая планета
КВН Живая планета Биология калана, или выдры морской
Биология калана, или выдры морской Деревья леса
Деревья леса Адаптации организмов
Адаптации организмов Селекция животных
Селекция животных Белки. Качественный состав белков
Белки. Качественный состав белков Предмет і завдання мікробіології. Основні етапи розвитку. Систематика номенклатура і класифікація мікроорганізмів
Предмет і завдання мікробіології. Основні етапи розвитку. Систематика номенклатура і класифікація мікроорганізмів Побег - осевой орган высших растений
Побег - осевой орган высших растений Комнатные растения
Комнатные растения Размножение и индивидуальное развитие организмов. Бесполое размножение
Размножение и индивидуальное развитие организмов. Бесполое размножение Грегор Иоганн Мендель (20 июля 1822— 6 января 1884 гг.)
Грегор Иоганн Мендель (20 июля 1822— 6 января 1884 гг.) Земноводные. Среда обитания и внешнее строение
Земноводные. Среда обитания и внешнее строение Предмет и задачи микробиологии. Основные этапы развития. Систематика номенклатура и классификация микроорганизмов
Предмет и задачи микробиологии. Основные этапы развития. Систематика номенклатура и классификация микроорганизмов Головоногие моллюски
Головоногие моллюски Готовимся к ЕГЭ по биологии
Готовимся к ЕГЭ по биологии Общая вирусология. Систематика, морфология и физиология вирусов. Бактериофаги. Использование фагов в медицинской практике
Общая вирусология. Систематика, морфология и физиология вирусов. Бактериофаги. Использование фагов в медицинской практике Эволюция человека
Эволюция человека ЦНС. Головной мозг
ЦНС. Головной мозг Возрастная физиология. Физиологические особенности функционирования систем организма человека в различные возрастные периоды
Возрастная физиология. Физиологические особенности функционирования систем организма человека в различные возрастные периоды Экскурсия по Лапландскому заповеднику
Экскурсия по Лапландскому заповеднику