- Диплоидная линия эволюции

Содержание
- 2. Все высшие растения кроме мхов относят к диплоидной эволюционной ветви развития. В их жизненном цикле господствующим
- 4. Эволюция жизненного цикла У всех высших растений спорофит − зеленое автотрофное растение, в то время как
- 5. Споровые в наземных условия Приспособления наземных растений выразились в возрастающем расхождении спорофита и гаметофита, выполняющих различные
- 6. Массовое производство спор в наземных условиях В наземных условиях большая часть спор попадает в неблагоприятные условия
- 7. Расчленение спорофита Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением, так как для нормального
- 8. Появление проводящей системы Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной проводящей системы
- 9. Водное питание в наземных условиях Решение проблем водного режима привело к образованию ксилемы, или древесины, обеспечивающей
- 10. Стелярное строение Флоэма и ксилема в центральном цилиндре различных групп высших растений располагается по-разному. В связи
- 11. Стелы разных групп споровых Наиболее примитивным типом стелы является протостела риниофитов и некоторых других споровых. Позже
- 12. Эволюция стелы
- 13. Возникновение листьев В результате дальнейшего морфологического расчленения спорофита возникли специальные органы фотосинтеза − листья. У всех
- 14. Теломные листья Листья большинства споровых и семенных растений возникли в результате уплощения и срастания конечных веточек
- 15. Эволюция теломных листьев Теломные листья с самого начала были спороносными, выполняя как функции фотосинтеза так и
- 16. Происхождение листьев
- 18. Теломная теория
- 19. Эволюция ветвления Исходной формой ветвления спорофитов было равнодихотомичес-кое ( риниофиты, плауны, псилот), после чего растения выработали
- 20. Эволюция ветвления
- 21. Функции листьев у споровых В отличие от безлистных риниофитов на листьях высших споровых диплоидной линии эволюции
- 22. Разделение функций листьев В процессе эволюции происходило разделение этих функций. Так у ряда папоротников (ужовниковые, осмунда,
- 23. Два типа листьев страусопера Трофофиллы Спорофиллы
- 24. побеги со спорофиллами называются стробилами Стробилы споровых
- 25. Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения У равноспоровых морфологически одинаковые споры прорастают
- 26. Разноспоровые растения Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты, которые способны существовать лишь
- 27. Генеральная линия эволюции Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита. Редукция гаметофитов, т.е. полового поколения
- 28. Отделы высших споровых диплоидной линии эволюции
- 30. Риниофиты (RHYNIOPHYTA) или псилофитовидные Впервые они были найдены в 1859 г. канадским геологом Дж. Досоном в
- 31. Псилофит – голорос первичный (Psilophyton princeps)
- 32. Находка риниофитов Растение было названо Psilophyton princeps − голорос первичный. Необычное растение не укладывалось в существующие
- 33. Представители риниофитов Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
- 34. Представители риниофитов Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и др.).
- 35. Появление синангиев Ярравия
- 37. Скачать презентацию

Все высшие растения кроме мхов
относят к диплоидной эволюционной ветви
относят к диплоидной эволюционной ветви
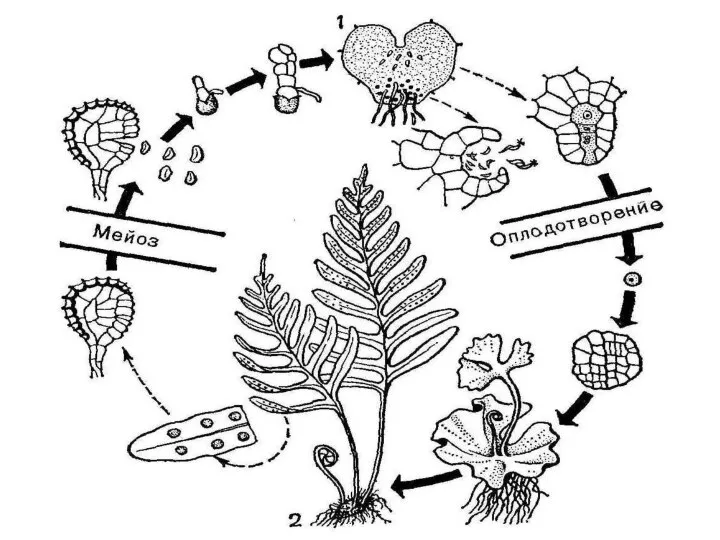

Эволюция жизненного цикла
У всех высших растений спорофит − зеленое автотрофное растение,
Эволюция жизненного цикла
У всех высших растений спорофит − зеленое автотрофное растение,

Споровые в наземных условия
Приспособления наземных растений выразились в возрастающем расхождении спорофита
Споровые в наземных условия
Приспособления наземных растений выразились в возрастающем расхождении спорофита

Массовое производство спор в наземных условиях
В наземных условиях большая часть спор
Массовое производство спор в наземных условиях
В наземных условиях большая часть спор

Расчленение спорофита
Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением,
Расчленение спорофита
Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением,

Появление проводящей системы
Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной
Появление проводящей системы
Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной

Водное питание в наземных условиях
Решение проблем водного режима привело к образованию
Водное питание в наземных условиях
Решение проблем водного режима привело к образованию

Стелярное строение
Флоэма и ксилема в центральном цилиндре различных групп высших растений
Стелярное строение
Флоэма и ксилема в центральном цилиндре различных групп высших растений

Стелы разных групп споровых
Наиболее примитивным типом стелы является протостела риниофитов и
Стелы разных групп споровых
Наиболее примитивным типом стелы является протостела риниофитов и
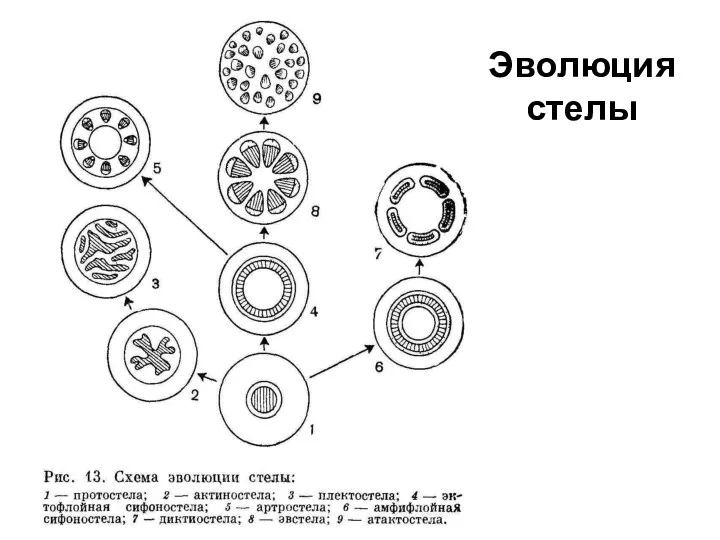
Эволюция
стелы
Эволюция
стелы

Возникновение листьев
В результате дальнейшего морфологического расчленения спорофита возникли специальные органы фотосинтеза
Возникновение листьев
В результате дальнейшего морфологического расчленения спорофита возникли специальные органы фотосинтеза

Теломные листья
Листья большинства споровых и семенных растений возникли в результате уплощения
Теломные листья
Листья большинства споровых и семенных растений возникли в результате уплощения

Эволюция теломных листьев
Теломные листья с самого начала были спороносными, выполняя как
Эволюция теломных листьев
Теломные листья с самого начала были спороносными, выполняя как
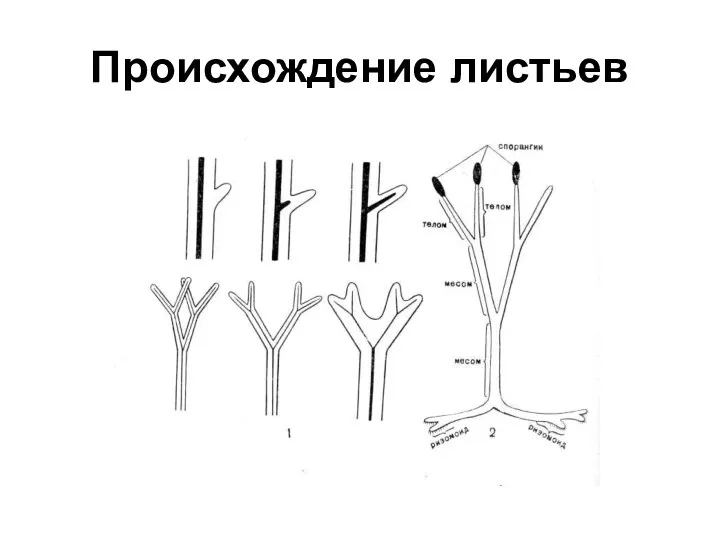
Происхождение листьев
Происхождение листьев

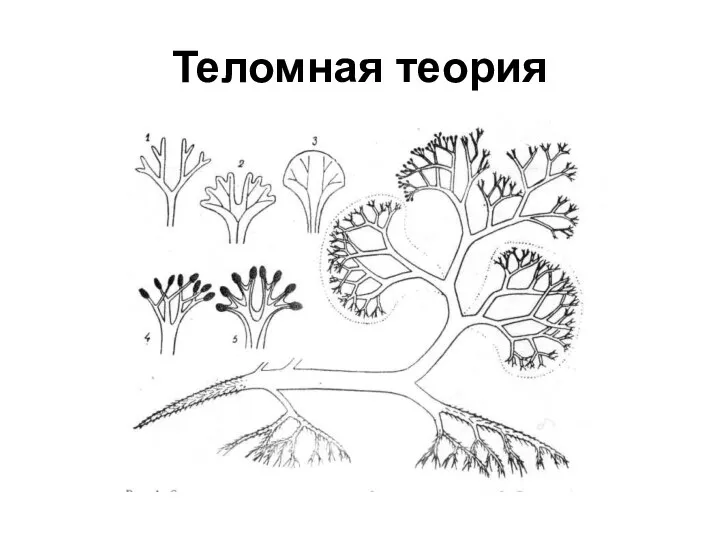
Теломная теория
Теломная теория

Эволюция ветвления
Исходной формой ветвления спорофитов было равнодихотомичес-кое ( риниофиты, плауны, псилот),
Эволюция ветвления
Исходной формой ветвления спорофитов было равнодихотомичес-кое ( риниофиты, плауны, псилот),
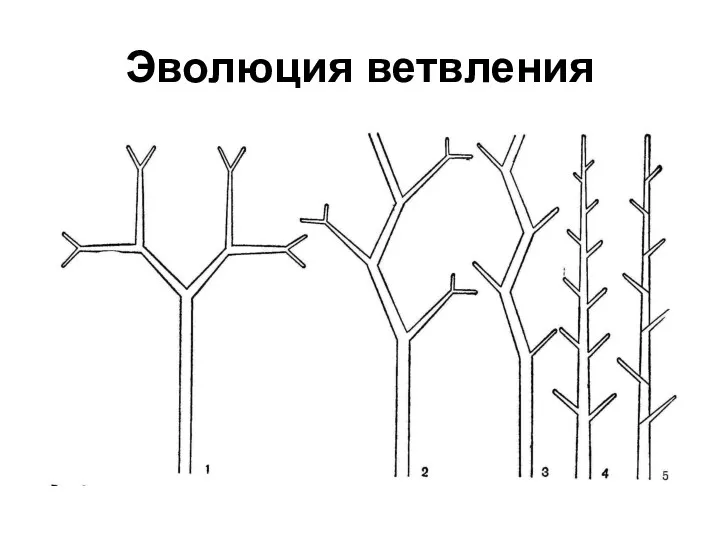
Эволюция ветвления
Эволюция ветвления

Функции листьев у споровых
В отличие от безлистных риниофитов на листьях высших
Функции листьев у споровых
В отличие от безлистных риниофитов на листьях высших

Разделение функций листьев
В процессе эволюции происходило разделение этих функций. Так у
Разделение функций листьев
В процессе эволюции происходило разделение этих функций. Так у

Два типа листьев страусопера
Трофофиллы
Спорофиллы
Два типа листьев страусопера
Трофофиллы
Спорофиллы

побеги со спорофиллами
называются стробилами
Стробилы споровых
побеги со спорофиллами
называются стробилами
Стробилы споровых
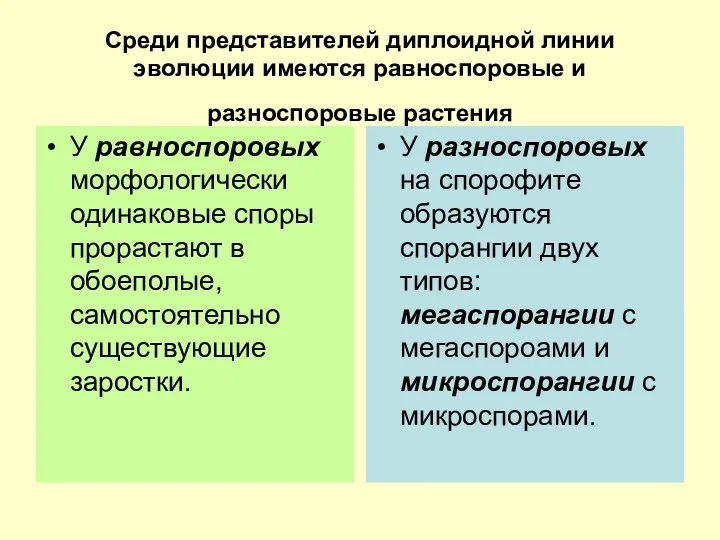
Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения
У
Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения
У

Разноспоровые растения
Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты,
Разноспоровые растения
Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты,

Генеральная линия эволюции
Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита.
Редукция гаметофитов,
Генеральная линия эволюции
Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита.
Редукция гаметофитов,

Отделы высших споровых диплоидной линии эволюции
Отделы высших споровых диплоидной линии эволюции
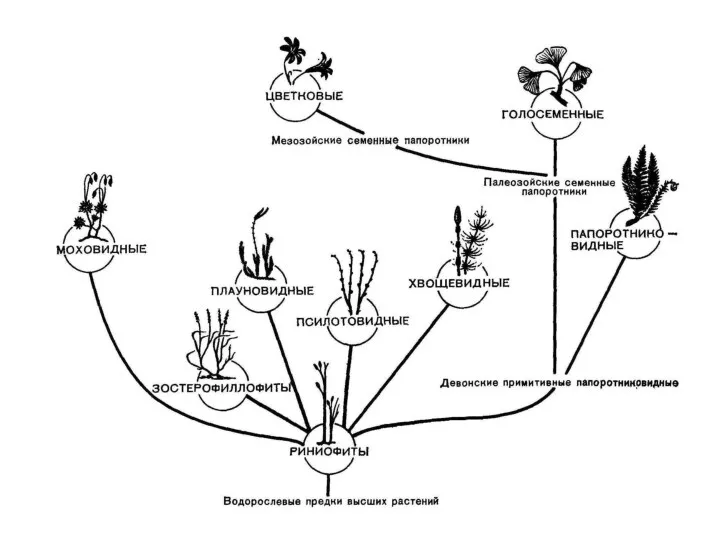

Риниофиты (RHYNIOPHYTA) или псилофитовидные
Впервые они были найдены в 1859 г. канадским
Риниофиты (RHYNIOPHYTA) или псилофитовидные
Впервые они были найдены в 1859 г. канадским
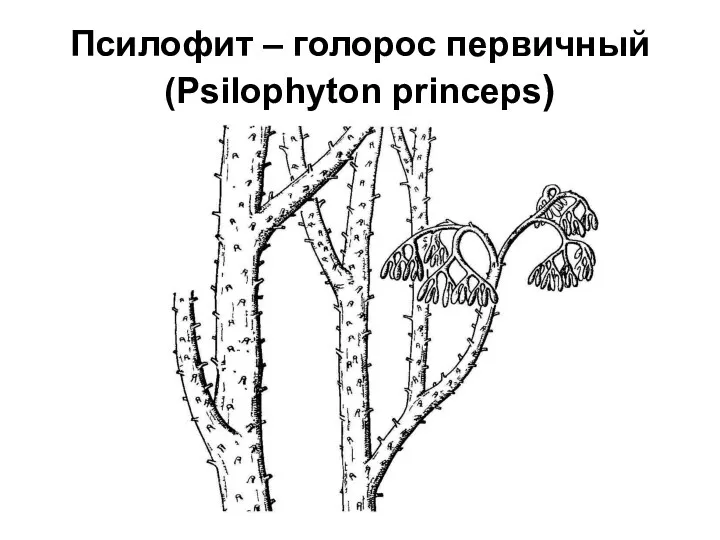
Псилофит – голорос первичный (Psilophyton princeps)
Псилофит – голорос первичный (Psilophyton princeps)

Находка риниофитов
Растение было названо Psilophyton princeps − голорос первичный. Необычное растение
Находка риниофитов
Растение было названо Psilophyton princeps − голорос первичный. Необычное растение
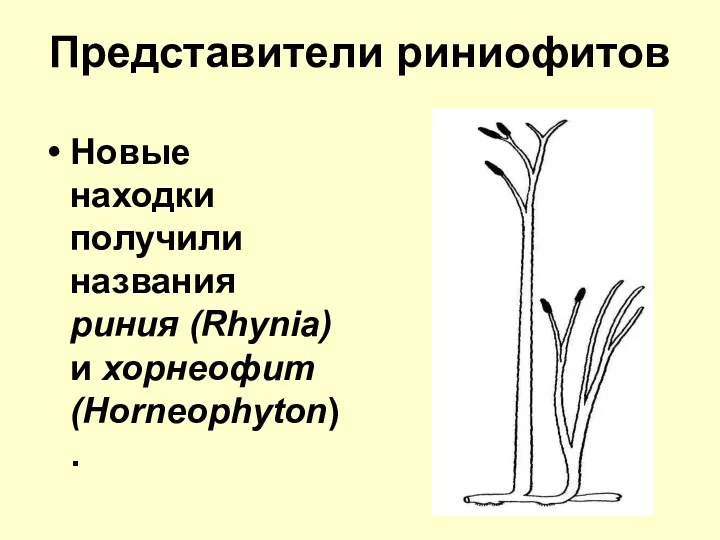
Представители риниофитов
Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
Представители риниофитов
Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
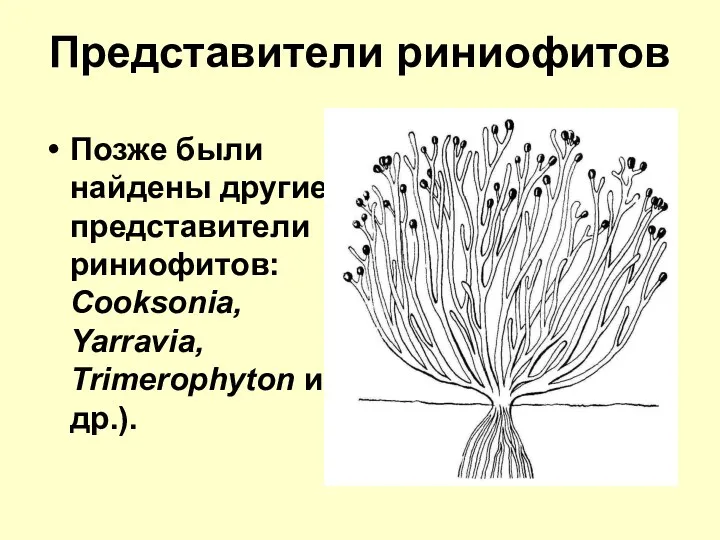
Представители риниофитов
Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и
Представители риниофитов
Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и

Появление синангиев
Ярравия
Появление синангиев
Ярравия
 О тех, кого не любят (Многообразие пресмыкающихся)
О тех, кого не любят (Многообразие пресмыкающихся) Расщепление пероксида водорода в клетках клубня картофеля. Лабораторная работа №1
Расщепление пероксида водорода в клетках клубня картофеля. Лабораторная работа №1 Ауыз су, өндірістік су
Ауыз су, өндірістік су Життя – це спосіб існування білкових тіл
Життя – це спосіб існування білкових тіл Процесс образования половых клеток - гаметогенез
Процесс образования половых клеток - гаметогенез Физические основы деятельности систем дыхания, кровообращения и энергообмена при мышечных движениях
Физические основы деятельности систем дыхания, кровообращения и энергообмена при мышечных движениях Вегетативное размножение растения
Вегетативное размножение растения Животные Красной книги Республики Коми
Животные Красной книги Республики Коми Өсімдіктерді қорғауда биологиялық препараттарды қолдану. (Лекция 13)
Өсімдіктерді қорғауда биологиялық препараттарды қолдану. (Лекция 13) Фотосинтез. Суммарная реакция фотосинтеза
Фотосинтез. Суммарная реакция фотосинтеза Метод максимальной парсимонии (продолжение). Метод максимального правдоподобия
Метод максимальной парсимонии (продолжение). Метод максимального правдоподобия Тварини-будівельники
Тварини-будівельники Генетика человека с основами медицинской генетики
Генетика человека с основами медицинской генетики Анатомия листа С3 и С4 растений. Особенности фиксация диоксида углерода в клетках мезофилла. Акцепторы диоксида углерода
Анатомия листа С3 и С4 растений. Особенности фиксация диоксида углерода в клетках мезофилла. Акцепторы диоксида углерода Морфология и классификация микроорганизмов (часть 3)
Морфология и классификация микроорганизмов (часть 3) Многообразие моллюсков
Многообразие моллюсков Отряд голенастые
Отряд голенастые Which statement about the properties of life is false?
Which statement about the properties of life is false? Сезонные изменения в природе и жизнедеятельности организмов
Сезонные изменения в природе и жизнедеятельности организмов Серцевий цикл. Робота серця
Серцевий цикл. Робота серця презентация Единство многообразия
презентация Единство многообразия Комнатные растения
Комнатные растения Молекулярный механизм генетических процессов. Репликация. (Лекция 10)
Молекулярный механизм генетических процессов. Репликация. (Лекция 10) Лікарська сировина тваринного походження
Лікарська сировина тваринного походження Ткани. Эпителиальная ткань. Соединительная ткань. Мышечная ткань
Ткани. Эпителиальная ткань. Соединительная ткань. Мышечная ткань Ядовитые грибы и ягоды
Ядовитые грибы и ягоды Свечение моря
Свечение моря Взаимосвязи животных в природе. Биологические сообщества
Взаимосвязи животных в природе. Биологические сообщества